Помогите решить тест по биологии (заранее спасибо). Процесс образования углеводов в клетках высших растений из
Углеводы образование в процессах фотосинтеза
Окисление органических веществ. В результате поглощения СО2 и дальнейших его преобразований в ходе фотосинтеза образуется молекула углевода, которая служит углеродным скелетом для построения всех органических соединений в клетке. Органические вещества, возникшие в процессе фотосинтеза, характеризуются высоким запасом внутренней энергии. Но энергия, аккумулированная в конечных продуктах фотосинтеза — углеводах, жирах, белках,— недоступна для непосредственного использования ее в химических реакциях. Перевод этой потенциальной энергии в активную форму осуществляется в процессе дыхания. Дыхание включает механизмы активации атомоп водорода органического субстрата, освобождения и мобилизации энергии в виде АТФ и генерации различных углеродных скелетов. В процессе дыхания углевод, жиры и белки в реакциях биологического окисления и постепенной перестройки органического скелета отдают спои атомы водорода с образованием восстановленных форм. Последние при окислении в дыхательной цепи освобождают энергию, которая аккумулируется в активной форме в сопряженных реакциях синтеза АТФ. Таким образом, фотосинтез и дыхание — это разли ные, но тесно связанные стороны общего энергообмена. [c.609] Таким образом, суммарный результат фотосинтеза состоит в связывании диоксида углерода, окислении воды до молекулярного кислорода и синтеза углеводов. Образование кислорода как побочного продукта фотосинтеза не является универсальным свойством фотосинтезирующих организмов. Например, у некоторых бактерий фотосинтеза процесс выражается схемой [c.162]Биосинтез начинается с фотосинтеза [1]. Вся жизнь на Земле зависит от способности некоторых организмов (зеленых растений, водорослей и фотосинтезирующих бактерий), содержащих характерные фотосинтезирующие пигменты, использовать энергию солнечной радиации для синтеза органических молекул из неорганических веществ — диоксида углерода, азота и серы. Продукты фотосинтеза служат затем не только исходными веществами, но и источником химической энергии для всех последующих биосинтетических реакций. Обычно принято описывать фотосинтез только как процесс образования углеводов в некоторых случаях основными продуктами фотосинтеза, действительно, являются исключительно крахмал, целлюлоза и сахароза, однако в других организмах на синтез углеводов идет, быть может, всего лишь третья часть углерода, связываемого и восстанавливаемого в процессе фотосинтеза. При ближайшем рассмотрении оказывается, что нельзя провести четкую границу между образованием продуктов фотосинтеза и другими биосинтетическими реакциями в клетке, в которых могут участвовать промежуточные вещества фотосинтетического цикла восстановления углерода. [c.396]
Углеводы — это обширный класс органических соединений с эмпирической формулой С (Н, 0) , образование которых связано с процессом фотосинтеза. Углеводы в растениях находятся в виде моносахаридов (глюкоза — С Н О ), олигосахаридов (крахмал) и полисахаридов (целлюлоза — (С Н О ) , где п > 10000. Целлюлоза — основной строительный материал растительных тканей. Она выполняет в растениях опорные функции и придает им механическую прочность. По распространенности органических веществ на земном шаре она занимает первое место. [c.47]
Магний — составная часть хлорофилла. Он участвует в процессе фотосинтеза, в образовании или распаде углеводов и жиров, в превращениях фосфорных соединений. Недостаток магния в почве вызывает заболевания растений (хлороз, мраморность листьев и др.). При низком содержании его в кормах наблюдаются заболевания сельскохозяйственных животных. Магниевым микроудобрением служит доломит. [c.259]
После завершения этих реакций наступает темновая стадия процесса фотосинтеза, сущность которой состоит в передаче водорода молекулой восстановленного хлорофилла молекуле СО2 с образованием органических соединений типа углеводов. Этот процесс совершается под действием соответствующих ферментов по схеме [c.178]
В данной главе мы бросили беглый взгляд не некоторые важнейшие составляющие биосферы-той части физического мира, в которой протекают жизненные циклы организмов. Наряду с соответствующими условиями окружающей среды для поддержания жизни необходим какой-либо источник энергии. Первичным источником необходимой энергии является Солнце. В процессе фотосинтеза растения превращают солнечную энергию в химическую. Солнечная энергия поглощается растительным пигментом хлорофиллом и затем используется для образования углевода глюкозы и О2 из СО2 и Н2О. [c.464]
Нетрудно видеть, что перенос макроэргических связей обеспечивает энергетическое сопряжение реакций и делает возможным использование одного или немногих ве- ществ для энергетического питания целой системы реакций. Понятно, что для обеспечения развитой кодированной формы управления потоком энергии лучше всего все порции энергии, получаемые извне, запасать в виде энергии макроэргических связей, а затем извлекать энергию по мере надобности и в той форме, в какой она будет нужна. В действительности нет необходимости всю энергию запасать именно в молекулах АТФ. Вполне достаточно иметь запас какого-либо вещества, которое, легко взаимодействуя с АДФ, может обеспечить быстрое образование активных и мобильных молекул АТФ тогда, когда потребуется произвести какую-либо работу. Так оно на самом деле и происходит. В процессе фотосинтеза, например, молекулы АТФ играют важную роль, улавливая энергию возбужденных светом электронов и направляя ее на синтез углеводов, но запасается в клетках не АТФ, а крахмал, который и служит для сохранения больших количеств энергии. [c.148]
Фотохимические процессы имеют огромное значение для жизни на Земле. Энергия Солнца утилизируется в процессе фотосинтеза, при этом из атмосферного углекислого газа образуются углеводы и освобождается кислород. Светоиндуцированные химические изменения, происходящие в атмосферных газах и распыленных частицах, также дают вклад в химический состав атмосферы, делая ее пригодной для существования жизни на Земле. В самом деле, образование из простейших элементов сложных биоорганических соединений — кирпичиков жизни, а затем и возникновение самой жизни тесно связаны с фотохимическими процессами. Важнейший для жизнедеятельности человека и многих других существ процесс — зрение — также имеет фотохимическое происхождение. Таким образом, природа использует свет для осуществления весьма важных химических процессов. Человек использует свет в различных областях от создания новых сложных органических соединений и различных систем передачи изображения (фотографии) до накопления солнечной энергии. [c.7]
При восстановлении 1 моля СО2 до углеводородного уровня поглощается энергия 469 кДж. При фотосинтезе выделяется свободный кислород, поступающий в атмосферу. Продуктами фотосинтеза являются не только углеводы, но и растительные белки. Весь процесс фотосинтеза в современных зеленых растениях имеет довольно сложный характер. Простым продуктом фотосинтеза является глюкоза, образование которой происходит согласно уравнению [c.359]
Образование углеводов у высших растений совершается в процессе фотосинтеза из углекислого газа и воды при участии зеленого пигмента растений — хлорофилла. [c.258]
Способность восстанавливать углеродные соединения с помощью солнечной энергии позволила появившемуся живому организму усваивать двуокись углерода, возможно в виде иона карбоната или бикарбоната, из окружающего первичного океана и использовать эту двуокись углерода в качестве источника атомов углерода для образования молекул сахаров и других питательных молекул. Эта реакция составляет сущность процесса фотосинтеза. В зеленых растениях при фотосинтезе происходит восстановление двуокиси углерода до альдегида (углевода), при котором вода служит первичным источником атомов водорода, а в атмосферу выделяется высвобождаемый при таком восстановлении кислород. Так же как и при анаэробном метаболизме (см. разд. Первичный метаболизм ), каждая реакция, входящая в процесс фотосинтеза, требует участия строго специфического фермента. [c.39]
Запасные полисахариды присутствуют в растениях в коллоидном состоянии или в водонерастворимой форме, благодаря чему они могут накапливаться в растительных клетках в большом количестве, не влияя на осмотическое давление. Крахмал — наиболее важный и накапливающийся в наибольшем количестве запасный полисахарид в мире растений. У всех растений — от низших водорослей до некоторых высших растений, главным образом двудольных,— углеводы, образовавшиеся в процессе фотосинтеза в хлоропластах, немедленно превращаются в крахмал (фото 46). Такой крахмал называют ассимиляционным. Согласно Смиту [160], у подсолнечника в крахмал превращается почти весь ассимилированный углерод. Однако ассимиляционный крахмал представляет собой довольно лабильную, переходную форму он либо довольно быстро используется в процессах метаболизма, либо превращается в ряде органов, например в семенах, плодах, стеблях, листовых влагалищах и корнях, в запасный крахмал. Эти общие метаболические особенности присущи так называемому крахмалистому листу. Напротив, в сахаристом листе злаков (однодольные растения) крахмал почти не обнаруживается. Сахара здесь представлены главным образом сахарозой и различными моносахаридами они транспортируются в другие части растения и превращаются в запасный крахмал в специальных органах. Например, энергичный синтез крахмала обычно имеет место в листовых влагалищах и в семенах злаков, начиная от периода цветения и кончая периодом созревания зерна. В ряде работ показано, что образование крахмала в зерне ячменя, риса и ржи в стадии налива специфически связано с ассимиляционной активностью верхних листьев и колоса, но не с ассимиляционной активностью расположенных ниже листьев [8, 144]. [c.140]
В книге Основы фотосинтеза в сжатой и доступной форме излагаются основные вопросы биофизики, биохимии и физиологии фотосинтеза представления об эволюция фотосинтеза, химяческ01м составе и структуре фотосинтетического аппарата, поглощении квантов света пигментами, окислительно-восстановительных реакциях, протекающих с участием хлорофилла, о поглощении и выделении нислорода хлоропластами, фотосинтетическом фосфорилировании и механизме образования АТФ, реакциях усвоения СОз и образовании в процессе фотосинтеза углеводов, органических кислот, аминокислот, о регуляции фотосинтеза, оттоке ассимилятов, связи фотосинтеза и урожая. [c.2]
УГЛЕВОДЫ. Органические вещества, состоящие из углерода, водорода и кислорода. У большинства углеводов водород и кислород содержатся в том же соотношении, что и в воде. Широко распространены в природе и играют очень важную роль в жизни растений, животных и человека. Образование У. в зеленом растении происходит в процессе фотосинтеза из углекислого газа и воды. В растениях на долю У. приходится до 85—90% сухого вещества. Они выполняют в растениях опорную (скелетную) функцию и являются основным энергетическим материалом. В животном организме в среднем содержится до 2% У., но в некоторых органах (печень) может содержаться до 10%. У.— основная составная часть пищи человека и животных. [c.299]
Многие углеводы и другие вещества, образованные при фотосинтезе, подвергаются сложным превращениям в процессе дыхания растений. Глюкоза — конечный продукт фотосинтеза — содержит значительное количество солнечной энергии, заключенной в ее молекуле. [c.11]
Реакции р -карбоксилирования, приводящие к образованию новых ди- и трикарбоновых органических кислот, играют большую роль в процессе фотосинтеза, в метаболизме углеводов, белков, жиров, в образовании органических кислот у суккулентов и т.д. [c.238]
Определенные формы живых организмов могут использовать эту энергию непосредственно для преобразования одних молекул в другие, более богатые энергией, с использованием углекислого газа как единственного источника углерода. Такими формами являются все фотосинтезирующие растения, которые в процессе фотосинтеза осуществляют образование углеводов и свободного кислорода из двуокиси углерода и воды. Это так называемые автотрофы. [c.111]
Система тилакоидных мембран хлоропласта превраш,а-ет энергию света в форму, которая может быть использована для осушествления химических реакций. Целиком процесс фотосинтеза был схематически представлен на рис. 10.1. В приводимом ниже обсуждении фотосинтеза рассматриваются три стадии. Первая стадия представляет собой световую реакцию — первичный процесс, с помош,ью которого энергия света поглощается светособирающими пигментами и переносится на фотохимические реакционные центры. На второй стадии поглощенная энергия света используется для осуществления транспорта электронов от воды до NADP+. В ходе электронного транспорта устанавливается градиент заряда, или концентрации протонов, через функциональные везикулы мембраны. Третья стадия представляет собой путь, по которому NADPH, образованный электронтранспортной системой, и АТР, генерируемый за счет различий электрохимического потенциала протонного градиента, используются для фиксации СО2 и синтеза углеводов. Хотя в целях упрощения процесс фотосинтеза разбит на три стадии, необходимо помнить, что поглощение света, транспорт электронов и генерация электрохимического градиента в действительности очень тесно сопряжены. [c.333]
Методом меченых атомов установлено, что в процессе фотосинтеза в растениях создаются разнообразные вещества, количество и состав которых изменяется в зависимости от вида растения, возраста, условий существования и др. Оказалось, что в фотосинтезе большую роль играет спектральный состав света и его интенсивность. Например, красно-желтая часть спектра способствует синтезу главным образом углеводов синяя часть спектра — образованию белков. Это создает возможность более эффективного использования фотосинтеза в тепличном хозяйстве, где можно создавать различные условия освещения и управлять качественной и количественной стороной этого процесса. [c.485]
Под действием света могут идти как реакции разложения молекул на атомы (фотолиз), так и реакции образования новых молекул, нередко более сложных, чем исходные (фотосинтез). Особенно большое значение имеют реакции фотосинтеза, происходящие в растениях под действием солнечного света. В результате фотосинтеза образуются различные органические соединения, главным образом углеводы — крахмал, клетчатка. Познание сущности процесса фотосинтеза — одна из важнейших проблем современного естествознания. [c.121]
После рассмотрения сравнительно простых реакций образования отдельных моносахаридов необходимо остановиться на весьма сложном, но исключительно важном процессе фотосинтеза. Это нужно сделать не только потому, что конечные продукты фотосинтеза — углеводы [c.204]
Радиоизотопы, которые можно использовать в качестве меток даже при очень небольших концентрациях, помогают изучать самые разнообразные процессы, происходящие в живых организмах. В живых организмах они усваиваются так же, как обычные нерадиоактивные атомы того же элемента. Например, соединения, содержащие углерод 0 , усваиваются точно так же, как и соединения, в состав которых входит только устойчивый изотоп углерода Введенные в живой организм меченые соединения принимают участие в образовании углеводов, жиров и белков, так как у обоих изотопов электронные оболочки, а следовательно, и химические свойства одинаковы. Углерод дает постоянное излучение и его путь в виде различных веществ в организме можно проследить с помощью счетчика Гейгера. Углерод С применяется для изучения процессов фотосинтеза и превращения жиров, углеводов и белков в живых тканях. С помощью изотопа железа Ре изучались процессы образования красных кровяных телец 145 [c.145]
Процесс фотосинтеза может быть выражен суммарным уравнением (1), которое отражает тот хорошо известный факт, что для осуществления в растениях фотосинтеза необходима вода и что в качестве побочного продукта реакции выделяется кислород (из воды). В фотосинтезирующих бактериях кислород не образуется и используются другие доноры водорода [НгХ например, h3S или лактат СИзСН (ОН) 0 см. уравнение (2)). Хилл в 1937 г. и Арнон в 1954 г. показали, что образование NADPH и АТР, необходимых для связывания диоксида углерода, не зависит от их использования в фотосинтетическом цикле восстановления углерода. Эти наблюдения позволили формально разделить реакцию фотосинтеза на световую реакцию (образование NADPH и АТР) и темновую реакцию, в которой диоксид углерода превращается в углевод. [c.397]
Ранее предполагалось, что описанный выше процесс полимеризации формальдегида до сахаров имеет также значение с физиологической точки зрения и что аналогичным образом происходит образование углеводов при процессах ассимиляции в зеленых растениях (Байер, Вильштеттер и Штолль, Варбург). Однако в настоящее время считают, что при быстром фотосинтезе в качестве одного из первоначальных продуктов реакции образуется фосфоглицериновая кислота Н20зР0СН2СН(0Н)С00Н (Кальвин), из которой в растениях получаются углеводы (стр. 984) [c.212]
Процесс Ф. состоит пз реакций двух типов — фотолиза воды (разложения ее под действием света) и восстановления углекислого газа. Свет необходим лишь для первой реакции, а реакции восстановления СОг — темповые , т. е. идут без доступа света. В настоящее время принимается, что первым продуктом фотосинтеза является фосфоглицериновая кислота, которая затем превращается в сахарозу, крахмал и другие углеводы. Процессы образования углеводов при Ф. состоят пз большого числа реакций, идущих при участии многочисленных ферментов. Наряду с углеводами в процессе Ф. образуются и другие соединения, в частности аминокислоты. Результатом первичной фотохимической реакции Ф. является фосфорилирование адепозиндифосфорной кислоты с образованием аденозинтрифосфорной кислоты (АТФ), богатой энергией. В ходе этой реакции используется только часть поглощенной световой энергии, а другая ее часть расходуется на образование фермента — восстановителя — п на выделение кислорода. Углеводы же синтезируются из СОг за счет энергии АТФ при участии восстановителя. [c.329]
Диффузия играет большую роль на многих стадиях процесса фотосинтети-ческого включения углерода СОг в углеводы. При этом углекислый газ диффундирует из атмосферы, достигая поверхности листа, а затем проходит через усть-ичные отверстия. Войдя в лист, СО2 диффундирует по межклеточным воздухоносным пространствам, а затем через клеточные оболочки и плазму клеток ме.зо-филла листа. Далее углекислый газ, по-виднмому, в форме НСОг диффундирует через цитоплазму и достигает хлоропластов. Затем СО2 оказывается в хлоропласте и попадает в зону действия ферментов, участвующих в образовании углеводов. Как видно, одну только эту сторону фотосинтеза можно расчленить на много стадий, в каждой из которых важную роль играет диффузия. Если бы с помощью ферментов фиксировался весь углекислый газ, находящийся в сфере их действия, и не происходила бы диффузия новых количеств углекислого газа из атмосферы, окружающей растение, процесс фотосинтеза прекратился бы. Диффузия важна также для многих других аспектов физиологии растений, особенно для проникновения веществ через мембраны. [c.17]
Несмотря на такое значение фоторадиолиза воды и биокаталитического ее образования при дыхании из свободного кислорода и атомов водорода, отнимаемых от молекул пищевых веществ при содействии дегидрогеназ, наука наша имеет пока все еще далеко не полные сведения о сложнейших тайнах протекания процессов фотосинтеза углеводов, белков и жиров, а также процессов дыхания. [c.350]
Известно, что процесс фотосинтеза протекает с участием хлорофилла, непременной составной частью которого являются ионы магния. Магний оказывает большое влияние на образование углеводов в растениях и, следовательно, на плодообразование. Недостаток магния в почвах выражается в появлении на листьях мраморо-видности — белесой пятнистости, в их скручивании и пожелтении. Это начинается с краев нижних листьев. Листья при недостатке магния становятся хрупкими. При недостатке магния замедляется рост и вегетация растений, а при большом его дефиците в почве — растение вовсе не вступает в фазу плодоношения. [c.127]
Процесс фотосинтеза чрезвычайно важен для жизни на нашей планете, поскольку он осуществляет утилизацию имеющейся в изобилии солнечной энергии в химически доступную форму путем фиксации СОг с образованием углеводов. Пигменты, и в первую очередь хлорофилл, играют главную роль в фотосинтезе, и поэтому данный процесс еще долго будет оставать- [c.365]
Для исследования процессов образования полисахаридов большое значение имеет использование меченых соединений, например Oq, меченых сахаров и их производных. Предполагается, что биосинтезирующая система растений не различает соединений, содержащих С и С, и в обоих случаях реакции протекают одинаково, Включение СОз в состав воздуха, окружающего растущее растение, и анализ образцов, собранных через определенное время после начала эксперимента, позволили сделать ряд интересных выводов. За очень короткие промежутки времени в процессе фотосинтеза образуется большое количество меченых соединений. Тем не менее у большинства растений основная масса С уже в первые минуты концентрируется в углеводах. Когда в питательную среду вводится радиоактивная глюкоза, радиоактивность прежде всего обнаруживается в полисахаридах и только потом — в иротеинах, липидах и других полимерах. Если учесть, что глюкоза и ее соединения являются исходным веществом для синтеза большинства биополимеров, то можно полагать, что путь иревра- [c.24]
Было высказано предположение, что первичная световая реак ция в фотосинтезе и реакции Хилла заключается в фотолизе водь для создания восстановительного потенциала водорода и окисли тельного потенциала гидроксила. При фотосинтезе водород в конеч ном счете восстанавливает углекислоту с образованием углеводов, а при реакции Хилла водород восстанавливает добавленный окислитель. В обоих случаях гидроксил в конечном счете освобождает молекулярный кислород. Эти реакции представлены в табл. 23. Согласно предложенной схеме, весь кислород, выделяемый при фотосинтезе, происходит из воды. Используя НгО , удалось показать, что кислород, выделяемый в процессе фотосинтеза, действительно происходит из воды, а не из углекислого газа. [c.261]
Магний входит в состав хлорофилла и, следовательно, участвует в процессе фотосинтеза. Он активирует ферменты — киназы, которые отщепляют фосфорную кислоту от аденозинтрифосфата (АТФ) и переносят ее на молекулы сахара и другие соединения с образованием фосфатных эфиров аденозиндифосфата (АДФ). Магний усиливает восстановительные процессы в растениях, оказывает сильное влияние на образование углеводов, плодооб-разование и превращение минеральных солей фосфора в слоншые органические соединения. До 50% его находится в растении в виде [c.29]
Ио предложенной авторами схеме, восстановление СО2 в процессе фотосинтеза до уровня сахаров протекает через ряд ступеней. Карбоксильная группа образовавшейся карбоновой кислоты на свету может восстановиться водородом воды в спиртовую группу -СН2ОН. К спиртовой группе может присоединиться новая молекула СО2, которая в свою очередь тоже восстановится на свету. Таким образом, происходит удлинение цепочки до образования соединений типа углеводов. Когда наращивается цепочка достаточной длины и ООразуется соединение типа гексозы или триозы, происходит отделение ее от радикала. Триоза затем конденсируется в глюкозу [c.241]
Указанные процессы, связанные с непосредственным образованием в процессе фотосинтеза других продуктов, кроме углеводов, нашли свое отражение в более поздних схемах фотосинтеза Кальвина с сотрудниками ( ualvin, ВаввЬаа, 1962 Бассем, Кальвин, 1962). В своих первых работах эти исследователи фиксировали основное вни1 ание на синтезе углеводов и приводили только один конечный продукт - сахарозу. В более поздних схемах указаны Оба возможные пути превращения 3-ФГК восстановление ее в 3-ФГА приводит к образованию углеводов превращение ее в Ш и ФЕП приводит к образованию органических кислот, аминокислот и др. [c.255]
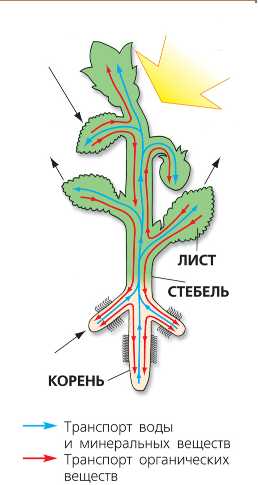
Выделительные ткани — вместилище смол, эфирных масел и других веществ. Ассимиляционная — паренхимная зеленая ткань (хлоденхима), в ее зеленых пластидах — хло-ропластах осуществляется важнейший процесс — фотосинтез (образование сложных органических веществ из воды и углекислоты при участии лучистой энергии). В результате этого процесса в листьях образуются в основном углеводы — сахара, которые, передвигаясь по ситовидным трубкам, поставляют образовательным тканям — камбию и другим сырье для построения новых растительных клеток и их оболочек. [c.24]
Восстановительная способность восстановленного НАДФ и энергия АТФ используются для удаления кислорода из ФГ (восстановление). Реакция протекает в две стадии на первой используется некоторое количество АТФ, произведенного в световых реакциях, а на второй — весь восстановленный НАДФ, полученный в этих реакциях. Суммарный итог — восстановление карбоксильной кислотной группы (—СООН) до альдегидной (—СНО). Продукт представляет собой ЗС-фосфат сахара (триозофосфат), т. е. сахар с присоединенной фосфатной группой. Это соединение обладает большей химической энергией и является первым углеводом, образованным в процесс фотосинтеза. [c.268]
Синтез органических веществ в зеленых растениях из углекислого газа и воды с использованием световой энергии носит иазвание фотосинтеза. Процесс фотосинтеза является основным источником образования органических веществ на нашей планете и, с этой точки зрения, вполне объясним интерес, который проявляют к нему представители различных отраслей естество- знания (биологи, химики, физики). Благодаря исследованиям М. Ненцкого, К. Тимирязева, Р. Вильштеттера, Г. Фишера, М. Цвета и др. изучена химическая природа хлорофилла, играющего роль фотосенсибилизатора. Хлорофилл, нерастворимый в воде зеленый иигмент, в зеленых растениях находится в особых образованиях — хлоропластах. Хлоропласты содержат до 75% воды. Сухое вещество хлоропластов состоит из белковой основы (стромы), хлорофилла, фосфатидов, каротиноидов, минеральных веществ, углеводов и т. д. Хлорофилл в хлоропластах содержится в отдель Ш1х гра нулах в сочетании с белками и липидами. [c.229]
Обилий энергообмен земных организмов можно упрош енно представить как образование в фотосинтезе сложных молекул углеводов из СОг и ПгО с после-дуюш ей деградацией продуктов фотосинтеза в процессах дыхания. Именно этот энергообмен обеспечивает суш ествование и развитие как отдельных организмов — звеньев в круговороте энергии, так и в жизни на Земле в целом. С этой точки зрения, уменьшение энтропии живых систем в процессах их жизнедеятельности обусловлено в конечном итоге поглош ением квантов света фотосинтезируюш ими организмами, что, однако, с избытком компенсируется образованием положительной энтропии в ядерных реакциях на Солнце. Этот принцип относится и к отдельным организмам, для которых поступление извне питательных веш еств, несуш их приток отрицательной энтропии, всегда сопряжено с продуцированием положительной энтропии при их образовании в других участках внешней среды, так что суммарное изменение энтропии в системе организм - - внешняя среда всегда положительно. Точно так же уменьшение энтропии в части клетки, где идет биохимический синтез, происходит за счет избыточного увеличения энтропии в других частях организма или среды. [c.125]
chem21.info
Образование углеводов и органических кислот
Образование углеводов и органических кислот
Углеводы являются важнейшей составной частью всех растений. Они образуются в зеленых растений в результате фотосинтеза из углекислого газа и воды под действием световой энергии. При этом углерод из неорганической формы переходит в органическую.
По современным представлениям фотосинтез представляет собой цепь окислительно-восстановительных реакций с поглощением немалого количества солнечной энергии. Суммарно уравнение фотосинтеза имеет вид
6 CO2 + 6 h3O С6h22O6 + 6 O2
Механизм этого процесса следующий: при фотосинтезе происходит разложение воды, и образующийся при этом водород идёт на восстановление углекислого газа. Происходит превращение кинетической энергии солнца в потенциальную химическую энергию, которая аккумулируется сложными органическими веществами.
Фотосинтез имеет огромное биологическое значение: благодаря ему ежегодно связывается около (15÷20)·1010 тонн углерода и выделяется в атмосферу 4·1011 тонн кислорода. Фотосинтез проходит в хлоропластах – клеточных органоидах, главной составной частью которых является хлорофилл.
Главным продуктом фотосинтеза являются углеводы (Гексозы). Они образуются в листьях виноградного куста, откуда транспортируются в ягоду. Гексозы (C6h22O6) представлены глюкозой и фруктозой. Это кристаллические вещества с молекулярной массой 180, хорошо растворимые в воде и спирте. Плоскость поляризации глюкоза вращает вправо, а фруктоза влево. Оба сахара хорошо сбраживаются дрожжами.
К другим (несбраживаемым) углеводам виноградной грозди относятся Пентозы (С5h20O5), Пентозаны (С5H8O4) n, Целлюлоза (C6h20O5) n, Пектиновые вещества, камеди, декстраны. Целлюлоза является основой твердых частей ягоды. Они состоят из большого числа (до 10000) остатков глюкозы. Пентозаны содержатся в твердых частях виноградной грозди: в семенах (4,0 – 4,5 % от всей массы), гребня (1,0 – 2,8 %), кожице (1,0 – 1,6 %).
При созревании винограда нерастворимый протопектин переходит в растворимый пектин, чем обусловлено в основном размягчение ягод. При брожении и спиртовании сусла он выпадает в осадок.
В последнее время проводятся исследования по изучению связи между метаболизмом углеводов и органических кислот в процессе роста и созревания ягод винограда. Первоначальный распад органических кислот и аккумуляция сахаров происходят одновременно в начале созревания. Обычно это длится 6 – 7 недель после цветения; это процесс интенсифицируется в середине срока созревания ягод винограда.
Органические кислоты образуются в процессе дыхания растений за счет неполного окисления углеводов, а также по пути синтеза аминокислот.
Органические кислоты образуются и при фотосинтезе, главным образом в листьях, откуда они транспортируются в ягоды винограда.
Ж. Риберо-Гайон и П. Риберо-Гайон, изучив механизм синтеза винной кислоты, установили, что она образуется из глюкозы по схеме:
CHO COOH
׀ ׀
CHOH CHOH COOH COOH
׀ ׀ ׀ ׀
CHOH CHOH CHOH CHO CHOH
׀ ׀ ׀ + ׀ ׀
CHOH CHOH CHOH Ch3OH CHOH
׀ ׀ ׀ ׀
CHOH CO CHO COOH
׀ ׀
Ch3OH Ch3OH
Глюкоза Кето – 5- Альдегид Гликолевый Винная
глюконовая винной альдегид кислота
кислота кислоты
Синтез яблочной кислоты происходит несколькими путями. Один из них – карбоксилирование пировиноградной кислоты:
Ch4–CO–COOH + CO2 COOH–Ch3–CO–COOH
Пировиноградная Щавелевоуксусная
Кислота кислота
COOH–Ch3–CO–COOH+HAD·h3COOH–Ch3–CHOH–COOH+HAD
яблочная кислота
При помощи радиоактивного изотопа углерода 14С было доказано, что винная кислота расходуется главным образом на дыхание, тогда как яблочная кроме дыхания идёт на образование сахаров, других органических кислот и аминокислот.
В соке зрелого винограда в зависимости от его сорта и района произрастания винной кислоты содержится 40-95 % от суммы всех кислот. В сортах винограда, в которых преобладает винная кислота, в ходе созревания величина титруемой кислотности ниже, но более постоянна и не так зависти от метеорологических условий года.
В ходе созревания винограда содержание яблочной кислоты снижается значительно быстрее винной и в некоторых сортах винограда её может вовсе не остаться. Снижение яблочной кислоты происходит особенно сильно при высоких температурах воздуха в период созревания обычно в винограде, созревшем для приготовления столовых вин, яблочной кислоты остается 20-50 % от всех кислот. То же самое наблюдается и при уборке шампанских сортов винограда. Поэтому проведение процесса ЯМБ (яблочно-молочное брожение) является обязательным этапом в технологии первичного виноделия этих вин.
Наиболее глубоким и продолжительным неферментативным окисляющим действием характеризуются фенольные вещества.
Добавить комментарий
vinograd-vino.ru
Образование углеводов в процессах фотосинтеза и хемосинтеза — Мегаобучалка
 Фотосинтез. Фотосинтез — это совокупность процессов, в ходе которых запасается солнечная энергия в виде химических связей органических соединений, синтезируемых из неорганических веществ.
Фотосинтез. Фотосинтез — это совокупность процессов, в ходе которых запасается солнечная энергия в виде химических связей органических соединений, синтезируемых из неорганических веществ.
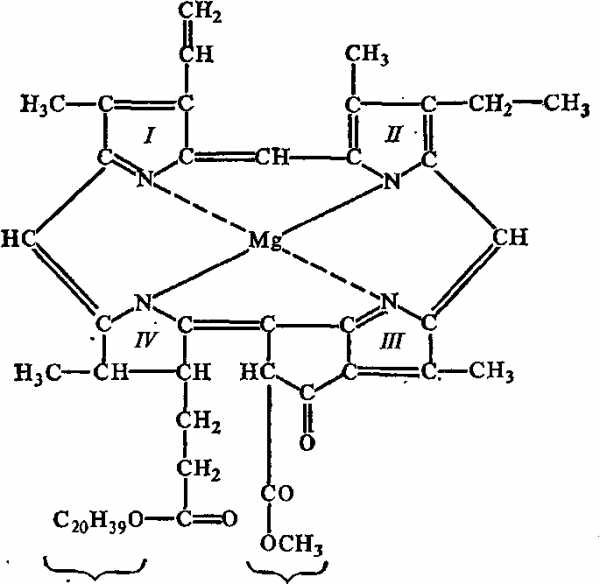
Рис. 15. Строение хлорофилла а: I—IV — пиррольные циклы; пунктиром показаны координационные связи
Фотосинтез состоит из двух фаз: световой (фотофизический и фотохимический этапы) и темновой. В ходе световой фазы происходит поглощение солнечной энергии хлорофиллом и передача ее в «реакционный центр», где в результате химических реакций, включающих транспорт электронов между различными переносчиками и сопряженного с ним фосфорилирования, образуются восстановительные и энергетические эквиваленты (НАДФН и АТФ). Для нормального функционирования реакций световой фазы кроме света необходимы хлорофилл и вода (или другой источник водорода). Различные хлорофиллы способны поглощать кванты света с определенной длиной волны и переходить при этом в возбужденное состояние. Переход в исходное состояние приводит к высвобождению энергии, которая через ряд промежуточных стадий запасается в форме АТФ и НАДФН.
Все известные в настоящее время фотосинтезирующие организмы содержат хлорофиллы — зеленые магнийпорфириновые пигменты. Известно свыше десяти их видов, различающихся природой химических группу присоединенных к пиррольным структурам порфиринового ядра, окраской, распространением среди живых организмов. Так, у всех зеленых растений содержатся хлорофиллы а и b (рис 17), в диатомовых водорослях — хлорофилл с, в красных водорослях — хлорофилл d. В клетках пурпурных бактерий обнаружены бактериохлорофиллы a и b, а в зеленых бактериях — бактериохлорофиллы с и d. Важным свойством молекул хлорофилла является их способность взаимодействовать с белками и друг с другом, образуя агрегированные формы с различными спектрами поглощения.
Наряду с зелеными пигментами в хлоропластах и хроматофорах содержатся каротиноиды — желтые и оранжевые пигменты
Структура β-каротина
изопреновой природы. Каротиноиды могут быть разделены на ряд групп по своему строению: собственно каротиноиды, гидроксилсодержащие каротиноиды и каротиноиды, содержащие карбонильные группы. Основными представителями у высших растений являются β-каротин и ксантофилл. Предполагают, что каротиноиды, используя лучи, не поглощаемые хлорофиллом, передают их энергию на молекулы хлорофилла. Существуют данные, что каротиноиды предохраняют молекулы хлорофилла от разрушения в процессе фотоокисления, а также играют определенную роль в половом процессе при прорастании пыльцы и росте пыльцевых трубок, у высших растений и при созревании половой клетки у водорослей и грибов.
Третья группа пигментов — фикобилины. Это красные и синие пигменты (фикоэритрины, фикоцианины), содержащиеся в хроматофорах некоторых водорослей. В основе химического строения фикобилинов лежит та же тетрапиррольная структура, но пиррольные группы расположены линейно. Фикобилины поглощают энергию света в зеленой и желтой областях спектра и передают ее на молекулу хлорофилла, после чего она используется в процессе фотосинтеза. Наличие фикобилинов у водорослей — пример приспособления в ходе эволюции к поглощению тех лучей солнечного света, которые проникают через толщу морской воды.
Темновая фаза фотосинтеза — это фиксация и восстановление СО2 с образованием углеводов и других конечных продуктов фотосинтеза. На этой стадии свет не нужен, а используются образованные в световой фазе восстановительные и энергетические эквиваленты. Во время темновой фазы атомы водорода, поставляемые световыми реакциями, используются для восстановления СО2 до углеводов согласно общему уравнению фотосинтеза:
Свет
6С02 + 12Н20 → С6Н12О6 + 602 + 6Н20
Хлорофилл
При этом на каждый моль синтезированного углевода запасается ~160 кДж энергии.
Процесс восстановления СО2 начинается с его присоединения к пятиуглеродному акцептору рибулозо-1,5-бисфосфату (РуБФ). Образующееся шестиуглеродное соединение очень нестойко. Опытами М. Кальвина с меченым оксидом углерода (IV) установлено, что при длительности фотосинтеза 2 с первым фиксируемым меченым соединением является не С6-соединение, а 3-фосфоглицериновая кислота (ФГК), причем метка обнаруживается только в карбоксиле. Реакция карбоксилирования катализируется рибулозобисфосфат-карбоксилазой (РБФ-карбоксилаза). Затем образующаяся ФГК фосфорилируется при участии фермента фосфоглице-раткиназы с использованием АТФ и превращается в 1,3-бисфосфо-D-глицериновую кислоту, которая более реакционноспособна и легче восстанавливается в глицеральдегид-3-фосфат (ГАФ).
В реакции восстановления принимают участие образовавшийся в световой стадии НАДФН (никотинамидадениннуклеотид) и фермент глицеральдегидфосфатдегидрогеназа. Часть образовавшихся молекул ГАФ под действием фермента триозофосфатизомеразы превращается в дигидроксиацетонфосфат (ДГАФ). Эти два триозофосфата конденсируются под действием альдолазы. Сначала образуется фруктозо-1,6-дифосфат (ФДФ), далее — монофосфаты фруктозы (Ф6Ф) и глюкозы (Г6Ф) и, наконец, — сахароза и крахмал. Для того чтобы процесс фотосинтеза продолжался, необходима постоянная регенерация РуБФ, поэтому в одном обороте цикла участвуют 6 молекул РуБФ, которые фиксируют 6 молекул С02. Образующиеся 12 молекул ФГК, а после ее восстановления 12 молекул ГАФ расходуются следующим образом:
5ГАФ → 5ДГАФ
ЗГАФ + ЗДГАФ -+ ЗФДФ →ЗФ6Ф
2ГАФ + 2Ф6Ф-→2 Ксилулоза-5Ф + 2Эритрозо-4Ф
2ГАФ + 2 Седогептулозо-7Ф 2 Ксилулозо-5Ф + 2 Рибозо-5Ф
Таким образом, две молекулы глицеральдегид-3-фосфата образуют одну молекулу гексозы, которая выходит из цикла, а 10 идут на регенерацию шести молекул рибулозо-1,5 бисфосфата, и цикл замыкается.
У некоторых растений в качестве первичных продуктов фотосинтеза сначала образуются оксалоацетат и малат в результате карбоксилирования ФЕП под действием ФЕП-карбоксилазы. Поскольку первичные продукты в этом случае содержат четыре атома углерода, его называют С4-путь фотосинтеза в отличие от цикла Кальвина, где образующаяся ФГК содержит три атома углерода (С3-путь). С4-Путь фотосинтеза, в котором принимают участие два типа клеток и два типа хлоропластов, называется кооперативным.
Хемосинтез. Гетеротрофная фиксация С02. Известный русский микробиолог С. Н. Виноградский показал (1856—1953), что органические вещества синтезируются в природе не только путем фотосинтеза в зеленых растениях, но и бактериями, не содержащими хлорофилла. Энергию, необходимую для синтеза органических соединений, эти бактерии получают при окислении различных неорганических соединений: Fe, N, S, Н, Sb, Мn. Этот процесс называется хемосинтезом. Некоторые хемосинтетики используют в качестве доноров водорода простейшие органические вещества — метан, метанол и пр.
Исследование химизма ассимиляции меченого оксида углерода (IV) (14С02) различными хемосинтезирующими бактериями показало, что первым стойким продуктом хемосинтеза является фосфоглицериновая кислота, а присоединение СO2 к рибулозобисфосфату, т. е. цикл Кальвина, — основным механизмом ассимиляции СО2. У многих хемосинтезирующих бактерий цикл Кальвина — главный, но не единственный путь образования органических веществ.
Таким образом, процессы фотосинтеза и хемосинтеза — источники органического вещества на Земле.
А. Ф. Лебедевым еще в 1914 г. была высказана мысль, что гетеротрофные организмы могут частично ассимилировать углерод не только из готовых органических соединений, но и из СО2, связывая последний с некоторыми кетокислотами. Впоследствии это получило экспериментальное подтверждение в исследованиях А. Л. Курсанова, Г. Вуда, С. Очоа и др.
Однако, как уже указывалось, между автотрофами и гетеротрофами существует коренное отличие: первые способны синтезировать органическое вещество полностью за счет неорганических веществ (СО2 и Н20), вторые усваивают СО2, используя готовое органическое соединение, например пируват.
6.6. Превращение углеводов в процессе пищеварения
Человек и животные не способны к первичному биосинтезу углеводов из неорганических веществ, они могут лишь образовывать их в процессе глюконеогенеза из других органических веществ (органических кислот, жиров, аминокислот), но главным источником углеводов является пища. Некоторые насекомые получают с пищей преимущественно одни углеводы (питающиеся древесиной, цветочным нектаром). Они настолько приспособились к подобному питанию, что погибают без углеводов. У ряда беспозвоночных животных существует хорошо выраженная сезонная периодичность содержания углеводов в организме. Так, у мидий и креветок осенью резко увеличивается количество углеводов, происходит запасание на зиму питательных веществ. Этому способствует и повышенное содержание углеводов в планктоне — пище мидий и креветок. Углеводы составляют существенную часть пищевого рациона человека и многих животных. На их долю приходится 60—70% общей суммы калорий пищи человека. Особенно много углеводов содержат крупы и макароны (65—75%), хлеб (около 50%), картофель (до 25%). Очень мало их в мясомолочных продуктах (0,5-2%).
Углеводы всасываются через слизистую оболочку кишечника только в виде моносахаридов. Даже хорошо растворимые дисахариды сахароза и лактоза не могут в неизмененном виде всасываться в кишечнике. Если их ввести в кровь, минуя желудочно-кишечный тракт, дисахариды не будут утилизироваться клетками различных тканей. Это еще в большей степени относится к нерастворимым в воде полисахаридам: крахмалу, гликогену. Таким образом, в процессе переваривания углеводов пищи должно происходить их расщепление до моносахаридов — той единственной формы, которая может всасываться и использоваться в организме.
Переваривание углеводов начинается в ротовой полости. В составе слюны содержатся два фермента: α-амилаза и мальтаза.
Специфической особенностью α-амилазы слюны (старое название — птиалин) является способность гидролитически расщеплять крахмал только тех пищевых продуктов, которые подвергались термической обработке при изготовлении пищи. «Сырой» крахмал во рту почти не гидролизуется, только при очень медленном жевании. α-Амилаза слюны млекопитающих в отличие от птиц высоко активна. Интересно, что у некоторых приматов (бабуины, резусы) этот фермент отсутствует, хотя у человека он очень активен.
Пища в ротовой полости находится недолго, после попадания ее в желудок постепенно пищевой комок пропитывается кислым желудочным соком. Низкие значения рН инактивируют α-амилазу слюны. Таким образом, ее гидролитическое действие осуществляется кратковременно, в ротовой полости расщепление крахмала и гликогена только начинается. В желудке амилолитические ферменты отсутствуют.
Основным местом переваривания крахмала и гликогена является тонкий кишечник, где на них действует α-амилаза поджелудочной железы. Она может расщеплять крахмал, не подвергшийся термической обработке при приготовлении пищи. Поэтому появление в кале непереваренных гранул крахмала — признак либо нарушения выделения, либо нарушения функции панкреатической α-амилазы. Установлено, что активность α-амилазы в кишечнике отдельных видов рыб зависит от типа питания: у видов, питающихся фитопланктоном, который богат крахмалом, она значительно выше, чем у потребляющих зоопланктон.
В поджелудочном соке содержится также фермент мальтаза, расщепляющий дисахарид мальтозу. Однако основная масса дисахаридов, образовавшихся в результате действия α-амилазы и поступивших с пищей, гидролизуется ферментами тонкого кишечника. Этот процесс происходит не в просвете кишечника, а в клетках слизистой оболочки. Там действуют мальтаза, изомальтаза и сахараза (инвертаза). Мальтаза и изомальтаза находятся обычно в прочном комплексе с инвертазой. Эти ферменты функционируют в щеточной кайме эпителия слизистой оболочки кишечника в количествах, обеспечивающих переваривание и усвоение пищи взрослого человека. Помимо перечисленных ферментов кишечный эпителий содержит β- галактозидазу с оптимумом рН 4,5, гетерогалактозидазу и лактазу.
В результате последовательного воздействия перечисленных ферментов углеводы превращаются в моносахариды. Они хорошо всасываются кишечной стенкой. Скорость их всасывания различна: наибольшая для галактозы, близка для глюкозы, примерно половину последней составляет для фруктозы, у маннозы и ксилозы около четверти от скорости всасывания глюкозы, самая малая у арабинозы. Пентозы и манноза проникают через эпителий путем облегченной диффузии, их поступление против градиента концентрации невозможно. Интенсивность всасывания пентоз и маннозы зависит поэтому от скорости удаления этих сахаров путем диффузии наружу, на серозную сторону клеток, откуда они быстро уносятся с током крови.
У человека и млекопитающих животных отсутствует фермент целлюлаза, вызывающий гидролиз клетчатки (целлюлозы). Не атакуются ферментами желудочно-кишечного тракта млекопитающих и растительные пентозаны. Только некоторые из них частично расщепляются бактериями в толстом кишечнике с образованием органических кислот, С02 и других веществ. Поскольку многие млекопитающие растительноядны, переваривание целлюлозы для них крайне необходимо. В связи с этим они имеют специализированный пищеварительный тракт, в высокой степени приспособленный для симбиотического переваривания целлюлозы. Так, у жвачных желудок состоит из нескольких отделов, первый и самый большой из которых называется рубцом. Он содержит в больших количествах бактерии и простейших типа инфузорий, продуцирующих фермент целлюлазу. Рубец можно сравнить с бродильным чаном, в котором пища, смешанная со слюной, подвергается интенсивной ферментации. Продукты брожения (главным образом, масляная, уксусная и пропионовая кислоты), всасываются и используются организмом, С02 и СН4 выводятся с отрыжкой. Очень важно, что образовавшиеся из целлюлозы органические кислоты могут непосредственно использоваться на биосинтез жиров, они являются у жвачных главным источником энергии (у крупного рогатого скота составляют до 70% всей необходимой организму энергии).
Реакция содержимого рубца у здорового животного при нормальной организации кормления постоянно поддерживается в пределах рН 6,5-7,4 и смещается в кислую сторону в период наиболее интенсивного сбраживания корма. В этот момент образование органических кислот брожения превалирует над их всасыванием и нейтрализацией.
Среда рубца чрезвычайно благоприятна для размножения микроорганизмов. рН находится в пределах 6,5-7,4, температура колеблется от 39 до 40 °С. Кислород, который токсичен для многих видов бактерий, в рубце почти отсутствует.
Интенсивность бродильных процессов в рубце очень велика. За сутки в нем образуется до 4 л летучих жирных кислот (ЛЖК). Общее количество ЛЖК и соотношение отдельных кислот зависит от структуры рациона. В большинстве случаев в рубце преобладает уксусная кислота. Наибольшее ее количество образуется при даче рационов, богатых клетчаткой. Обилие в рационе крахмалистых и сахаристых кормов благоприятствует образованию пропионовой кислоты.
В процессе жизнедеятельности микроорганизмов в рубце образуются также газы. Они необходимы для дальнейших реакций, в результате которых синтезируется ряд ценных питательных веществ. Количество и состав газов, образующихся через 2 - Зч после кормления, достигает у крупного рогатого скота до 25-35 л/ч. За сутки может образовываться до 700 л газов.
В рубце образуются аммиак, метан, двуокись углерода, азот, небольшое количество водорода, сероводорода и кислорода.
В итоге деятельности микрофлоры рубца каждый день в нем вырабатывается до 2,5кг бактериального протеина (400 г азота). Этот бактериальный протеин переваривается в тонком кишечнике и служит главным источником аминокислот для коровы.
Улучшение переваривания растительного материала с помощью симбиотического микробного брожения встречается и у некоторых птиц, например у белой куропатки, которая несколько зимних месяцев питается в основном почками и молодыми ветками. У большинства птиц семейства куриных имеются две слепые кишки для сбраживания целлюлозы. Локализация этого процесса в заднем отделе кишечника менее выгодна, чем в рубце, так как исключается возможность дальнейшего переваривания пищи при прохождении кишечника. У многих грызунов, кроликов и зайцев это компенсируется капрофагией, поеданием кала.
6.7. Регуляция постоянства содержания глюкозы в крови
Всосавшиеся из кишечника моносахариды (преимущественно глюкоза) через воротную вену доставляются прежде всего в печень, где часть глюкозы используется на образование запасных отложений гликогена. Содержание последнего в печени человека и животных зависит от режима питания; чем больше углеводов в рационе, тем больше гликогена в печени. Синтез и распад гликогена катализируются различными ферментами и контролируются независимо. Часть глюкозы используется самой печенью для получения энергии, необходимой для многочисленных реакций, протекающих в ней. Кроме того, определенное количество глюкозы в печени превращается в жиры. Так, при нормальном смешанном питании только 3—5% глюкозы превращается в гликоген (его среднее содержание в печени колеблется от 5 до 7%), 30% глюкозы идет на образование жиров и 60—70% окисляется до С02 и Н20. При обильном углеводном питании 10% глюкозы откладывается в печени в виде гликогена, 40%—превращается в жиры и 50% окисляется.
Сахар в крови, лимфе, спинномозговой жидкости представлен глюкозой, уровень которой очень тонко регулируется. Содержание глюкозы в артериальной и капиллярной крови здорового человека составляет 54—96 мг/100 мл при определении глюкозооксидазным методом и 80—120 мг/100 мл при определении по Хагедорну и Иенсену (вторым методом кроме глюкозы определяются одновременно некоторые другие редуцирующие вещества в крови). В венозной крови содержится меньше глюкозы, так как она потребляется тканями из крови. Единственный орган, выделяющий глюкозу в общую циркуляцию, — печень.
Инсулин секретируется β-клетками островков Лангерганса поджелудочной железы в форме проинсулина. Основным проявлением недостаточности инсулина является повышение уровня глюкозы в крови (гипергликемия), избыточное выведение глюкозы с мочой (глюкозурия) и понижение содержания гликогена в печени. Недостаточность инсулина подавляет биосинтез жирных кислот из глюкозы и ацетата, а также биосинтез белков; усиливает синтез ферментов, участвующих в глюконеогенезе; нарушает соотношение между гликолизом и глюконеогенезом.
Мышцы и печень при недостатке инсулина не способны использовать глюкозу даже при высоком ее содержании в крови. Недостаточность выделения инсулина лежит в основе возникновения заболевания— диабета. Многие последствия диабета напоминают эффект углеводного голодания, хотя содержание глюкозы в крови при этом заболевании резко повышается.
Глюкагон — гормон, продуцируемый α-клетками поджелудочной железы, активирует расщепление гликогена в печени, в результате чего освобождается глюкоза, поступающая в кровь.
Адреналин — гормон мозгового слоя надпочечников. Совместно с глюкагоном активирует гликоген-фосфорилазу печени и мышц, увеличивает содержание глюкозы в крови. Фермент встречается в двух формах — активной (фосфорилаза а) и неактивной (фосфорилаза b). Молекулярная масса фосфорилазы а составляет 380 000. Фосфорилаза а состоит из четырех субъединиц, каждая из которых содержит остаток фосфосерина и молекулу кофермента — пиридоксальфосфата, ковалентно связанную с остатком лизина. Фосфатаза фосфорилазы вызывает гидролитическое расщепление связи фосфатной группы с остатком серина и образование димеров.
Механизмы действия адреналина и глюкагона на поддержание уровня глюкозы в крови аналогичны.
Глюкокортикоиды (гормоны коркового слоя надпочечников) усиливают глюконеогенез — новообразование сахара из неуглеводов. Это приводит к увеличению количества глюкозы в крови и гликогена в печени. Соматотропный гормон гипофиза уменьшает утилизацию глюкозы периферическими тканями, усиливает глюконеогенез, угнетает продукцию инсулина, в итоге повышает содержание глюкозы в крови. Тироксин и трийодтиронин (гормоны щитовидной железы) в умеренных дозах усиливают всасывание моносахаридов в кишечнике. Избыток этих гормонов тормозит окислительное фосфорилирование и способствует повышению концентрации глюкозы в крови.
Содержание гормонов в крови находится под контролем центральной нервной системы, осуществляющей контроль за уровнем глюкозы в крови через эндокринную систему. Однако помимо опосредованного влияния на уровень глюкозы в крови центральная нервная система (ЦНС) осуществляет и прямое воздействие.
За счет контроля со стороны ЦНС и эндокринной системы в крови автоматически поддерживается постоянный уровень глюкозы, происходит саморегуляция, которая может осуществляться на различных уровнях. Так, например, кровь с избыточным содержанием глюкозы, достигая поджелудочной железы, непосредственно стимулирует выделение инсулина. Последний усиливает поглощение глюкозы тканями, использование ее на синтез гликогена, в результате концентрация глюкозы снижается до нормы.
6.8. Превращения углеводов, связанные с дыханием и брожением
Гликолиз
Стадии и реакции гликолиза. Гликолиз — это анаэробный процесс, приводящий к распаду одной молекулы глюкозы на две молекулы молочной кислоты. При этом освобождается энергия, которую организм аккумулирует в форме АТФ. Реакции гликолиза протекают в цитозоле, без потребления кислорода. В анаэробных условиях гликолиз — единственный процесс в организме животных, растений и многих бактерий, поставляющий энергию.
При аэробном расщеплении глюкозы один из конечных продуктов гликолиза — пировиноградная кислота окисляется до С02 и Н20 в цикле трикарбоновых кислот. Реакция этого цикла осуществляется в митохондриях (у бактерий — в соответствующих мембранных образованиях) при участии кислорода.
Гликолиз протекает в две стадии.
Первая стадия — подготовительная, или собирательная. На этой стадии различные гексозы вовлекаются в гликолиз. Хотя в реакциях гликолиза окисляется главным образом глюкоза, наряду с ней могут вступить в этот процесс другие различные гексозы, например фруктоза, манноза. При этом инертные молекулы гексоз активируются, фосфорилируются за счет АТФ, превращаются в глюкозо-6-фосфат. Этот этап заканчивается образованием глицеральдегид-3-фосфата.
Вторая стадия — окислительная. Глицеральдегид-З-фосфат окисляется до пировиноградной кислоты (пируват) или молочной кислоты (лактат). Энергия окисления накапливается в АТФ, образуются восстановительные эквиваленты НАДН.
Гексокиназа
АТФ + Глюкоза → АДФ + Глюкозо-6-фосфат
Эту реакцию катализируют ферменты двух типов, различающиеся по своей специфичности в отношении cахаров, — гексокиназа и глюкокиназа. Гексокиназа играет основную роль, именно она функционирует в большинстве клеток. Гексокиназа способна катализировать фосфорилирование не только D-глюкозы, но также и многих других гексоз, например D-фруктозы, D-маннозы и D-глюкозамина.
Глюкокиназа фосфорилирует только D-глюкозу, к тому же обладает к ней более низким сродством, чем гексокиназа. Глюкокиназа содержится в печени. Она вступает в действие только в том случае, когда концентрация глюкозы в крови очень высока. Обе киназы нуждаются в присутствии двухвалентных катионов (Mg24+ или Мп2+), которые связываются с АТФ, образуя истинные субстраты— MgАТФ2- или МnАТФ2-. В ходе гексокиназной реакции происходит значительное уменьшение свободной энергии, что обеспечивает необратимость этой реакции внутри клетки. Скорость реакции тормозится ее продуктом — глюкозо-6-фосфатом.
Образовавшийся при гликолизе в цитоплазме НАДН не может проникать внутрь митохондрий. В аэробных условиях он передает электроны и протон в дыхательную цепь митохондрий посредством глицерофосфатного челночного механизма. В цитоплазме НАДН реагирует с дигидроксиацетонфосфатом, образуя глицерол-3-фосфат, который легко проникает через мембрану митохондрий. Реакция катализируется цитоплазматической глицерол-3-фосфат — дегидрогеназой (НАД+):
Дигидроксиацетонфосфат + НАДН + Н+ ⇆ Глицерол-З-фосфат + НАД+
Внутри митохондрии глицерол-3-фосфат снова окисляется до дигидроксиацетонфосфата, но уже с помощью флавинового фермента— митохондриальной глицерол-3-фосфат — дегидрогеназы:
Глицерол-З-фосфат + ФАД⇆ Дигидроксиацетонфосфат + ФАДН2
Цитоплазматический НАДН восстанавливает щавелевоуксусную кислоту до яблочной, легко проникающей в митохондрии и там снова окисляющейся до щавелевоуксусной кислоты с передачей водорода в дыхательную цепь.
У растений в некоторых тканях гликолиз протекает даже при достаточном доступе кислорода. Особенно характерно такое однофазовое, «анаэробное дыхание» (или «аэробное брожение») для зародышевой ткани семян и внутренних тканей плодов растений вследствие недостатка в них кислорода. Гликолиз является основным путем катаболизма глюкозы гомоферментативных молочнокислых бактерий, дрожжей.
Брожение. Многие микроорганизмы (дрожжевые грибы, плесневые грибы, бактерии) получают энергию за счет брожения. Это наиболее примитивный способ получения энергии. Разнообразные брожения совпадают с гликолизом во многих своих стадиях.
Часто встречающийся тип брожения — молочнокислое. В результате гомоферментативного молочнокислого брожения из глюкозы образуется молочная кислота с почти 100%-ным выходом. Реакции этого процесса идентичны реакциям гликолиза. При гетероферментативном (смешанное) молочнокислом брожении из глюкозы кроме молочной кислоты образуются в значительном количестве другие продукты — уксусная кислота, этиловый спирт и оксид углерода (IV). Химизм этого процесса совершенно иной, в основе его лежит окисление глюкозы по пентозофосфатному пути.
Молочнокислое брожение протекает при силосовании кормов для сельскохозяйственных животных, квашении капусты. Образующаяся молочная кислота предотвращает развитие гнилостных бактерий, плесневых грибов, т. е. служит консервантом.
Пропионовая кислота представляет собой продукт сбраживания углеводов пропионовыми бактериями. Наряду с ней образуются уксусная кислота и оксид углерода (IV). На каждую молекулу ПВК, окисленную до уксусной кислоты и С02, приходится две молекулы ПВК, которые превращаются в пропионовую кислоту.
Уксуснокислые бактерии (рода Acetobacter) окисляют этиловый спирт до уксусной кислоты. Эти бактерии способны окислять и другие спирты, в том числе предельные многоатомные. Такое явление называется неполным окислением.
6.9. Глюконеогенез.
Анаэробная фаза расщепления глюкозы — гликолиз — заканчивается образованием ПВК или молочной кислоты. Они при определенных условиях могут вновь ресинтезироваться в глюкозу. Из двух молекул молочной кислоты образуется одна молекула глюкозы, т. е. происходит как бы обращение гликолиза. Этот процесс называется глюконеогенезом. Если гликолиз — центральный путь катаболизма углеводов, то глюконеогенез — анаболический процесс, наиболее важный общий путь биосинтеза моносахаридов и полисахаридов у человека, животных, многих бактерий. У фотосинтезирующих организмов он играет обычно второстепенную роль.
Большинство стадий глюконеогенеза представляет собой обращение реакций гликолиза. Однако существуют три необратимые стадии гликолиза, протекающие с выделением значительного количества энергии, поэтому глюконеогенез идет в обход этих стадий.
megaobuchalka.ru
Синтез углеводов и других веществ в растениях
В противоположность только что описанным процессам существует другой путь распада крупных молекул органических веществ на более мелкие частицы — анаэробное разложение. Он происходит лишь в отсутствии кислорода воздуха. Этот процесс известен также под названием гниения . В нем принимают участие живые организмы. Однако, в то время как в процессе аэробного разложения участвуют многие виды организмов, процессы гниения осуществляются лишь некоторыми видами бактерий. При поступлении воздуха в достаточном количестве бактерии гниения подавляются другими организмами, поглощающими кислород. Лишь когда эти организмы из-за недостатка кислорода погибают, могут размножаться анаэробные бактерии. Конечные продукты процессов гниения не похожи на продукты, образующиеся при аэробном разложении, так как при гниении кислород имеется лишь в таких количествах, в каких он уже имелся в исходных веществах. Напомним здесь еще раз о том, что при гниении не происходит окисления. Так, органически связанный азот не окисляется в нитрат, а появляется в конце процесса гниения в виде аммиака. Но в конце концов образующиеся вследствие анаэробного разложения частицы высокомолекулярных органических соединений вновь используются для синтеза новых белков, углеводов и жиров с помощью растений. Таким образом, мы видим, что и при втором пути распада круговорот органи- [c.23] В связи с важной ролью углеводов в обмене веществ и в жизнедеятельности растений изучение их синтеза и обмена постоянно находилось в центре внимания биохимиков. Некоторые из этих процессов были изучены довольно давно, другие — лишь Б последнее время, а детали химизма ряда сложных процессов еще неясны. [c.141]Синтез крахмала. Крахмал — основное запасное вещество больщинства растений, в которых он очень легко образуется как в процессе ассимиляции СО2, так и из других углеводов. Как мы видели, крахмал состоит из амилозы, представляющей собой неразветвленную цепь остатков а-глюкозы, соединенных [c.144]
Биосинтез начинается с фотосинтеза [1]. Вся жизнь на Земле зависит от способности некоторых организмов (зеленых растений, водорослей и фотосинтезирующих бактерий), содержащих характерные фотосинтезирующие пигменты, использовать энергию солнечной радиации для синтеза органических молекул из неорганических веществ — диоксида углерода, азота и серы. Продукты фотосинтеза служат затем не только исходными веществами, но и источником химической энергии для всех последующих биосинтетических реакций. Обычно принято описывать фотосинтез только как процесс образования углеводов в некоторых случаях основными продуктами фотосинтеза, действительно, являются исключительно крахмал, целлюлоза и сахароза, однако в других организмах на синтез углеводов идет, быть может, всего лишь третья часть углерода, связываемого и восстанавливаемого в процессе фотосинтеза. При ближайшем рассмотрении оказывается, что нельзя провести четкую границу между образованием продуктов фотосинтеза и другими биосинтетическими реакциями в клетке, в которых могут участвовать промежуточные вещества фотосинтетического цикла восстановления углерода. [c.396]
СИНТЕЗ УГЛЕВОДОВ И ДРУГИХ ВЕЩЕСТВ В РАСТЕНИЯХ [c.234]
Стереохимическая специфичность действия ферментов проявляется не только в реакциях гидролитического расщепления, но и в любых реакциях. При искусственном синтезе простейших углеводов, аминокислот и некоторых других веществ образуется смесь изомеров (рацематов), тогда как при ферментативных синтезах всегда образуется только один из оптических изомеров вешества. Растения и животные из -кислот и аммиака синтезируют аминокислоты -ряда. При фотосинтезе из углекислого газа и воды образуется >-глюкоза и т. д. [c.132]
Синтез углеводов. Синтез простых углеводов. Простые углеводы возникают главным образом при первичном биосинтезе органического вещества на Земле. Этот процесс осуществляется автотрофньшш организмами— растениями, а также фотосинтезирующими и хемосинтезирующими бактериями. Первичный синтез органического вещества в природе идет путем восстановления СОг атмосферы с одновременным формированием органических молекул, содержащих цепи углеродных атомов. В связях между атомами углерода и других элементов образуюхцихся органических соединений заключена энергия, поэтому их новообразование сопровождается ее поглощением. В общем виде процесс первичного новообразования органического вещества принято изображать в виде следующей схемы (см. с. 358). [c.357]
Калий, натрий и кальций находятся в растениях в виде солей органических и неорганических кислот. Соли калия оказывают влияние на синтез углеводов, белков и других веществ и способствуют оттоку образовавшихся в листе углеводов. При недостатке калия плохо развиваются опорные ткани и на растениях появляются бурые пятна. [c.295]
Магний входит в состав хлорофилла, усиливает синтез белков, углеводов, липидов и других веществ. Железо участвует в образовании хлорофилла, а также содержится в ряде дыхательных ферментов. Микроэлементы (например, молибден, марганец, медь) играют очень важную роль в жизни растений, так как входят в состав ферментов, катализирующих многие процессы обмена веществ. [c.9]
Например, в живых организмах из веществ относительно простого состава (вода, углекислый газ и сахар), в которых упорядоченность очень низка, образуются многие сложно построенные соединения (белки и т. д.), имеющие весьма высокую упорядоченность атомов. Такие процессы (например, синтез белков) идут с уменьшением энтропии. Зато одновременно другие вещества (молекулы сахара, жиры и т. д.) в результате окисления распадаются на более простые соединения (в конечном счете на углекислый газ и воду), а при этом снова происходит значительный рост энтропии. Конечно, в растениях из углекислого газа и воды в процессе фотосинтеза образуются также сахар и другие углеводы, но этот процесс идет не изолированно, а с поглощением энергии излучения Солнца. [c.126]
В результате работ И. П. Бородина выяснилось, что синтез аспарагина протекает в растениях с большой скоростью в условиях недостатка углеводов, когда происходит интенсивный окислительный распад белков. Д. Н. Прянишников выполнил очень важные исследования по обмену амидов (аспарагина и глютамина) в растениях. В исследованиях Д. Н. Прянишникова было показано, что синтез аспарагина и глютамина в растениях является процессом, аналогичным синтезу мочевины в животном мире. В обоих случаях достигается обезвреживание аммиака, но при этом все же имеется существенная разница. Мочевина является неактивным веществом в отношении дальнейшего участия в процессах обмена она выводится из организма без изменений, являясь типичным примером конечного продукта обмена. Аспарагин же и глютамин способны к дальнейшим превращениям и могут вовлекаться в процессы синтеза белка и других азотистых соединений (стр. 378). В. Л. Кретович показал, что в обмене аспарагина и глютамина в растениях имеется существенное различие. [c.375]
Фосфорорганические соединения, так же как и углеводы, в обработанных дефолиантами листьях распадаются на более подвижные соединения и интенсивно оттекают в другие органы растений. Это говорит о том, что дефолианты сильно нарушают ход физиологических и биохимических процессов в листьях растений, в частности ослабляют процесс синтеза органических веществ и усиливают процесс распада. Такой характер обмена веществ способствует образованию отделительного слоя и опадению листа. При-этом отделительный слой образуется вследствие деления и обособления клеток указанной зоны. [c.274]
Одновременно с распадом органического азотсодержащего вещества в почве происходят и вторичные процессы синтеза, когда минеральные соединения азота вновь переходят в органические, не усвояемые для растений. Эти процессы носят биологический характер. Микроорганизмы строят белок своих тел, используя углеводы и азот. Различные бактерии, актиномицеты и плесневые грибы разлагают клетчатку, пентозаны и другие вещества растительных и животных остатков. В качестве азотной пищи им необходимы в первую очередь легкорастворимые соединения азота почвы, которые они переводят в белок плазмы клеток. [c.181]
Распад нуклеиновых кислот под влиянием нуклеаз происходит и в животном, и в растительном организмах. Образовавшиеся пуриновые и пиримидиновые основания, углевод и фосфорная кислота подверга.ются в организме дальнейшим превращениям. Рибоза и дезоксирибоза могут окисляться до углекислоты и воды. Пуриновые и пиримидиновые основания превращаются в конечные продукты обмена, которые у животных выделяются из организма, а у растений могут быть использованы для синтеза других веществ. [c.372]
Одним из методов синтеза меченых соединений является биосинтез. Биосинтез широко используется при введении метки в сложные органические вещества природного происхождения (белки, углеводы и т. п.). Простейшим примером является получение меченой глюкозы и других углеводов в процессе фотосинтеза. Освещенные зеленые листья живого растения по мещают в атмосферу меченной по углероду (С) углекислоты. Затем сахар экстрагируют из растений и очищают, используя обычные химические и биохимические процедуры. [c.176]
Превращения углеводов, жиров и белков, их распад и синтез в организме теснейшим образом связаны друг с другом. Нельзя представить себе изолированно превращение отдельных органических, а также и неорганических веществ в организме. Только как исключение можно наблюдать преимущественный синтез углеводов (у зеленых растений на свету), распад углеводов с образованием этилового спирта и углекислого газа (в дрожжевых клетках при спиртовом брожении) и молочной кислоты (при работе мышц), синтез жиров (при откорме животных), синтез белков (при усиленном росте). Но даже и в этих случаях обмен веществ не сводится к превра[це-пиям т0JПзK0 одной какой-либо группы веществ. Обмен веществ между любым живым организмом п окружающей его средой является чрезвычайно слюж-ным процессом, в который вовлекаются химические составные части организма и вещества, поступающие в пего извне (пищевые вещества, включая кпс лород и воду). Обмен веществ у человека и животных регулируется централыюй нервной системой. При изучении превращений углеводов, жиров и белков приводились данные о регуляторной деятельности центральной нервной системы. Было бы ошибочным полагать суп.1,ествование в центральной нервной системе отдельных механизмов, регулирующих превращения отдельных групп веществ. Процесс обмена веществ между организмом и внешней средой, лежащей в основе проявления жизни,— единый биологический процесс и если его расчленяют на процессы превращения отдельных веществ, то это делают только с целью более глубокого его познания и изучения. [c.459]
Если представить себе работу живой клетки в целом, то можно сказать, что клетки организмов животных, растений и даже одноклеточные микроорганизмы представляют собой поразительные по своему совершенству химические заводы. В них вырабатываются, например, сложнейшие полимерные частицы с самыми разнообразными свойствами, различного состава, разной прочности, эластичности, различной термоустойчивости и окраски. В клетках очень постоянно, одновременно, или, если это нужно, то в необходимой последовательности, происходят тонкие синтезы таких сложных и разнохарактерных веществ, как нуклеиновые кислоты, белки и в том числе ферменты, сложные углеводы, жиры, витамины, гормоны и множество других сложных соединений. Эти заводы в клетках смонтированы очень компактно, все процессы в них точно и четко регулируются, режим оптимальный. В основе работы их лежит согласованное действие ферментных систем, осуществляющих цепи управляемых химических реакций. Естественно, что в будущем подобные принципы работы должны быть использованы и химической, и биохимической промышленностью. [c.337]
Промышленный органический синтез (получение топлива, растворителей, высокополимеров и т. д.) основывается на минеральном углеродистом сырье нефти, каменном угле и природных газах. Кроме того, многие органические веш,ества могут быть получены из природного сырья, т. е. из органических соединений, синтезируемых растениями и животными. Такие соединения являются представителями трех основных групп природных веществ углеводов, жиров и белков. Одни из них, а именно углеводы, весьма широко используются в химической и пищевой промышленности (переработка древесины различными способами, добывание и переработка крахмалистых и сахаристых продуктов), другие — жиры — занимают несравненно меньшее, но все же значительное место в промышленности (главным образом пищевой и мыловаренной), третьи — белки — широко используются в пищевой промышленности и в незначительной степени в других производствах (некоторые виды пластмасс, кожевенное производство). [c.5]
Растение и животное в создании органического вещества. Основная задача сельскохозяйственного производства— получение органических веществ растительного и животного происхождения углеводов, белков, жиров и других органических соединений. Синтез органического вещества осуществляется в природных условиях в организме растений и животных. [c.20]
Медь участвует в углеводном и белковом обмене в растениях. Под влиянием меди повышается как активность пероксидазы, так и синтез белков, углеводов и жиров. Недостаток меди вызывает у растений понижение активности синтетических процессов и ведет к накоплению растворимых углеводов, аминокислот и других продуктов распада сложных органических веществ. [c.21]
Уничтожая главные составные части растений — лигнин, целлюлозу и другие углеводы, сапрофиты синтезируют вещества другой природы — белки, жиры, хитин. Поэтому вопрос об исходном материале горючих ископаемых не так прост, как это казалось лет 20 тому назад. Горючие ископаемые образуются не непосредственно из растительных остатков и не из продуктов их химического распада, а из продуктов сложного биохимического синтеза, осуществляемого микробами. Характер продуктов этого синтеза зависит не столько от химического состава первичного растительного материала, сколько от условий, в которые попадает этот материал после его отмирания. [c.372]
При недостатке или отсутствии углеводов для дыхания используются другие органические соединения, обладающие необходи-мым. запасом энергии. Однако в этом случае синтез всех других веществ будет в значительной степени нарушен, а рост и развитие растений будут подавлены. [c.12]
Продукты распада сине-зеленых водорослей. Сине-зеленые водоросли относятся к группе низших, наиболее примитивных растений. В большинстве случаев это одноклеточные организмы, обычно соединяющиеся в кс-лонии. У некоторых клетки при помощи слизи и выростов соединены в ценобии в виде нитей, давая внешнюю картину многоклеточности (рис. 9.1). Размножаются они преимущественно путем деления клеток. Живут сине-зеленые водоросли не только в воде, но и на суше (на берегах водоемов, в почвах и на их поверхности). Это самые распространенные растения земного шара. Они первыми заселяют бесструктурные почвы и совместно с бактериями подготавливают их для освоения другими растениями. Эти водоросли вообще аэробные организмы. Они способны к синтезу углеводов, но используют и распадающиеся органические вещества. [c.253]
Переаминирование имеет большое значение для синтеза белков, а также для дезаминирования аминокислот. Дезаминирование — это отщепление аминогруппы от аминокислоты, в результате образуются аммиак и кетокис-лота. Кетокислота используется растением для переработки в углеводы, жиры и другие вещества аммиак же вступает в реакцию прямого аминиро-вания кетокислот, возникающих из углеводов, и дает аминокислоты. Кроме [c.184]
Все эмпирически известное по поводу энзимов в клетках растений не имеет непосредственного отношения к этим гипотетическим катализаторам. Данные, имеющиеся в нашем распоряжении, касаются хорошо известных энзимов, вроде каталазы, карбоангидразы, фос-форилазы, амилазы, мальтазы и инвертазы, которые или вообще не имеют отношения к синтезу углеводов, или участвуют лишь в его заключительных стадиях (образование и разлоягение сахарозы и крахмала). О наличии энзимов, превращающих углеводы в зеленых листьях, кратко говорилось в главе П1. Здесь мы добавим немногие данные по другим энзималг, найденным в выделенном веществе хлоропластов [97, 98, 105]. [c.381]
Во время дыхания растительный организм, постепенно расщепляя молекулу глюкозы, извлекает ее энергию для накоплепия во вновь образующихся фосфатйых связях АТФ. Молекулы ад но-зинтрифосфата (АТФ) переносят полученную за счет питательных веществ или солнечного света свободную энергию от центров дыхания (митохондрий) или фотосинтеза (пластид) во все участки клетки, обеспечивая прохождение всех процессов, связанных с поглощением энергии. Углеводы при дыхании окисляются не сразу до углекислого газа и воды, а постепенно, образуя десятки промежуточных соединений, которые растения используют для синтеза всех необходимых им других органических соединений. Получающиеся при дыхании органические кислоты, спирты и другие соединения служат, как и при фотосинтезе, для образования белков, жиров и других веществ. Суммарно и схематически процесс дыхания растений можно представить уравнением, как бы обратным уравнению фотосинтеза [c.11]
Переаминирование имеет большое значение для синтеза белков, а также для дезаминирования аминокислот. Дезаминирование — это отщепление аминогруппы от аминокислоты, в результате образуются аммиак и кетокис-лота. Кетокислота используется растением для переработки в углеводы, жиры и другие вещества аммиак же вступает в реакцию прямого аминирова-ния кетокислот, возникающих из углеводов, и дает аминокислоты. Кроме того, аммиак реагирует с аспарагиновой и глютаминово кислотами, способными связать еще но одной его молекуле, давая таким образом амиды амино дикарбоновых кислот [c.173]
Фотохимические (фотокаталитически е) реакции происходят в природе и сравнительно давно используются промышленностью. Фотохимическими называют реакции, вызываемые и ускоряемые действием света их элементарный механизм состоит в активации молекул при поглощении фотонов. Большинство промышленных фотохимических реакций происходит по цепному механизму, т. е. молекулы, поглотившие фотон, диссоциируют, и активированные атомы или группы атомов служат инициаторами вторичных реакций. По такому типу протекают галогенирование углеводородов и других веществ, синтез полистирола, сульфохлорирование парафинов и т. п. Природный фотосинтез требует непрерывного подвсда световой энергии синтез углеводов из диоксида углерода воздуха совершается под действием солнечного света, поглощаемого пигментом растений хлорофиллом (аналог гемоглобина крови). При фотокаталитических реакциях фотоны поглощаются не реагентами, а катализаторами, ускоряющими химическую реакцию, т. е. реакция ускоряется в результате суммарного действия катализатора и световой энергии. [c.150]
ФОТОСИНТЕЗ — синтез растениями органических веществ (углеводов, белков, жиров) из диоксида углерода, воды, азота, ( юсфора, минеральных солей и других компонентов с помощью солнечной энергии, поглощаемой пигментом хлорофиллом. Ф.— основной процесс образования органических веществ на Земле, определяющий круговорот углерода, кислорода и других элементов, а также основной механизм трансформации солнечной энергии на нашей планете. В процессе Ф, растения усваивают вгод4 101 туглерода, разлагают 1,2 х X 10 т воды, выделяют 1 10 т кислорода и запасают 4-102° кал солнечной энергии в виде химической энергии продуктов Ф. Это количество энергии намного превышает годовую потребность человечества в ней. Ф.—сложный окис-лительно-восстановительный процесс, сочетающий фотохимические реакции с ферментативными. Вследствие Ф. происходит окисление воды с выделением молекулярного кислорода и восстановление диоксида углерода, что выражается [c.268]
В организмах человека и животных происходят процессы синтеза различных органических веществ. Следует, однако, отметить, что процессы синтеза в этих организмах не столь разнообразны, как в зеленых растениях, и известным образом ограничены. Следует подчеркнуть, что животные организмы неспособны использовать энергию солнечных лучей для синтеза органических соединений. Из этого отнюдь не вытекает, что в организме животных не используется для синтеза углекислый газ, вода и аммиак. Уже с давних пор известно, что выделяющаяся нз организмов человека, млекопитающих животных, амфибий и рыб мочевина синтезируется из углекислого газа, воды и аммиака применение метода меченых молекул позволило выявить участие воды, углекислого газа и аммиака в процессах синтеза сложных органических веществ — составных частей организма. После введения в организм животных тяжелой воды можно обнаружить тяжелый водород (дейтерий) в составе жиров, углеводов, белков и других веществ организма. Введение в организм животных карбонатов, меченных i позволяет проследить, как различные органические вещества приобретают радиоактивную метку, благодаря включению в их состав углерода углекислого газа. После введения в организм животных аммонийных солей, меченных стабильным изотопом азота (N ), появляется в составе белков и других азотистых веществ. Все эти данные с несомненностью показывают, что в организме животных для синтеза органических веществ используются минеральные вещества — аммиак, вода и углекислый газ. Было бы, однако, ошибочным считать, что в организмах животных и в зеленых растениях отсутствуют различия в использовании минеральных веществ для синтетических целей. Различия эти прежде всего количественного характера. Объем использования углекислого газа, воды и аммиака для синтеза органических веществ в организмах животных, но сравнению с организмами зеленых растений, незначителен. Далее обращают на себя внимание и различия качественного характера-, ряд веществ, синтезирующихся в растениях, вовсе не синтезируются в организмах человека и животных, и эти вещества должны доставляться человеку и животным в готовом виде с продуктами питания. Так, оргагшзмы человека и животных не способны к синтезу ряда аминокислот, входящих в состав белков, не могут синтезировать различные витамины и т. д. Отсутствие этих веществ в пище приводит к их гибели. [c.235]
Известно, что при фотосинтезе и минеральном питании растений, этих двух взаимосвязанных автотрофных процессах, происходят поглощение и ассимиляция органогенов С, О, Н, Ы, Р, 5 и других элементов (металлов и неметаллов) и синтез углеводов, белков, липидов и веществ вторичного происхождения (органические кислоты, гликозиды, фенольные соединения, алкалоиды, каучук и др.), т, е. синтез органических веществ. Установленные закономерности н корреляционные зависимости между световым и углеродным питанием (фотосинтез), транспирацией, дыханием, минеральным питанием и урож айность10 составляют основу теории объединения воздушного и минерального питания растений как интегрального процесса. Поэтому для протекания в растениях эффективного продукционного процесса необходимы следующие условия [c.350]
Для исследования процессов образования полисахаридов большое значение имеет использование меченых соединений, например Oq, меченых сахаров и их производных. Предполагается, что биосинтезирующая система растений не различает соединений, содержащих С и С, и в обоих случаях реакции протекают одинаково, Включение СОз в состав воздуха, окружающего растущее растение, и анализ образцов, собранных через определенное время после начала эксперимента, позволили сделать ряд интересных выводов. За очень короткие промежутки времени в процессе фотосинтеза образуется большое количество меченых соединений. Тем не менее у большинства растений основная масса С уже в первые минуты концентрируется в углеводах. Когда в питательную среду вводится радиоактивная глюкоза, радиоактивность прежде всего обнаруживается в полисахаридах и только потом — в иротеинах, липидах и других полимерах. Если учесть, что глюкоза и ее соединения являются исходным веществом для синтеза большинства биополимеров, то можно полагать, что путь иревра- [c.24]
Заканчивая рассмотрение аминокислотного обмена, следует сказать, что обычно в растениях в свободном состоянии содержится 20—30 различных аминокислот, которые подвергаются непрерывным превращениям используются для синтеза белков, нуклеиновых кислот, алкалоидов и других азотистых веществ, превращаются в безазотистые соединения — органические кислоты, углеводы, жиры. Содержание аминокислот в растениях может резко меняться в зависимости от возраста растений, от ряда внешних условий (температуры, длины дня, увлажнения и т. д.), а также от питания. При этом изменяется ке только концентрация, но и качественный состав аминокислот. Различные внешние воздействия, нарушая течение азотного обмена, часто направляют его по другим путям, что приводит к уменьшению или даже к исчезновению ряда аминокислот, характерных для данного растения, или, наоборот, к повышенпю общего содержания аминокислот, или появлению ряда нехарактерных продуктов азотного обмена. При обычных условиях выращивания количество свободных аминокислот с возрастом растений понижается. В вегетативных органах растений свободных аминокислот обычно больше, чем в репродуктивных, в то время как для белков наблюдается обратная зависимость. При различных условиях минерального питания содержание индивидуальных аминокислот в растениях и соотношение между ими могут быть резко различными. Увеличение общего количества свободных аминокислот в растениях и усиленное накопление отдельных аминокислот наблюдается при пониженном питании растений калием, фосфором, серой, кальцием и магнием, а также при недостатке ряда микроэлементов цинка, меди, марганца, железа. Увеличение содержания аминокислот наблюдалось также при лучших условиях азотного питания. При недостатке молибдена количество свободных аминокислот и амидов в растениях уменьшалось вследствие ослабления восстановления нитратов. В настоящее время проводятся широкие исследования [c.264]
Одновременно с минерализацией органического азотсодержащего вещества в почве происходят и вторичные процессы синтеза, когда минеральные соединения азота вновь переходят в органические, не усвояемые для растений. Эти процессы носят биологический характер. Микроорганизмы строят белок своих тел, используя углеводы и азот. Различные бактерии, лктиномицеты и плесневые грибы разлагают клетчатку, пентозаны и другие [c.192]
При. попытке выделения продуктов метаболизма 2,4-Д в растениях обнаружено вещество кислого характера, извлекаемое эфиром [936], в которое пе >еходит до 60% радиоактивного углерода. Предполагали, что это может быть один из изомеров окси-2,4-Д [936]. Синтез и изучение некоторых таких соединений показали, что они физиологически неактивны [937, 938]. Обработка растений фасоли малыми дозами 2,4-Д, меченной С , показала, что в результате метаболизма 2,4-Д превращаег-ся в весьма разл1ичные продукты, так как С обнаружен в белках, углеводах, кислотах и других соединениях [939, 940]. [c.375]
В настоящее время следовало бы попытаться произвести синтез более сложных азотистых соединений, исходя из формамида и формальдегида или других углеводов. Получаясь при соединении двух характерных групп СНО и Nh3 и являясь типичным альдегидамидом, формамид должен обладать способностью вступать в соединения посредством либо одной, либо другой из своих групп или посредством обеих. Если бы можно было достигнуть таким образом синтеза аспарагина или аналогичного вещества, то вопрос о синтезе белковых веществ в растениях существенно продвинулся бы. Я сделал в этом направлении некоторые опыты, которые до настоящего времени не дали удовлетворительных результатов я предполагаю продолжить эти исследования. [c.11]
chem21.info
Помогите решить тест по биологии (заранее спасибо)
1.Определение гомеостаз характеризует:А. процесс разрушения клеток путем их растворения Б. состояние динамического равновесия клетки, обеспеченное деятельностью регуляторных системВ. процесс расщепления углеводов в отсутствии кислорода Г. общее снижение жизнеспособности организма2. Метаболизм заключается в протекании двух взаимосвя¬занных и противоположно направленных процессов:А. возбуждения и торможения Б. жизни и смертиВ. синтеза и расщепления органических веществ Г. поглощения кислорода и выделения углекислого газа3.Генетический код един для всех существ, обитающих на Земле, и представляет собой:А. систему «записи» Б. способностьнаследственной информации воспроизводить себе в молекулах ДНК подобныхВ. процесс образования живыми организмами органических молекул из неорганических веществ Г. доклеточные образования, обладающие некоторыми свойствами клетки
4. Для какой фазы важнейшего процесса, осуществляемого зелеными клетками растений, характерно образование углеводов в результате последовательного превращения воды и углекислого газа?А. фазытрансляции Б. фазы кислородного этапа обмена
В. темновой Г. световой5. Процессы анаэробного окисления глюкозы протекают в: А. ядре Б. пластидахВ. цитоплазме Г. митохондриях.6. При каком из процессов выделяется наибольшее коли¬чество энергии?А. гликолиз Б. клеточное дыханиеВ. испарение Г. фотолиз7. Какой газ является побочным продуктом реакции фотолиза, происходящего в зеленых клетках растений?А. водород Б. азотВ. углекислый газ Г. кислород8. К какому процессу жизнедеятельности клетки относит¬ся распад рибосомы на субъединицы с высвобождением энергии информационной РНК, а также с синтезом пептидов? А .Третьему этапу Б. завершению синтеза белка энергетического обмена В. транскрипции Г. реакции матричного синтеза
9.Между атомами каких элементов создается пептидная связь в процессе формирования первичной структуры белка при его синтезе?А. углерод — углерод Б. углерод — кислород —углеродВ. углерод — азот Г. азот — азот10. На мембранах этого органоида осуществляется синтез жиров и углеводов, благодаря его функционированию происхо¬дит обновление и рост плазматической мембраны, он называется:А. гладкая эндоплазматическая сеть Б. аппарат Гольджи В. шероховатая эндоплазматическая сеть Г. клеточный центр
otvet.mail.ru
Клетки высших растений - Справочник химика 21
Подобно бактериям, клетки высших растений и животных часто покрыты внеклеточным материалом. Так, растительные клетки имеют жесткую стенку, содержащую в большом количестве целлюлозу и другие полимерные углеводы. Клетки, расположенные на наружных поверхностях растений, бывают покрыты восковым слоем. Клетки животных снаружи обычно защищены гликопротеидами — комплексами углеводов со специфическими белками клеточной поверхности. Пространство между клетками заполнено такими цементирующими веществами , как пектины у растений и гиалуроновая кислота у животных. Нерастворимые белки —коллаген и эластин — секретируются клетками соединительной ткани. Клетки, лежащие на поверхности (эпителиальные или эндотелиальные), нередко граничат с другой стороны с тонкой, содержащей коллаген базальной мембраной (рис. 1-3). Часто в результате совместного действия клеток различного типа происходит отложение неорганических соединений — фосфата кальция (в костях), карбоната кальция (скорлупа яиц и спикулы губок), окиси кремния (раковины Диатомовых водорослей) и т. п. Таким образом, обмен веществ в значительной мере протекает вне клеток. [c.37] Ориентированное движение цитоплазмы, наблюдаемое в клетках высших растений, скорее всего связано с функцией актина. Гораздо более полно это явление изучено на примере гигантских клеток некоторых водорослей. [c.194]Сине-зеленые водоросли, фотосинтезирующие бактерии, зеленые клетки высших растений [c.460]
Эукариотические клетки высших растений (рис. 2-21) несколько отличаются от клеток высших животных, несмотря на сходство их основных особенностей. Пожалуй, наиболее явное различие состоит в том, что большинство растительных клеток содержит пластиды. Пластиды-это расположенные в цитоплазме специализированные органеллы, окруженные двумя мембранами. К самым типичным пластидам, характерным для всех клеток зеленых растений, относятся хлоропласты (рис. 2-22). Подобно митохондриям хлоропласты можно рассматри- [c.46]
В фотосинтезирующих клетках высших растений всегда присутствуют хлорофиллы двух типов. Один из них-это хлорофилл а, а второй представлен у многих видов хлорофиллом Ь, отличающимся от хлорофилла а тем, что вместо метильной группы при кольце II в нем содержится альдегидная группа (рис. 23-8). Хлорофиллы аиЬ могут быть выделены в чистом виде из экстрактов листьев хроматографическими методами. Хотя оба они окрашены в зеленый цвет, но их спектры поглощения слегка различаются. У большей части высших растений количество хлорофилла а примерно вдвое превышает количество хлорофилла Ь. [c.691]
Изучение лизогенных бактерий позволило по-новому подойти к проблеме происхождения и природы разного типа вирусов. Хотя некоторые исследователи рассматривают вирусы как химические соединения и проявления их жизнедеятельности считают исходящими от клеток хозяина, чрезвычайно интересно, что эти вирусы имеют особый генотип — специфическое строение ДНК, Как показывает изучение состава ДНК вируса, она очень похожа на ДНК бактерии-хозяина и может быть присоединена в определенном месте к хромосоме бактерии-хозяина. Изучение лизогенных бактерий ставит также вопрос о том, не присутствуют ли такие скрытые формы вирусов также и в клетках высших растений и животных, где они при особых обстоятельствах переходят Б активное состояние и оказывают вредное действие. Относительно этого нет достоверных сведений, однако можно утверждать, что в течение многих поколений некоторые растительные вирусы могут сохраняться в организме определенных видов насекомых, которые передают этот вирус растениям, когда сосут сок из их зеленых частей. Таким образом, этот вирус присутствует в организме насекомого, внешне ничем не проявляясь, [c.254]
В клетках высших растений может быть до нескольких сотен хлоропластов. Они постоянно находятся в движении и реагируют на условия освещенности. Общая поверхность хлоропластов в сотни раз больше поверхности листьев. [c.12]
Значительная часть энергии, заключенной в углеводородной цепи с высокой степенью насыщения, посредством реакций р-окисления аккумулируется в тиоэфирной связи ацетил-КоА. В клетках высших растений ацетил-КоА может затем включаться в реакции цикла трикарбоновых кислот с последующим выделением запасенной энергии. Кроме того, ацетил-КоА принимает участие в глиоксилатном цикле. Ацетил-КоА участвует и во многих других реакциях, которые здесь не указаны. [c.185]
Другая важная закономерность переноса ионов состоит в том, что клетки высших растений, активно накапливающие ионы, характеризуются очень низкой проницаемостью мембран для свободных ионов [5, 27]. При перенесении тканей высших растений в воду не происходит большой потери ионов при условии, что в воде имеется в незначительной концентрации кальций. Даже изотопный обмен происходит во много раз медленнее, чем поглощение ионов клетками, что было достоверно установлено в опытах, в которых ткани, меченные радиоактивными изотопами, переносили в раствор, содержащий [c.261]
Помимо основных, важнейших метаболитов, присутствующих во всех живых клетках, высшие растения содержат значительные количества вторичных метаболитов, таких, как фенилпропановые соединения, описанные нами в этой главе, а также флавоноиды и другие пигменты, терпены, алкалоиды и т. д. Френкель [19] рассмотрел некоторые гипотезы о причинах существования этих вторичных соединений, многие из которых характерны только для ограниченного числа видов. Полагают, что новые метаболиты возникали в ходе эволюции в результате случайных мутаций. Если эти соединения имели хотя бы небольшое значение для выживания мутанта — пусть это было даже совсем незначительное преимущество,— то этот мутант постепенно вытеснял родительский штамм из его экологической ниши или даже распространялся в другие ниши. Новые метаболиты могут повышать выживаемость растений, защищая их от грибов, бактерий или насекомых или делая их несъедобными для животных. Другие вещества могут способствовать опылению, привлекая к цветкам насекомых. Кроме того, могут возникать вещества, которые повышают устойчивость растений к засухе или морозу. Преимущества, обусловленные одревеснением, [c.370]
Фото 1. Эксперимент, указывающий на полуконсервативный характер репликации ДНК в экспоненциально растущих клетках высших растений. [c.625]
Безусловно, наша попытка проникнуть в клетку при помощи электронного микроскопа не даст нам никаких знаний относительно процесса фотосинтеза, но, быть может, позволит нам кое-что узнать о тонкой структуре хлоропластов. Однако начнем мы со светового микроскопа. Ведь хлоропласты достаточно велики, и их очень удобно наблюдать в световой микроскоп. В клетках высших растений хлоропласты в общем однотипны по форме, напоминая диски или линзы диаметром чаще всего примерно 5 микрон. У некоторых видов мхов в клетках присутствует только по одному хлоропласту. Однако, как правило, в распоряжении ассимилирующей клетки их имеется 10—20, а иногда и больше. [c.244]
Гуляев Б. А Литвин Ф. Ф. К вопросу о единой системе агрегированных (полимерных) форм фотосинтетических пигментов в клетках высших растений, водорослей и бактерий.— Биофизика, т. 1 № 5. [c.226]
Уже отмечалось, что внедрение в клетки высшего растения гиф паразита, установление контакта между выделяемыми им физиологически активными веществами и протоплазмой хозяина вызывает у последнего ряд ответных реакций, совокупность которых и составляет сущность того, что известно под названием иммунитет, либо, наоборот, восприимчивость, или поражаемость. [c.648]
| Рис. 13-71. Ход цитокинеза в клетках высших растений, имеюш,их жесткую клеточную стенку. |  |
В клетках высших растений помимо хлоропластов содержатся еще и другие типы пластид, лишенные характерных для хлоропластов ламеллярной структуры и фотосинтетического аппарата. Среди этих пластид различают бесцветные лейкопласты и окрашенные хромопласты, окраска которых определяется высокой концентрацией каротиноидных пигментов. Подобно хлоропластам, эти тельца, по-видимому, передаются по наследству через какие-то структуры типа пропластид, присутствующие в материнской цитоплазме. Лейкопласты играют в клетке роль [c.57]
Подобно всем прочим эукариотическим клеткам, клетки высших растений содержат окруженное оболочкой ядро, эндоплазматический ретикулум, диктиосомы и митохондрии. Рибосомы встречаются в них как в свободном виде — в цитоплазме, так и прикрепленными к эндоплазматическому ретикулуму кроме того, рибосомы обнаруживаются в некоторых клеточных органеллах. Репликация ДНК, ДНК-зависимый синтез РНК и РНК-за-висимый синтез белка протекают в растительных клетках так же, как и в других клетках. Многие растительные белки обладают каталитической активностью, т. е. являются ферментами другие белки выступают как важные структурные компоненты клетки. [c.76]
После этого гексоза из цикла может направляться либо на синтез сахарозы и полисахаридов, либо — через дыхательный-путь (см. гл. 5) — на построение углеродных скелетов любых других органических соединений клетки. Таким образом, сахар,, образующийся в процессе фотосинтеза из СОг, — это основное органическое вещество, которое в клетках высших растений служит источником как энергии, так и необходимых клетке строительных блоков. [c.127]
Типичные величины трансмембранного потенциала и концентрации К+, Na+ и С1- внутри вакуоли клетки высшего растения показаны ниже. Концентрация Na+ внутри клетки меньше величины, рассчитанной по уравнению Нернста, а концентрация [c.225]
Фотосинтез (Photosyntesis) Процесс превращения клетками высших растений энергии видимого света в энергию химических связей, сопровождаемый образованием органических соединений и кислорода из диоксида углерода и воды. [c.563]
Уилки [80] в 1983 г. предложил называть гемицеллюлозами полисахариды, находящиеся в одревесневших клетках высших растений и образующие в них гель, в котором размещаются фибриллы целлюлозы. ГМЦ растворяются в растворах щелочей и гораздо легче гидролизуются минеральными кислотами, чем целлюлоза, образуя при этом -арабинозу, 0-ксилозу, 0-маннозу, в меньшем количестве — О- и -галактозу, Д-глюкозу, О-рамнозу, О-глюку-роновую, 4-0-метил-0-глюкуроиовую и Д-галактуроиовую кислоты и некоторые другие сахара в небольшом количестве. Как отмечает Уилки, такое определение охватывает также пектиновые вещества, характерными соединениями среди которых являются га-лактуроновая кислота, арабиноза и галактоза. В то же время оно исключает полисахариды с иными химическими свойствами, содержащиеся в клеточной оболочке в самые начальные периоды ее развития, когда еще не начиналась лигнификация клетки. Исключены в определении Уилки также полисахариды, сопровождающие при выделении трудногидролизуемую часть полисахаридов — целлюлозу, но при полном гидролизе образующие ксилозу и маннозу. [c.16]
Клетки высших растений Высшие растения (порядка 300 ООО видов) — это дифференцированные многоклеточные, преимущественно наземные организмы Способы их бесполого и полового размножения хорошо описаны в учебниках ботаники В процессе дифференциации и специализации клетки растений группировались в ткани (простые — из однотипных клеток, и сложные — из разных типов клеток) Ткани, в зависимости от функции, подразделяют на образовательные, или меристемные (от греч menstos — делимый), покровные, проводящие, механические, основные, секреторные (выделительные) Из всех тканей лишь меристематические способны к делению и за их счет образуются все другие ткани Это важно для получения клеток, которые затем должны быть включены в биотехнологический процесс (см специальную часть). [c.37]
Клетки трибов и водорослей по своей организации похожи на клетки высших растений. Основными частями клетки являются оболочка, протоплазма (цитоплазма) и ядро (нуклеус). В состав оболочки входит целлюлоза. Протоплазма представляет собой сложное коллоидное образование с резко выраженным поверхностным натяжением. В этой коллоидной системе непрерывной фазой является вода, а дисперсной фазой — липопротеиновые соединения. В протоплазме одноклеточных грибных организмов — дрожжей— легко обнаруживаются вакуоли, представляющие собой пустоты, заполненные клеточным соком. При делении вакуоли дочерней клетки образуются путем отпочковы-вания от вакуоли материнской клетки. В протоплазме имеются также мельчайшие гранулы-—рибосомы (микросомы), размеры которых составляют 200 ммк, обнаружить их можно лишь методом электронной микроскопии. Б рибосомах, состоящих из рибонуклеиновой кислоты и белка, происходит белковый синтез- [c.113]
Для ирижизненно-] о флуорохромирования митохондрий у микроорганизмов и сферозом в клетках высших растений для выявления ядер и нервных волокон на фиксированных препаратах [c.372]
У растений фотосинтез происходит внутри высокоспециализированных органелл, называемых пластидами. Если клетки разрушить и выделить из них чистую фракцию пластид, то оказывается, что пластиды в присутствии соответствующих субстратов и кофакторов способны осуществлять все реакции фотосинтеза. В зеленых растениях, в том числе и в зеленых одноклеточных организмах, например у водорослей hlorella и S enedesmus, а также у простейшего Euglena, пластиды содержат хлорофилл и называются хлоропластами. Число, размеры и форма хлоропластов у разных организмов различны. В клетках хлореллы, например, имеется только одна пластида чашеобразной формы большинство же других водорослей, а также зеленые клетки высших растений содержат много хлоропластов. [c.315]
Другим важным аспектом биологии хлоропластов является их самовоспроизведение. Существуют, но-видимому, два способа воспроизведения хлоропластов. Первый из пих состоит в делении зрелых хлоропластов, а второй — в делении предшественников хлоропластов — пропластид, из которых возникают зрелые хлоропласты. Деление зрелых хлоропластов обычно имеет место у многих водорослей, например у хлореллы (фото 25), а также у таких организмов, как гаметофиты папоротников, стоящих на весьма высокой ступени эволюционного развития. В то же время о делении зрелых пластид у высших растений имеется мало данных. В клетках высших растений пластиды обычно возникают из телец, называемых пропластидами, которые подвергаются делению (делящаяся пронластида представлена на фото 26). Развитие хлоропластов из пропластид является одним из чрезвычайно интересных аспектов биохимии хлоропластов. В пропластиде, как правило, присутствует очень мало (или вообще не имеется) внутренних хлорофиллсодержащих ламелл. В нормальных условиях эти ламеллы возникают нутем впячивания внутренней части двойной мембраны, окружающей пластиду [22]. Этот процесс отчетливо виден в пластидах Ni otiana (фото 26). При других условиях внутренняя мембрана может давать начало пузырькам, из которых в дальнейшем образуются ламеллы. [c.83]
Во многих отношениях хлоропласт проявляет свойства независимого организма, зара-зивнгего клетку высших растений. К этим свойствам относятся наличие в хлоропластах специфической ДНК и РНК, способность к синтезу белка и способность к самовоспроизведению путем деления. Такая автономность хлоропластов позволила генетику Ледербергу заявить, что клетки, обесцвеченные стрептомицином, излечиваются от своих хлоропластов . Возможно, дальнейшие исследования хлоропластов, пусть даже по-прежнему касающиеся наиболее удивительного их свойства — фотосинтеза, еще более подчеркнут биологическую независимость хлоропластов в растительной клетке. [c.83]
Лелуар и его сотрудники показали, что переносчиком глюкозильных групп в реакции биосинтеза гликогена у высших животных служит уридинфосфат, а в клетках высших растений и микроорганизмов эту функцию выполняет АДФ, ГДФ или ЦДФ. На второй стадии синтеза гликогена в печени, катализируемой глюкозо-1-фоофат-уридилтрансферазой, образуется УДФ-глюкоза. [c.175]
Необходимая для жизнедеятельности организмов энергия в условиях отсутствия доступа кислорода получается одноклеточными организмами и частично клетками высших растений в процессах так называемых брожений. При брол-сении в ряде следующих друг за другом реакций происходит распад углеводов, конечными продуктами которых обычно оказываются вещества, содержащие еще значительные количества энергии. Важнейшими типами чистых форм брожений являются спиртовое, молочнокислое и маслянокислое. Лучше других изучено спиртовое брожение. [c.384]
Внутри ядра имеются длинные образованпя — хроматиновые нити, с трудом видимые в покоящейся клетке, т. е. существующей в период между двумя делениями. Перед делением клетки хроматиновые нити образуют палочкообразные тельца — хромосомы. Число и форма их постоянны для данного вида организмов. В соматических (не половых) клетках высших растений и животных хромосомы существуют попарно, т. е. но две копии каждого типа. Такая пара хромосом называется гомологичной, подобный пабор хромосом — диплоидным. Клетки человека содержат 23 нары хромосом (46). Из общего правила, что в клетках с диплоидным набором каждая хромосома представлена двумя копиями, имеется исключение. Оно заключается в том, что в клетках женских особей одна пара хромосом содержит две одинаковые Х-хромосомы, а в клетках мужских особой — одну X- и одну У-хромосому, отсутствующую в женских клетках. В половых клетках (гаметах) число хромосом вдвое меньше — по одной хромосоме от каждой пары. Этот набор хромосом называют гаплоидным. [c.11]
Способность использовать разнообразные питательные вещества свойственна факультативным паразитам. Особенно выражено это свойство у полусапрофитов, которые не в состоянии входить в симбиотические отношения с живыми клетками высшего растения и распространяются лишь по тканям, предварительно убитым токсическими выделениями гриба. Факультативные паразиты образуют большое количество разнообразных экстрацеллюлярных ферментов, расщепляющих полимерные пластические вещества растений и переводящих тем самым эти вещества в усвояемую форму. [c.30]
Можно думать, что энергия, используемая грибом для собственных синтетических процессов, заключена в макроэргических связях нестойких соединений, возникающих на первых этапах цепи переноса электронов, до звена, нарушаемого динитрофенолом. Указанием на различия в энергетическом обмене может служить то, что у шампиньона молекула фосфата присоединяется к неидентифи-цированному пока соединению, не сходному ни с одним из веществ, фосфорилируемых в клетках высших растений. [c.171]
Для организованного роста растения необходимо, чтобы клетки в специализированных участках делились в определенной плоскости и в определенное время, иными словами, чтобы цепочки клеток были правильно ориентированы. Микротрубочки с самого начала принимают участие в ориентации плоскости деления клеток. Таким образом, первые заметные признаки того, что клетка высшего растения собирается делиться в определенной плоскости, появляются сразу же после интерфазы, когда микротрубочки кортикального слоя исчезают в ходе подготовки к митозу. В это время непосредственно под плазматической мембраной появляется узкая окружающая всю клетку зона микротрубочек (рис. 20-63). Так как микротрубочки, входящие в состав этой зоны, появляются в Сг перед началом профазы, данная зона называется иреирофазиым пучком. Хотя она исчезает перед метафазой, но граница плоскости деления каким-то образом запоминается когда позже в процессе цитокинеза образуется новая клеточная пластинка, го она начинает расти наружу, сливаясь с родительской стенкой гочно в гом месте, которое первоначально было занято нрепрофазным пучком (рис. 20-64). Даже если содержимое клетки после исчезновения препрофазного пучка сместить посредством центрифугирования, растущая клеточная пла- [c.431]
Приобретение цианелл клетками высших растений никогда не наблюдалось. Видимо, они слишком дифференцирован-ны для этого, а быть может, в этом случае просто не удается отличить цианеллы от хлоропластов. Хлоропласты, по-видимому, могут неделями жить как симбиотические органеллы в клетках пищеварительного тракта слизней [963, 1846, 1872]. Эти животные имеют специальные механизмы для приема хлоропластов в клетку. Удается также искусственно вво- [c.189]
Полный двойной набор хромосом называют диплоидным (2 п), а набор, получаемый от каждого из родителей через половые клетки, — гаплоидным (п). Все клетки высшего растения, за исключением гаплоидных половых, как минимум диплоидны. Гаплоидные половые клетки находятся преимущественно в зрелых пыльцевых зернах и в зародышевом мешке семязачатка. В жизненном цикле растения гаплоидный набор" получается из диплоидного в результате редукционного деления, или мейоза (см. стр. 34), протекающего в материнских клетках микро- и мегаспор, находящихся соответственно в пыльниках и семязачатке цветка. Возникшие таким путем гаплоидные клетки де лятся и дают начало мужским и женским гаметофитам, в которых в конце концов и образуются половые клетки, или гаметы, т. е. спермии и яйцеклетки. Когда — при половом размножении— женские и мужские гаметы сливаются в зиготу, происходит восстановление диплоидного числа хромосом, свойственного спорофиту. Прослеживая изменения в числе хромосом и в содержании ДНК, мы видим, что в цветковом растении совершается цикл, в котором диплоидия сменяется гаплоидией, а последующее слияние гаплоидных клеток разного генетического происхождения в новый диплоидный организм порождает новые комбинации генетических признаков. [c.33]
chem21.info