Фотосинтез и первичная биологическая продуктивность. Фотосинтез растений физиология
Физиология растений Фотосинтез Жизненно важные процессы
 Физиология растений Фотосинтез
Физиология растений Фотосинтез
 Жизненно важные процессы
Жизненно важные процессы

 Схема физиологических процессов в растении
Схема физиологических процессов в растении
 Фотосинтез Это процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов. (у растений - хлорофилл) Фотосинтез происходит на уровне клеток и идет в хлоропластах – особых частях клетки. Фотосинтез состоит из 2 процессов: 1) Накопление энергии за счет ее запасания из солнечного света 2) Синтез органических веществ (сахаров) за счет накопленной энергии.
Фотосинтез Это процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов. (у растений - хлорофилл) Фотосинтез происходит на уровне клеток и идет в хлоропластах – особых частях клетки. Фотосинтез состоит из 2 процессов: 1) Накопление энергии за счет ее запасания из солнечного света 2) Синтез органических веществ (сахаров) за счет накопленной энергии.
,") Строение хлоропластов В клетке растения обычно 10 -20 хлоропластов – крупных органоидов (органов клетки), легко отличимых по зеленому цвету. Когда-то хлоропласты были свободными бактериями, но стали жить вместе с растительными клетками и сейчас уже не могут выжить сами.
Строение хлоропластов В клетке растения обычно 10 -20 хлоропластов – крупных органоидов (органов клетки), легко отличимых по зеленому цвету. Когда-то хлоропласты были свободными бактериями, но стали жить вместе с растительными клетками и сейчас уже не могут выжить сами.
 Фотосинтез
Фотосинтез

 Где больше всего фотосинтезирующих растений?
Где больше всего фотосинтезирующих растений?
present5.com
| Согласно законам фотохимии, при поглощении кванта света атомом или молекулой какого-либо вещества электрон переходит на другую, более удаленную орбиталь, т. е. на более высокий энергетический уровень. Наибольшей энергией обладает электрон, отдаленный от ядра атома и находящийся на достаточно большом расстоянии от него. Вместе с тем, чем ближе к ядру, тем меньше энергия электрона. Каждый электрон переходит на более высокий энергетический уровень под влиянием одного кванта света, если энергия этого кванта равна разнице между этими энергетическими уровнями. Все фотосинтезирующие организмы содержат какой-либо тип хлорофилла. Исключением являются бактерии (Halobacterium halobium), содержащие бактериородопсин, с помощью которого они поглощают кванты света.
В молекуле хлорофилла два уровня возбуждения. Именно с этим связано и то, что он имеет две основные линии поглощения. Первый уровень возбуждения обусловлен переходом на более высокий энергетический уровень электрона в системе сопряженных двойных связей, а второй — с возбуждением неспаренных электронов атомов азота и кислорода в порфириновом ядре. При поглощении света электроны переходят в колебательное движение. Наиболее подвижными в молекуле являются делокализованные электроны, орбитали которых как бы размазаны, обобщены между двумя ядрами. Особенно легко возбуждаются электроны сопряженных двойных связей. Электрон, кроме того, что он находится на определенной орбитали и вращается вокруг ядра, обладает еще спином (вектором магнитного момента) — характеристикой, которую можно трактовать как направление вращения электрона вокруг своей оси. Спин электрона может принимать два значения. Спины двух электронов, находящихся на одной орбитали, противоположны. Когда в молекуле все электроны расположены попарно, их суммарный спин равен нулю. Это основное синглетное состояние (S0). В основном энергетическом состоянии S0 молекула находится в тепловом равновесии со средой, все электроны попарно занимают орбитали с наименьшей энергией. При поглощении света электроны переходят на следующие орбитали с более высоким энергетическим уровнем. При этом имеются две возможности: если электрон не меняет спина, то это приводит к возникновению первого и второго синглетного состояния. Если же один из электронов меняет спин, то такое состояние называют триплетным. Наиболее высокий энергетический уровень — это второй синглетный уровень. Электрон переходит на него под влиянием сине-фиолетовых лучей, кванты которых содержат больше энергии. В первое возбужденное состояние электроны могут переходить, поглощая более мелкие кванты красного света. Из возбужденного, первого синглетного и триплетного состояния молекула хлорофилла также может переходить в основное. При этом ее дезактивация (потеря энергии) может проходить: 1) путем выделения энергии в виде света (флуоресценция и фосфоресценция) или в виде тепла; 2) путем переноса энергии на другую молекулу пигмента; 3) путем затрачивания энергии на фотохимические процессы (потеря электрона и присоединение его к акцептору. В любом из указанных случаев молекула пигмента дезактивируется и переходит на основной энергетический уровень. Энергия, испускаемая в виде флуоресценции или в виде теплоты, не может быть использована.
В настоящее время показано, что хлорофилл имеет две функции — поглощение и передачу энергии. При этом основная часть молекул хлорофилла — более 90% всего хлорофилла хлоропластов входит в состав светособирающего комплекса (ССК). Светособирающий комплекс выполняет роль антенны, которая эффективно поглощает свет и переносит энергию возбуждения к реакционному центру. Кроме большого числа (до нескольких сотен) молекул хлорофилла ССК содержит каротиноиды, а у некоторых водорослей и цианобактерий фикобилины. Каротиноиды и фикобилины увеличивают эффективность усвоения света за счет того, что они поглощают свет в тех областях спектра, в которых молекулы хлорофилла поглощают свет сравнительно слабо. Такое устройство позволяет значительно полнее использовать энергию света. Подсчитано, что каждая молекула хлорофилла на прямом солнечном свету поглощает квант света не чаще чем 10 раз в секунду. Между тем скорость последующих реакций фотосинтеза значительно больше. Из сказанного следует, что при непосредственной связи каждой молекулы хлорофилла с последующей реакцией процесс фотосинтеза шел бы прерывисто. Необходимо учитывать также, что использование энергии света в химических реакциях требует большого количества ферментов. Если бы каждая молекула хлорофилла отдавала энергию света непосредственно на фотохимические процессы, то в листе не хватило бы места для размещения всех необходимых для этого ферментных систем. В процессе эволюции в растениях выработался механизм, позволяющий наиболее полно использовать кванты света, падающие на лист подобно каплям дождя. Механизм этот заключается в том, что энергия квантов света улавливается 200—400 молекулами хлорофилла и каротиноидами ССК и как бы стекается к реакционному центру. Передача энергии между молекулами пигментов идет главным образом резонансным путем, без разделения зарядов с большой скоростью. Так, время переноса энергии от одной молекулы хлорофилла к другой составляет 10-12 — 10-12 с, а от молекулы каротиноидов к хлорофиллу 4 x 10-10 с. Таким образом, время переноса энергии значительно меньше времени жизни возбужденной молекулы (10-8 с). Такой перенос может осуществляться только при близком расстоянии между молекулами пигментов.
Расчеты показали, что в одном хлоропласте до 1 млрд молекул хлорофилла. Расстояние между молекулами хлорофилла в мембранах составляет всего 1 нм. Перенос энергии происходит только от пигментов, поглощающих свет с меньшей длиной волны, к пигментам, поглощающим свет с большей длиной волны. Дело в том, что хотя передача энергии от одной молекулы пигмента к другой идет с большой эффективностью (от хлорофилла b к хлорофиллу а — 90%, от каротиноидов к хлорофиллу — 40%), однако все же это связано с некоторой ее потерей. Вместе с тем кванты света с меньшей длиной волны обладают большей энергией. Потеря энергии приводит к превращению квантов в более мелкие (с большей длиной волны). Именно поэтому основные формы хлорофилла, к которым стекается энергия, являются более длинноволновыми. Обратный перенос энергии невозможен. В том случае, если молекулы пигментов имеют одинаковый максимум поглощения возможен другой путь передачи энергии, получивший название экситонного. При этом пути электронное возбуждение распространяется в результате межмолекулярной вибрации на комплекс пигментов. Строение ССК зависит не только от систематического положения фотосинтезирующего организма, но и от условий произрастания. Теневыносливые растения имеют, как правило, больший размер ССК по сравнению с растениями, растущими в условиях высокой освещенности. Таким образом, в первичных процессах фотосинтеза, связанных с поглощением молекулой хлорофилла кванта света, важную роль играют процессы передачи энергии. Фотофизический этап фотосинтеза и заключаемая в том, что кванты света поглощаются и переводят молекулы пигментов в возбужденное состояние. Затем эта энергия переносится на реакционный центр, осуществляющий первичные фотохимические реакции: разделение зарядов. Дальнейшее превращение энергии света в химическую энергию проходит ряд этапов, начиная с окислительно-восстановительных превращений хлорофилла и включая как фотохимические (световые), так и энзиматические (темновые) реакции. |



fizrast.ru
Реферат Фотосинтез
скачатьРеферат на тему:
План:
- Введение
- 1 Типы фотосинтеза
- 1.1 Бесхлорофильный фотосинтез
- 1.2 Хлорофильный фотосинтез
- 1.2.1 Аноксигенный
- 1.2.2 Оксигенный
- 2 Пространственная локализация
- 3 Световая (светозависимая) стадия
- 3.1 Фотохимическая суть процесса
- 3.2 Светособирающие комплексы
- 3.3 Основные компоненты электронтранспортной цепи
- 3.3.1 Фотосистема II
- 3.3.2 b6f или b/f-комплекс
- 3.3.3 Фотосистема I
- 3.4 Циклический и псевдоциклический транспорт электрона
- 4 Темновая стадия
- 4.1 С3-фотосинтез, цикл Кальвина
- 4.2 С4-фотосинтез
- 4.3 САМ фотосинтез
- 5 Значение фотосинтеза
- 6 Изучение фотосинтеза
- 7 Прочие факты ЛитератураПримечания
Введение
Лист растения
Фотосинтез (от греч. φωτο- — свет и σύνθεσις — синтез, совмещение, помещение вместе) — процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл и бактериородопсин у бактерий). В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества.
1. Типы фотосинтеза
1.1. Бесхлорофильный фотосинтез
Осуществляется археями рода Halobacterium, является наиболее примитивным типом фотосинтеза, кванты света поглощаются белком-бактериородопсином, имеющим сходство с родопсином в виде наличия ретиналя, этот тип фотосинтеза отличается отсутствием электрон-транспортной цепи,синтез АТФ осуществляется через создание электрохимического градиента протонов или ионов хлора при помощи бактериородопсиновой и галородопсиновой ионной помпы.
1.2. Хлорофильный фотосинтез
1.2.1. Аноксигенный
Осуществляется пурпурными и зелёными бактериями, а также геликобактериями.
1.2.2. Оксигенный
Оксигенный фотосинтез распространён гораздо шире. Осуществляется растениями, цианобактериями и прохлорофитами.
Этапы фотосинтеза:
- фотофизический
- фотохимический
- химический
На первом этапе происходит поглощение квантов света пигментами, их переход в возбуждённое состояние и передача энергии к другим молекулам фотосистемы. На втором этапе происходит разделение зарядов в реакционном центре, перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом АТФ и НАДФН. Первые два этапа вместе называют светозависимой стадией фотосинтеза. Третий этап происходит уже без обязательного участия света и включает в себя биохимические реакции синтеза органических веществ с использованием энергии, накопленной на светозависимой стадии. Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюкогенез, образование сахаров и крахмала из углекислого газа воздуха.
Первые процессы фотосинтеза у цианобактерий появились ещё в архейскую эру.
2. Пространственная локализация
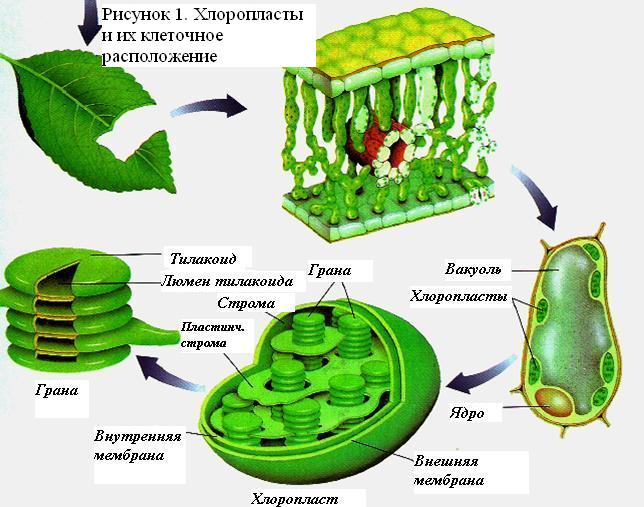
Хлоропласты в клетках листа
Фотосинтез растений осуществляется в хлоропластах: обособленных двухмембранных органеллах клетки. Хлоропласты могут быть в клетках плодов, стеблей, однако основным органом фотосинтеза, анатомически приспособленным к его ведению, является лист. В листе наиболее богата хлоропластами ткань палисадной паренхимы. У некоторых суккулентов с вырожденными листьями (например, кактусы) основная фотосинтетическая активность связана со стеблем.
Свет для фотосинтеза захватывается более полно благодаря плоской форме листа, обеспечивающей большое отношение поверхности к объёму. Вода доставляется из корня по развитой сети сосудов (жилок листа). Углекислый газ поступает отчасти посредством диффузии через кутикулу и эпидермис, однако большая его часть диффундирует в лист через устьица и по листу по межклеточному пространству. Растения, осуществляющие С4 и CAM фотосинтез сформировали особые механизмы для активной ассимиляции углекислого газа.
Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые соединяясь друг с другом образуют тилакоиды, которые в свою очередь группируются в стопки, называемые граны. Внутритилакоидное пространство отделено и не сообщается с остальной стромой, предполагается также что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (70s типа), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их предшественниками свободных цианобактерий, вошедших в состав эукариотической клетки в процессе симбиогенеза.
Цианобактерии, таким образом, как бы сами являются хлоропластом и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды, однако, не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе .
3. Световая (светозависимая) стадия
В ходе световой стадии фотосинтеза образуются высокоэнергетические продукты: АТФ, служащий в клетке источником энергии, и НАДФН, использующийся как восстановитель. В качестве побочного продукта выделяется кислород. В общем роль световых реакций фотосинтеза заключается в том, что в световую фазу синтезируются молекула АТФ и молекулы-переносчики протонов, то есть НАДФ Н2.
3.1. Фотохимическая суть процесса
Хлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на спектре его поглощения): первый связан с переходом на более высокий энергетический уровень электрона системы сопряжённых двойных связей, второй — с возбуждением неспаренных электронов азота и магния порфиринового ядра. При неизменном спине электрона формируются синглетные первое и второе возбуждённое состояние, при изменённом — триплетное первое и второе.
Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно и хлорофилл за 10-12 сек переходит с него на первое, с потерей 100 кДж/моль энергии только в виде теплоты. Из первого синглетного и триплетного состояний молекула может переходить в основное с выделением энергии в виде света (флуоресценция и фосфоресценция соответственно) или тепла, с переносом энергии на другую молекулу, либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносом электрона на другое соединение.
Первая возможность реализуется в светособирающих комплексах, вторая — в реакционных центрах, где переходящий в возбужденное состояние под воздействием кванта света хлорофилл становится донором электрона (восстановителем) и передаёт его на первичный акцептор. Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передаёт его вторичному. Кроме того, время жизни полученных соединений выше чем у возбуждённой молекулы хлорофилла. Происходит стабилизация энергии и разделения зарядов. Для дальнейшей стабилизации вторичный донор электронов восстанавливает положительно заряженный хлорофилл, первичным донором же является в случае оксигенного фотосинтеза вода.
Проблемой, с которой сталкиваются при этом проводящие оксигенный фотосинтез организмы, является различие окислительно-восстановительных потенциалов воды (для полуреакции h3O → O2 E0=+0,82 В) и НАДФ+ (E0=-0,32 В). Хлорофилл при этом должен иметь в основном состоянии потенциал больший +0,82 В чтобы окислять воду, но при этом иметь в возбуждённом состоянии потенциал меньший чем −0,32 В чтобы восстанавливать НАДФ+. Одна молекула хлорофилла не может отвечать обоим требованиям. Поэтому сформировалось две фотосистемы и для полного проведения процесса необходимо два кванта света и два хлорофилла разных типов.
3.2. Светособирающие комплексы
Хлорофилл выполняет две функции: поглощения и передачи энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.
Передача энергии идёт резонансным путём (механизм Фёрстера) и занимает для одной пары молекул 10−10—10−12 с, расстояние на которое осуществляется перенос составляет около 1 нм. Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каратиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины (с максимумом поглощения при 495—565 нм), за ними фикоцианины (550—615 нм) и аллофикоцианины (610—670 нм), последовательно передающие энергию на хлорофилл a (680—700 нм) реакционного центра.
3.3. Основные компоненты электронтранспортной цепи
3.3.1. Фотосистема II
Фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. Светособирающий комплекс II содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр фотосистемы II представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофилла a с максимумом поглощения при 680 нм (П680). На него в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула П680 становится сильным восстановителем (E0=-0,7 В).
П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС II и далее на пластохиноны, транспортируемые в восстановленной форме к b6f комплексу. Одна молекула пластохинона переносит 2 электрона и 2 протона, которые берутся из стромы.
Заполнение электронной вакансии в молекуле П680 происходит за счёт воды. В состав ФС II входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутритилакоидного пространства и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС II — это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве и 2 восстановленных пластохинонов в мембране.
3.3.2. b6f или b/f-комплекс
b6f комплекс является насосом, перекачивающим протоны из стромы во внутритилакоидное пространство и создающий градиент их концентрации за счёт выделяющейся в окислительно-восстановительных реакциях электронтранспортной цепи энергии. 2 пластохинона дают перекачку 4 протонов. В дальнейшем трансмембранный протонный градиент (pH стромы около 8, внутритилакоидного пространства — 5) используется для синтеза АТФ трансмембранным ферментом АТФ-синтазой.
3.3.3. Фотосистема I
Светособирающий комплекс I содержит примерно 200 молекул хлорофилла.
В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор — хлорофилл a, тот — вторичный (витамин K1 или филлохинон), после чего электрон передаётся на ферредоксин, который и восстанавливает НАДФ с помощью фермента ферредоксин-НАДФ-редуктазы.
Белок пластоцианин, восстановленный в b6f комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700.
3.4. Циклический и псевдоциклический транспорт электрона
Помимо полного нециклического пути электрона, описанного выше, обнаружены циклический и псевдоциклический.
Суть циклического пути заключается в том, что ферредоксин вместо НАДФ восстанавливает пластохинон, который переносит его назад на b6f комплекс. В результате образуется больший протонный градиент и больше АТФ, но не возникает НАДФН.
При псевдоциклическом пути ферредоксин восстанавливает кислород, который в дальнейшем превращается в воду и может быть использован в фотосистеме II. При этом также не образуется НАДФН.
4. Темновая стадия
В темновой стадии с участием АТФ и НАДФН происходит восстановление CO2 до глюкозы (C6h22O6). Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
4.1. С3-фотосинтез, цикл Кальвина
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий:
- Карбоксилирования
- Восстановления
- Регенерация акцептора CO2
На первой стадии к рибулозо-1,5-бифосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа. Этот белок составляет основную фракцию белков хлоропласта и предположительно наиболее распространённый фермент в природе. В результате образуется промежуточное неустойчивое соединение, распадающееся на две молекулы 3-фосфоглицериновой кислоты (ФГК).
Во второй стадии ФГК в два этапа восстанавливается. Сначала она фосфорилируется АТФ под действием фосфороглицерокиназы с образованием 1,3-дифосфоглицериновой кислоты (ДФГК), затем при воздействии триозофосфатдегидрогеназы и НАДФН ацил-фосфатная группа ДФГК дефосфорилируется и восстанавливается до альдегидной и образуется глицеральдегид-3-фосфат - фосфорилированный углевод (ФГА).
В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 5-углеродных рибулозо-1,5-бифосфата, для чего необходимы 3АТФ.
Наконец, две ФГА необходимы для синтеза глюкозы. Для образования одной её молекулы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН и 18 АТФ.
4.2. С4-фотосинтез
При низкой концентрации растворённого в строме CO2 рибулозобифосфаткарбоксилаза катализирует реакцию окисления рибулозо-1,5-бифосфата и его распад на 3-фосфоглицериновую кислоту и фосфогликолевую кислоту, которая вынужденно используется в процессе фотодыхания.
Для увеличения концентрации CO2 растения С4 типа изменили анатомию листа. Цикл Кальвина у них локализуется в клетках обкладки проводящего пучка, в клетках мезофилла же под действием ФЕП-карбоксилазы фосфоенолпируват карбоксилируется с образованием щавелеуксусной кислоты, которая превращается в малат или аспартат и транспортируется в клетки обкладки, где декарбоксилируется с образованием пирувата, возвращаемого в клетки мезофилла.
С4 фотосинтез практические не сопровождается потерями рибулозо-1,5-бифосфата из цикла Кальвина, поэтому более эффективен. Однако он требует не 18, а 30 АТФ на синтез 1 молекулы глюкозы. Это оправдывает себя в тропиках, где жаркий климат требует держать устьица закрытыми, что препятствует поступлению CO2 в лист, а также при рудеральной жизненной стратегии.
4.3. САМ фотосинтез
При CAM (англ. Crassulaceae acid metabolism — кислотный метаболизм толстянковых) фотосинтезе происходит разделение ассимиляции CO2 и цикла Кальвина не в пространстве как у С4, а во времени. Ночью в вакуолях клеток по аналогичному вышеописанному механизму при открытых устьицах накапливается малат, днём при закрытых устьицах идёт цикл Кальвина. Этот механизм позволяет максимально экономить воду, однако уступает в эффективности и С4, и С3. Он оправдан при стресстолерантной жизненной стратегии.
5. Значение фотосинтеза
Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия, получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф), также является запасённой в процессе фотосинтеза.
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь свободный кислород атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу.
6. Изучение фотосинтеза
Первые опыты по фотосинтезу были проведены Джозефом Пристли в 1770—1780-х годах, когда он обратил внимание на «порчу» воздуха в герметичном сосуде горящей свечой (воздух переставал быть способен поддерживать горение, помещённые в него животные задыхались) и «исправление» его растениями. Пристли сделал вывод что растения выделяют кислород, который необходим для дыхания и горения, однако не заметил что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз.
Позже было установлено что помимо выделения кислорода растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 Роберт Майер на основании закона сохранения энергии постулировал что растения преобразуют энергию солнечного света в энергию химических связей. В 1877 В. Пфеффер назвал этот процесс фотосинтезом.
Хлорофиллы были впервые выделены в 1818 П. Ж. Пельтье и Ж. Кавенту. Разделить пигменты и изучить их по отдельности удалось М. С. Цвету с помощью созданного им метода хроматографии. Спектры поглощения хлорофилла были изучены К. А. Тимирязевым, он же, развивая положения Майера, показал что именно поглощенные позволяют повысить энергию системы, создав вместо слабых связей С-О и О-Н высокоэнергетические С-С (до этого считалось что в фотосинтезе используются жёлтые лучи, не поглощаемые пигментами листа). Сделано это было благодаря созданному им методу учёта фотосинтеза по поглощённому CO2, в ходе экспериментов по освещению растения светом разных длин волн (разного цвета) оказалось что интенсивность фотосинтеза совпадает со спектром поглощения хлорофилла.
Окислительно-восстановительную сущность фотосинтеза (как оксигенного, так и аноксигенного) постулировал Корнелис ван Ниль. Это означало что кислород в фотосинтезе образуется полностью из воды, что экспериментально подтвердил в 1941 А. П. Виноградов в опытах с изотопной меткой. В 1937 г. Роберт Хилл установил что процесс окисления воды (и выделения кислорода), а также ассимиляции CO2 можно разобщить. В 1954—1958 Д. Арнон установил механизм световых стадий фотосинтеза, а сущность процесса ассимиляции CO2 была раскрыта Мельвином Кальвином с использованием изотопов углерода в конце 1940-х, за эту работу в 1961 ему была присуждена Нобелевская премия.
В 1955 году был выделен и очищен фермент рибулозобисфосфат-карбоксилаза/оксигеназа. С4 фотосинтез был описан Ю. С. Карпиловым в 1960 и М. Д. Хэтчем и К. Р. Слэком в 1966.
7. Прочие факты
- Морской слизень Elysia chlorotica ассимилирует хлоропласты водоросли Vaucheria litorea в клетки пищеварительного тракта. Хлоропласты способны фотосинтезировать в организме слизня в течение нескольких месяцев, что позволяет слизню жить за счет глюкозы, полученной в результате фотосинтеза. Геном слизня кодирует некоторые белки, необходимые хлоропластам для фотосинтеза.[1]
- Предполагается, что в естественной среде фотосинтезирующие бактерии могут использовать не только свет Солнца, но и другие источники света, а потому могут находиться в местах, не подвергающихся облучению звезды[2] В 2005 году Томас Битти из университета Британской Колумбии и Роберт Блейкеншип из университета Аризоны в глубоководных пробах, взятых в окрестностях глубоководного термального источника у побережья Коста Рики обнаружили серобактерию GSB1, сходную с серобактериями родов Chlorobium и Prosthecochloris, содержащую бактериохлорофилл. Они предположили, что вероятность контаминации образца невелика и, следовательно, GSB1 использует для фотосинтеза не солнечный свет (который не проникает сквозь 2,4-километровую толщу моря), а тусклый длинноволновый (~750 нм) свет, испускаемый гидротермальными источниками[2].
- По состоянию на конец 70-х годов XX века, мощность солнечной энергии, перерабатываемой земной растительностью при фотосинтезе, всего лишь на порядок превосходила мощность всех электростанций в мире.[3]
Литература
- Холл Д., Рао К. Фотосинтез: Пер. с англ. — М.: Мир, 1983.
- Физиология растений / под ред. проф. Ермакова И. П. — М.: Академия, 2007
- Молекулярная биология клетки / Альбертис Б., Брей Д. и др. В 3 тт. — М.: Мир, 1994
- Рубин А. Б. Биофизика. В 2 тт. — М.: Изд. Московского университета и Наука, 2004.
- Чернавская Н. М., Чернавский Д. С. Туннельный транспорт электронов в фотосинтезе. М., 1977.
- В. Любименко. Влияние света на усвоение органических веществ зелёными растениями // Известия Императорской Академии наук. VI серия. — 1907. — № 12. — С. 395—426, с 6 табл.
Примечания
- Rumpho ME, Worful JM, Lee J, et al. (November 2008). «Horizontal gene transfer of the algal nuclear gene psbO to the photosynthetic sea slug Elysia chlorotica - www.pnas.org/content/105/46/17867.abstract». Proc. Natl. Acad. Sci. U.S.A. 105 (46): 17867–17871. DOI:10.1073/pnas.0804968105 - dx.doi.org/10.1073/pnas.0804968105. PMID 19004808 - www.ncbi.nlm.nih.gov/pubmed/19004808?dopt=Abstract. Проверено 2008-11-24.
- ↑ 12Beatty, J. Thomas; Jörg Overmann, Michael T. Lince, Ann K. Manske, Andrew S. Lang, Robert E. Blankenship, Cindy L. Van Dover, Tracey A. Martinson, F. Gerald Plumley (2005). «An obligately photosynthetic bacterial anaerobe from a deep-sea hydrothermal vent - www.pnas.org/content/102/26/9306.abstract». Proceedings of the National Academy of Sciences of the United States of America 102 (26): 9306 -9310. DOI:10.1073/pnas.0503674102 - dx.doi.org/10.1073/pnas.0503674102. Проверено 2011-02-10.
- Углекислый газ в атмосфере земли - ufn.ru/ru/articles/1978/11/f/, Б.М. Смирнов, Объединенный институт высоких температур РАН, 126 (11) (1978), Москва
wreferat.baza-referat.ru
Фотосинтез и первичная биологическая продуктивность
Фотосинтез (от греч. φωτο- — свет и σύνθεσις — синтез, совмещение, помещение вместе) — процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл и бактериородопсин у бактерий). В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества.
 Рис.1. Фазы фотосинтеза
Рис.1. Фазы фотосинтеза
Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф) также является запасённой в процессе фотосинтеза.
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь свободный кислород атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу.
Cветовая фаза фотосинтеза
Осуществляется в хлоропластах, в которых на мембранах располагаются молекулы хлорофилла. Хлорофилл поглощает энергию солнечного света, которая затем используется при синтезе молекул АТФ из АДФ и фосфорной кислоты, а также способствуют расщеплению молекул воды: 2h30 = 4H+ + 4e- + O2. Кислород, образующийся при расщеплении выделяется в окружающую среду в свободной форме. Под влиянием энергии солнечного света молекула хлорофилла возбуждается, в результате чего один из её электронов переходит на более высокий энергетический уровень. Этот электрон, проходя по цепи переносчиков (белков мембраны хлоропласта), отдаёт избыточную энергию на окислительно-восстановительные реакции (синтез молекул АТФ). Молекулы хлорофилла, потерявшие электроны, присоединяют электроны, образующиеся при расщеплении молекулы воды. Под действием света электрон в реакционном центре переходит в возбуждённое состояние «перескакивая» на высокий энергетический уровень молекулы хлорофилла. Часть электронов, захваченных ферментами способствует образованию АТФ путём присоединения остатка фосфорной кислоты (Ф) и АДФ. Другая часть электронов принимает участие в разложении воды на молекулярный кислород, ионы водорода и электроны. Образовавшийся водород с помощью электронов присоединяется к веществу, способному транспортировать водород в пределах хлоропласта. Темновая фаза фотосинтеза
Для темновой фазы фотосинтеза обязательным компонентом является углекислый газ – СО2. Поэтому растение должно постоянно его поглощать из атмосферы. Для этой цели на поверхности листа имеются специальные структуры – устьица. Когда они открываются, СО2 поступает именно внутрь листа, растворяется в воде и вступает в реакцию световой фазы фотосинтеза. В ходе световой фазы у большинства растений СО2 связывается с пятиуглеродным органическим соединением (которое представляет собой цепочку из пяти молекул углерода), в результате чего образуются две молекулы трехуглеродного соединения (3-фосфоглицериновая кислота). Т.к. первичным результатом являются именно эти трехуглеродные соединения, растения с таким типом фотосинтеза получили название С3-растений. Дальнейший синтез, происходящий в хлоропластах, довольно сложен. В конечном итоге образуется шестиуглеродное соединение, из которого потом могут синтезироваться глюкоза, сахароза или крахмал. Именно в виде этих органических веществ растение накапливает энергию. Только небольшая их часть остается в листе и используется для его нужд. Остальные же углеводы путешествуют по всему растению и поступают именно туда, где больше всего нужна энергия, например, в точки роста.
Биологическая продуктивность, образование биомассы организмами, выражаемое потоками органического вещества и его потенциальной химической энергии на единицу площади за единицу времени. Понятие биологическая продуктивность применимо к растительности, сообществам (фитоценозам), к их отдельным ярусам, к отдельным популяциям растений и животных. Продуктивность всех популяций организмов на единицу площади характеризует биологическая продуктивность биогеоценозов и экосистем. Специфику процесса изучает физиология растений. Биологическая продуктивность иногда отождествляют с запасами биомассы на единицу площади, что может лишь отчасти характеризовать биологическая продуктивность. Различают первичную и вторичную биологическую продуктивность. Первичная биологическая продуктивность характеризуется образованием биомассы (первичной продукции) в процессе фотосинтеза зелёными растениями (автотрофами), которые образуют первый трофический уровень экосистемы и служат началом всех цепей питания. К первичным продуцентам относят и некоторые хемосинтезирующие бактерии. В процессе утилизации вещества и энергии первичной продукции образуется биомасса всех гетеротрофных организмов (бактерий, грибов и животных), называемых консументами. Продукция консументов характеризует вторичную биологическая продуктивность, к которой относят и массу хищных животных, питающихся растительноядными и другими хищниками.
 Рис.2. Первичная биологическая продуктивность
Рис.2. Первичная биологическая продуктивность
При исследованиях первичной биологической продуктивности наземных биогеоценозов определяют ряд показателей, которые затем используют в качестве отдельных статей баланса органические вещества на конкретных участках. Продукция, определяемая с учётом затрат вещества и энергии на процессы метаболизма самих организмов-продуцентов, называется первичной брутто-продукцией, или валовой продукцией (обычно обозначают GPP — от англ, gross primary production). Разность между первичной брутто-продукцией и затратами растений на дыхание (Ra) определяет первичную нетто-продукцию — NPP (от англ, net primary production). В лесном фитоценозе NPP включает в себя не только чистую продукцию прироста за учитываемый период (истинный прирост фитомассы) — NEP (net ecosystem production), но и продукцию, перешедшую за то же время в опад (листья, цветки, семена и др.) и отпад (отмершие деревья, сучья и др.), которые суммарно обозначаются L, а также часть продукции живых растений, пошедшую на корм животных-фитофагов (консумпцию) — Сa. Сумму этих показателей часто называют гетеротрофным дыханием (Rh), поскольку энергия в обоих этих потоках (Rh=L+Ca) освобождается главным образом с участием гетеротрофных организмов.
Для консументов, независимо от их трофической специализации, применяют иную схему. Отчуждаемая при консумпции фитофагами продукция растений в некотором количестве поедается животными, остальная (огрызки, объедки) поступает в опад. Съеденная пища частично ассимилируется организмами, частично экскре-тируется и поступает в детрит. За счёт продуктов ассимиляции происходит прирост биомассы, т. е. формируется продукция и поддерживаются процессы метаболизма. В продукцию включаются вещество или энергия прироста (привеса) животных за изучаемый период и прироста потомства. Эти величины, с учётом вещества и энергии элиминированных особей, характеризуют прирост биомассы животных. Биомасса животных-иммигрантов в продукцию не включается. При этом ассимилированная пища и прирост биомассы животных соответствуют общей (брутто) и чистой (нетто) продукции автотрофов.
Первичная биологическая продуктивность зависит от интенсивности фотосинтеза растениями и продолжительности его периода, фотосинтезирующей поверхности фитоценозов и древостоев, выражаемой индексом листовой поверхности и характером расположения листвы в толще полога, а также от кол-ва поступающей фотосинтетической радиации, условий увлажнения и минерального питания. Кульминация первичной продукции насаждений, выражаемая чистой продукцией (NPP) или истинным приростом фитомассы (NEP), приходится на возраст 20—40 лет. Однако в искусств, насаждениях она наступает раньше, чем в естественных, хотя с возрастом различия Б. п. лесных культур и естественных древостоев сглаживаются.
biofile.ru
Световая фаза фотосинтеза — Физиология растений
Фотосинтез имеет две фазы световую и темновую. В световой фазе молекулы пигментов поглощают фотоны, передают поглощенную энергию молекулам хлорофилла а, происходит трансформация энергии света в химическую энергию АТФ и восстановленного НАДФН, выделяется кислород в результате фоторазложения воды. Все эти процессы происходят на мембранах хлоропластов. При темновой фазе фотосинтеза в строме хлоропластов восстанавливается поглощенный СО2 с образованием углеводов и других органических соединений.
5.2.1. Поглощение света и возбуждение хлорофилла
Электрон представляет собой заряженную частицу, которая движется по орбите вокруг ядра атома. Электрон обладает вектором магнитного момента спином, который отражает направление вращения электрона вокруг своей оси. В стабильном состоянии валентные электроны занимают самые низкие энергетические уровни и распределены по ним согласно принципу Паули: два электрона с антипараллельными спинами на каждой орбитали. Такое состояние молекулы называют основным синглетным состоянием S0. При поглощении молекулой пигмента фотона электрон переходит на более высокую орбиталь за счет энергии фотона. Если у возбужденного электрона сохраняется направление спина, то считают, что молекула находится в синглетном возбужденном состоянии S*. Если возбужденный электрон меняет спин, то такое возбужденное состояние называется триплетным Т*.
Поглощение молекулой хлорофилла кванта красного света приводит к синглетному возбужденному состоянию S*1. При поглощении кванта синего цвета с более высоким уровнем энергии электрон переходит на более высокую орбиталь S*2. Время жизни электрона на S*2 составляет не более 10-12 секунды. За такое короткое время энергия электронного возбуждения не может быть использована и электрон возвращается на S*1, потеряв немного энергии в виде тепла. Время жизни электрона в состоянии S*1 равно 10-9. Если молекула пигмента не участвует в фотохимической реакции, то электрон возвращается в стабильное состояние. При этом часть энергии теряется в виде тепла и излучается квант света с большей по сравнению с поглощенным фотоном длиной волны явление флуоресценции.
Время жизни электрона в состоянии Т* равно 10-4-10-2 секунды. Из триплетного возбужденного состояния молекула может вернуться в стабильное состояние, излучив кроме тепла еще более длинноволновый, чем при флуоресценции, квант света. Это слабое свечение называют фосфоресценцией. Если молекула в возбужденном состоянии участвует в фотохимических реакциях с передачей электрона акцептору, то флуоресценции и фосфоресценции не происходит.
Для более полного использования падающего на листья света энергия фотонов улавливается 200-400 молекулами пигментов светособирающего (антенного) комплекса и передается к одной молекуле хлорофилла а, являющейся реакционным центром, которая и участвует в фотохимических реакциях. Передача энергии молекулами пигментов происходит по принципу индуктивного резонанса. Молекула пигмента, поглотившая фотон, создает переменное электрическое поле, которое усиливает колебания электрона в соседней молекуле. Это происходит из-за перекрытия частот колебаний и очень маленького расстояния между молекулами пигментов. Тем более, что при освещении листьев тилакоиды и граны сдвигаются и уплотняются, поэтому хлоропласты уменьшаются в объеме. Передача энергии молекулами пигментов идет с большой эффективностью. Так, от хлорофилла b к хлорофиллу а передается примерно 90 %, а от каротиноидов к хлорофиллу а 40 % поглощенной энергии.
Совокупность молекул светособирающего комплекса и реакционного центра составляет фотосистему. Предположение о существовании в хлоропластах двух фотосистем высказал Р. Эмерсон в 1957 г, изучая влияние света на квантовый выход фотосинтеза у водоросли хлореллы. Квантовый выход фотосинтеза это количество выделившегося кислорода или связанного углекислого газа на 1 квант поглощенной энергии. Он установил, что при одновременном освещении хлореллы коротковолновым (650 нм) и длинноволновым (700 нм) красным светом эффект выше, чем суммарный квантовый выход при воздействии красным светом этих длин в отдельности. Это явление получило название эффекта усиления Эмерсона. Позже предположение Р. Эмерсона о наличии двух фотосистем получило экспериментальное подтверждение.
5.2.2. Нециклический и циклический транспорт электронов
В фотосистеме II реакционный центр, состоящий из димера хлорофилла а с максимумом поглощения при 680 нм (П680), последовательно поглотив энергию, равную 2 квантам коротковолнового красного света, и перейдя в синглетное возбужденное состояние, передает 2 электрона феофитину. От феофитина электроны последовательно передаются на пластохиноны QA и QB, расположенные на другой стороне мембраны тилакоида, затем на две липидорастворимые молекулы пластохинона PQ. Он является переносчиком протонов и электронов, выполняющего в мембране роль челнока. Принимая 2 электрона и захватывая из стромы 2 Н+, 2 молекулы PQh3 диффундируют через липидную фазу мембраны. Электроны от 2 PQh3 поступают в цепь:
железосерный белок FeSr цитохром f Cu-содержащий белок пластоцианин П700,
а протоны попадают в полость тилакоида.
Вакантные места в П+680 заполняются 2 электронами из содержащего Mn переносчика электронов, который восстанавливается с участием белкового комплекса S. Окисленный комплекс S связывает воду и восстанавливается за счет электронов воды. При фотоокислении воды комплексом S вторая пара протонов освобождается в полость тилакоида, а атом кислорода диффундирует из хлоропласта.
После возбуждения реакционного центра фотосистемы I - молекулы хлорофилла а с максимумом поглощения при 700 нм (П700), 2 квантами длинноволного красного света 2 электрона передаются мономерной форме хлорофилла а А1 и затем переносчикам электронoв железосерным белкам А2 и АВ, ферредоксину на наружной стороне мембраны тилакоида, НАДФ-редуктазе. Этот фермент восстанавливает НАДФ в НАДФН. На вакантные места в П+700 переходят электроны с пластоцианина и нециклическая цепь транспорта электронов замыкается.
В мембранах хлоропластов может функционировать циклический транспорт электронов, когда действует только фотосистема I. Возбужденные молекулы П700 передают электроны на А1, железосерные белки А2 и АВ, переносящие электроны на другую сторону мембраны, ферредоксин, пластохинон, цитохром b6, железосерный белок FeSr, цитохром f, пластоцианин и П700 (рис. 5.4). Энергия, освобождающаяся при транспорте электронов, используется для фосфорилирования АДФ.
Рис. 5.4. Локализация электрон- и протонтранспортных реакций в тилакоидной мембране хлоропласта (по В. В. Полевому).
Механизм фотофосфорилирования АДФ объясняет теория П. Митчелла. Согласно этой теории, трансмембранный перенос пластохинонами электронов и протонов в одну сторону чередуется с переносом цитохромной системой в обратную сторону электронов. Поэтому по одну сторону мембраны накапливается избыток протонов и возникает электрохимический мембранный потенциал. Его энергия используется для синтеза АТФ при разрядке мембраны в результате транспорта протонов через мембрану посредством Н+-АТФазы, которая действует как АТФсинтетаза. Этот фермент выглядит как грибовидная частица на поверхности мембраны тилакоидов. Его молекула состоит из двух частей: головки, выступающей с наружной стороны мембраны и содержащей активный центр фермента, и ножки, погруженной в мембрану и представляющей канал, через который передвигаются протоны.
ifreestore.net
Влияние внутренних и внешних факторов на фотосинтез — Физиология растений
Фотосинтез осуществляется в полуавтономных органеллах хлоропластах. Однако он в значительной мере контролируется процессами, происходящими в растении, и факторами внешней среды.
Отток ассимилятов. Накопление фотоассимилятов в хлоропластах и в околопластидном пространстве приводит к ингибированию ферментов, участвующих в фотосинтезе.
Содержание хлорофилла. С увеличением содержания в клетке хлорофилла увеличивается интенсивность фотосинтеза.
Возраст листа и растения. В ходе роста листа интенсивность фотосинтеза увеличивается. После окончания роста листа она постепенно снижается. У многих однолетних растений интенсивность фотосинтеза достигает максимума в фазу бутонизации и цветения, а затем снижается.
Свет. Имеется нижний порог освещенности, при котором растения начинают фотосинтезировать. Затем зависимость интенсивности фотосинтеза от освещенности имеет логарифмический характер с последующим выходом на плато. Угол наклона кривой зависимости интенсивности фотосинтеза от освещенности зависит от влияния других факторов. Так, у светолюбивых растений она выходит на плато при значительно более высокой освещенности, чем у теневыносливых растений. Уровень освещения, при котором поглощение СО2 в ходе фотосинтеза равно выделению СО2 в процессе дыхания, называется компенсационным пунктом.
Важен и спектральный состав света. При освещении красным светом образуются преимущественно углеводы, синим - амино- и органические кислоты.
Температура. При низкой освещенности фотосинтез идет с одинаковой скоростью при 15 и 25оС. Это объясняется тем, что при низкой освещенности интенсивность фотосинтеза зависит от скорости световых реакций. При высокой освещенности интенсивность фотосинтеза лимитируется скоростью темновых реакций и Q10 примерно равен 2. Для большинства растения С3-типа оптимальная температура 20-25оС, для растений С4-типа она равна 25-40оС. При температуре выше оптимальной интенсивность фотосинтеза снижается из-за инактивации хлоропластов и закрытия устьиц.
Содержание СО2 в воздухе. Повышение содержания СО2 с 0,03 % до 0,3 % вызывает увеличение интенсивности фотосинтеза. Дальнейшее возрастание концентрации СО2 до 1 % не сказывается на фотосинтезе, но более высокий уровень СО2 в воздухе приводит к депрессии фотосинтеза. Высокие концентрации СО2 особенно неблагоприятны при высокой освещенности, так как происходит ингибирование темновых реакций. Влияние содержания углекислого газа на фотосинтез зависит от вида растения.
Снабжение водой. При большом водном дефиците интенсивность фотосинтеза снижается из-за закрытия устьиц, что уменьшает поступление СО2 в листья, снижает транспирацию и приводит к повышению температуры листа. Кроме того, обезвоживание изменяет конформацию и, следовательно, активность ферментов.
Содержание кислорода в воздухе, в среднем, равно 21 %. Повышение концентрации или отсутствие кислорода для фотосинтеза неблагоприятны. Кислород снижает активность рибулозодифосфаткарбоксилазы.
Минеральное питание. Исключение любого элемента минерального питания отрицательно сказывается на фотосинтезе. Особенно важны такие элементы как фосфор, магний, железо, марганец, медь, калий и азот. На всех этапах фотосинтеза участвуют фосфорилированные соединения. Калий активирует процессы фосфорилирования и участвует в открывании устьиц. Магний входит в состав хлорофиллов, активирует реакции карбоксилирования и восстановления НАДФ. Железо необходимо для синтеза хлорофиллов. Марганец участвует в фоторазложении воды. Медь входит в состав пластоцианина. Азот необходим для формирования хлоропластов и образования пигментов.
5.5. b
Выделяют пять основных аспектов планетарной роли фотосинтезирующих организмов.
Накопление органической массы. В процессе фотосинтеза наземные растения образуют до 170 млрд. тонн, а растения мирового океана до 70 млрд. тонн биомассы в год в пересчете на сухое вещество, используемой гетеротрофными организмами.
Обеспечение постоянства содержания СО2 в воздухе. Связывание СО2 в ходе фотосинтеза в значительной мере компенсирует его выделение в результате других процессов (дыхание, брожение, деятельность вулканов, производственная деятельность человечества).
Препятствие развитию парникового эффекта. Часть солнечного света отражается от поверхности Земли в виде тепловых инфракрасных лучей. СО2 поглощает инфракрасное излучение и тем самым сохраняет тепло на Земле. Повышение содержания СО2 в атмосфере может способствовать увеличению температуры, то есть создавать парниковый эффект. Это приведет к затоплению прибрежных зон из-за поднятия уровня мирового океана в результате таяния ледников в горах и на полюсах. Однако высокое содержание СО2 в воздухе активирует фотосинтез и, следовательно, концентрация СО2 в воздухе опять уменьшится.
Накопление кислорода в атмосфере. Первоначально в атмосфере Земли кислорода было очень мало. Сейчас его содержание составляет 21 % по объему воздуха. В основном, этот кислород является продуктом фотосинтеза. Ежегодно растения и другие фотосинтезирующие организмы поставляют в атмосферу примерно 120 млрд. тонн кислорода.
Озоновый экран. Озон (О3) образуется в результате фотодиссоциации молекул кислорода под действием солнечной радиации на высоте около 25 км. Озон задерживает большую часть ультрафиолетовых лучей, губительных для всего живого.
ifreestore.net
Реферат на тему фотосинтез | Kursak.NET
Растения превращают солнечный свет в запасенную химическую энергию в два этапа: сначала они улавливают энергию солнечного света, а затем используют ее для связывания углерода с образованием органических молекул.
Зеленые растения — биологи называют их автотрофами — основа жизни на планете. С растений начинаются практически все пищевые цепи. Они превращают энергию, падающую на них в форме солнечного света, в энергию, запасенную в углеводах, из которых важнее всего шестиуглеродный сахар глюкоза. Этот процесс преобразования энергии называется фотосинтезом. Другие живые организмы получают доступ к этой энергии, поедая растения. Так создается пищевая цепь, поддерживающая планетарную экосистему.
Кроме того, воздух, которым мы дышим, благодаря фотосинтезу насыщается кислородом. Суммарное уравнение фотосинтеза выглядит так:
вода + углекислый газ + свет —> углеводы + кислород
Растения поглощают углекислый газ, образовавшийся при дыхании, и выделяют кислород — продукт жизнедеятельности растений. К тому же, фотосинтез играет важнейшую роль в круговороте углерода в природе.
История открытия фотосинтеза
В начале XVII в. фламандский врач Ван Гельмонт вырастил в кадке с землей дерево, которое он поливал только дождевой водой. Он заметил, что спустя пять лет, дерево выросло до больших размеров, хотя количество земли в кадке практически не уменьшилось. Ван Гельмонт, естественно, сделал вывод, что материал, из которого образовалось дерево произошел из воды, использованной для полива. В 1777 английский ботаник Стивен Хейлс опубликовал книгу, в которой сообщалось, что в качестве питательного вещества, необходимого для роста, растения используют главным образом воздух. В тот же период знаменитый английский химик Джозеф Пристли (он был одним из первооткрывателей кислорода) провел серию опытов по горению и дыханию и пришел к выводу о том, что зелёные растения способны совершать все те дыхательные процессы, которые были обнаружены в тканях животных. Пристли сжигал свечу в замкнутом объеме воздуха, и обнаруживал, что получавшийся при этом воздух уже не может поддерживать горение. Мышь, помещенная в такой сосуд, умирала. Однако веточка мяты продолжала жить в воздухе неделями. В заключение Пристли обнаружил, что в воздухе, восстановленном веточкой мяты, вновь стала гореть свеча, могла дышать мышь. Теперь мы знаем, что свеча, сгорая, потребляла кислород из замкнутого объема воздуха, но затем воздух снова насыщался кислородом благодаря фотосинтезу, происходившему в оставленной веточке мяты. Спустя несколько лет голландский врач Ингенхауз обнаружил, что растения окисляют кислород лишь на солнечном свету и что только их зеленые части обеспечивают выделение кислорода. В 1817 г. два французских химика, Пельтье и Каванту, выделили из листьев зеленое вещество и назвали его хлорофиллом. Следующей важной вехой в истории изучения фотосинтеза было сделанное в 1845 г. немецким физиком Робертом Майером утверждение о том, что зеленые растения преобразуют энергию, солнечного света в химическую энергию.
История изучения фотосинтеза
В течение тысячелетий люди считали, что питается растение исключительно благодаря корням, поглощая с их помощью все необходимые вещества из почвы. Проверить эту точку зрения взялся в начале девятнадцатого века голландский натуралист Ян Ван Гельмонт. Он взвесил землю в горшке и посадил туда побег ивы. В течение пяти лет он поливал деревце, а затем высушил землю и взвесил её и растение. Ива весила семьдесят пять килограмм, а вес земли изменился всего на несколько сот граммов. Вывод учёного был таков – растения получают питательные вещества, прежде всего, не из почвы, а из воды.
На два столетия в науке утвердилась теория водного питания растений. Листья, по этой теории, лишь помогали растению испарять излишнюю влагу.
К самому неожиданному, но правильному предположению о воздушном питании растений ученые пришли лишь к началу девятнадцатого века. Важную роль в понимании этого процесса сыграло открытие, совершенное английским химиком Джозефом Пристли в 1771 году. Он поставил опыт, в результате которого он сделал вывод: растения очищают воздух и делают его пригодным для дыхания. Позднее выяснилось: для того, чтобы растение очищало воздух, необходим свет.
Десять лет спустя, учёные поняли, что растение не просто превращает углекислый газ в кислород. Углекислый газ необходим растениям для жизни, он служит для них настоящей пищей (вместе с водой и минеральными солями).
Воздушное питание растений называется фотосинтезом. Кислород в процессе фотосинтеза выделяется в качестве необычного продукта.
Миллиарды лет назад на земле не было свободного кислорода. Весь кислород, которым дышат почти все живые существа нашей планеты, выделен растениями в процессе фотосинтеза. Фотосинтез сумел изменить весь облик нашей планеты!
Начиная с семидесятых годов прошлого столетия, крупные успехи в области фотосинтеза были получены в России. Работами русских учёных Пуриевича, Ивановского, Риктера, Иванова, Костычева были изучены многие стороны этого процесса .
Значение фотосинтеза не осознавалось до сравнительно недавнего времени. Аристотель и другие учёные Греции, наблюдая, что жизненные процессы животных зависят от потребления пищи, полагали, что растения добывают свою «пищу» из почвы.
Немногим более трехсот лет назад в одном из первых тщательно продуманных биологических экспериментов голландский врач Ян Ван Гельмонт представил доказательства того, что не одна почва кормит растение. Ван Гельмонт выращивал маленькое дерево ивы в глиняном горшке, добавляя в него только воду.
Через пять лет масса игл увеличилась на 74,4 кг, в то время, как масса почвы уменьшилась только на 57 гр.
В конце XVIII века английский ученый Джозеф Пристли сообщил, что он «случайно обнаружил метод исправления воздуха, который был испорчен горением свечей». 17 августа 1771 г. Пристли «… поместил живую веточку мяты в закрытый сосуд, в котором горела восковая свеча», а 21 числа того же месяца обнаружил, что «… другая свеча снова могла гореть в этом же сосуде». «Исправляющим началом, которым для этих целей пользуется природа, – полагал Пристли, – было растение». Он расширил свои наблюдения и скоро показал, что воздух, «исправляемый» растением, не был «совсем не подходящим для мыши».
Опыты Пристли впервые позволили объяснить, почему воздух на Земле остается «чистым» и может поддерживать жизнь, несмотря на горение бесчисленных огней и дыхание множества живых организмов. Он говорил: «Благодаря этим открытиям мы уверены, что растения произрастают не напрасно, а очищают и облагораживают нашу атмосферу».
Позднее голландский врач Ян Ингенхауз (1730-1799) подтвердил работу Пристли и показал, что воздух «исправляется» только на солнечном свету и только зелеными частями растения. В 1796 году Ингенхауз предположил, что углекислота разлагается при фотосинтезе на С и О2, а О2 выделяется в виде газа. В последствие было обнаружено, что соотношение атомов углерода, водорода и кислорода в сахарах и крахмале таково, что один атом углерода приходится на одну молекулу воды, на что и указывает слово «углеводы». Считалось общепринятым, что углеводы образуются из С и Н2О, а О2 выделяется из углекислоты. Это вполне разумная гипотеза была широко признана, но, как позднее выяснилось, она была совершенно неверной.
Исследователем, который опроверг эту общепринятую теорию, был Корнелиус ван Ниль из Стамфордского университета, когда он, будучи еще студентом – дипломником, исследовал метаболизм различных фотосинтезирующих бактерий. Одна группа таких бактерий, а именно пурпурные серные бактерии, восстанавливает С до углеводов, но не выделяет О2. Пурпурным серным бактериям для фотосинтеза необходим сероводород. В результате фотосинтеза внутри бактериальных клеток накапливаются частицы серы. Ван Ниль обнаружил, что для этих бактерий уравнение фотосинтеза может быть записано как:
свет
С О2 + 2Н2S (Ch3O) + Н2О + 2S
Этот факт не привлекал внимания исследователей до тех пор, пока ван Ниль не сделал смелого сообщения и не предложил следующего суммарного уравнения фотосинтеза:
свет
С О2 + 2Н2А (Ch3O) + Н2О + 2А
В этом уравнении Н2А представляет собой либо воду, либо другое окисляемое вещество, например, сероводород или свободный Н2. У зеленых растений и водорослей Н2А = Н2О. То есть ван Ниль предположил, что Н2О, а не углекислота, разлагается при фотосинтезе. Эта блестящая идея, выдвинутая в тридцатые годы, экспериментально была доказана позднее, когда исследователи, использую тяжелый изотоп О2(18О2), проследили путь кислорода от воды до газообразного состояния:
свет
С О2 + 2Н218О2 (Ch3O) + Н2О + 18О2
Таким образом, для водорослей или зеленых растений, у которых вода служит донором электронов, суммарное уравнение фотосинтеза записывается следующим образом:
свет
6СО2 + 12Н2О C6h22O6 + 6О2 + 6Н2О
Пространственная локализация фотосинтеза
Хлоропласты в клетках листа
Фотосинтез растений осуществляется в хлоропластах: обособленных двухмембранных органеллах клетки. Хлоропласты могут быть в клеткахплодов, стеблей, однако основным органом фотосинтеза, анатомически приспособленным к его ведению, является лист. В листе наиболее богата хлоропластами ткань палисадной паренхимы. У некоторых суккулентов с вырожденными листьями (например, кактусы) основная фотосинтетическая активность связана со стеблем.
Свет для фотосинтеза захватывается более полно благодаря плоской форме листа, обеспечивающей большое отношение поверхности к объёму. Вода доставляется из корня по развитой сети сосудов (жилок листа). Углекислый газ поступает отчасти посредством диффузии через кутикулу иэпидермис, однако большая его часть диффундирует в лист через устьица и по листу по межклеточному пространству. Растения, осуществляющие С4 и CAM фотосинтез, сформировали особые механизмы для активной ассимиляции углекислого газа.
Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые соединяясь друг с другом образуют тилакоиды, которые в свою очередь группируются в стопки, называемые граны. Внутритилакоидное пространство отделено и не сообщается с остальной стромой, предполагается также что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (70s типа), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их потомками свободных цианобактерий, вошедших в состав эукариотической клетки в процессе симбиогенеза.
Цианобактерии, таким образом, как бы сами являются хлоропластом и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды, однако, не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе.
Процессы, происходящие в листе
Лист осуществляет три важных процесса – фотосинтез, испарение воды и газообмен. В процессе фотосинтеза в листьях из воды и двуокиси углерода под действием солнечных лучей синтезируются органические вещества. Днем, в результате фотосинтеза и дыхания, растение выделяет кислород и двуокись углерода, а ночью – только двуокись углерода, образующуюся при дыхании.
Большинство растений способно синтезировать хлорофилл при слабом освещении. При прямом солнечном освещении хлорофилл синтезируется быстрее.
Необходимая для фотосинтеза световая энергия в известных пределах поглощается тем больше, чем меньше затемнен лист. Потому у растений в процессе эволюции выработалась способность поворачивать пластину листа к свету так, чтобы на нее падало больше солнечных лучей. Листья на растении располагаются так, чтобы не притеснять друг друга.
Тимирязев доказал, что источником энергии для фотосинтеза служат преимущественно красные лучи спектра. На это указывает спектр поглощения хлорофилла, где наиболее интенсивная полоса поглощения наблюдается в красной, и менее интенсивное – в сине-фиолетовой части.
В хлоропластах вместе с хлорофиллом имеются пигменты каротин и ксантофилл. Оба этих пигмента поглощают синие и, отчасти, зеленые лучи и пропускают красные и желтые. Некоторые ученые приписываю каротину и ксантофиллу роль экранов, защищающих хлорофилл от разрушительного действия синих лучей.
Процесс фотосинтеза слагается из целого ряда последовательных реакций, часть которых протекает с поглощением световой энергии, а часть – в темноте. Устойчивыми окончательными продуктами фотосинтеза являются углеводы (сахара, а затем крахмал), органические кислоты, аминокислоты, белки.
Фотосинтез при различных условиях протекает с разной интенсивностью.
Интенсивность фотосинтеза также зависит от фазы развития растения. Максимальная интенсивность фотосинтеза наблюдается в фазе цветения.
Обычное содержание углекислоты в воздухе составляет 0,03% по объему. Уменьшение содержания углекислоты в воздухе снижает интенсивность фотосинтеза. Повышение содержания углекислоты до 0,5% увеличивает интенсивность фотосинтеза почти пропорционально. Однако при дальнейшем повышении содержания углекислоты, интенсивность фотосинтеза не возрастает, а при 1% – растение страдает.
Растения испаряют или трансперируют очень большое количество воды. Испарение воды является одной из причин восходящего тока. Вследствие испарения воды растением в нем накапливаются минеральные вещества, и происходит полезное для растения понижение температуры во время солнечного нагрева. Иногда трансперация снижает температуру растения на 6о.
Растение регулирует процесс испарения воды посредством работы устьиц. Отложение кутикулы или воскового налета на эпидерме, образование его волосков и другие приспособления направлены к сокращению нерегулируемой трансперации.
Процесс фотосинтеза и постоянное протекающее дыхание живых клеток листа требуют газообмена между внутренними тканями листа и атмосферой. В процессе фотосинтеза из атмосферы поглощается ассимилируемый углекислый газ и возвращается в атмосферу кислородом.
Применение изотопного метода анализа показало, что кислород, возвращаемый в атмосферу (16О) принадлежит воде, а не углекислому газу воздуха, в котором преобладает другой его изотоп – 15О. При дыхании живых клеток (окисление свободным кислородом органических веществ внутри клетки до углекислого газа и воды) необходимо поступление из атмосферы кислорода и возвращение углекислоты. Этот газообмен также в основном осуществляется через устьичный аппарат.
Световые и темновые реакции
Еще в 1905 г. английский физиолог растений Ф. Ф. Блекмэн, интерпретируя форму кривой светового насыщения фотосинтеза, высказал предположение, что фотосинтез представляет собой двухстадийный процесс, включающий фотохимическую, т.е. светочувствительную реакцию и нефотохимическую, т. е. темновую, реакцию. Темновая реакция, будучи ферментативной, протекает медленнее, чем световая реакция, и поэтому при высоких интенсивностях света скорость фотосинтеза полностью определяется скоростью темновой реакции. Световая реакция либо вообще не зависит от температуры, либо зависимость эта выражена очень слабо, тогда темновая реакция, как и все ферментативные процессы, зависит от температуры в довольно значительно и степени. Следует ясно представлять себе, что реакция, называемая темновой, может протекать как в темноте, так и на свету. Световую и темновую реакции можно разделить, используя вспышки света, длящиеся краткие доли секунды. Вспышки света длительностью меньше одной миллисекунды (10-3 с) можно получить либо с помощью механического приспособления, поставив на пути пучка постоянного света вращающийся диск со щелью, либо электрически, заряжая конденсатор и разряжая его через вакуумную или газоразрядную лампу. В качестве источников света пользуются также рубиновыми лазерами с длиной волны излучения 694 нм. В 1932 г. Эмерсон и Арнольд освещали суспензию клеток вспышками света от газоразрядной лампы с длительностью около 10-3с. Они измеряли скорость выделения кислорода в зависимости от энергии вспышек, длительности темнового промежутка между вспышками и температуры суспензии клеток. При увеличении интенсивности вспышек насыщение фотосинтеза в нормальных клетках наступало, когда выделялась одна молекула 02 на 2500 молекул хлорофилла. Эмерсон и Арнольд сделали вывод, что максимальный выход фотосинтеза определяется не числом молекул хлорофилла, поглощающих свет, а числом молекул фермента, катализирующего темновую реакцию. Они также обнаружили, что при увеличении темновых интервалов между последовательными вспышками за пределы 0,06 с выход кислорода в расчете на одну вспышку уже не зависел от длительности темнового интервала, тогда как при более коротких промежутках он возрастал с увеличением длительности темнового интервала (от 0 до 0,06 с). Таким образом, темновая реакция, которая определяет уровень насыщения фотосинтеза, завершается примерно за 0,06 с. На основе этих данных было рассчитано, что среднее время, характеризующее скорость реакции, составило около 0,02 с при 25°С.
Лимитирующие факторы
Интенсивность, или скорость процесса фотосинтеза в растении зависит от ряда внутренних и внешних факторов. Из внутренних факторов наиболее важное значение имеют структура листа и содержание в нем хлорофилла, скорость накопления продуктов фотосинтеза в хлоропластах, влияние ферментов, а также наличие малых концентраций необходимых неорганических веществ. Внешние параметры – это количество и качество света, попадающего на листья, температура окружающей среды, концентрация углекислоты и кислорода в атмосфере вблизи растения.
Скорость фотосинтеза возрастает линейно, или прямо пропорционально увеличению интенсивности света. По мере дальнейшего увеличения интенсивности света нарастание фотосинтеза становится все менее и менее выраженным, и, наконец, прекращается, когда освещенность достигает определенного уровня 10000 люкс. Дальнейшее увеличение интенсивности света уже не влияет на скорость фотосинтеза. Область стабильной скорости фотосинтеза называется областью светонасыщения. Если нужно увеличить скорость фотосинтеза в этой области, следует изменять не интенсивность света, а какие-либо другие факторы. Интенсивность солнечного света, попадающего в ясный летний день на поверхность земли, во многих местах нашей планеты составляет примерно 100000 люкс. Следовательно, растениям, за исключением тех, которые растут в густых лесах и в тени, падающего солнечного света бывает достаточно для насыщения их фотосинтетической активности (энергия квантов, соответствующих крайним участкам видимого диапазона – фиолетового и красного, различается всего лишь в два раза, и все фотоны этого диапазона в принципе способны осуществить запуск фотосинтеза).
В случае низких интенсивностей света скорость фотосинтеза при 15 и 25°С одинакова. Реакции, протекающие при таких интенсивностях света, которые соответствуют области лимитирования света, подобно истинным фотохимическим реакциям, не чувствительны к температурам. Однако при более высоких интенсивностях скорость фотосинтеза при 25°С гораздо выше, чем при 15°С. Следовательно, в области светового насыщения уровень фотосинтеза зависит не только от поглощения фотонов, но и от других факторов. Большинство растений в умеренном климате хорошо функционируют в интервале температур от 10 до 35°С, наиболее благоприятные условия – это температура около 25°С.
В области лимитирования светом скорость фотосинтеза не изменяется при уменьшении концентрации СО2 . Отсюда можно сделать вывод, что С02 участвует непосредственно в фотохимической реакции. В то же время при более высоких интенсивностях освещения, лежащих за пределами области лимитирования, фотосинтез существенно возрастает при увеличении концентрации СО2. У некоторых зерновых культур фотосинтез линейно возрастал при увеличении концентрации СО2 до 0,5%. (эти измерения проводили в кратковременных опытах, поскольку длительное воздействие высоких концентраций СО2 повреждает листы). Высоких значений скорость фотосинтеза достигает при содержании С02 около 0,1%. Средняя концентрация углекислоты в атмосфере составляет от 0,03%. Поэтому в обычных условиях растениям не хватает СО2 для того, чтобы с максимальной эффективностью использовать попадающий на них солнечный свет. Если помещенное в замкнутый объем растение освещать светом насыщающей интенсивности, то концентрация СО2 в объеме воздуха будет постепенно уменьшаться и достигнет постоянного уровня, известного под названием «С02 компенсационного пункта». В этой точке появление СО2 при фотосинтезе уравновешивается выделением О2 в результате дыхания (темнового и светового). У растений разных видов положения компенсационных пунктов различны.
Бактериальный фотосинтез
Наряду с фотосинтезом высших растений и водорослей, сопровождаемым выделением O2, в природе осуществляется бактериальный фотосинтез, в котором окисляемым субстратом является не вода, а другие соединения, обладающие более выраженными восстановительными свойствами, например h3S, SO2. Кислород при бактериальном фотосинтезе не выделяется, например:
Фотосинтезирующие бактерии способны использовать не только видимое, но и ближнее ИК излучение (до 1000 нм) в соответствии со спектрами поглощения преобладающих в них пигментов – бактериохлорофиллов. Бактериальный фотосинтез не имеет существенного значения в глобальном запасании солнечной энергии, но важен для понимания общих механизмов фотосинтеза Кроме того, локально бескислородный фотосинтез может вносить существенный вклад в суммарную продуктивность планктона. Так, в Черном море количество хлорофилла и бактериохлорофилла в столбеводы в ряде мест приблизительно одинаково.
Фотосинтез галобактерий
Единственный известный в природе нехлорофилльный способ запасания энергии света осуществляют бактерииHalobacterium halobium. Ha ярком свету при пониженной концентрации O2 они образуют в своих мембранах пурпурный белок бактериородопсин. В результате индуцированной светом цис-транс-изомеризации ретиналя (хромофора этого пигмента) происходит поглощение H+ и синтез АТФ. Последний используется для частичного обеспечения энергетических потребностей клетки.
Современные представления о фотосинтезе
В настоящее время известно, что фотосинтез проходит две стадии, но только одна из них – на свету. Доказательства двухстадийности процесса впервые были получены в 1905 году английским физиологом растений Ф.Ф. Блэклином, который исследовал влияние освещенности и температуры на объем фотосинтеза.
На основании экспериментов, Блэклин сделал следующие выводы.
1. Имеется одна группа светозависимых реакций, которые не зависят от температуры. Объем этих реакций в диапазоне низких освещенностей мог возрастать с увеличением освещенности, но не с увеличением температуры.
2. Имеется вторая группа реакций, зависимых от температуры, а не от света. Оказалось, что обе группы реакций необходимы для осуществления фотосинтеза. Увеличение объема только одной группы реакций увеличивает объем всего процесса, но только до того момента, пока вторая группа реакций не начнет удерживать первую. После этого необходимо ускорить вторую группу реакций, чтобы первые могли проходить без ограничений.
Таким образом, было показано, что обе стадии светозависимы: «световая и темновая». Важно помнить, что темновые реакции нормально проходят на свету и нуждаются в продуктах световой стадии. Выражение «темновые реакции» просто означает, что свет как таковой в них не участвует.
Объем темновых реакций возрастает с увеличением температуры, но только до 30о, а затем начинает падать. На основании этого факта предположили, что темновые реакции катализируются ферментами, поскольку обмен ферментативных реакций, таким образом, зависит от температуры. В последствие оказалось, что данный вывод был сделан неправильно.
На первой стадии фотосинтеза (световые реакции) энергия света используется для образования АТР (молекула аденозин-трифосфата) и высокоэнергетических переносчиков электронов. На второй стадии фотосинтеза (темновые реакции) энергетические продукты, образовавшиеся в световых реакциях, используются для восстановления СО2 до простого сахара (глюкозы).
Процесс фотосинтеза все больше и больше привлекает к себе внимание ученых. Наука близка к разрешению важнейшего вопроса – искусственного создания при помощи световой энергии ценных органических веществ из широко распространенных неорганических веществ. Проблема фотосинтеза усиленно разрабатывается ботаниками, химиками, физиками и другими специалистами.
В последнее время уже удалось искусственно получить синтез формальдегида и сахаристых веществ из водных растворов карбонатной кислоты; при этом роль поглотителя световой энергии играли вместо хлорофилла карбонаты кобальта и никеля. Недавно синтезирована молекула хлорофилла.
Успехи науки в области синтеза органических веществ наносят сокрушительный удар по идеалистическому учению – витализму, который доказывал, что для образования органических веществ из неорганических необходима особая «жизненная сила» и что человек не сможет синтезировать сложные органические вещества.
Фотосинтез в растениях осуществляется в хлоропластах. Он включает преобразования энергии (световой процесс), превращение вещества (темновой процесс). Световой процесс происходит в гилакоидах, темновой – в строме хлоропластов. Обобщенное циркулирование фотосинтеза выглядит следующим образом:
свет
6СО2 + 12Н2О C6h22O6 + 6Н2О + 6О2
Два процесса фотосинтеза выражаются отдельными уравнениями
свет
12Н2О 12h3 + 6О2 + энергия АТР
(световой процесс)
свет
12h3 + 6О2 + энергия АТР С6Н12О6 + Н2О
(темновой процесс)
Значение фотосинтеза в природе
Фотосинтез – единственный процесс в биосфере, ведущий к увеличению ее свободной энергии за счет внешнего источника. Запасенная в продуктах фотосинтеза энергия – основной источник энергии для человечества.
Ежегодно в результате фотосинтеза на Земле образуется 150 млрд. тонн органического вещества и выделяется около 200 млн. тонн свободного кислорода.
Круговорот кислорода, углерода и других элементов, вовлекаемых в фотосинтез, поддерживает современный состав атмосферы, необходимый для жизни на Земле. Фотосинтез препятствует увеличению концентрации СО2, предотвращая перегрев Земли вследствие так называемого «парникового эффекта».
Поскольку зеленые растения представляют собой непосредственную или опосредованную базу питания всех других гетеротрофных организмов, фотосинтез удовлетворяет потребность в пище всего живого на нашей планете. Он – важнейшая основа сельского и лесного хозяйства. Хотя возможности воздействия на него еще не велики, но все же и они, в какой то мере используются. При повышении концентрации углекислого газа в воздухе до 0,1% (против 0,3% в естественной атмосфере) удалось, например, повысить урожайность огурцов и томатов втрое.
Квадратный метр поверхности листьев в течение одного часа продуцирует около одного грамма сахара; это значит, что все растения, по приблизительной оценке, изымают из атмосферы от 100 до 200 млрд. тонн С в год. Около 60% этого количества поглощают леса, занимающие 30% непокрытой льдами поверхности суши, 32% – окультуренные земли, а оставшиеся 8% – растения степей и пустынных мест, а также городов и поселков.
Зеленое растение способно не только использовать углекислый газ и создавать сахар, но и превращать азотные соединения, и соединения серы в вещества, слагающие его тело. Через корневую систему растение получает растворенные в почвенной воде ионы нитратов и перерабатывает их в своих клетках в аминокислоты – основные компоненты всех белковых соединений. Компоненты жиров также возникают из соединений, образующихся в процессах обмена веществ и энергии. Из жирных кислот и глицерина возникают жиры и масла, которые служат для растения, главным образом, запасными веществами. В семенах приблизительно 80% всех растений, в качестве богатого энергией запасного вещества, содержатся жиры. Получение семян, жиров и масел играет важную роль в сельскохозяйственной и пищевой промышленности.
Прочие факты
- Морской слизень Elysia chlorotica ассимилирует хлоропласты водоросли Vaucheria litorea в клетки пищеварительного тракта. Хлоропласты способны фотосинтезировать в организме слизня в течение нескольких месяцев, что позволяет слизню жить за счет глюкозы, полученной в результате фотосинтеза. Геном слизня кодирует некоторые белки, необходимые хлоропластам для фотосинтеза.
- Предполагается, что в естественной среде фотосинтезирующие бактерии могут использовать не только свет Солнца, но и другие источники света, а потому могут находиться в местах, не подвергающихся облучению звезды. В 2005 году Томас Битти из университета Британской Колумбии и Роберт Блейкеншип из университета Аризоны в глубоководных пробах, взятых в окрестностях глубоководного термального источника у побережья Коста Рики обнаружили серобактерию GSB1, сходную с серобактериями родов Chlorobium иProsthecochloris, содержащую бактериохлорофилл. Они предположили, что вероятность контаминации образца невелика и, следовательно, GSB1 использует для фотосинтеза несолнечный свет (который не проникает сквозь 2,4-километровую толщу моря), а тусклый длинноволновый (~750 нм) свет, испускаемый гидротермальными источниками.
- По состоянию на конец 70-х годов XX века, мощность солнечной энергии, перерабатываемой земной растительностью при фотосинтезе, всего лишь на порядок превосходила мощность всех электростанций в мире.
Заключение
Во второй половине XIX столетия было установлено, что энергия солнечного света усваивается и трансформируется при помощи зеленого пигмента хлорофилла.
На основе проведенных опытов можно сказать что, зеленая окраска хлорофилла определяется наличием в нем атома металла вне зависимости от того, будет ли это магний, медь или цинк.
Современная наука подтвердила правильность взглядов К. А. Тимирязева относительно исключительной важности для фотосинтеза именно красных лучей солнечного спектра. Оказалось, что коэффициент использования красного света в ходе фотосинтеза выше, чем синих лучей, которые также поглощаются хлорофиллом. Красные лучи, по представлениям К. А. Тимирязева, играют основополагающую роль в процессе мироздания и созидания жизни.
Как известно растения поглащают углекислый газ, который присоединяется к пятиуглеродному веществу под названием рибулезодифосфат, где потом он в дальнешем участвует во многих других реакциях.
Изучение особенностей фотосинтеза у разных растений, безусловно, будет способствовать расширению возможностей человека в управлении их фотосинтетической деятельностью, продуктивностью и урожаем. В целом фотосинтез это один из основополагающих процессов жизни, на котором основана большая часть современной растительной фауны на поверхности земли.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
- Холл Д., Рао К. «Фотосинтез»: Пер. с англ. — М.: Мир, 1983.
- под ред. проф. Ермакова И. П. «Физиология растений» — М.: Академия, 2007.
- Альбертис Б., Брей Д. и др. «Молекулярная биология клетки» В 3 тт. — М.: Мир, 1994.
- Чернавская Н. М., Чернавский Д. С. «Туннельный транспорт электронов в фотосинтезе». – М., 1977.
- Медведев С. С. «Физиология растений» — СПб,: СПбГУ, 2004.
- Сергеев И. И. «История фотосинтеза». – М.: Наука, 1989.
- Пчелов А. М. «Природа и ее жизнь». – Л.: Жизнь, 1990.
kursak.net