
Агрономическое сопровождение и консультации. Стресс у растений
1.2 Основные группы факторов, способных вызвать стресс у растений. Устойчивость растений к недостатку кислорода, стрессу, температуре
Похожие главы из других работ:
Биоценоз и экосистема
4. Экологические группы: продуценты, консументы и редуценты
Для поддержания круговорота веществ в экосистеме необходимо наличие запаса неорганических веществ в усвояемой форме и трех функционально различных экологических групп организмов: продуцентов, консументов и редуцентов...
Изучение растительного покрова Черноморского побережья Туапсинского района
4.2.1 Экологические группы по отношению к абиотическим факторам
В результате проведенного экологического анализа был выделен ряд экоморф по отношению к влажности почвы. Преобладают мезофиты (72 %) - 116 видов (Sonchus arvensis, Achillea millefolium, Ulmus laevis и др.). Hа втором месте стоят ксерофиты (22 %) - 35 видов (Hypericum perforatum, Quercus petraea...
Микробиологическая индикация загрязнения окружающей среды
3.2 Почвенные микроорганизмы. Группы почвенных микроорганизмов
Почвенные микроорганизмы оказывают непосредственное влияние на собственно образование и формирование почвы, на минерализацию (разложение) органических остатков и образование гумуса. Поэтому, не имея представления об основных экологических...
Некоторые вопросы по экологии
4.2 каковы источники и основные группы загрязняющих веществ атмосферы России
В последние годы содержание в атмосферном воздухе российских городов и промышленных ненов таких вредных примесей, как взвешенные вещества, диоксид серы, существенно уменьшилось...
Организм и условия его обитания
1.2 Экологические группы гидробионтов
Толща воды, или пелагиаль (pelages -- море) заселена пелагическими организмами, которые обладают способностью плавать или удерживаться в определенных слоях. В связи с этим данные организмы подразделяются на две группы: нектон и планктон...
Организм и условия его обитания
3.2 Экологические группы почвенных организмов
Неоднородность почвы приводит к тому, что для организмов разных размеров она выступает как разная среда. По степени связи с почвой как средой обитания животных объединяют в три экологические группы: геобионты, геофилы и геоксены...
Организм и условия его обитания
3.4 Роль эдафических факторов в распределении растений и животных
На наземных животных эдафические факторы оказывают меньшее влияние. Вместе с тем животные тесно связаны с растительностью, и она играет решающую роль в их распределении. Среди крупных позвоночных легко обнаружить формы...
Пищевые цепи и трофические уровни
1.1 Группы организмов, участвующие в биогеоцинозе
Биотический круговорот происходит в природной системе, объединяющей на основе обмена веществ и энергии сообщество живых организмов (биоценоз) с неживыми компонентами -- условиями обитания. Такая система получила название биогеоценоз (греч...
Приспособления растений к водному режиму
2. ЭКОЛОГИЧЕСКИЕ ГРУППЫ НАЗЕМНЫХ РАСТЕНИЙ ПО ОТНОШЕНИЮ К ВОДЕ
Различные виды растений по-разному реагируют на изменяющиеся условия водоснабжения. Одни приспособлены к резким колебаниям содержания воды в среде и при этом сохраняют способность к поглощению воды из почвы...
Растительный мир Омской области
1.1 Основные виды покрытосеменных растений Омской области
Покрытосеменные, или цветковые, наиболее совершенные, разнообразные, распространенные, интересные с научной и ценные с хозяйственной точки зрения растения. Покрытосеменные - самый большой отдел растительного мира. Это молодая...
Устойчивость растений к недостатку кислорода, стрессу, температуре
Глава I. Общие представления о стрессе и факторах, вызывающих стресс у растений
...
Устойчивость растений к недостатку кислорода, стрессу, температуре
1.1 Стресс и "триада" Селье
Термин стресс (от англ. "stress" - напряжение) был предложен выдающимся канадским ученым-физиологом Гансом Селье в 1936 году для описания реакции организма на любое сильное неблагоприятное воздействие...
Физико-химические методы очистки сточных вод
2. Анализ реализации исследуемой технологии на выбранном производственном участке. Действие опасных факторов и вредных факторов на рабочем месте
...
Экология водорослей
ГЛАВА 2. ЭКОЛОГИЧЕСКИЕ ГРУППЫ
...
Экосистемы
4. Основные законы действия абиотических факторов.
...
eco.bobrodobro.ru
АМИНОКИСЛОТЫ В БОРЬБЕ СО СТРЕССОМ У РАСТЕНИЙ
АМИНОКИСЛОТЫ В БОРЬБЕ СО СТРЕССОМ У РАСТЕНИЙ
Стресс – это реакция растительного организма на воздействие неблагоприятных факторов, которая сопровождается замедлением метаболических процессов, затратами энергии на преодоление негативных факторов внешней среды в ущерб формированию урожая. Основной принцип действия антистрессантов заключается в том, что специально подобранные органические биологически активные вещества запускают, поддерживают и стимулируют физиологические процессы растительного организма, улучшают усвоение питательных элементов и в кратчайшие сроки восстанавливают рост и развитие растений.
В практике сельскохозяйственного производства в условиях наступления ранней весны, когда идет активная вегетация озимых культур, на Северном Кавказе нередко наблюдается возврат холодов в апреле и мае. Это отрицательно сказывается на росте, развитии растений и закладке их потенциальной продуктивности. Низкие положительные температуры в дневное время (+6, +8 0С) и отрицательные в ночное (по прогнозу метеорологов в первой декаде апреля температура ночью будет опускаться до -2, -4 0С) не позволят провести в короткие агротехнические сроки обработку посевов гербицидами. Период борьбы с сорной растительностью может растянуться в крае до 20-25 дней.
Растения озимых культур будут в это время переходить к фазе трубкования и на данном этапе онтогенеза действие гербицидов особенно отрицательно скажется на процессах формирования элементов продуктивности колоса и обмене веществ, поскольку снизится интенсивность фотосинтеза, замедлится синтез важнейших аминокислот (структурных компонентов белков), приостановятся ростовые процессы растений. Стрессовый эффект в зависимости от роста и развития растений, характеру температурного режима в период до и после обработки посевов гербицидами может проявляться в течение 8-15-ти дней.
Применение аминокислот во внекорневых подкормках, хотя и может вызвать некоторые вопросы и дискуссии, без сомнения, в настоящее время является одним из самых эффективных способов нивелирования влияния вредных условий окружающей среды на сельскохозяйственные растения.
Стрессовые факторы в зависимости от генезиса действия, можно разделить на 2 группы:
- Абиотические
- Излучение
- Температура (повышенная, пониженная)
- Вода (засуха, чрезмерная влажность)
- Газы (дефицит СО2, О2)
- Минеральные компоненты (дефицит, избыток, тяжелые металлы)
- Механические (ветер, снежный покров, ледяная корка)
- Биотические
- Микроорганизмы (вирусы, бактерии, грибы)
- Животные (грызуны, паразиты, вытаптывание животными)
- Антропогенные (промышленное загрязнение, пестициды)
- Растения (перенасыщение, конкуренция, аллелопатия, растения-паразиты)
Таким образом, стрессы могут иметь различную природу и воздействовать на растения в коротком или длинном временном промежутке. Стрессовые факторы, обусловленные погодой (например, засуха, чрезмерная влажность, низкая и высокая температуры), могут длиться долгое время или иметь кратковременное действие, оказывая в большинстве случаев негативное влияние на величину и качество урожая.
ПУТИ МЕТАБОЛИЗМА АЗОТА В РАСТЕНИИ И СОХРАНЕНИЕ ЭНЕРГИИ
Как известно, растения синтезируют собственные аминокислоты из неорганического азота. Данный процесс включает превращение нитратов в нитриты и аммонийные соединения и их последующее усвоение в органические молекулы глутаминовой кислоты. Основываясь на этой аминокислоте, растения синтезируют остальные через процесс переаминирования.
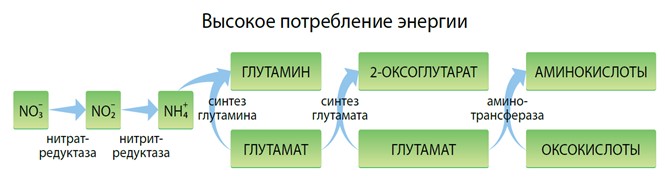
Вышеуказанные метаболические процессы поглощают много энергии, это и является причиной того, что при стрессовых ситуациях растения необходимо напрямую снабжать аминокислотами, высвобождая энергию для прочих физиологических реакций. Доказано, что аминокислоты легко усваиваются на листовом и корневом уровнях и могут напрямую использоваться растениями.
Подвергнутые стрессу растения, сразу активируют защитные механизмы и производят разнообразные субстанции, являющиеся активатором для продуцирования аминокислот, а впоследствии - белков. Использование препаратов, которые в своем составе содержат вещества, активизирующие синтез аминокислот и сами аминокислоты способствует тому, что не тратится энергия на их выработку - они сразу включаются в метаболизм растения.
 Аминокислоты (L-глутаминовая, глицин, лизин и др.), входящие в состав препарата Аминокат 10%, 30% представляют собой готовый строительный материал для биосинтеза молекул белка. Аминокислоты, попадая на лист, практически сразу включаются в обмен веществ без дополнительных затрат времени и энергии на преобразование неорганических веществ в органические. Поступая в растения через листовой аппарат, они в кратчайшие сроки - от нескольких часов – до 2-3-х дней - восстанавливают обмен веществ в растениях после воздействия любых стрессовых факторов (химических, физических, биологических) и активизируют функциональную деятельность всего растительного организма.
Аминокислоты (L-глутаминовая, глицин, лизин и др.), входящие в состав препарата Аминокат 10%, 30% представляют собой готовый строительный материал для биосинтеза молекул белка. Аминокислоты, попадая на лист, практически сразу включаются в обмен веществ без дополнительных затрат времени и энергии на преобразование неорганических веществ в органические. Поступая в растения через листовой аппарат, они в кратчайшие сроки - от нескольких часов – до 2-3-х дней - восстанавливают обмен веществ в растениях после воздействия любых стрессовых факторов (химических, физических, биологических) и активизируют функциональную деятельность всего растительного организма.
Среди большого количества аминокислот, которые выполняют в растениях различные функции, наряду с глутаминовой, особого внимания заслуживают ещё две – пролин и глицин. Пролин – способствует повышению иммунитета растений в стрессовых ситуациях и накоплению азота, усиливает способность семян к прорастанию, улучшает эффективность фотосинтеза и увеличивает содержание хлорофилла. Его действие заключается в улучшении генеративного развития растений и их плодоношения. Пролин влияет на завязывание плодов, регулирует водообмен в растении – он накапливается в клетках растений в случае водного и осмотического стресса. Глицин выполняет роль комплексообразующего вещества (хелата), которое является прекурсором хлорофилла, благодаря чему влияет на увеличение эффективности фотосинтеза.
МЕХАНИЗМ ДЕЙСТВЯ АНТИСТРЕССОВЫХ ПРЕПАРАТОВ НА ОСНОВЕ ДРУГИХ ОРГАНИЧЕСКИХ ВЕЩЕСТВ
В отличие от аминокислот, полимерные соединения различной органической природы - гуматы; Альбит и др. действуют на растения опосредованно и значительно медленнее, поскольку это крупные органические молекулы, которые из-за значительных размеров не способны проникать в клетки растений и принимать непосредственное участие в процессах биосинтеза.
Препараты на основе указанных соединений способны стимулировать обмен веществ у растений после достаточно длительного процесса микробиологического расщепления в течение 8-12 дней, а также за счет воздействия продуктов метаболизма микроорганизмов, оказывающих разлагающее воздействие на указанные вещества.
 Однако длительный процесс разложения сопряжен с более длительным периодом снижения у растений метаболической и фотосинтетической активности. Сложная и разнообразная химическая структура органических полимерных соединений различного происхождения, огромное разнообразие эпифитной микрофлоры, обитающей на поверхности растений, к сожалению, не позволяет точно прогнозировать состав и количество продуктов разложения, а, соответственно, и характер их влияния на растительный организм.
Однако длительный процесс разложения сопряжен с более длительным периодом снижения у растений метаболической и фотосинтетической активности. Сложная и разнообразная химическая структура органических полимерных соединений различного происхождения, огромное разнообразие эпифитной микрофлоры, обитающей на поверхности растений, к сожалению, не позволяет точно прогнозировать состав и количество продуктов разложения, а, соответственно, и характер их влияния на растительный организм.
Так, например, точные молекулярные формулы гуминовых кислот отсутствуют. Все предложенные учеными варианты имеют вид схем, в которых учтены лишь состав и некоторые свойства соединений. Расположение атомов, атомных групп в них определяется гипотетически. Отсутствие у гуминовых кислот формул объясняется их переменным составом и полидисперсностью даже, казалось бы, в однородных органических соединениях. Поэтому обычные методы определения формул биоорганических молекул для гуминовых кислот оказались недейственными.
Действующее вещество препарата Альбит – биополимер поли-бета-гидроксимасляная кислота. Это естественное запасное вещество полезных почвенных бактерий (подобно крахмалу у растений, жиру и гликогену у животных), которое служит субстратом для роста и развития микроорганизмов, но не способно использоваться растениями напрямую в биохимических процессах.

ПРОЦЕССЫ, АКТИВИЗИРУЮЩИЕСЯ В РАСТЕНИЯХ ПОД ДЕЙСТВИЕМ АМИНОКИСЛОТ
Осморегуляция
Свободные аминокислоты, накопленные в клетках во время стрессовых ситуаций, являются результатом синтеза или расщепления белков. Эти аминокислоты вместе с растворенными органическими веществами имеют высокую степень клеточной осморегуляции, которая увеличивает устойчивость клеток к неблагоприятным факторам. Примером является стресс под воздействием засоленности. Накопление в клеточной цитоплазме свободных аминокислот, особенно пролина, позволяет поддерживать низкий уровень ионов, которые могут быть токсичными, например Cl– и Na+.
Регулирование открытия устьиц и активности фотосинтеза
 Закрытие устьиц растений, провоцируемое стрессовыми условиями (высокой температурой, низкой относительной влажностью и т.п.), уменьшает активность фотосинтеза, замедляет все процессы обмена веществ. Экзогенное внесение аминокислот в растения при стрессовых условиях способствует открытию устьиц (рисунок 1, 2). Этот эффект позволяет увеличить удержание влаги в тканях, активизировать фотосинтез, скорректировать скорость испарения и отсрочить увядание.
Закрытие устьиц растений, провоцируемое стрессовыми условиями (высокой температурой, низкой относительной влажностью и т.п.), уменьшает активность фотосинтеза, замедляет все процессы обмена веществ. Экзогенное внесение аминокислот в растения при стрессовых условиях способствует открытию устьиц (рисунок 1, 2). Этот эффект позволяет увеличить удержание влаги в тканях, активизировать фотосинтез, скорректировать скорость испарения и отсрочить увядание.
Антиоксидантная активность
Во время стрессовых ситуаций растения реагируют накоплением веществ с большой оксидативной активностью (О2–, ОН, Н2О2 и О2), что ухудшает структуры клеток. При этих условиях растения используют, среди прочих, азотные соединения, которые действуют как естественные антиоксиданты: аминокислоты, амины, полиамины и определенные ферменты (пероксидазы, каталазы, пероксид дисмутазы). Среди аминокислот антиоксидантными свойствами выделяются аргинин, гистидин, цистеин, триптофан, лизин, метионин и треонин. Применение препарата АМИНОКАТ (10%, 30%) способствует увеличению содержания в растительных тканях антиоксидантов (SOD).
Стимуляция гормональной регуляции
Было замечено, что аминокислоты оказывают влияние на уровни гормонов в растениях. Растения испытывают гормональный дисбаланс из-за внешних факторов, особенно в критические фазы развития. Аминокислоты способны самостоятельно влиять на синтез гормонов, чтобы сбалансировать потребности растения. В то же время комбинированное применение аминокислотных препаратов с гиберрелинами и ауксинами проявляет важный эффект синергизма, когда аминокислоты и гормоны взаимно усиливают действие друг друга. Пуриновые и пиримидиновые основания, присутствующие в составе АМИНОКАТ (10%, 30%) способны стимулировать формирование цитокинов в растении, поскольку они являются прекурсорами этих гормонов. Они воздействуют на деление клеток и воспроизводство, что приводит к усилению роста.
Активизация и защита процессов опыления и плодоношения
Доступность аминокислот в органах цветения является основой успешного прорастания пыльцы и надлежащего и быстрого развития пыльцевой трубки. Развитие пыльцевой трубки происходит благодаря аминокислотам и сахарам, потребляемым пестиком цветка. Из-за этого растению важно иметь в своем распоряжении эти вещества. Особенно способствуют прорастанию пыльцы пролин и глутаминовая кислота.
Опыление и оплодотворение цветов — явление, существенно подвергающееся внутренним и внешним факторам, способным отрицательно повлиять на завязывание и формирование плодов. Было замечено, что внешняя обработка аминокислотами уменьшает время оплодотворения. Сокращение времени оплодотворения увеличивает возможность успешного завязывания плодов при неблагоприятных условиях, особенно экстремальных температурах, как высоких, так и низких. Кроме того, аминокислоты способствуют сбалансированному питанию растения, что увеличивает его способность к удержанию плодов и уменьшает сброс завязи.
Повышение эффективности средств защиты растений
Эффективность средств защиты растений зависит от интенсивности их поглощения через поверхностные структуры растения. Данный процесс, главным образом, идет через клеточные мембраны, а также через каналы и поры растения. Клеточные мембраны содержат высокий процент мембранных белков, способных к изменению проводимости в соответствии со структурой, в которой они находятся. Различные структуры определяются последовательностью аминокислот, с которыми они сочетаются. Обработка аминокислотными препаратами вместе со средствами защиты растений, регуляторами роста, листовыми подкормками, увеличивает проводимость мембраны, повышая скорость усвоения активных веществ.
РЕЗУЛЬТАТЫ ПРИМЕНЕНИЯ ПРЕПАРАТА АМИНОКАТ В ХОЗЯЙСТВАХ КРАЯ
На сегодняшний день одним из наиболее эффективных и востребованных с/х производством антистрессовым препаратом на основе аминокислот является препарат Аминокат 10%, 30%. Его применение в технологии возделывания с/х культур становится обязательным приемом, который прочно входит в практику выращивания различных культур на Ставрополье. Данный препарат является наиболее доступным и относительно недорогим средством управления стрессом у растений.
Высокую эффективность препарат демонстрирует в случае необходимости активизации роста и развития вегетативной массы, предупреждении стрессовых состояний растений (пестицидные обработки), а так же для ликвидации последствия различных повреждающих факторов - повреждение градобоем, избыточной дозой или последействием гербицидов. Даже при крайне угнетенном состояним посевов, обработка данным препаратом позволяет в короткие сроки восстановить обмен веществ растений, получить активно растущий листовой аппарат и достичь уровня продуктивности посева близкого к продуктивности неповрежденных растений. Урожайность сахарной свеклы серьезно поврежденной градобоем после применения аминокислотных препаратов составила 486 ц/га, при дегистии 16,1% (рисунок 4).
В одном из хозяйств Красногвардейского района обследование посевов кукурузы свидетельствовало о серьезном стрессовом состоянии растений, вызванным, по данным специалистов агрослужбы хозяйства, последействием гербицида Фенизан (состав: дикамба+хлорсульфорон). Данный гербицид обладает последействием в том случае, если между применением гербицидов и высевом последующей культуры выпало малое количество осадков. У растений отмечалась задержка роста, слабое развитие корневой системы, хотя по развитию они уже находились в фазе 6-7 листьев (рисунок 3). При проведении функциональной экспресс-диагностики была выявлена очень низкая активность хлоропластов (8,7 единиц) и существенный дефицит цинка и серы.
Применение препарата Аминокат 10% позволило вывести растения кукурузы из состояния стресса, при этом повторное проведение ФЭД через 16 дней после обработки указывало на то, что активность хлоропластов увеличилась до 26 единиц, наблюдался активный прирост биомассы, увеличилась площадь листовой поверхности, цвет растений стал более темно-зеленым, начала интенсивно развиваться корневая система (в том числе и воздушные корни). В результате на полях, где растения были подвержены стрессу, урожайность зерна составила 65 ц/га при средней урожайности кукурузы по хозяйству 80 ц/га.


agro-analiz.com
Раневой стресс растений
Трудно, пожалуй, найти другой научный термин, который так прочно вошел бы в жизнь современного общества, как стресс.
Впервые этот термин ввел известный канадский физиолог Ганс Селье в 1936 г. для объяснения реакции организма на самые различные невзгоды, для преодоления которых требуется напряжение всех его сил.
Согласно определению самого Селье, стресс представляет собой совокупность всех неспецифических изменений, возникающих под влиянием любых сильных воздействий, и сопровождается перестройкой защитных сил организма. Уже из определения следует, что неспецифические реакции имеют прямое отношение к иммунитету против инфекционных болезней, хотя в целом иммунитет рассматривается как проблема специфичности. Ведь что может быть более специфичным, чем реакция антиген—антитело. И все же очень большую роль, особенно в иммунитете растений, играют неспецифические реакции. Более того, они в ряде случаев могут определить исход взаимоотношений между хозяином и паразитом. К тому же изучение неспецифических реакций может приблизить нас к пониманию специфических и, следовательно, к проблеме иммунитета в целом.
К числу самых распространенных неспецифических защитных реакций, по крайней мере в растительном мире, относится реакция на механические поранения, которые неизбежны при ветре, граде и других неблагоприятных условиях. Такого рода реакции, несомненно, возникли еще до появления паразитов, и можно думать, что они в какой-то мере явились основой для развития у растений защитных реакций против паразитов. Ведь многие паразиты способны проникать в растительную ткань только лишь после того, как механически ее повредят.
Реакцию растений на механическое повреждение часто называют еще процессами репарации, залечивания, заживления, образования раневой перидермы. Особенно хорошо исследована раневая реакция клубней картофеля, которые являются излюбленным объектом для проведения такого рода работ. Неповрежденный клубень покрыт так называемой естественной перидермой, точной копией которой является раневая перидерма, которая образуется на пораненном участке. Естественная перидерма защищает клубни от неблагоприятных воздействий и излишнего испарения.
Под микроскопом видно, что естественная перидерма представляет собой многослойные ряды уплощенных клеток, расположенных одна под другой. Оболочки таких клеток содержат особое вещество типа пробки, носящее название суберина. Суберин адкрустирует клеточную оболочку, накладываясь на нее изнутри. Под электронным микроскопом суберинизация клеточных стенок выглядит слоистой из-за чередования слоев суберина и воска.
Суберин представляет собой полимер оксимонооксиди — и дикарбоновых кислот. В составе его находятся также некоторые полициклические вещества терпеновой природы. Вещества, входящие в состав пробки, делают ее высокоустойчивой к влиянию множества химических воздействий, таких, как органические кислоты, разбавленная соляная кислота, слабые щелочи. Наличие суберина в составе покровных тканей усиливает их механические свойства, поскольку большинство микроорганизмов не содержит ферментов, расщепляющих эти полимеры.
В процессе суберинизации происходит закупоривание пор в клеточных оболочках, в результате чего прерываются межклеточные контакты. Содержимое клеток, оболочки которых пропитываются суберином, постепенно отмирает, их полости наполняются воздухом, в результате чего клетки становятся термо — и влагонепроницаемыми.
В ответ на механическое поранение растения отвечают защитной раневой реакцией, суть которой сводится к образованию на пораненном участке точно такой же перидермы, какой является естественная. В результате поранения нарушается структурная и функциональная целостность растения или отделенного от него органа. В ответ на нарушение включаются системы иммунологического контроля, имеющие целью ликвидировать повреждение и восстановить нарушенную целостность. Образующаяся на месте поранения раневая перидерма по своим структурным и функциональным характеристикам ничем не отличается от естественной. Ее образование имеет целью как можно скорее закрыть ворота в кладовые растительных тканей, которые оказались открытыми в результате поранения. Неспособность растительных тканей быстро реагировать на повреждение может оказаться гибельной для организма в целом.
Как же происходит образование раневой перидермы? Взрослые клетки, давно прекратившие клеточные деления и находящиеся на поверхности поранения или залегающие непосредственно под ним, вновь приобретают меристематическую активность (способность к делению). Клетки обычно делятся параллельно пораненной поверхности, в них закладывается феллоген или пробковый камбий. Феллоген начинает делиться, отделяя наружу себе подобные по форме клетки, которые называются феллемой. В большинстве случаев это мертвые клетки, оболочки которых пропитаны суберином. Внутрь клубня феллоген также отпочковывает часть клеток такой же формы, которые остаются живыми и носят название феллодермы. Весь перидермальный комплекс состоит из приблизительно одинаковых расположенных друг под другом клеток феллемы, феллогена и феллодермы.
Для того чтобы взрослые клетки на поверхности поранения приобрели меристематическую активность, они должны получить стимул к делению или раневому возбуждению. Вокруг природы этого стимула развернулась длительная дискуссия. Существование раневого гормона предсказал еще 60 лет тому назад Г. Габерланд на основании своих опытов. На разрезанной поверхности кольраби, как и на клубне картофеля, образовывалась раневая перидерма. В том случае, если поверхность ломтика промывалась водой, образование перидермы задерживалось. Если же на предварительно промытую поверхность наносили кашицу из растертых тканей, перидерма вновь начинала образовываться. Это позволило Габерланду прийти к заключению, что из продуктов отмерших и разрушенных клеток образуются вещества, вызывающие раневое возбуждение. Он назвал их раневыми гормонами или некрогормонами.
Вещество, получившее название раневого гормона, было выделено в конце 30-х годов Д. Инглишем и Д. Боннером. Им оказалась Δ-децен-1,10-дикарбоновая кислота, получившая название травматиновой кислоты, или травматина. Это вещество было обнаружено в картофеле, брюссельской капусте, люцерне, рисе, томатах, апельсинах и других растениях, но отсутствовало в тканях животных. В качестве раневого гормона испытывались также иные кислоты жирного ряда. Выяснилось, что активность коррелировала с присутствием двойной связи в молекуле и возрастала при увеличении числа углеродных атомов.
В самое последнее время другое вещество, напоминающее по своим свойствам раневой гормон, было обнаружено у ряда растений. Под влиянием различных стрессовых воздействий, в том числе и поранения, это вещество освобождалось из клеточных стенок растений, быстро передавалось по проводящей системе и индуцировало в здоровых тканях, удаленных от места повреждения, образование ингибиторов протеиназ, т. е. веществ, подавляющих ферментные системы, расщепляющие белки. Растения, таким образом, как бы заранее готовятся отражать атаку ферментов паразитарного микроорганизма, который может проникнуть через места механических поранений. Поскольку образующийся сигнал принадлежит самому растению, а не вводится паразитом, он подходит под определение раневой гормон. По предварительным данным, вещество представляет собой рамногалактуронан.
Вещество широко распространено в растительном мире. Если экстракты из 37 видов растений наносили на молодые листья томатов, то они индуцировали в них накопление ингибиторов протеиназ. Только два вида экстрактов из сельдерея и кабачка не вызвали их накопления.
Еще 40 лет назад А. Л. Курсанов установил, что процесс раневого залечивания сопровождается возрастанием дыхания. Субстратом возросшего дыхания в залечивающихся клубнях картофеля являлся крахмал, количество которого в тканях, прилегающих к месту поранения, постепенно уменьшалось, взамен чего возрастали моносахара, сгорающие в акте дыхания.
Процессы репарации нуждаются в источниках энергии. Показано, что в зоне поранения возрастало число митохондрий. Дополнительно образованные митохондрии генерировали энергию с такой же скоростью, как и предсуществующие в свеженарезанных клубнях. Очевидно, энергия, необходимая растениям для залечивания поранений, реализуется за счет дополнительно образовавшихся митохондрий.
В ответ на механическое поранение в клубнях картофеля происходило также образование нуклеиновых кислот. Показано, например, что через 10 минут после разрезания в бобе гороха в 10 раз возрастало количество рибонуклеиновой кислоты. В зоне поранения увеличивалось также и содержание аскорбиновой кислоты. К примеру, в раневой зоне клубня картофеля содержание аскорбиновой кислоты в 4—5 раз выше, чем ее количество в интактных клубнях.
Неоднократно предпринимались попытки установить корреляцию между устойчивостью клубней картофеля и толщиной их покровов. В конце концов исследователи вынуждены были признать, что такой зависимости не существует. Не обнаружено связи и между способностью клубней картофеля образовывать раневую перидерму и ее устойчивостью к ряду паразитарных микроорганизмов. По-видимому, образование перидермы на поверхности поранения клубней картофеля является второстепенным фактором в устойчивости клубней к ряду фитопатогенов, поскольку для ее образования требуется время, в течение которого паразит может проникнуть в клубень. Однако уже сформированная раневая перидерма вполне способна надежно защитить клубни от проникновения инфекции.
Еще сравнительно недавно считалось, что защитная роль перидермы по отношению к фитопатогенным микроорганизмам определяется только ее механическими свойствами. Позднее стало ясным, что перидерма является не только механическим, но и токсикологическим барьером на пути проникновения инфекции. Дело в том, что в зоне поранения накапливаются антибиотические вещества, которые подавляют проникновение инфекции. Эти вещества различной химической природы и, следовательно, обладают различным механизмом действия. Такие антибиотические вещества присутствуют как в составе раневой, так и естественной перидермы. Они как бы защитным чехлом покрывают растение и его органы снаружи. Вот, например, как располагаются ингибиторы в составе перидермы клубня картофеля: в наружных отмерших клетках феллемы находятся некоторые фенольные соединения, в том числе кофейная кислота. Под ними в меристематической ткани залегают высокотоксические гликоалкалоиды, а еще ниже располагается хлорогеновая кислота. Таким образом, проникающему паразиту, помимо механических покровов, приходится преодолевать несколько последовательно расположенных токсических барьеров из ингибиторов различной химической природы с различными спектрами антибиотического действия. К тому же расположение ингибитора в отмерших тканях наружных растительных покровов позволяет создать их чрезвычайно высокие локальные концентрации.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Помогаем Растениям, Перенесшим Стресс
Вредители, болезни и неправильный уход ослабляют растения, заставляют их страдать и испытывать стресс.
Одно из первоочередных правил ухода за растениями -проведение регулярного о смотра, в ходе которого можно заметить как радостные изменения в состоянии ваших зеленых питомцев -появление бутонов, так и неприятные -например, внезапное изменение окраски листьев. Эта процедура помогает вовремя обнаружить и устранить проблему, возникшую с самочувствием растения, которое, как и человек, в период болезни переживает стресс.
{LikeAndRead}
Такие разные
У каждого вида растений, которые вы культивируете у себя дома, собственные требования к температуре и уровню влажности воздуха, освещению, вентиляции и подкормке. К тому же режим полива варьируется в зависимости от времени года и размера вашего зеленого друга. Кроме того, повышение температуры требует увеличения объема полива и частоты проветриваний. Необходимо учитывать эти требования при уходе за вашими зелеными питомцами, среди которых наверняка есть и тропические экземпляры, и виды, в природных условиях растущие в странах с умеренным климатом. Иначе в течение нескольких месяцев одни из растений получат слишком много влаги и удобрений, а другие - слишком мало, да еще южные неженки могут пострадать от холода.
Вредители и инфекции
Если вовремя не принять меры, вредители и заболевания быстро распространятся в тесной компании растений. К примеру, тля в жаркие дни достигает половой зрелости в течение пяти дней, значит, всего за две недели пораженными могут оказаться все ваши зеленые друзья. Понять, болеет ли растение, иногда бывает достаточно сложно. Особенно если у него пестрые листья, как, например, у абутилона полосатого. Для того чтобы определить, является ли изменение окраски листьев обычным явлением или обусловлено болезнью, лучше воспользоваться лупой, с помощью которой можно рас смотреть вредителей -переносчиков инфекций (клещей, белокрылок, трипсов). Многие заболевания возникают при сочетании нескольких факторов, неблагоприятных для развития растений (излишняя влажность воздуха, чрезмерный полив и т. д.). Если не допускать создания таких условий, можно избежать негативных последствий для здоровья ваших зеленых друзей. Очень важно также вовремя удалять омертвевшие ткани (увядшие цветки, поблекшие листья и т. д.), в которых любят поселяться вредители.
Примите соответствующие меры
Время отпуска - период треволнений для цветовода. Ведь приятель, которому поручен уход за растениями, даже из лучших побуждений может случайно сделать что-то неверно и погубить их. Чтобы этого не случилось, принимают соответствующие меры.
Переезд
Переезд становится настоящим потрясением для растений, о которых просто забывают в суете.
ПРОФИЛАКТМЧЕСКИЕ МЕРЫ
Профилактика лучше, чем лечение
У многих растений есть предрасположенность к грибковым заболеваниям, например, некоторые виды роз очень страдают от настоящей мучнистой росы. Листьями и стеблями фуксин любит лакомиться тля,
радермахеры -мучнистые червецы. Необходимо заранее провести мероприятия, предотвращающие появление заболеваний и вредителей: обработать растения соответствующими инсектицидами, уменьшить уровень влажности в помещении и др.

Особенно страдают цветочные: из-за стресса они могут даже сбросить бутоны. Если вы перебираетесь на новую квартиру в холодную погоду, не забудьте обернуть бумагой (газетами) или картоном ваших зеленых друзей. Лиственным растениям может понадобиться время для адаптации. На новом месте такие разновидности стараются не перемещать хотя бы в течение одного месяца (если, конечно, они чувствуют себя хорошо).
Снятие стресса
Ослабленное растение (независимо от факторов, которые повлекли за собой стресс) легче поражается заболеваниями и вредителями. Чтобы ваш зеленый питомец поскорее поправился, ему нужно помочь снять стресс.

Исправление ошибок
Когда растение начинает болеть, необходимо выяснить, какие ошибки были допущены, и поскорее исправить сложившуюся ситуацию. Если все условия ухода четко соблюдались, нужно вспомнить, не менялась ли погода в течение последних нескольких недель. Однолетники реагируют на такие изменения незамедлительно, а реакция кустарников может проявиться позже. Из-за неблагоприятных условий однолетники, высаженные в открытый грунт, чаще всего погибают - у этих растений с неодеревенелыми побегами мало шансов на выживание. Неприхотливые кустарники могут сбросить листья из-за жары, пересыхания земляного кома и вследствие чрезмерного полива. Установив причину плохого самочувствия, растения по возможности пересаживают. Так, если вайи папоротников из-за недостаточной влажности воздуха засохли, их обрезают, а растения пересаживают. Можно сделать тепличку для восстановления ослабленных папоротников. Перед пересадкой луковицы луковичных (или клубневых) культур подсушивают, что особо важно для капризных лилий.

Нужные процедуры
Растениям, страдающим одновременно из-за неправильного ухода и поражения вредителями заболеваниями, помочь очень трудно. Дело в том, что экземпляры, ослабленные из-за неправильного ухода, нельзя обрабатывать препаратами, помогающими бороться с болезнями или вредителями. Поэтому сначала устраняют причину, вызвавшую стресс. В зависимости от вида растения проводят пересадку, обрезку, осушают компост или увеличивают объем и частоту полива. Подходящим инсектицидом - или фунгицидом зеленого друга обрабатывают, только когда восстанавливается его рост. При необходимости процедуру повторяют.

Стоит ли надеяться?
Растениям, которые уже несколько раз выхаживали, вряд ли удастся помочь: скорее всего, они истощили свой защитный потенциал, и в их тканях уже произошли необратимые изменения, негативно влияющие на способность восстанавливать тургор. На всякий случай можно еще раз убедиться, стоит ли тратить время на реабилитацию растения, перенесшего стресс. Для этого делают небольшой надрез (для однолетников достаточно неглубокой царапины) на коре стебля. Если под корой ткани зеленого или зеленовато-белого цвета, то не все еще потеряно, а если коричневого, с зеленым питомцем придется расстаться.
Когда ничего не помогло...
Иногда необходимо проявить нетерпимость по отношению к растениям, которым вы напрасно пытались помочь. Чахлые экземпляры не только плохо выглядят - они опасны для других зеленых друзей, поэтому от них лучше избавиться. Не стоит особо грустить по этому поводу: любое растение в какой-то момент достигает пика своего развития, после чего его выбрасывают.
{/LikeAndRead}
xn--80ai2adfh.com
Глава 7. ФИЗИОЛОГИЯ СТРЕССАСпособность к защите от повреждающих и неблагоприятных факторов среды – обязательное свойство любого, в том числе и растительного, организма. Ответные реакции, индуцируемые в организме внешними воздействиями, часто объединяют терминам «адаптационный синдром», а также широко распространившимся термином «стресс». Основные понятия и положения учения о стрессе были разработаны (в приложении к медицине) в 1936 г. канадцем Гансом Селье. Он полагал, что адаптивная реакция организма на различные неблагоприятные факторы (стрессоры) развивается по единому сценарию. Комплекс ответных реакций организма на стрессоры Г. Селье назвал, «генерализованным адаптационным синдромом», в котором он выделил три стадии (триады): 1) тревога и торможение большинства процессов; 2) адаптация, в течение которого организм приспосабливается к стрессору; 3) истощение, если адаптивный потенциал организма недостаточен для преодоления влияния стрессора (рис. 7.1).
По Г. Селье, стресс – это совокупность всех неспецифических изменений, возникающих в организме под влиянием любых неблагоприятных и повреждающих факторов (стрессоров). Селье считал, что одной из отличительных характеристик живых организмов является способность адаптироваться к стрессорам путем «концентрирования усилий, или напряжения». Предложенный Г. Селье термин «стресс» быстро завоевал популярность в физиологии, поскольку позволял одним словом объединить воздействие на организм разнокачественных повреждающих факторов без количественной оценки эффекта, вызываемого каждым из стрессоров в отдельности. При сопоставлении фаз триад у растений и животных наибольшим сомнениям подверглась идентичность первой фазы. Судя по доминирующим в ней реакциям, она не могла быть названа фазой тревоги. Ее предлагается именовать первичной стрессовой реакцией. Стрессовыми называют те внешние факторы, которые оказывают неблагоприятное воздействие на растение. В большинстве случаев стрессовое воздействие оценивают по его влиянию на выживание растительного организма, процессы роста, ассимиляции углекислоты или элементов минерального питания. Разные виды растений устойчивы (или неустойчивы) к различным стрессовым воздействиям, т. е. характер стрессового воздействия зависит и от вида (сорта) растения, и от стрессового фактора. Приобретение устойчивости под воздействием одного из неблагоприятных факторов может вызывать повышение устойчивости растительного организма к другим стрессовым воздействиям. Это явление называется кросс-устойчивостью, или кросс-адаптацией. В ответных реакциях растений на повреждающие факторы выделяют элементы неспецифической устойчивости (включающиеся в самых различных стрессовых ситуациях) и специфические процессы, инициируемые в растении только определенным типом стрессовых воздействий. На формирование неспецифических элементов устойчивости (синтез белков теплового шока, полиаминов) требуется гораздо меньшее время, чем для прохождения специфических адаптивных реакций, например, таких, как синтез белков-антифризов, фитохелатинов, переключение фотосинтеза на САМ-метаболизм. Растения очень часто подвергаются стрессовым воздействиям в естественных условиях. Некоторые стрессовые факторы, например высокая или низкая температура воздуха, могут действовать в течение нескольких минут, другие оказывают на растение неблагоприятное влияние в течение нескольких дней, недель (затопление) или месяцев (дефицит или избыток некоторых минеральных элементов). Именно устойчивость к неблагоприятным условиям среды определяет характер распределения различных видов растений по климатическим зонам. Большинство же сельскохозяйственных культур вынуждены постоянно находиться в стрессовых условиях, поэтому обычно реализуется только 20 % их генетического потенциала. Для эффективного выращивания большинства растений в условиях агрокультуры важна не только их потенциальная продуктивность, но и способность противостоять и адаптироваться к различным стрессовым ситуациям. Важное значение при этом имеет акклиматизация растений, т. е. увеличение их устойчивости к стрессовому фактору путем закаливания или серии подпороговых стрессовых воздействий. Процесс акклиматизации связан с экспрессией определенных генов и появлением таких генных продуктов, активность которых повышает устойчивость растения к стрессовому воздействию. Акклиматизацию не следует понимать как адаптацию, которая определяется как генетически детерминированный уровень устойчивости определенных видов растений и, как правило, является результатом процесса отбора в течение многих поколений. Адаптация – это совокупность морфологических, физиологических и химических приспособительных реакций, обеспечивающих возможность выживания определенного вида растений при действии неблагоприятных условий среды.^ Недостаток воды в растительных тканях возникает тогда, когда расход начинает превышать поступление. Засухи бывают почвенные и атмосферные. Атмосферная засуха характеризуется низкой относительной влажностью воздуха; почвенная засуха, как правило, следует за атмосферной и возникает при длительном отсутствии дождей. Водный дефицит вызывает серьезные нарушения в первую очередь процессов роста и фотосинтеза. Во время засухи наряду с обезвоживанием происходит перегрев растений, поэтому засуха является наиболее жестким стрессовым воздействием. В зонах, где среднегодовое количество осадков позволяет получать очень высокие урожаи сельскохозяйственных культур, главным фактором, лимитирующим рост растений, является дефицит воды. Более устойчивы к засухе такие виды растений, которые способны расходовать воду более экономно или запасать ее. У растительных организмов имеется несколько адаптивных стратегий, с помощью которых удается переживать засушливые периоды. Одни виды растений способны накапливать и удерживать воду, поддерживая, таким образом, необходимую степень гидратации своих клеток и органов (например, суккуленты, имеющие САМ-путь углеродного метаболизма). Другие приспособились функционировать в условиях дефицита воды в организме (ксерофиты, для которых, как правило, характерен С4-путь фотосинтеза). У некоторых видов растений в условиях засухи (или засоления) даже индуцируется переход фотосинтеза к САМ-пути углеродного метаболизма. К третьей группе относят растения, которые «избегают» засухи за счет организации своего жизненного цикла таким образом, чтобы вегетировать в период достаточного обеспечения водой. Многие виды растений резко ускоряют свой рост весной, когда в почве накапливается большое количество влаги, до наступления засушливого лета. У таких растений формируется большая листовая поверхность и быстро развивается мощная корневая система, которая выбирает всю влагу из почвы, накопившуюся в зимний период. При этом растения или быстро завершают свой жизненный цикл (эфемероиды), или запасают воду для завершения репродуктивного этапа развития. Такой тип роста и развития растений называют детерминированным, поскольку он определяется периодами дождей или засухи. При резком наступлении засушливого периода растения, как правило, сбрасывают часть листьев за счет формирования у черешка отделительного слоя под влиянием фитогормона этилена. Если водный дефицит развивается постепенно, то растительному организму лучше удается приспособиться к недостатку воды, заранее замедлив темпы ростовых процессов. Совсем другая стратегия развития у растений, произрастающих в условиях нормальной обеспеченности водой. В этом случае развиваются большие листья, а вегетация и цветение идут весь летний период. Такой тип развития растений называют недетерминированным. У растений с этим типом роста дефицит воды лимитирует не только размеры отдельных листьев, но и их количество. Одной из самых ранних реакций растительных организмов на водный дефицит является синтез фитогормона абсцизовой кислоты, которая вызывает закрывание устьиц и таким образом снижает потерю воды при транспирации. При водном дефиците транспорт ассимилятов из листьев угнетается в меньшей степени, чем их синтез. Недостаток воды инициирует рост корней в более влажные участки почвы при условии, что в растущие кончики корней поступает необходимое количество ассимилятов. В этом случае возникает конкуренция за ассимиляты между растущими корнями и формирующимися плодами. В условиях водного дефицита активируется синтез низкомолекулярных осмотически активных веществ (моно- и олигосахаридов, аминокислот, в первую очередь пролина, бетаина, многоатомных спиртов) и различных стрессовых белков, таких, как осмотин, дегидрины, а также белков, удерживающих воду и ионы. В состав стрессовых белков входят аквапорины – каналоформеры, которые облегчают водный транспорт через мембраны в осмотически зависимых ситуациях. У растений найдены два типа аквапорины – в плазматической и вакуолярной мембранах. Особое значение в осморегуляции имеет пролин, содержание которого при засухе резко возрастает. К функциональным белкам, способствующим устойчивости к засухе, относятся и многочисленные гидролазы, в том числе ферменты протеолиза, ингибиторы протеаз, ферменты биосинтеза осмотиков. При водном стрессе синтезируются также регуляторные белки, которые участвуют в экспрессии генов и сигнальной трансдукции. Пути приспособления растений к засухе приведены на рис. 7.2.
^ Под устойчивостью к низким температурам подразумевают холодостойкость и морозоустойчивость, т. е. способность растений выживать в условиях низких положительных температур и при заморозках. В листьях растений, повреждающихся при охлаждении, нарушаются процессы фотосинтеза, транспорта ассимилятов, дыхания, синтеза белков. Диапазон повреждающих температур сильно варьирует в зависимости от вида растения. Особенно устойчивы к низким температурам древесные виды в состоянии покоя. Семена, другие обезвоженные ткани растений и споры грибов способны переживать температуры около абсолютного нуля. Тургесцентные вегетирующие клетки также могут выдерживать замораживание, если их охлаждать очень быстро, поскольку формирующиеся в таких условиях кристаллы льда очень малы и не вызывают механических повреждений. Холодостойкость. Большинство видов тропических и субтропических растений не выдерживают не только заморозков, но даже низких положительных температур. Такие растения, как кукуруза, фасоль, рис, томаты, огурцы и хлопок, останавливаются в росте при температуре 10–15 °С и повреждаются, если температура падает ниже 10 °С. Холодостойкость теплолюбивых растений можно также повысить предпосевным закаливанием. Для этого наклюнувшиеся семена (например, огурцов, томатов) в течение нескольких суток выдерживают в чередующихся (через 12 ч) условиях низких положительных (около 5 °С) и более высоких (10–20 °С) температур. Таким же образом можно закалять и рассаду. Обычное время для закаливания, например, картофеля составляет 15 дней обработки низкими положительными температурами. После такой закалки растения картофеля хорошо переносят даже слабые заморозки. В процессе закаливания изменяется гормональный баланс растения, ростовые процессы затормаживаются, возрастает количество ненасыщенных жирных кислот, включаются механизмы защиты клеток от обезвоживания и замерзания. Основной причиной повреждающего действия низких положительных температур на теплолюбивые растения является нарушение функционирования клеточных мембран из-за их «затвердевания», связанного с фазовыми переходами жирных кислот, поскольку при достаточно низких температурах липидные бислои ведут себя, как твердые тела. При температуре выше фазового перехода структура бислоя сохраняется, однако при этом жирные кислоты «плавятся», в результате чего вращение и скручивание молекул происходит легче, чем при низких температурах. Затвердевание или «плавление» мембранных липидов зависит от состава содержащихся в них жирных кислот. Для насыщенных жирных кислот с длинной цепью характерны более высокие температуры фазовых переходов. Однако фазовые переходы в мембране индуцируются не только изменениями температуры. Они могут быть вызваны сдвигами рН и мембранного потенциала, двухвалентными катионами и гормонами. Различная реакция устойчивых и неустойчивых растений на низкие температуры определяется в первую очередь различиями в составе жирных кислот, входящих в состав мембранных фосфолипидов. Выявлено, что у холодостойких растений содержание ненасыщенных жирных кислот (таких, как линоленовая и линолевая) гораздо выше, чем у растений, чувствительных к холоду. Увеличение количества ненасыщенных жирных кислот в составе мембран приводит к снижению температуры фазового перехода мембранных липидов. У растений, чувствительных к холоду, в составе мембранных липидов велико содержание насыщенных жирных кислот (пальмитиновой, стеариновой). Мембраны такого типа стремятся к затвердеванию до квазикристаллического состояния уже при низких положительных температурах. При этом они становятся менее текучими, что нарушает функционирование многих белков-каналоформеров, переносчиков, рецепторов, ферментов и т. п. Схема влияния низких температур на мембраны от силы воздействия приведена на рис. 7.3.
Итак, холодостойкими называются растения, которые не повреждаются и не снижают своей продуктивности при температуре от 0 до +10 оС. Следует отметить, что холодостойкость свойственна и растениям умеренной зоны. Для большинства сельскохозяйственных культур низкие положительные температуры почти безвредны. Наиболее холодоустойчивыми являются яровые пшеница, ячмень, овес. Картофель, томаты, гречиха, просо, кукуруза, соя переносят температуры +5 оС и ниже без значительных повреждений. Фасоль, сорго, клещевина, арахис, рис, арбуз, дыня, тыква, огурцы, кабачки, перец – теплолюбивые растения, погибающие при температуре ниже +10 оС. В ходе холодового воздействия синтезируются стрессовые белки холодового шока. Так, в проростках озимой пшеницы трехчасовая гипотермия (+3 оС) индуцирует синтез большого набора стрессовых низкомолекулярных и гидрофильных белков. Морозоустойчивость. Заморозки вызывают повреждение клеток кристаллами льда, образующимися при температуре ниже 0 °С. Повреждающий эффект усугубляется также обезвоживанием тканей, которое при этом происходит. На первых этапах замерзания сильных повреждений растительных клеток не наблюдается, так как вначале кристаллы льда формируются в межклетниках и сосудах ксилемы. Если же заморозки длятся долго, вода переходит из цитоплазмы в апопласт, клетки обезвоживаются, кристаллы льда растут, и повреждают их. Для предотвращения замерзания в растительных клетках и тканях функционирует система антифризов, представленная различными белками, углеводами и гликопротеинами. Криопротекторы, синтезирующиеся в растительных клетках при понижении температуры, могут предотвратить или резко замедлить рост кристаллов льда. Гидрофильные белки, моно- и олигосахариды, обладающие криопротекторным эффектом, способны связывать значительные количества воды. Связанная таким образом вода уже не замерзает и не транспортируется. Белки и углеводы, обладающие криопротекторным эффектом, способны стабилизировать другие белки и клеточные мембраны при дегидратации клеток, инициированной низкими температурами. Криопротекторы начинают синтезироваться, прежде всего, в эпидермисе и клетках, окружающих межклеточные полости, где происходит наиболее интенсивное образование кристаллов льда при замерзании тканей. Важная роль в адаптации растений к низким температурам принадлежит фитогормону АБК. Устойчивость многих растений к заморозкам может быть повышена не только закаливанием, но также обработкой их АБК. При воздействии низких положительных температур и (или) при уменьшении светового периода концентрация этого гормона в растительных тканях резко возрастает. Мутанты арабидопсиса, нечувствительные к АБК (abi 1) или с нарушенным синтезом АБК (аbа 1), не могут приспособиться к заморозкам и не закаливаются. Активность многих генов и белков, которые экспрессируются при низкой температуре или водном дефиците, может быть индуцирована обработкой АБК. При изучении этих генов обнаружилось, что белки, появляющиеся при охлаждении тканей, имеют высокую степень гомологии с семейством белков RAB/LEA/DHN (responsive to ABA, late embryo abudant and dehydrin), синтезируются при созревании семян, водном дефиците и обработке растений АБК. Они очень гидрофильны и обладают необычным свойством: сохраняют свою структуру при резких колебаниях температуры. Продуктами генов LЕА являются гидрофильные белки, способные очень прочно удерживать воду. Считается, что именно эта группа белков обеспечивает защиту клетки при обезвоживании и заморозках, стабилизируя другие белки и клеточные мембраны. У морозостойких растений при действии низких температур усиливается гидролиз крахмала и накопление в цитоплазме растворимых cахаров. У озимых форм пшеницы и многих других видов растений функцию криопротектора выполняет сахароза, накапливающаяся в больших концентрациях. В качестве антифризов могут также выступать и другие растворимые сахара: рафиноза, фруктозиды, сорбит или маннит, которые накапливаются в клеточных стенках растений и таким образом предотвращают образование кристаллов льда. Наиболее чувствительны к морозу корни, корневища, клубни и луковицы большинства растений, которые повреждаются при температуре ниже –10 – –15°С. Покоящиеся почки древесных растений способны выдерживать морозы до –25 – –30 °С. У многолетних растений, устойчивых к заморозкам, в период подготовки к зиме в клубнях, луковицах и корневищах накапливаются запасные вещества, которые начинают использоваться с началом вегетационного периода.^ При повышении температуры выше оптимальной, при которой физиологические процессы идут с максимальной скоростью, наблюдается их снижение скорости вплоть до остановки. Только несколько видов высших растений могут постоянно существовать при температуре выше 45 °С. При высоких температурах способны выживать лишь нерастущие и обезвоженные клетки или ткани, например семена и пыльца. Активно вегетирующие ткани редко выживают при температурах более 45 °С, в то время как сухие семена и пыльца некоторых видов могут выдерживать температуру 120 и 70° С соответственно. К высоким температурам хорошо приспособились некоторые суккуленты, для которых характерен САМ-тип углеродного метаболизма. Поскольку у этих растений устьица днем закрыты, они не могут регулировать температуру за счет транспирации, но способны выдерживать перегрев до 60–65 °С. Тепловой стресс представляет потенциальную угрозу при выращивании растений в теплице, где в условиях высокой влажности и неподвижного воздуха плохо функционирует система естественного охлаждения растений путем транспирации. Одной из основных причин теплового шока, вызываемого высокими температурами, является потеря воды растительными тканями. Вероятно, поэтому продуктивность различных сортов сельскохозяйственных культур (например, хлопка и пшеницы) тем выше, чем больше устьичное сопротивление и соответственно меньше потери воды при транспирации. При перегреве растительных тканей в первую очередь повреждаются белки и мембраны. Повышается степень ненасыщенности жирных кислот, входящих в состав мембранных липидов, и их текучесть. Снижается способность к образованию водородных и ионных связей между полярными группами биополимеров. Происходит нарушение системы слабых взаимодействий, обеспечивающих поддержание пространственной структуры в белковых молекулах и мембранных липидах. Интегральные мембранные белки стремятся целиком перейти в гидрофобную фазу мембраны, что сказывается на процессах мембранного транспорта, рецепции, энергетике. В результате нарушения третичной структуры элементов электрон-транспортной цепи хлоропластов и митохондрий угнетаются процессы фотосинтеза и дыхания. Наиболее чувствителен к высоким температурам процесс фотосинтеза. Одним из основных элементов адаптации растительных организмов к перегреву являются белки теплового шока, которые синтезируются при повышении температуры и помогают растениям выдерживать этот тип стрессового воздействия. Впервые они были выявлены у дрозофилы, позже идентифицированы у животных (включая человека), микроорганизмов и растений. Если клетки или проростки растений быстро нагреть до температуры 40 °С, синтез большинства белков и мРНК будет подавлен. Именно в этих условиях происходит активация синтеза около 30–50 белков, называемых белками теплового шока (БТШ). Новые транскрипты БТШ, т.е. соответствующие мРНК, обнаруживаются уже через 3–5 мин после воздействия высоких температур. Их образование наблюдается также в естественных условиях при постепенном повышении температуры (рис. 7.4).
Для растительных организмов характерно накопление низкомолекулярных БТШ (от 15 до 31 кДа). После их синтеза отдельные клетки и целый растительный организм приобретают устойчивость к повышенным температурам и даже к таким, которые ранее были летальными. Нарушения в синтезе БТШ, как правило, лишают растения устойчивости к тепловому шоку. В экспериментах in vitro выявлено, что белковые фракции, обогащенные БТШ с молекулярной массой 15 и 18 кДа, обладают высокой устойчивостью к повышенной температуре и предотвращают денатурацию других белков при нагревании. Некоторые БТШ не имеют отношения к тепловому шоку, а инициируются другими типами стрессовых воздействий (засухой, низкими температурами, механическими повреждениями, засолением) и обработкой АБК. Таким образом, клетки, подвергшиеся воздействию одного вида стресса, приобретают устойчивость (через образование БТШ) к другим повреждающим факторам, т. е. происходит кросс-адаптация. Например, тепловой шок (48 ч при 38 °С) стимулирует образование ряда БТШ у томатов, что позволяет им без заметных повреждений выдерживать почти 3-недельное пребывание при температуре 2 °С. Многие БТШ обнаруживаются и в нестрессированных клетках. Различные виды БТШ и их гомологи обнаружены в ядре, митохондриях, хлоропластах, эндоплазматическом ретикулуме, цитоплазме. Выявлено, что БТШ с молекулярной массой 60, 70, 90 кДа и другие работают как молекулярные шапероны, обеспечивая посттрансляционное хранение, созревание (процессинг) и пространственную укладку (фолдиг) единичных белков, а также сборку олигомеров белковых молекул. Некоторые БТШ обеспечивают мембранный транспорт полипептидов в клеточные компартменты и защиту от протеазной активности. Ряд БТШ обладает способностью временно связываться с некоторыми ферментами и высвобождать их только на определенной стадии развития клетки, когда необходимо проявление энзиматической активности.^ В зависимости от типа анионов выделяют хлоридный, сульфатный, хлоридно-сульфатный и карбонатный типы засоления почв. Растения, приспособленные к существованию в условиях избыточного засоления, называют галофитами (от греч. gа1оs – соль). Галофиты (в отличие от гликофитов – растений незасоленных почв и водоемов) способны противостоять засолению. Галофиты способны нормально развиваться на растворах с содержанием хлорида натрия от 100 до 500 мМ (рис. 7.5).
Представление о способности культурных растений произрастать на почвах разной степени засоления дает табл. 7.1. |





userdocs.ru
Снятие стресса у растений — Центр чудес
Стресс у комнатных растений. Причины и средства борьбы
Стресс — сейчас очень модное и популярное слово. Оказывается, стресс могут испытывать не только люди, но и растения, причем он действует на них также разрушительно, как и на нас((.
Стресс— это общая реакция растения на действие любых неблагоприятных факторов.
Внешние признаки стресса у растений: прекращение роста, желтеют опадают листья, не завязываются и опадают бутоны, нарушается процесс корнеобразования у черенков и т.д.
Выделяют три основные группы факторов, вызывающих стресс у растений:
физические, к которым относят
-недостаточную или избыточную влажность,
-температуру (воздействие высоких или низких температур),
-травмирующие механические воздействия.
—ими являются различные соли,
-ксенобиотики (гербициды, инсектициды, фунгициды, промышленные отходы). И
Биологические, к которым относят
-вредителей, а также
-другие растения, если с ними приходится конкурировать.
Во многих случаях растения способны со стрессом справиться самостоятельно и адаптироваться к существующим условиям. Но это, к сожалению, происходит не всегда. Стресс может носить длительный характер и сильно подорвать силы растения. К моменту адаптации растение может прийти ослабленным, часто потерявшим привлекательный вид. Очевидно, следует помочь растению и стимулировать в нем основные адаптивные процессы.
К числу антистрессовых препаратов, которые могут помочь растению, относятся: Эпин Экстра, Циркон, Крезацин, Фитоспорин, Мивал, Планриз и другие. Эти препараты имеют иммуностимулирующую и антистрессовую активность.
Остановлюсь на некоторых из них.
1. Эпин-экстра — это прозрачный препарат с запахом спирта, действующее вещество препарата – эпибрассинолид (спиртовый раствор 0,25 г/л, в раствор добавляется шампунь для лучшей смачиваемости поверхности листьев). Является синтезированным аналогом природного вещества (производится по микробиологической технологии). Он работает как антистрессовый адаптоген: активирует собственные силы растения. При засухе опрыскивание эпином-экстра усиливает способность корня поглощать влагу, при избытке почвенной воды — увеличивает испаряющую способность листьев, при недостатке света — ускоряет и наращивает синтез хлорофилла. Применение его имеет смысл для повышения устойчивости растений перед ожидающим его стрессом. Например, стоит опрыскать сеянцы перед пикировкой или взрослое растение перед пересадкой или обрезкой. Замечено, что черенки после опрыскивания лучше укореняются. Обработку эпином-экстра следует производить примерно за сутки до намеченной работы.
Эпин корнями не всасывается, поэтому лить его в почву бессмысленно. Препарат практически не опасен для человека, теплокровных животных, рыб, пчел и других полезных насекомых. Не загрязняет окружающей среды. Класс опасности 3.
2. Циркон. Действующее вещество — гидроксикоричные кислоты, изготавливается из природного сырья — эхинацеи пурпурной. Препарат фирмы НЭСТ-М. Раствор продается в полипропиленовых ампулах в виде 0,1 г/л спиртового раствора с добавкой шампуня для лучшей смачиваемости листьев.
Циркон проявляет антистрессовую активность в условиях засухи, защищает клетки от УФ-излучения , эффективно помогает растениям восстановиться при пересадке (уменьшает транспирацию, повышает всасывание воды и питательных веществ). Препарат оказывает выраженное защитное действие против фитопатогенов различной природы (грибов и бактерий), есть данные, что препарат обладает противовирусным действием.
Циркон быстро впитывается листьями растений и корнями , но внутри растения перемещается медленно. Поэтому опрыскивать растение нужно полностью, добиваясь смачивания всех листьев. Остатки раствора есть смысл вылить в почву. К препарату нет привыкания. Нельзя допускать передозировку циркона.
Препарат практически не опасен для человека, теплокровных животных, рыб, пчел и других полезных насекомых. Не загрязняет окружающей среды. Класс опасности 4.
Рекомендации по применению эпина-экстра и циркона:
• Оба эти вещества разрушаются в щелочной среде. Поэтому для получения раствора нужно использовать только чистую кипяченую воду и подкислить ее лимонной (уксусной) кислотой.
• При использовании и циркона или эпина-экстра как стимуляторов корнеобразования к ним добавляют другие стимуляторы, но нужно иметь в виду, что с гетероауксином смешивать их можно, а с корневином нельзя.
• Эпин-экстра и циркон требуется хранить в темноте, обрабатывать ими растения лучше вечером (они разрушаются на свету).
• Препараты хранят сутки: они окисляются и приходят в негодность (окисляются кислородом, растворенным в воде). После вскрытия ампулы препарат лучше набрать в шприц (в инсулиновом шприце 40 капель).
Эпин разводится: 1 мл (40 кап.) на 5 литров воды или 8 капель на 1 л.
Циркон разводится: 1мл (40 кап.) на 10 литров воды или 4 капли на 1 л.
Нужно не забыть добавить в воду каплю лимонной или уксусной кислоты (можно каплю лимонного сока).
• Эпин усваивается и утилизируется растением за 14 суток, циркон всего за 18 часов.
• В отличие от эпина цирконом надо тщательно обрабатывать все растение, поскольку циркон передвигается по растению медленно, а эпин — быстро;
• В отличие от эпина циркон усваивается корнями, поэтому можно поливать землю его раствором.
tsvetiki.com
Снятие стресса у растений
- Травяная добавка
- Разработанный Майклом Тьерра, доктором восточной медицины, лицензированным иглотерапевтом
Stress Free от Planetary Herbals – это уникальное сочетание седативных, успокоительных и тонизирующих средств. Этот трехэтапный подход предоставляет внутреннюю поддержку против внешних стрессов.
По 1 таблетке три раза в день между приемами пищи.
Двухосновный фосфат кальция, стеариновая кислота, камедь акации, модифицированная камедь целлюлозы и диоксид кремния.
Не применять при беременности, планировании беременности или кормлении грудью.
Не употреблять, если какой-либо из изолирующих слоев упаковки поврежден или отсутствует. Хранить в месте, недоступном для детей.
Семена злаков, Черепковые аэрозольные части, Хроп-стробиль, Древесные бетонированные части, экстракт корня валерианы, экстракт корня женьшеня, экстракт из ягорей боярышника, экстракт корня лисицы, экстракт корня солодки, экстракт цветка ромашки, экстракт корня черной кохоши и экстракт корня элеутеро.
Отзыв оставлен aMIR3785 28 марта 2018
Отзыв оставлен Клиент iHerb 23 марта 2018
Отзыв оставлен Клиент iHerb 26 января 2018
— Бонус лояльности равен 5% от стоимости текущего заказа в iHerb, не включая стоимость доставки, и доступен для использования в следующем заказе после отправления текущего заказа. — При размещении каждого нового заказа, Вам будет начисляться бонус лояльности равный 5% от стоимости заказа. — Бонус лояльности истекает через 90 дней после последней покупки в iHerb.
Товары с пометкой «Эконом-доставка» имеют меньшие затраты на доставку, таким образом, мы можем предложить Вам экономить больше! Это означает, что при добавлении товара с отметкой «Эконом-доставка», Ваша стоимость доставки будет уменьшаться.
Обеспечение качества, прозрачности и лучших условий можно увидеть во всех домашних брендах iHerb, которые производятся и упаковываются исключительно в США или Канаде. Квалифицированные сырьевые материалы поставляются со всего мира и соответствуют самому высокому уровню стандартов индустрии:
Этап 1 — Сырье — на идентичность, чистоту и действие
Этап 2 — Производство — Тестирование процесса обеспечения качества
Этап 3 — Готовая продукция — соответствие этикетки и требованиям безопасности для потребителей
Лаборатории компании и сторонние лаборатории следуют принципам НЛД (Надлежащая лабораторная практика)
Наши производители полность соответствуют стандартам НЛД (Надлежащая лабораторная практика, 21 CFR часть 111), и зарегистрированы в NSF и/или NPA. Некоторые из производителей так же могут иметь дополнительные сертификаты, такие как производство лекарственных средств OTC и сертификаты органической продукции.
Если Вы не полностью удовлетворены Вашим товаром, Вы можете вернуть его в течении 90 дней с момента покупки и Вы получите 100% гарантию возврата денег.
ru.iherb.com
Кафедра физиологии и биохимии растений СПбГУ является одним из первых научных центров, где в начале 1960-х годов под руководством профессора С.В.Солдатенкова были начаты исследования особенностей обмена веществ растений в условиях недостатка кислорода. Эти работы были продолжены его ученицей Т.В.Чирковой в направлении изучения физиолого-биохимических механизмов адаптации растений к гипо- и аноксии.
В 1986 году под ее руководством на кафедре была создана научно-исследовательская группа «Физиология стресса и адаптации растений». В работе группы принимают участие студенты, аспиранты и сотрудники кафедры. В группе защищена 1 докторская, 11 кандидатских и более 80 студенческих работ (дипломных, выпускных квалификационных и магистерских). Опубликовано 225 научных работ, в том числе несколько монографий. Исследования механизмов адаптации проводятся группой преимущественно на двух модельных объектах: проростках пшеницы и риса, контрастных по устойчивости к недостатку кислорода. Кроме того, для изучения конкретных механизмов гормональной регуляции активно привлекаются мутантные и трансгенные растения резуховидки и табака, а для проведения эколого-физиологических исследований – представители дикорастущей гидро- и гелофитной флоры. Эксперименты по изучению механизмов перцепции и трансдукции анаэробного сигнала проводятся на клеточном уровне: на протопластах и клеточных культурах. Члены группы владеют современными методами физиологического, биохимического, молекулярно-биологического и микроскопического анализа растительного материала.
В результате многолетних исследований влияния недостатка или отсутствия кислорода на особенности дыхания, фотосинтеза, белкового, липидного, гормонального обменов впервые предложена и экспериментально обоснована оригинальная концепция физиолого-биохимических основ адаптации растений к гипо- и аноксии, что представляет собой существенный вклад в разработку общей теории стресса у растений. Она базируется на представлении о том, что длительная устойчивость определяется сформированной в процессе эволюции способностью растений к координации метаболических защитных реакций с помощью комплекса систем регуляции на организменном, межклеточном, и внутриклеточном уровнях. Было установлено также, что растения, устойчивые к гипо- и аноксии, способны лучше противостоять также и окислительному пост-аноксическому стрессу.
Рис.1. Схема путей приспособления растений к гипо- и аноксии (Чиркова, 1988, 1998, 2002).
Показано, что для устойчивых растений характерна меньшая интенсивность ростовых процессов. Удлинение корней останавливается под действием анаэробиоза у всех растений, а побегов – только у неустойчивых. Изучение зон роста корня обнаружило меньшую чувствительность к аноксии процесса деления клеток по сравнению с растяжением. В анаэробной среде выявлены различия в сдвигах гормонального баланса у растений, контрастных по устойчивости к кислородной недостаточности: продемонстрировано преобладание абсцизовой кислоты у чувствительного растения и индолилуксусной – у устойчивых. Такие изменения гормонального статуса вызывают остановку роста у неприспособленных растений, тогда как накопление ауксина в надземной части проростков устойчивого риса позволяет сохранять ее рост, а в корнях, наоборот, ингибирует его.
В настоящее время активно изучается роль так называемых “новых” гормонов растений (полиаминов, брассиностероидов, салици- ловой и жасмоновой кислот) в регуляции адаптационных процессов растений при дефиците кислорода и последующей ре-аэрации. В работе группы большое внимание уделено исследованию окислительного стресса, возникающего сразу после выхода растений из анаэробной среды в условия аэрации, и, в частности, роли деструктивных окислительных реакций и защитных антиоксидантных систем.
Рис.2. Фотографии традиционных экспериментальных объектов – проростков риса (слева) и пшеницы (справа).
Впервые показано, что под влиянием аноксии и особенно в пост-аноксический период происходит снижение содержания восстановленных форм низкомолекулярных антиоксидантов (аскорбиновой кислоты, глутатиона, каротина) и возрастание их окисленных форм. Сделан принципиальный вывод о том, что снижение уровня восстановленных форм антиоксидантов может служить одним из возможных редокс-сигналов аноксического повреждения тканей, подвергнутых затем и окислительному стрессу. Преобладание прооксидантов над антиоксидантами свидетельствует о переходе от аноксического к окислительному стрессу. Более того, соотношение прооксидантов и антиоксидантов может лежать в основе внутриклеточной редокс-регуляции и восприятия уровня кислорода. Изучение продукции активных форм кислорода (супероксидный анион-радикал, пероксид водорода), а также комплекса антиоксидантных ферментов (гваякол-пероксидаза, аскорбатпероксидаза, глутатион-пероксидаза, каталаза) впервые позволило доказать, что растения, устойчивые к кислородной недостаточности, способны эффективно противостоять и окислительному стрессу именно благодаря хорошо работающей антиоксидантной системе.
В последнее время группой активно разрабатываются и способы восприятия и передачи гипо- и аноксических сигналов в растительной клетке, темы почти не изученной, но совершенно необходимой для понимания механизмов адаптации растений к гипо- и аноксии Результаты исследования начальных этапов трансдукции анаэробного сигнала показало большую интенсивность его у устойчивого растения по сравнению с неустойчивым. Аккумуляция важнейшего участника процесса трансдукции – ионов цитоплазматического кальция под действием аноксии в протопластах из проростков риса была также больше, чем в протопластах пшеницы, и происходила за счет участия как внутриклеточных, так и внеклеточных депо ионов кальция. Недавно получены данные о том, что активные формы кислорода в субтоксических концентрациях также могут участвовать в трансдукции анаэробного сигнала у растений, однако работ в этом направлении почти нет. В связи с этим в настоящее время проводятся эксперименты по получению культуры клеток риса и пшеницы, синтезирующих генетически кодируемые флуоресцентные внутриклеточные белковые сенсоры для измерения уровня активных форм кислорода и ионов кальция. Подобран оптимальный состав питательных сред для индукции каллусообразования и получены каллусные культуры риса и пшеницы. Каллусная культура пшеницы переведена в суспензионную культуру для выполнения дальнейших экспериментов по получению трансгенных клеточных линий. Для создания трансгенных клеточных линий пшеницы и риса было апробировано несколько методов агробактериальной трансформации, среди которых был выбран наиболее оптимальный. С помощью данного метода были получены трансгенные каллусы риса и пшеницы, селекцию которых проводили на среде, содержащей канамицин. После завершения этапа первичной селекции трансгенная природа полученных каллусов была подтверждена с помощью ПЦР-анализа.
Рис.3. Микрофотографии протопластов, полученных из побегов риса (сверху) и пшеницы (снизу), загруженных Ca-чувствительным флуоресцентным красителем Fura2-AM. Слева – протопласты в проходящем свете, справа – флуоресценция (возбуждение при 436 нм, эмиссия – при 510-550 нм). Масштабная линейка – 20 мкм (Yemelyanov et al., 2011).
Основные темы исследований, предлагаемые студентам и аспирантам для работы в группе «Физиология стресса и адаптации растений»:
Окислительные процессы в растениях в условиях аноксии и последующего окислительного стресса.
Молекулярные механизмы антиоксидантной защиты растений в условиях недостатка кислорода и последующего окислительного стресса
Трансдукция аноксического сигнала в растительных клетках.
Роль фитогормонов в регуляции адаптации растений к недостатку кислорода и последующему окислительному воздействию.
Вторичный обмен растений в условиях аноксии и последующего окислительного стресса.
Скрининг устойчивых к недостатку кислорода форм сельскохозяйственных растений.
Сотрудничество с другими научными организациями:
Группа проводит комплексные научные исследования в сотрудничестве как с другими подразделениями СПбГУ, так и с научными учреждениями Санкт-Петербурга (Ботанический институт РАН), России (Казанский институт биохимии и биофизики РАН, Институт биохимии и генетики Уфимского НЦ РАН) и зарубежных стран (кафедра физиологии растений университета Хельсинки, кафедра ботаники Стокгольмского университета и кафедра физиологии растений технического университета Ахена).
С университетом Хельсинки (Helsinki University, Department of Biosiences, Division of Plant physiology – Ass. Prof. Fagerstedt K.V., Dr. Blokhina O.B.) в течение ряда лет проводятся совместные исследования по проблеме анаэробиоза в рамках международного договора между университетами Санкт-Петербурга и Хельсинки. В университете Хельсинки проводили экспериментальные работы аспиранты кафедры Блохина О.Б. и Куличихин К.Ю., на основании которых были защищены кандидатские диссертации.
Совместная работа со Шведским сельскохозяйственным университетом в Упсале (Swedish Agricultural University, Department of Plant Biology and Forest Genetics (Uppsala), Prof. Lindberg S.), а затем со Стокгольмским университетом (University of Stockholm,
Рис.4. Фотографии каллусов, полученных из проросших зерновок риса.
Department of Botany, Prof. Lindberg S.), проводилась доцентом Емельяновым В.В. по теме «Трансдукция аноксического сигнала растительными клетками». Проект финансировался Шведским институтом и Министерством образования и науки РФ. На протопластах клеток пшеницы и риса показано участие ионов кальция и калия в трансдукции анаэробного сигнала. Совместную работу с Университетом Западной Австралии (School of Biomedical, Biomolecular and Chemical Sciences, Faculty of Life and Physical Science, The University of Western Australia) проводил аспирант Куличихин К.Ю. Экспериментальные данные оформлены в виде статьи, опубликованной в J. of Exp. Botany, 2009.
В 2010 г. доцентом Емельяновым В.В. проведена совместная работа с кафедрой физиологии растений Технического университета г. Ахена, Германия (Rheinisch-Westfalische technische Hochschule Aachen, Institut fur Biologie III, Pflanzenphysiologie). В результате ее изучен эндогенный уровень салициловой кислоты в растениях пшеницы и риса в условиях аэрации, аноксии и последующей реаэрации. Работа финансировалась DAAD и Министерством образования и науки РФ. На основании полученных результатов готовится к публикации статья.
Научная работа группы «Физиология стресса и адаптации растений» неоднократно поддерживалась грантами различных отечественных и международных организаций. Научные сотрудники, аспиранты и студенты регулярно получают гранты для молодых учёных правительства Санкт-Петербурга. Участие сотрудников группы в международных симпозиумах также неоднократно поддерживалось отечественными и зарубежными фондами. В настоящее время работа финансируется грантом Министерства образования и науки в рамках реализации ФЦП «Научные и научно-педагогические кадры инновационной России» на 2009-2013 годы (ГК 2010-1.3.2-203-002-008). Члены группы «Физиология адаптации растений» состоят в Санкт-Петербургском обществе естествоиспытателей природы, Скандинавском и Всероссийском Обществах физиологов растений, Федерации Европейских обществ биологов растений и Международном обществе по анаэробиозу растений.
Чиркова Т.В. Роль клеточных мембран в устойчивости растений к недостатку кислорода // Успехи соврем. биол. 1983. Т. 95. №1. С. 44-56.
Чиркова Т.В. Пути адаптации растений к гипоксии и аноксии. Л., Изд-во Ленингр. ун-та. 1988. 244 с.
Чиркова Т.В. О путях приспособления растений к гипоксии и аноксии. // Физиология растений. 1988. Т. 35. №2. С. 393-411.
Чиркова Т.В., Жукова Т.М. Способ оценки устойчивости растений к недостатку кислорода. Авт. Свид-во на изобретение, № 1565403. Бюлл.№19.1990.
Чиркова Т.В. Клеточные мембраны и устойчивость растений к стрессовым воздействиям. // Соросовский Образовательный Журнал. 1997. №9. С. 12-17.
Чиркова Т.В., Иванова Т.И., Маслов Ю.И., Магомедов И.М. и др. Практикум по фотосинтезу и дыханию растений. Учеб. пособие / Под ред. В.В.Полевого и Т.В.Чирковой. СПб., Изд-во С-Петербург. ун-та. 1997. 162 с.
Чиркова Т.В. Растение и анаэробиоз. // Вестник СП6ГУ.1998. Сер. З. Вып. 2 (№10). С. 41-52.
Чиркова Т.В., Новицкая Л.О., Блохина О.Б. Перекисное окисление липидов и активность антиоксидантных систем при аноксии у растений с разной устойчивостью к недостатку кислорода. //Физиология растений. 1998. Т. 45. №1. С. 65-73.
Чиркова Т.В., Войцековская С.А. Синтез белка в растениях в условиях гипо- и аноксии. // Успехи современной биологии. 1999. Т. 119. №2. С. 178-189.
Полевой В.В., Чиркова Т.В., Лутова Л.А. и др. Практикум по росту и устойчивости растений. Учеб. пособие / Под ред. В.В.Полевого и Т.В.Чирковой. СПб., Изд-во С.-Петерб. ун-та. 2001. 212 с.
Семихатова О.А., Чиркова Т.В. Физиология дыхания растений. Учеб. пособие. СПб. Изд-во С.-Петерб. ун-та. 2001. 224 с.
Чиркова Т.В. Физиологические основы устойчивости растений. Учеб. пособие. СПб. Изд-во С.-Петерб. ун-та. 2002. 244 с.
Емельянов В.В., Кирчихина Н.А., Ласточкин В.В., Чиркова Т.В. Гормональный баланс проростков пшеницы и риса в условиях аноксии. // Физиология растений. 2003. Т. 50. С. 922-929.
Куличихин К.Ю., Чиркова Т.В., Фагерстедт К.В. Активность ферментов биохимического рН-стата в кончиках корней злаков в условиях кислородной недостаточности. // Физиология растений. 2009. Т. 56. №3. С. 418-430.
Состав группы «Физиология стресса и адаптации растений» в 2007 г.
Слева направо: студ. Арешева О.М., проф. Чиркова Т.В., студ. Шикова К.К., студ. Жминько О.С., доц. Емельянов В.В.
Состав группы в 2011 г.
Слева направо, верхний ряд: студ. Ма Г., доц. Емельянов В.В., н.с. Ласточкин В.В., нижний ряд: асп. Злотина М.М., проф. Чиркова Т.В.
www.bio.spbu.ru
centrchudes.ru
Стрессы растений - система защиты
Стрессом для растения (и всех других живых организмов) является резкие изменение внешних условий – температуры, pH, ионизации и т.д. В ответ на это живые существа реагируют специальным инструментом защиты – стрессовыми белками.
Крайне интересно, что в процессе эволюции практически не прослеживается изменения структуры этих белков и даже механизма запуска их образования в организме. То есть они одни и те же у грибов, растений, моллюсков, пауков, рыб, человека…
Стрессовые белки очень разнообразны и, соответственно, выполняют разную работу. Одна группа (белки теплового шока) обеспечивает выживание в условиях повышенной температуры. Их описано более ста. Эти белки препятствуют разрушению сложных белковых комплексов, противодействуют слипанию развернувшихся белковых глобул. Другая группа (белки синикации) связывает воду, обеспечивая высыхание семян в частности и оводненность тканей вообще. Именно эти белки определяют устойчивость растений к осмотическому и солевому стрессу, резким потерям воды при усиленной транспирации. Одновременно эти белки снижают температуру замерзания воды в клетках растений или, если порог кристаллизации уже пройден, способствуют образованию аморфного льда, не повреждающего клеточные мембраны.
Стрессовые белки начинают образовываться в организме уже через несколько минут после начала стресса и достигают максимальной концентрации примерно через 2 часа. Их уровень, обеспечивающий защиту организма, сохраняется в течение примерно 48 часов.
В практике растениеводства давно используется закаливание. Этот прием позволяет, постепенно изменяя силу воздействия внешних факторов, подготовить растение к пересадке в более жесткие условия выращивания. Физиологическая суть закаливания заключается именно в постепенном повышении уровня стрессовых белков, что обеспечивает выживание при изменении условий. Гормоны растений тоже умеют стимулировать образование стрессовых белков. При этом цитокинины и брассиностероиды повышают преимущественно уровень белков теплового шока, а абсцизовая кислота – белков синикации.
Стрессовые воздействия часто сопровождаются взрывной активацией окислительных процессов, приводящих к выбросу большого количества свободных радикалов. Это вызывает повреждение клеточных мембран. У растений есть соответствующий механизм защиты, поскольку в большинстве растительных тканей существует собственный значительный запас антиоксидантов, в основном – в виде витаминов Си Е.
 Стрессы растений
Стрессы растенийОцените статью:
[Всего голосов: 0 Средний: 0/5]dachniki.info










