
Формула хлорофилла и его роль в процессе фотосинтеза. Хлорофилл у растений
ХЛОРОФИЛЛЫ — Большая Медицинская Энциклопедия
ХЛОРОФИЛЛЫ (греческий chloros зеленый + phyllon лист) — пигменты растений, а также некоторых микроорганизмов, с помощью которых улавливается энергия солнечного света и осуществляется процесс фотосинтеза. Участвуя в фотосинтезе (см.), хлорофиллы играют огромную биол. роль.
Существует четыре вида хлорофиллов: а, Ь, с и d. Высшие растения содержат хлорофиллы а и Ь, бурые и диатомовые водоросли — хлорофиллы а и с, красные водоросли — хлорофилл d. Кроме того, некоторые фотосинтезирующие бактерии содержат аналоги хлорофиллов — бак-териохлорофиллы. В основе молекул хлорофиллов лежит магниевый комплекс порфиринового цикла (см. Пор-фирины). К одному из пиррольных колец присоединен остаток многоатомного спирта фитола, благодаря чему хлорофиллы получили возможность встраиваться в липидный слой мембраны хлоропластов.
Выделение хлорофиллов в чистом виде и разделение их на два компонента (хлорофиллы а и Ь) впервые было осуществлено русским ботаником М. С. Цветом с помощью разработанного им метода хроматографии (см.). Им же было доказано, что в листьях растений хлорофиллы сопровождает ряд желтых спутников — каротиноидов (см.). Структурная формула хлорофиллов установлена Фишером (Н. Fischer) в 1940 году М. В. Ненцкий и его ученики доказали хим. родство гемоглобина (см.) и хлорофиллов растений. В изучении физиологической роли хлорофиллов большое значение имели исследования К. А. Тимирязева. Полный синтез хлорофиллов произвели независимо друг от друга Штрелль (М. Strell) и Вудворд (R. В. Wood-worcl) в I960 году.
Хлорофиллы являются главной составной частью пигментного аппарата высших растений, мхов, водорослей, фотосинтезирующих бактерий. Содержание их в растениях зависит от вида растения, обеспеченности минеральным питанием и других условий. Количество хлорофиллов в растениях колеблется от 1,7 до 5% в пересчете на сухой вес. Концентрация их на поверхности листа определяет интенсивность поглощения растением света, если уровень хлорофиллов не превышает 2 мг/дм2. При содержании хлорофиллов от 3 мг/дм2 и выше коэффициент поглощения света приближается к 97 —100% и не зависит от количества пигмента.
В клетках зеленого листа хлорофиллы находятся в особых органеллах — пластидах, которые называются также хлорофилловыми зернами, или хлоропластами. Каждый хлоропласт растения Mnium medium имеет объем 4,1 X 10-11 см3 и содержит 1,3-109 молекул хлорофилла, ограничен двойной липопротеидной мембраной и заполнен белковой стромой. Чередующиеся пластинки белка и окрашенных пигментно-липидных слоев образуют включения в строме (граны). Расстояния между молекулами пигмента в тонком моно- или бимолекулярном слоях невелики; каждая из пары молекул может быть связана с ферментами типа цитохрома (см. Цитохромы), способного отдавать электрон хлорофиллу, а другая — с акцептором электрона типа ферредоксина.
Процесс фотосинтеза начинается с поглощения кванта света пигментной системой растения. Участие промежуточных систем в цепи переноса электрона показано на схеме: где X — хлорофилл, ЦИТ — цитохромы, ФД — ферредоксин, ФЛ — флавиновые системы, hv — квант света.
Важное значение в функционирующей фотосинтетической единице имеет процесс миграции энергии между различными формами хлорофилла. Активно функционирующая фотосинтетическая единица содержит 200—400 молекул хлорофилла, которые работают как единая светоулавливающая система, поглощающая один квант света. За один цикл работы на каждые 3000 молекул хлорофилла высвобождается одна молекула кислорода. Установлено, что спектрально различные формы хлорофилла образуют лестницу энергетических уровней, по которой поглощенная энергия «стекает» к реакционным центрам. Спектральные исследования позволили расчленить формы хлорофилла на три основные группы (коротковолновые, длинноволновые и промежуточные) в соответствии с их ролью в поглощении и переносе энергии.
У фотосинтетических бактерий также обнаружены субклеточные частицы, содержащие бактериохлорофилл. Это уплощенные диски диаметром 100 нм, носящие название хроматофоры.
Структуры пигментобелковых комплексов в организации фотосинтетических мембран различных организмов, включая бактерии, водоросли и высшие растения, сходны. Полипептиды хлорофиллобелкового комплекса синтезируются внутри хлоропластов; они состоят из главного полипептида с мол. весом (массой) 73 000 и трех минорных с молекулярным весом (массой) 47 000, 30 000 и 15 000 единиц.
Синтез и обновление пигмента в растущей зеленой ткани протекают с высокой скоростью. С возрастом ткани процесс биосинтеза хлорофилла замедляется. На первых этапах биосинтеза хлорофилла путем конденсации двух молекул Ь-аминолеву-линовой кислоты формируется порфо-билиноген — производное пиррола, которое в результате ряда превращений дает соединение, содержащее порфириновое ядро — протопорфирин. Из протопорфирина образуется непосредственный предшественник хлорофилла — протохлорофиллид, содержащий атом магния. Затем после присоединения многоатомного спирта фитола образуется хлорофилл.
Этапы от порфобилиногена до протопорфирина и от протопорфирина до хлорофилла а осуществляются по одной из двух схем:
Первая реакция преобладает в листьях этиолированных (то есть выросших в темноте) растений, вторая — в зеленых. Терминальные стадии биосинтеза пигментного аппарата ускоряются при участии единого полиферментного хлорофилл-синтетазного комплекса. В связи с этим естественна зависимость биосинтеза хлорофилла от скорости белкового синтеза и торможения его ингибиторами синтеза белка. Синтез пигментов замедляется также при снижении температуры и полностью прекращается при температуре ниже —2°, тогда как фотосинтез продолжается и при отрицательных температурах, вплоть до —24°. Процесс нарушается при недостаточности железа и избытке марганца.
Образование хлорофилла Ь происходит последовательно через хлорофилл а путем окисления. Реакция превращения идет на свету; промежуточной стадией является образование фермент-белкового комплекса.
Есть указания на зависимость скорости реакции от работы электронно-транспортной цепи и соотвественно скорости генерации НАДФН и НАДН как доноров водорода. Остаются неясными стадии синтеза на участке включения магния, превращения Mg-порфиринов, а также этерификации фитолом остатка нропионовой кислоты IV пиррольного кольца.
Способность зеленых растений образовывать в процессе фотосинтеза сложные органические вещества из двуокиси углерода и воды определяется присутствием в них хлорофиллов. При этом содержание пигментов хлорофилла а и хлорофилла Ъ не зависит от географических особенностей местности. Содержание хлорофилла а в большей степени подвержеко влиянию физиологических и экологических условий, чем содержание хлорофилла Ъ.
Описаны изменения хлорофиллов в онтогенезе растений. Их содержание возрастает в фазу кущения, в фазу цветения и завязывания плодов. По уровню хлорофилла можно определить готовность растений к цветению. После завершения ростовых процессов накопление хлорофилла прекращается, и обновление молекул пигмента происходит внутри хлоропласта, не будучи связано с образованием новых хлоропластов.
Принцип фотосенсибилизирующего действия хлорофиллов при фотосинтезе был обоснован К. А. Тимирязевым и включает возбуждение пигмента светом с переходом пигмента в синглетное или триплетное состояние и последующими обратимыми фотохимическими изменениями. Хлорофилл на разных этапах может служить фотохимическим донором или акцептором электронов.
Поскольку тетрапиррольным структурам, содержащим комплексно связанный атом железа, принадлежит важная роль в тканевом дыхании млекопитающих (см. Гемоглобин), хлорофилл и его металлопроизводные (т. е. соединения, в структуру которых вместо магния введены медь, железо, цинк, кадмий или серебро) используют в медицине в качестве антигипоксических средств. Металлопроизводные хлорофилла получили название «феофитинаты». Их антигипоксический эффект связывают с тетрапиррольной структурой и присутствием атома металла. Водорастворимые препараты хлорофилла обладают антибактериальной и противовирусной активностью, особенно Ag-феофитинат. Гемопоэтические, общетонизирующие свойства присущи хлорофиллину натрия, который используют также в качестве биостимулятора.
См. также Ассимиляция, Пигментный обмен, Пигменты, Порфирины.
Библиогр.: Годнев Т. Н. Хлорофилл, Его строение и образование в растении, Минск, 1963, библиогр.; К р а с-новский А. А. Уровни светового регулирования фотосинтеза, в кн.: Теоретические основы фотосинтетической продуктивности, под ред. А. А. Ничипорови-ча, с. 23, М., 1972; Мецлер Д. Э. Биохимия, Химические реакции в живой клетке, пер. с англ., т. 1—2, М., 1980; Проблемы биосинтеза хлорофиллов, под ред. А. А. Шлыка, Минск, 1971; Шлык А. А. Метаболизм хлорофилла в зеленом растении, Минск, 1965, библиогр.; Е igenberg К. Ё., С г о a s-m u n W. R. a. Chan S. I. Chlorophyll a in bilayer membranes, Biochim. biophys. Acta, v. 679, p. 353, 1982; Metabolic pathways, ed. by D. M. Greenberg, v. 2, N. Y.— L., 1967; Olson J. M. Chlorophyll organization in green photosynthetic bacteria, Biochim. biophys. Acta, v. 594, p. 33, 1980.
II. А. Верболович, В. П. Верболович.
xn--90aw5c.xn--c1avg
Формула хлорофилла и его роль в процессе фотосинтеза
Почему трава, а также листья на деревьях и кустах зеленые? Виной всему хлорофилл. Можно взять прочную веревку знаний и завязать с ним крепкое знакомство.
История
Проведем небольшой экскурс в сравнительно недалекое прошлое. Жозеф Бьенеме Каванту и Пьер Жозеф Пеллетье – вот кому нужно пожать руку. Мужи науки постарались отделить зеленый пигмент из листьев разных растений. Старания увенчались успехом в 1817 году.
Пигмент наименовали хлорофилл. От греческого chloros – зеленый, и phyllon - лист. Независимо от вышесказанного, в начале 20 века Михаил Цвет и Рихард Вильштеттер пришли к выводу: оказывается, в хлорофилл входит несколько компонентов.
Засучив рукава, Вильштеттер принялся за работу. Очистка и кристаллизация выявили два компонента. Назвали их просто, альфа и бета (а и b). За труды в поле исследования данного вещества в 1915 году ему торжественно вручили премию Нобеля.
В 1940 Ханс Фишер предложил всему миру окончательную структуру хлорофилла «а». Король синтеза Роберт Бернс Вудворд и несколько ученых из Америки получили в 1960 году ненатуральный хлорофилл. Так и приоткрылась завеса тайны – появление хлорофилла.

Химические свойства
Формула хлорофилла, определенная из опытных показателей, выглядит так: C55H72O5N4Mg. В конструкцию входит органическая дикарбоновая кислота (хлорофиллин), а также спирты метиловый и фитол. Хлорофиллин – это металлорганическое соединение, имеющее прямое отношение к магнийпорфиринам и содержащее азот.
COOH
MgN4Oh40C32
COOH
Хлорофилл значится сложным эфиром из-за того, что оставшиеся части метилового спирта Ch4OH и фитола C20h49OH заместили водород карбоксильных группировок.
Выше размещена структурная формула хлорофилла альфа. Разглядев ее внимательно, можно увидеть, что у бета-хлорофилла на один атом кислорода больше, но на два атома водорода меньше (группа CHO вместо Ch4). Отсюда молекулярная масса альфа-хлорофилла ниже, чем у бета.
В середине частицы интересующего нас вещества обосновался магний. Он соединяется с 4 атомами азота пиррольных формирований. Систему элементарных и сменяющихся двойных связей можно наблюдать в пиррольных связках.
Хромофорное формирование, удачно вписываемое в состав хлорофилла - это и есть N. Оно делает возможным впитывание отдельных лучей солнечного спектра и его цвет, независимо от того, что днем солнце горит, как пламя, а вечером похоже на тлеющие угли.

Перейдем к размерам. Порфириновое ядро в диаметре 10 нм, фитольный фрагмент оказался длинной в 2 нм. В ядре хлорофилл составляет 0,25 нм, меж микрочастицами пиррольных группок азота.
Хотелось бы отметить, что атом магния, который входит в состав хлорофилла, в диаметре всего 0,24 нм и практически полностью заполняет свободное место между атомами пиррольных группировок азота, что помогает ядру молекулы быть более крепким.
Можно прийти к выводу: из двух составляющих под нехитрым названием альфа и бета и состоит хлорофилл (a и b).
Хлорофилл a
Относительная масса молекулы - 893,52. Создают в отделенном пребывании микрокристаллы черного цвета с голубым отливом. При температуре 117-120 градусов Цельсия они расплавляются и перевоплощаются в жидкость.
В этаноле такие же хлороформы, в ацетоне, а еще бензолах растворяются охотно. Результаты принимают сине-зеленую окраску и имеют отличительную особенность - насыщенная красная флуоресценция. Плохо растворяются в петролейном эфире. В воде не распускаются вовсе.
Формула хлорофилла альфа: C55H72O5N4Mg. Вещество по своей химической конструкции относят к хлоринам. В кольце к пропионовой кислоте, а именно к ее остатку, прикреплен фитол.
Кое-какие растительные организмы, вместо хлорофилла a, образуют его аналог. Здесь этильную группу (-Ch3-Ch4) во II пиррольном кольце сменила винильная (-CH=Ch3). Такая молекула заключает в себе первую винильную группу в кольце один, вторую в кольце два.
Хлорофилл b
Формула хлорофилла-бета имеет следующий вид: C55H70O6N4Mg. Молекулярный вес вещества составляет 903. У атома углерода C3 в пиррольном кольце два, обнаруживается немного спирта, лишенного водорода –H-C=O, который обладает желтым цветом. Это и есть отличие от хлорофилла a.
Смеем заметить, что в специальных постоянных частях клетки, жизненно важных для ее дальнейшего существования пластидах-хлоропластах, пребывают несколько типов хлорофиллов.

Хлорофиллы c и d
У криптомонад, динофлагеллятов, а также у бациллариофициевых и бурых водорослей найден хлорофилл с. Классический порфирин – вот чем отличается этот пигмент.
У водорослей красной окраски хлорофилл d. Некоторые сомневаются в его существовании. Полагается, что он является только продуктом вырождения хлорофилла a. На данный момент можно уверенно сказать, что хлорофилл с литерой d – это основной краситель кое-каких фотосинтезирующих прокариотов.
Свойства хлорофилла
После продолжительных исследований возникло доказательство, что в особенностях хлорофилла, пребывающего в растении и добытого из него, замечена несхожесть. Хлорофилл в растениях соединен с белком. Об этом свидетельствуют следующие наблюдения:
- Спектр впитывания хлорофилла в листе другой, если сравнить его с извлеченным.
- Чистым спиртом из высохших растений предмет описания достать нереально. Протекает экстракция благополучно при хорошо увлажненных листьях, либо следует долить в спирт воду. Именно она разбивает связанный с хлорофиллом белок.
- Материал, вытянутый из листьев растений, быстро разрушается под влиянием кислорода, концентрированной кислоты, световых лучей.
Зато хлорофилл в растениях устойчив ко всему вышеперечисленному.

Хлоропласты
В растениях хлорофилла содержится 1% от сухого вещества. Найти можно в особых органеллах клетки – пластидах, что показывает неравномерное распределение его в растении. Пластиды клеток, окрашенные в зеленый цвет и имеющие в себе хлорофилл, имеют название хлоропласты.
Количество h3O в хлоропластах колеблется от 58 до 75%, содержимое сухого вещества состоит из белков, липидов, хлорофилла и каротиноидов.
Функции хлорофилла
Удивительное сходство обнаружили ученые в устройстве молекул хлорофилла и гемоглобина – главного дыхательного компонента человеческой крови. Отличие состоит в том, что в клешневидном соединении посередине в пигменте растительного происхождения размещен магний, а в гемоглобине - железо.
В ходе фотосинтеза растительность планеты поглощает углекислый газ, выделяет кислород. Вот еще одна замечательная функция хлорофилла. По деятельности его можно сравнить с гемоглобином, но объем воздействия на человеческий организм несколько больше.

Хлорофилл – это растительный пигмент, чувствительный к свету и покрытый зеленым цветом. Далее идет фотосинтез, при котором его микрочастицы преобразовывают энергию солнца, поглощаемую клетками растений, в химическую энергию.
Можно прийти к следующим умозаключениям, что фотосинтез – это процесс преобразования энергии солнца. Если доверять современным сведениям, замечено, что протекание синтеза органических веществ из газа углекислого и воды с использованием световой энергии разложено на три этапа.
Этап №1
Данная фаза вершится в процессе фотохимического распада воды, при содействии хлорофилла. Отмечается выделение молекулярного кислорода.
Этап №2
Здесь наблюдается несколько окислительно-восстановительных реакций. В них берут активное содействие цитохромы и иные переносчики электронов. Реакция происходит за счет световой энергии, переносимой электронами от воды на NADPH и образующей ATP. Тут запасается световая энергия.

Этап №3
Уже образовавшиеся NADPH и ATP пускаются в ход для преобразования углекислого газа в углевод. Поглощенная энергия света участвует в реакциях 1 и 2 этапов. Реакции последнего, третьего, происходят без участия света и называются темновыми.
Фотосинтез – это единственный биологический процесс, проходящий с возрастанием свободной энергии. Прямо или косвенно обеспечивает доступной химической предприимчивостью обитающих на земле двуногих, крылатых, бескрылых, четвероногих и прочие организмы.
Гемоглобин и хлорофилл
Молекулы гемоглобина и хлорофилла имеют сложную, но в то же время схожую атомарную структуру. Общим в их строении является профин – кольцо из маленьких колечек. Различие замечено в отросточках, присоединённых к профину, и в атомах, расположенных внутри: атом железа (Fe) у гемоглобина, у хлорофилла магний (Mg).
Хлорофилл и гемоглобин по строению похожи, но формируют разные белковые структуры. Вокруг атома магния сформирован хлорофилл, вокруг железа - гемоглобин. Если взять молекулу жидкого хлорофилла и отсоединить фитольный хвост (20 углеродную цепь), поменять атом магния на железо, то зеленый цвет пигмента станет красным. В итоге - готовая молекула гемоглобина.

Усваивается хлорофилл легко и быстро, благодаря именно такому сходству. Хорошо поддерживает организм при кислородном голодании. Насыщает кровь нужными микроэлементами, отсюда она лучше транспортирует важнейшие для жизни вещества к клеткам. Происходит своевременный выброс отработанных материалов, токсинов, отходов, возникающих в результате естественного обмена веществ. Имеет воздействие на спящие лейкоциты, пробуждая их.
Описываемый герой без страха и упрека защищает, укрепляет мембраны клетки, помогает восстановиться соединительной ткани. К заслугам хлорофилла можно отнести быстрое заживление язв, разных ран и эрозий. Улучшает иммунную работу, выделена способность купировать патологические нарушения молекул ДНК.
Положительная тенденция при лечении инфекционных и простудных заболеваний. Это не весь список добрых дел рассмотренного вещества.
fb.ru
их состав, структура и свойства
Впервые хлорофилл в кристаллическом виде был описан русским физиологом и ботаником И. П. Бородиным в 1883 г. В дальнейшем оказалось, что это не сам хлорофилл, а несколько видоизмененная его форма — этилхлорофиллид. Польские биохимики М. Ненцкий и Л. Мархлевский (1897) обнаружили, что основу молекулы хлорофилла, как и гема гемоглобина, составляет порфириновое кольцо. Таким образом было показано принципиальное структурное сходство этих пигментов у растений и животных.
Немецкий химик Р. Вильштеттер в 1906—1914 гг. установил элементарный состав хлорофилла а — C 55 H 72 O 5 N 4 Mg и хлорофилла b — C 55 H 70 O 6 N 4 Mg, а немецкий биохимик Г. Фишер в 1930—1940 гг. полностью расшифровал структурную формулу хлорофилла. В 1960 г. химики-органики Р. Б. Вудворд (США) и М. Штрель (ФРГ) осуществили искусственный синтез хлорофилла. Хлорофилл — сложный эфир дикарбоновой кислоты хлорофиллина, у которой одна карбоксильная группа этерифицирована остатком метилового спирта, а другая остатком одноатомного непредельного спирта фитола. Структурная формула хлорофилла а представлена на рисунке 1.
 Рис.1. Структурная формула хлорофилла а и b
Рис.1. Структурная формула хлорофилла а и b
Четыре пиррольных кольца ( I — IV ) соединены между собой метановыми мостиками (а, р, у, 5), образуя порфириновое ядро. Наружные атомы углерода в тетрапиррольном цикле обозначены цифрами 1 — 10. Атомы азота пиррольных колец четырьмя координационными" связями взаимодействуют с атомом магния. В структуре порфиринового ядра есть также цикло-пёнтановое кольцо (V), образованное остатком кетопропионовой кислоты и содержащее химически активную карбонильную группу у С9 и метилированную карбоксильную группу у С10. Структура, состоящая из тетрапиррольного и циклопентанового колец, получила название форбина. Боковая цепь IV пиррольного кольца включает в себя пропионовую кислоту, связанную сложноэфирной связью с полиизопреновым непредельным спиртом фитолом (С20Н39ОН). У 1, 3, 5 и 8-го углеродов пиррольных колец имеются метальные группы, у 2-го — винильная, у 4-го — этильная группа. Порфириновое кольцо представляет собой систему из девяти пар конъюгированных (сопряженных) чередующихся двойных и одинарных связей с 18 делокализованными p-электронами. Хлорофилл b отличается от хлорофилла а тем, что у 3-го углерода вместо метильной находится формильная (—СНО) группа. Структура хлорофилла, лишенная фитола, называется хлорофиллидом. При замещении атома магния протонами в молекуле хлорофиллов образуются соответствующие феофетины.
У высших растений и водорослей обнаружены хлорофиллы а, b, с, d. Все фотосинтезирующие растения, включая все группы водорослей, а также цианобактерии, содержат хлорофиллы группы а. Хлорофилл b представлен у высших растений, у зеленых водорослей и эвгленовых. У бурых и диатомовых водорослей вместо хлорофилла b присутствует хлорофилл с, а у многих красных водорослей — хлорофилл d. В фотосинтезирующих бактериях, осуществляющих фоторедукцию, найдены различные бактериохлорофиллы.
Биосинтез хлорофиллов
Первый этап биосинтеза хлорофиллов у растений — образование 5-аминолевулиновой кислоты (А Л К) из С5-дикарбоновых кислот. Показано, что глутаминовая кислота через 2-гидроксиглутаровую превращается в 4,5-диоксовалериановую, которая затем аминируется за счет аланина или других аминокислот (рис. 2). Реакция переаминирования катализируется АЛК-трансаминазой с участием пиридоксальфосфата в качестве кофермента. Для синтеза АЛК может использоваться и оскетоглутаровая (2-оксоглутаровая) кислота. Циклизация двух молекул АЛК приводит к образованию пиррольного соединения — порфобилиногена. Из четырех пиррольных колец формируется уропорфириноген, который превращается в протопорфирин IX. Дальнейший путь превращений протопорфирина может быть различным. С участием железа формируется гем, входящий в состав цитохромов, каталазы, пероксидазы и гемоглобина. Если молекулу протопорфирина включается магний, затем карбоксильная группа у С10 этерифицируется метильной группой 6-аденозил-b-метионина и замыкается циклопентановое кольцо (К), то образуется протохлорофиллид.
 Рис.2. Биосинтез молекулы хлорофилла
Рис.2. Биосинтез молекулы хлорофилла
Под действием света в течение нескольких секунд протохлорофиллид превращается в хлорофиллид а в результате гидрирования двойной связи у С7 — С8 в IV пиррольном ядре. У низших растений и некоторых голосеменных (у хвойных) хлорофиллид может образовываться в темноте. Хлорофиллид обладает теми же спектральными свойствами, что и хлорофилл. Последний этап в формировании молекулы хлорофилла а — этерификация фитолом, который, как и все полиизопреновые соединения, синтезируется из ацетил-СоА через мевалоновую кислоту. По данным А. А. Шлыка (1965), хлорофилл b может образовываться из вновь синтезированных молекул хлорофилла а. Все описанные процессы, начиная с образования АЛК, осуществляются в хлоропластах. Первые признаки зеленения выращенных в темноте этиолированных покрытосеменных растений наблюдаются через 2 — 4 ч после начала освещения.
Физико-химические свойства хлорофиллов
В твердом виде хлорофилл а представляет собой аморфное вещество сине-черного цвета. Температура плавления хлорофилла а 117 — 120 °С. Хлорофиллы хорошо растворимы в этиловом эфире, бензоле, хлороформе, ацетоне, этиловом спирте, плохо растворимы в петролейном эфире и нерастворимы в воде. Раствор хлорофилла а в этиловом эфире имеет сине-зеленый цвет, хлорофилла b — желто-зеленый. Резко выраженные максимумы поглощения хлорофиллов лежат в красной и синей частях спектра (рис.3).
 Рис.3. Спектры поглощения пигментов пластид
Рис.3. Спектры поглощения пигментов пластид
В этиловом эфире максимумы поглощения хлорофиллов группы а в красной части спектра — в пределах 660 — 663 нм, в синей — 428 — 430 нм, хлорофилла b — соответственно в пределах 642 — 644 и 452 — 455 нм. Хлорофиллы очень слабо поглощают оранжевый и желтый свет и совсем не поглощают зеленые и инфракрасные лучи. Поглощение в сине-фиолетовой части спектра обусловлено системой конъюгированных одинарных и двойных связей порфиринового кольца молекулы хлорофилла. Интенсивная полоса поглощения в красной области связана с гидрированием двойной связи у С7 — С8 в IV пиррольном ядре (при переходе от протохлорофиллида к хлорофиллиду) и присутствием магния в порфириновом кольце. Эти же условия способствуют снижению поглощения в желтой и зеленой частях спектра. Замещение магния протонами при обработке хлорофилла кислотой приводит к образованию феофетина, имеющего буро-зеленый цвет и ослабленный красный максимум поглощения. Удаление остатков фитола и метилового спирта путем щелочного гидролиза мало сказывается на спектре поглощения хлорофилла. У хлорофиллина те же спектральные свойства, что и у хлорофилла: он зеленый, имеет красный и синий максимумы поглощения.
На положение максимумов спектра поглощения оказывают влияние природа растворителя и взаимодействие молекул хлорофилла друг с другом, а также с другими пигментами, липидами и белками. У агрегированных молекул хлорофилла (например, в твердых пленках и у хлорофилла, находящегося в хлоропластах) красный максимум поглощения сдвинут в более длинноволновую область (до 680 нм).
Растворы хлорофиллов в полярных растворителях обладают яркой флуоресценцией (люминесценцией). В этиловом эфире у хлорофилла а наблюдается рубиново-красная флуоресценция с максимумом 668 нм, у хлорофилла b — 648 нм т. е. максимумы флуоресценции в соответствии с правилом Стокса несколько сдвинуты в более длинноволновую часть спектра по отношению к максимумам поглощения. Агрегированный хлорофилл и хлорофилл в нативном состоянии (в живом листе) флуоресцируют слабо. Растворы хлорофиллов способны также к фосфоресценции (т. е. длительному послесвечению), максимум которого лежит в инфракрасной области.
Механизмы флуоресценции и фосфоресценции, в том числе молекул хлорофилла, достаточно хорошо изучены. Наиболее устойчивы те состояния атомов и молекул, в которых валентные электроны занимают самые низкие энергетические уровни и распределены по ним согласно принципу Паули (не более двух электронов с антипараллельными спинами на каждой орбитали). Такое состояние молекулы называют основным синглетным ( S0 ) энергетическим состоянием (рис.4). Суммарный спин (вектор магнитных моментов) всех электронов молекулы в этом случае равен нулю. Когда молекула поглощает квант света, то за счет этой энергии электрон переходит на более высокие вакантные орбитали. Если у возбужденного электрона сохраняется то же направление спина, то молекула находится в синглетном возбужденном состоянии ( S*). Если при переходе на более высокую орбиталь спин электрона обращается, то такое возбуждение называется триплетным (T*). Прямой переход из основного состояния в триплетное — маловероятное событие, так как для этого необходимо обращение ориентации спина. Переходу молекулы из основного состояния в возбужденное, происходящему при поглощении кванта света, соответствует полоса в спектре поглощения. Поглощение молекулой хлорофилла кванта красного света приводит к синглетному электронвозбужденному состоянию — S*1.
 Рис.4. Энергетические состояния молекулы хлорофилла
Рис.4. Энергетические состояния молекулы хлорофилла
При поглощении кванта синего света с более высоким уровнем энергии электрон переходит на более высокую орбиту ( S*2). Возбужденная молекула хлорофилла возвращается в основное состояние различными путями. Отдав часть энергии в виде теплоты (особенно с уровня S*1), молекула может излучить квант света с большей длиной волны (правило Стокса), что проявляется в виде флуоресценции. Как уже отмечалось, хлорофилл а поглощает свет в красной и синей областях спектра, но флуоресцирует лишь в красной. Время жизни синглетного возбужденного состояния 10-13 — 10-9 с.
Другой путь потери энергии состоит в переходе молекулы из синглетного возбужденного состояния в метастабильное триплетное (с обращением спина). Этот переход безизлучательный, т. е. энергия теряется в виде теплоты. Триплетное возбужденное состояние имеет гораздо большее время жизни (>10-4 с). Из триплетного состояния молекула может вернуться в основное, излучив еще более длинноволновый (чём в случае флуоресценции) квант света. Это обычно более слабое свечение и есть фосфоресценция.
Наконец, энергия возбужденного состояния может быть использована на фотохимические реакции. В этом случае флуоресценции и фосфоресценции хлорофилла не наблюдается.
Структура молекулы хлорофилла, отобранная в процессе эволюции из многих других органических пигментов, прекрасно приспособлена к своим функциям сенсибилизатора фотохимических реакций. В ее состав входят 18 делокализованных π-электронов (представленных в структурной формуле хлорофилла в виде 18-членного кольца из конъюгированных двойных связей), что делает молекулу хлорофилла легко возбудимой при поглощении квантов света.
Еще Тимирязев предположил, что хлорофилл способен к окислительно-восстановительному превращению. Впервые реакция фотовосстановления хлорофилла была осуществлена в модельных опытах А. А. Красновским в 1948 г. Хлорофилл, растворенный в пиридине, в анаэробных условиях под действием света восстанавливается аскорбиновой кислотой или другими донорами электронов. При этом образуется восстановленная («красная») форма хлорофилла с максимумом поглощения при 525 нм.
После выключения света реакция идет в обратном направлении. Фотовосстановленный хлорофилл в свою очередь может восстанавливать различные акцепторы электронов. В той же модельной системе, но с добавлением акцептора электронов хлорофилл при освещении действует как сенсибилизатор. В этих условиях происходит восстановление NAD+, рибофлавина, хинона, Fe3+, кислорода. Таким образом, молекула хлорофилла может выступать не только в роли первичного акцептора электрона, но и в роли его первичного донора.
Из всего сказанного следует, что молекула хлорофилла благодаря структурным и физико-химическим особенностям способна выполнять три важнейшие функции: 1) избирательно поглощать энергию света, 2) запасать ее в виде энергии электронного возбуждения, 3) фотохимически преобразовывать энергию возбужденного состояния в химическую энергию первичных фотовосстановленных и фотоокисленных соединений.
Для функционирования молекулы хлорофилла в процессах фотосинтеза существенное значение имеет ее пространственная организация. Магний-порфириновое кольцо молекулы представляет собой почти плоскую пластинку толщиной 0,42 нм и площадью 1 нм2. Это гидрофильная часть молекулы хлорофилла. Длинный алифатический остаток фитола (2 нм), образующий угол с порфириновым кольцом, — ее гидрофобный полюс, необходимый для взаимодействия молекулы хлорофилла с гидрофобными участками мембранных белков и липидов.
biofile.ru
Физические свойства хлорофилла
Хлорофилл способен к избирательному поглощению света и к флюоресценции. Спектр поглощения данного соединения определяется его способностью поглощать свет определенной длины волны (определенного цвета). Для того чтобы получить спектр поглощения, К. А. Тимирязев пропускал луч света сначала через раствор хлорофилла, а затем через призму. В этом случае часть лучей поглощалась. Было показано, что хлорофилл в той же концентрации, как в листе, имеет две основные линии поглощения в красных и сине-фиолетовых лучах. При этом хлорофилл а в растворе имеет максимумы поглощения 429 и 660 нм, тогда как хлорофилл b — 453 и 642 нм. Однако необходимо учитывать, что в листе спектры поглощения хлорофилла меняются в зависимости от его состояния, степени агрегации, адсорбции на определенных белках. В настоящее время показано, что есть формы хлорофилла, поглощающие свет с длиной волны 700, 710 и даже до 720 нм. Формы хлорофилла, поглощающие свет с большой длиной волны, имеют особенно важное значение в процессе фотосинтеза. Хлорофилл обладает способностью к флюоресценции. Флюоресценция представляет собой свечение тел, возбуждаемое освещением и продолжающееся очень короткий промежуток времени (10-8—10-9 с). Свет, испускаемый при флюоресценции, имеет всегда большую длину волны по сравнению с поглощенным. Это связано с тем, что часть поглощенной энергии выделяется в виде тепла. Хлорофилл обладает красной флюоресценцией.
Биосинтез хлорофилла
Синтез хлорофилла происходит в две фазы: темновую — до протохлорофиллида и световую — образование из протохлорофиллида хлорофилла. Для превращения протохлорофиллида в хлорофиллид необходимо его связывание с белком голохромом и присоединение двух атомов водорода. Именно последняя реакция для большинства растений протекает с использованием энергии света (фотовосстановление). Водороды присоединяются к 7-му и 8-му атомам углерода. На последнем этапе к хлорофиллиду присоединяется спирт фитол. Поскольку синтез хлорофилла — процесс многоэтапный, в нем участвуют различные ферменты, составляющие, по-видимому, полиферментный комплекс. Интересно заметить, что образование многих белков-ферментов ускоряется на свету. Содержание хлорофилла в листе колеблется незначительно. Это связано с тем, что идет непрерывный процесс разрушения старых молекул и образование новых молекул хлорофилла. Причем эти два процесса уравновешивают друг друга. При этом предполагается, что вновь образовавшиеся молекулы хлорофилла не смешиваются со старыми и имеют несколько иные свойства.
Условия образования хлорофилла
Проростки, выросшие в отсутствии света, называют этиолированными. Такие проростки, как правило, характеризуются измененной формой (вытянутые стебли, неразвившиеся листья) и слабой желтой окраской (хлорофилла в них нет). Известно, что в некоторых случаях хлорофилл образуется и в отсутствии света. Способность образовывать хлорофилл в темноте характерна для растений, стоящих на нижней ступени эволюционного процесса. Так, при благоприятных условиях питания некоторые бактерии могут синтезировать в темноте желто-зеленый пигмент — бактериохлорофилл. Сине-зеленые водоросли при достаточном снабжении органическим веществом растут и образуют пигменты в темноте.
Способность к образованию хлорофилла в темноте обнаружена и у таких высокоорганизованных водорослей, как харовые. Лиственные и печеночные мхи сохраняют способность образовывать хлорофилл в темноте. Почти у всех видов хвойных при прорастании семян в темноте семядоли зеленеют. Более развита эта способность у теневыносливых пород хвойных деревьев. По мере роста проростков в темноте образовавшийся хлорофилл разрушается, и на 35—40-й день проростки в отсутствии света погибают. Интересно заметить, что проростки хвойных, выращенные из изолированных зародышей в темноте, хлорофилла не образуют. Однако достаточно присутствия небольшого кусочка нераздробленного эндосперма, чтобы проростки начинали зеленеть. Зеленение происходит даже в том случае, если зародыш соприкасается с эндоспермом другого вида хвойных деревьев. При этом наблюдается прямая корреляция между величиной окислительно-восстановительного потенциала эндосперма и способностью проростков зеленеть в темноте.
В целом рассмотрение этого вопроса приводит к заключению, что в эволюционном плане хлорофилл первоначально образовался как побочный продукт процесса темнового обмена. Однако в дальнейшем на свету растения, обладающие хлорофиллом, получили большее преимущество благодаря возможности использовать энергию солнечного света, и эта особенность была закреплена естественным отбором.
Исследования влияния света на накопление хлорофилла в этиолированных проростках показали, что первым в процессе зеленения появляется хлорофилл а. Нормальное соотношение хлорофилла а к хлорофиллу b (3:1) наступает только через несколько часов после начала освещения, при этом, хлорофилл b образуется из хлорофилла а.
Спектрографический анализ показывает, что процесс образования хлорофилла идет очень быстро. Так, уже через 1 мин после начала освещения выделенный из этиолированных проростков пигмент имеет спектр поглощения, совпадающий со спектром поглощения хлорофилла а.
При исследовании влияния качества света на образование хлорофилла в большинстве случаев проявилась положительная роль красного света. Большее значение имеет интенсивность освещения. Оказалось, что освещение электрической лампой мощностью 10 Вт на расстоянии 400 см было пределом, ниже которого образование хлорофилла прекращалось. Существует и верхний предел освещенности, выше которого образование хлорофилла тормозится.
Целый ряд исследований показывает, что образование хлорофилла идет интенсивнее на прерывистом свете. Это подтверждает, что в образовании хлорофилла имеется темновая и световая фазы. При этом световая фаза значительно короче темновой.
Образование хлорофилла зависит от температуры. Оптимальная температура для накопления хлорофилла 26—30°С. Как и следовало ожидать, от температуры зависит лишь образование предшественников хлорофилла (темновая фаза). При наличии уже образовавшихся предшественников хлорофилла процесс зеленения (световая фаза) идет с одинаковой скоростью независимо от температуры.
На скорость образования хлорофилла оказывает влияние содержание воды. Сильное обезвоживание проростков приводит к полному прекращению образования хлорофилла. Особенно чувствительно к обезвоживанию образование протохлорофиллида. Показана необходимость углеводов для протекания процесса зеленения. Именно с этим связано то, что зеленение этиолированных проростков на свету зависит от их возраста. После 7—9-дневного возраста способность к образованию хлорофилла у таких проростков резко падает. При опрыскивании сахарозой проростки снова начинают интенсивно зеленеть.
Важнейшее значение для образования хлорофилла имеют условия минерального питания. Прежде всего, необходимо достаточное количество железа. При недостатке железа даже листья взрослых растений теряют окраску. Это явление названо хлорозом. Железо — необходимый катализатор образования хлорофилла. Оно необходимо на этапе синтеза σ-аминолевулиповой кислоты из глицина и сукцинил-КоА, а также синтеза протопорфирина. Большое значение для обеспечения синтеза хлорофилла имеет нормальное снабжение растений азотом и магнием, так как оба эти элемента входят в состав хлорофилла. При недостатке меди хлорофилл легко разрушается. Это, по-видимому, связано с тем, что медь способствует образованию устойчивых комплексов между хлорофиллом и соответствующими белками.
Исследование процесса накопления хлорофилла у растений в течение вегетационного периода показало, что максимальное содержание хлорофилла приурочено к началу цветения. Есть даже мнение, что повышение образования хлорофилла может быть использовано как индикатор, указывающий на готовность растений к цветению. Синтез хлорофилла зависит от деятельности корневой системы. Так, при прививках содержание хлорофилла в листьях привоя зависит от свойств корневой системы подвоя. Возможно, что влияние корневой системы связано с тем, что там образуются гормоны (цитокинины). У двудомных растений большим содержанием хлорофилла характеризуются листья женских особей.
studfiles.net
Что такое хлорофилл? Зеленое вещество растений

Какая основная характеристика растений отличает их от животных? Растения — зеленые. Конечно, имеются и исключения, но основным требованием, предъявляемым к растениям, является то, что они должны быть зелеными.
Хлорофилл в растениях
Зеленый цвет растений является одним из основных требований. Зеленое вещество растений — хлорофилл — поглощает необходимые вещества из почвы и воздуха и производит продукты, необходимые для их существования. Если бы растения не обладали этим свойством, человек и животные не смогли бы существовать из-за отсутствия продуктов питания! Даже те существа, которые употребляют для еды животную пищу, зависят от наличия других существ, которые питаются растительной пищей. И действительно, растительна пища является началом всего живого на Земле!
Теперь тебе ясно, что хлорофилл, это удивительное зеленое вещество, которое обеспечивает человека и животных растительной пищей, очень важен для нашей жизни! Хлорофилл содержится в клетках листьев, а также в стеблях и цветках.
Для чего нужен хлорофилл
С помощью хлорофилла растение поглощает солнечный свет и использует полученную энергию для превращения неорганических элементов в органические, или «жизнетворные» химические элементы. Этот процесс называется «фотосинтезом». В переводе с греческого это слово означает «свет» и «соединение».
Существуют некоторые растения, которые не обладают зеленой окраской, то есть не имеют хлорофилла. Как же они существуют? Грибы и семейство растений, у которых отсутствует хлорофилл, не вырабатывают своей пищи. Поэтому они должны получать ее откуда-то еще. Если растение получает пищу за счет других растений или животных, их называют «паразиты». Если они получают пищу за счет продуктов распада растений и животных, мы относим их к «сапрофитам».
Хлорофилл может извлекаться из растений и использоваться человеком для различных целей. В некоторых случаях он убивает определенные бактерии.
Другие интересные статьи
Как был открыт кофе? История напитка...
9 головоломок, на разгадку которых может понадобиться целая ...
Еврипид, основатель драматургии
Поделиться ссылкой:
Понравилось это:
Нравится Загрузка...
intofact.ru
Хлорофилл - функция, отзывы, состав
Хлорофилл – зеленый светочувствительный пигмент растений. Молекулы хлорофилла обладают уникальной способностью преобразовывать энергию солнца, поглощаемую растительными клетками, в химическую энергию. Процесс преобразования солнечной энергии называется фотосинтезом.
Функции хлорофилла
Ученые обнаружили поразительное сходство в строении молекулы хлорофилла и молекулы гемоглобина – главного дыхательного компонента крови человека. Единственное отличие в их строении в том, что в центре хелатного комплекса в растительном пигменте располагается атом магния, а в гемоглобине – атом железа.
Растения, поглощая углекислый газ в процессе фотосинтеза, выделяют в воздух кислород. Образование кислорода в процессе фотосинтеза – еще одна удивительная функция хлорофилла. По выполняемым функциям хлорофилл можно сравнить с гемоглобином, однако спектр воздействия хлорофилла на организм человека значительно шире.
Во многих отзывах о хлорофилле подтверждается то, что данное вещество активизирует функцию поджелудочной железы и улучшает работу щитовидной железы. Пигмент регулирует артериальное давление, устраняет нервозность, способствует оздоровлению кишечника. Известный врач Б.С. Тайц рекомендовал добавлять хлорофилл в пищу детям при аллергии.
Отзывы о хлорофилле подтверждают, что это вещество значительно ускоряет заживление язвы двенадцатиперстной кишки и язвы желудка. Кроме того, он укрепляет мембраны клеток, ускоряет фагоцитоз, усиливает иммунитет.
Уникальный состав хлорофилла делает его прекрасным средством для устранения потенциально вредных для организма человека веществ. Пигмент связывает и выводит токсины из организма. Немаловажной функцией хлорофилла является защита клеток от разрушения свободными радикалами. Данное вещество способно оказывать противоопухолевое действие. В некоторых отзывах о хлорофилле говорится, что он оказывает благотворное влияние на пищеварительную, сердечно-сосудистую и дыхательную системы.
Благодаря своему составу хлорофилл является мощным антиоксидантом и тонизирующим средством, замедляющим процессы старения.
Витамин К в составе хлорофилла, регулируя скорость образования кристаллов оксалата кальция в моче, предотвращает развитие мочекаменной болезни.
В фармацевтической промышленности на основе хлорофилла производят различные биологически активные добавки. В пищевой промышленности его используют в качестве пищевой добавки Е140. Натуральный краситель Е140 добавляют при изготовлении мороженого, кремов, молочных десертов, майонеза и различных соусов для придания продуктам приятного оливкового оттенка.
Получение жидкого хлорофилла
Чаще всего жидкий хлорофилл получают из люцерны. Из листьев растения выжимают сок, после чего сок высушивают по специальной технологии. Из готового продукта делают раствор либо изготавливают капсулы.
Жидкий хлорофилл, полученный из люцерны, наделен всеми полезными свойствами растения. Люцерна – самый лучший источник жидкого хлорофилла. Большая и глубокая система корней позволяет растению доставать из недр земли все самое полезное. Все витамины и минералы в люцерне находятся в легкой для усваивания организмом форме. В ней достаточно много железа, магния и калия. В растении в четыре раза больше аскорбиновой кислоты, чем в цитрусовых.
Хлорофилловые добавки очень полезны любому человеку, поскольку они помогают обезвредить канцерогены, попадающие в организм вместе с пищей.
www.neboleem.net
фотосинтез, хлорофилл, хлоропласты и производительность растений — Сириус агро плант
ФОТОСИНТЕЗ
Фотосинтез является единственным механизмом получения энергии для биосферы (за исключением процессов, происходящих у некоторых хемосинтезирующих бактерий, которые получают энергию от окисления неорганических субстратов). Фотосинтез включает в себя окислительно-восстановительные реакции. В общем, в процессе окисления молекулы воды высвобождаются электроны с выделением кислорода и восстановлением диоксида углерода с образованием углеводов (Salisbury и Ross, 1994).
Фотосинтез важен для человека, среди прочего, потому что через него производится пища и кислород (Barcelo и др., 1983). Кроме того, он прямо или косвенно кормит почти весь живой мир на планете (Campbell и Reece, 2005).
Первые организмы-фотосинтезаторы, вероятно, появились 3 миллиарда или 3,5 миллиардов лет назад. По мере их распространения они устанавливали взаимозависимости и отношения, которые не только трансформировали их, но и изменили внешний облик планеты. Древние организмы жили в среде без свободного кислорода в атмосфере, в состав которой входил большой процент водорода. Кислород, даже был токсичным для этих примитивных организмов, так же, как в настоящее время для анаэробов. Энергия приобреталась такими процессами, как ферментация или гликолиз, что должно было привести к постепенному накоплению углекислого газа в ранней атмосфере. Если бы живые существа использовали сразу солнечную энергию, используя углекислый газ и выделяя кислород, они, безусловно, более эффективно использовали бы ресурсы своей окружающей среды, отметив веху в истории и эволюции жизни на планете. Условием фотосинтеза является поглощение фотонов, квантовых частиц определенными пигментами (Curtis и Barnes, 2001).
Фотосинтез представляет собой эндергонический процесс, потому что необходимо участие энергии солнечного излучения, чтобы начать цепные реакции, которые приводят к образованию органических соединений, которые накапливает организм.
Общая реакция фотосинтеза:

(Ch3O)n- это сокращенная форма представления крахмала или других углеводов по эмпирической формуле. Крахмал является наиболее распространенным продуктом фотосинтеза. Фотосинтез зависит от различных факторов окружающей среды, таких как интенсивность и качество освещения, влажность воздуха, температура, наличие воды, минеральных веществ и углекислого газа. Фотосинтез одного листа опирается на более, чем 50 индивидуальных реакций, каждая из которых зависят от параметров окружающей среды (Cogua, 2011).
Способность растений компенсировать воздействия окружающей среды зависит от их производительности и выживания в конкретной среде. Скорость фотосинтеза может измениться в течение следующих десятилетий в зависимости от приспособительных реакций к изменению уровней CO2, начиная от прямых реакций собственно на CO2, вплоть до косвенных реакций в связи с изменением температуры и водного режима, которые могут произойти в будущем (Cogua, 2011).
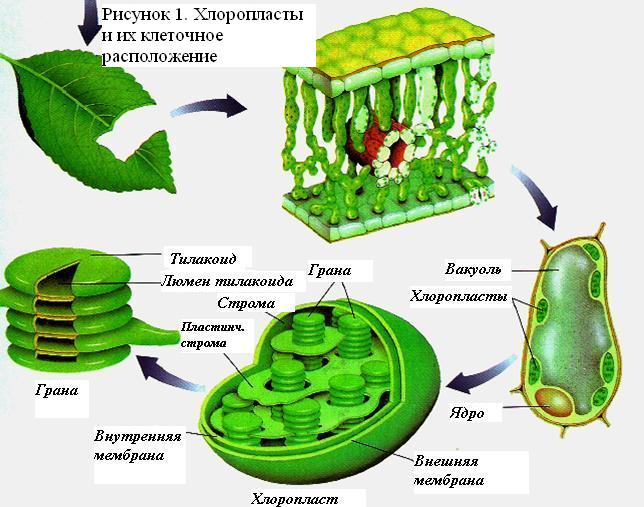
Хлоропласты
Клеточными органеллами, в которых происходит фотосинтез, являются хлоропласты.
Размер и форма хлоропластов изменчивы; они происходят из структур, известных как протопластиды (молодые хлоропласты), которые делятся по мере развития эмбриона. Хлоропласты окружены двойным набором мембран, контролирующих движение наружу и внутрь молекул. Внутренне они состоят из студенистого материала, богатого ферментами, называемого строма. Здесь происходит реакция превращения диоксида углерода в углеводы. Хлоропласты имеют пластинчатые мембраны и плоские закрытые «мешочки», похожие на пузырьки, называемые тилакоидами. Тилакоиды образуют стопки, называемые гранами, которые соединены между собой другими тилакоидами более вытянутой формы. В тилакоидных мембранах находятся хлорофиллы и другие пигменты, которые участвуют в поглощении света, ферменты для переноса электронов и фактор связи для образования АТФ (ATP). Основные пигменты, присутствующие в тилакоидных мембранах — хлорофилл a и хлорофилл b. В свою очередь, там найдены и другие пигменты, называемые каротинами и ксантофиллами (Salisbury и Росс, 1994).
Пигменты фотосинтеза
Чтобы световая энергия могла быть использована живыми существами, она должна быть поглощена фотосинтезирующими пигментами. Эти пигменты представляют собой вещества, способные поглощать свет на определенных длинах волн, например, хлорофилл – пигмент, который придает растениям характерный зеленый цвет, поглощает свет на длинах волн, соответствующих фиолетовому, синему и красному цвету, отражая зеленый цвет (Azcon и Talon, 1993).
Хлорофилл
Структура молекулы хлорофилла
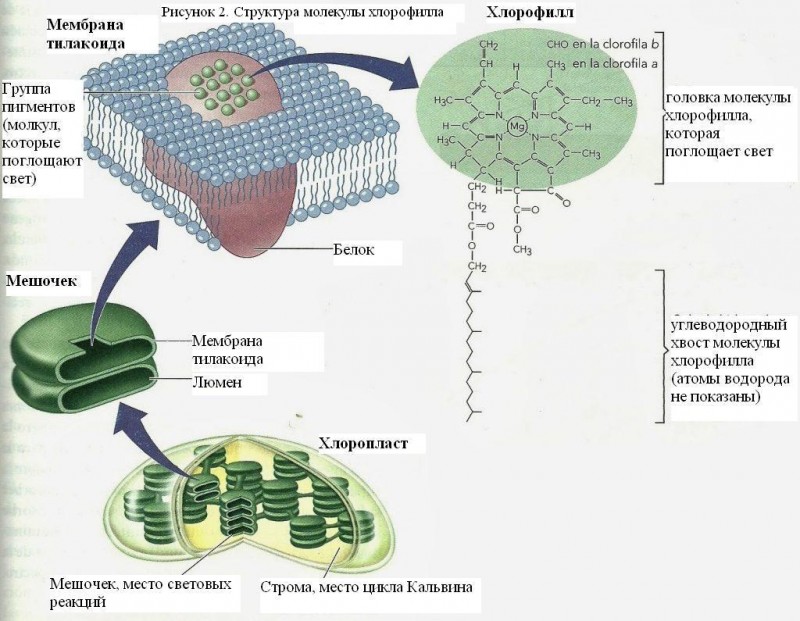
Хлорофилл является тетрапиррольным соединением, т.е. состоит из четырех пиррольных колец, соединенных метильными мостиками, которые образуют порфирин. Тетрапиррол является основной структурой порфирина, который, как и в хлорофилле, является частью гемоглобина и цитохрома.
В центре структуры находится металл (магний). Кольцо IV этерифицируется спиртом (фитолом), состоящим из серии из двадцати атомов углерода с двойной связью. Это «хвост» восковой природы, который приводит к тому, что пигмент является нерастворимым в воде, но растворим в некоторых органических растворителях. Отсюда и двойственная природа хлорофилла с центром гидрофильного порфирина и гидрофобным фитолом (Azcon и Talon, 1993).

Хлорофиллы a и b
Как упоминалось выше, хлорофилл a – это пигмент, непосредственно связанный с превращением световой энергии в химическую. Тем не менее, существует еще один тип хлорофилла, хлорофилл b, и пигменты, называемые каротиноидами, которые бывают красными, оранжевыми или желтыми, и которые в зеленых листьях маскируются обилием хлорофиллов. Эти дополнительные пигменты хлорофилла позволяют растениям поглощать более широкий диапазон спектра света, доступного для фотосинтеза, действуя в качестве рецепторов, которые передают энергию. Также существует хлорофилл с, найденный в бурых водорослях. В красных водорослях найден хлорофилл d. Наконец, существует тип хлорофилла (бактериохлорофилл), который является пигментом фототрофных бактерий (Cogua, 2011).
Каротиноиды
Желтые и красные пигменты, известные как каротиноиды, имеют систему сопряженных двойных связей, образованных атомами углерода, являются соединениями, нерастворимыми в воде, но растворимы в жировых растворителях. Они разделены на ненасыщенные углеводороды или каротины и кислородсодержащие производные, называемые ксантофиллами. В растениях каротиноид называется бета-каротином. Это пигменты красного, оранжевого или желтого цвета (Curtis и Barnes, 2001). Каротиноиды, участвующие в процессе фотосинтеза, называются первичными, в отличие от тех, которые находятся в цветах и плодах, формируя хромопласты, и в гетеротрофах, таких как бактерии, дрожжи и грибы (Cogua, 2011).
Организация пигментов в тилакоидах
Как отмечено выше, структурные единицы фотосинтеза у эукариот представляют собой хлоропласты, расположенные преимущественно в листьях. Внутренние ткани каждого листа покрыты эпидермальными клетками, покрытыми кутикулой, восковым слоем. Кислород и углекислый газ проникают в листья через специальные поры, называемые устьицами. Высокий процент фотосинтеза происходит в клетках палисадной паренхимы, которые расположены под эпидермисом и образуют мезофилл. Хлоропласты ориентированы внутри клеток, чтобы облегчить поглощение света тилакоидными мембранами (Curtis and Barnes, 2001).
Стадии фотосинтеза
Английский физиолог Ф. Блэкмэн (F. F. Blackman) после проведения различных экспериментов предположил, что в процессе фотосинтеза существует, по крайней мере, два ограничивающих фактора: интенсивность света и температура. Поэтому существует группа реакций, которые зависят от света, но не зависят от температуры, как видно из общей схемы фотосинтеза (Рисунок 3) (Curtis and Barnes, 2001).
В опытах Блэкмэна скорость реакций, которые зависят от температуры, увеличивалась лишь около 30°С. Затем скорость уменьшалась. На основании экспериментальных данных он сделал вывод, что эти реакции контролировались ферментами, так как это способ, по которому ферменты предположительно реагируют на температуру. Это способствовало дифференциации стадии световых реакций, зависящей от света и ферментативной стадии, не зависящей от света, или «темновых» реакций. Эти последние реакции нуждаются в химических веществах, синтезированных в «световых» реакциях, но они могут происходить и в светлых, и в темных условиях (Curtis и Barnes 2001).
Общая схема фотосинтеза

Поглощение света
Световые реакции можно резюмировать следующим образом. Светопоглощающие пигменты делятся на две группы: те, которые поглощают и передают энергию к реакционному центру, и те, которые образуют этот реакционный центр, представляющий собой определенный тип молекул хлорофилла (хлорофилл a P680 и Р700) и осуществляют фотохимическую реакцию.
Для обеспечения фотосинтеза требуется сотрудничество двух типов фотосистем, называемых I и II (PS I и PS II). В PS II вода используется в качестве донора электронов, выделяя кислород в качестве побочного продукта. В PS I восстанавливается терминальный акцептор, отдавая электрон ферредоксину, чтобы с его помощью восстановить НАДФ (NADP), которая, наконец, используется для превращения CO2 в углеводы (Melgarejo, 2010).
Фотосистемы I и II работают вместе, хотя в то же время фотосистема I может работать независимо, как это описано на рисунке.

В данной модели энергия света достигает фотосистемы II, где она захватывается молекулой хлорофилла a P680. Электрон молекулы Р680 высвобождается на более высоком энергетическом уровне, из которого он передается к молекуле первичного акцептора электронов. Электрон проходит вниз на фотосистему I вдоль цепи переноса электронов. В результате потока электронов вдоль транспортной цепи устанавливается протонный градиент по всей тилакоидной мембране; потенциальная энергия этого электрохимического градиента используется для формирования АТФ (ATP) из АДФ (ADP) – хемиосмотического процесса, аналогичного процессу у митохондрий. Этот процесс известен как фотофосфорилирование (Curtis и Barnes, 2001).
Одновременно выделяются три других действия:
1. Молекула хлорофилла P680, теряя свой электрон, ищет замену. Она находит ее в молекуле воды, которая, будучи прикрепленной к молекуле, содержащей марганец, лишается электрона, а затем распадается на протоны и газообразный кислород.
2. Дополнительно световая энергия захватывается в реактивной молекуле хлорофилла Р700 фотосистемы I. Молекула окисляется, и электрон высвобождается в первичный акцептор электронов, из которого он перемещается вниз по направлению к НАДФ+ (NADP+).
3. Электрон, удаленный из молекулы P700 фотосистемы I, замещается электроном, который перемещался вниз от первичного акцептора электронов фотосистемы II. Таким образом, на свету происходит непрерывный поток электронов от воды к фотосистеме II, к фотосистеме I, к НАДФ+ (CURTIS, Helena, BARNES, Sue, 2001). Энергия, восстановленная на этих этапах, представлена молекулой АТФ (ATP) (образование которой высвобождает молекулу воды) и НАДФH (NADPH), которые затем преобразуются в основные источники энергии для восстановления углекислого газа. Для генерации одной молекулы НАДФН необходимо поглотить 4 протона: 2 фотосистемой II и 2 фотосистемой I (Curtis и Barnes, 2001).
Восстановление и фиксация CO2
На второй стадии фотосинтеза энергия используется для восстановления углерода. Углерод доступен для фотосинтезирующих клеток в форме диоксида углерода. У растений двуокись углерода достигает фотосинтезирующих клеток через специализированные отверстия листьев и зеленых стеблей, называемые устьицами (Curtis и Barnes 2001).
На этой второй стадии фотосинтеза, АТФ и НАДФН, сформированные на первой стадии, используются для восстановления углерода из диоксида углерода до простого сахара. Затем химическая энергия, временно хранящаяся в молекулах АТФ и НАДФН, передается соответствующим молекулам для транспортировки и хранения энергии в клетках водорослей или органов растений. Результатом этого процесса является формирование углеродного скелета, из которого затем могут быть построены и другие органические молекулы (Cogua, 2011).
Реакции фиксации углерода проходят в условиях, которые не требуют света, но некоторые ферменты регулируются им и производятся в строме посредством цикла Кальвина (Рисунок 5). НАДФ и АТФ, образованные в реакциях, которые поглощают световую энергию, используются для восстановления углекислого газа. Цикл производит глицеральдегид фосфат, из которого может быть сформирована глюкоза и другие органические соединения (Cogua, 2011).
Цикл Кальвина

Это преобразование происходит в строме хлоропластов. CO2 поступает в хлоропласт, проходя через клеточные мембраны органеллы по определенным каналам. Этот газ поступает в тело растения, благодаря устьичному отверстию, которое в то же время облегчает выделение водяного пара в процессе, известном как транспирация (Cogua, 2011). Цикл Кальвина представляет собой центральный путь восстановления СО2 до органических молекул. Он начинается, когда фермент рибулоза 1,5 бифосфат карбоксилаза оксигеназа (Rubisco) выполняет карбоксилирование СО2 и дает начало двум молекулам из трех атомов углерода (Melgarejo, 2010). Чтобы выяснить это, было проведено несколько экспериментов, которые были успешными, благодаря использованию радиоизотопов в зеленых водорослях Chlorella pyrenoidosa и Scenedesmus obliquus. После получения 14СО2 в течение 60 секунд в культурах вышеупомянутых водорослей появляется несколько меченых соединений; фосфаты сахаров, органические кислоты и аминокислоты. Сокращая время применения С14О2 шаг за шагом, удалось восстановить образованные меченые продукты. Менее чем за 2 секунды появляется только одно меченое соединение, 3-фосфоглицериновая кислота (3-PGA), состоящая из трех атомов углерода. Таким образом, первая стадия восстановления СО2 состоит в карбоксилировании, в ходе которого СО2 включается в молекулу акцептора, имеющую 5 атомов углерода, рибулозо-1,5-дифосфат, расщепление которого приводит к образованию двух молекул 3-PGA. Механизм восстановления СО2 является циклическим процессом из нескольких частичных ферментативных стадий (Мельгарехо, 2010).
Метаболизм Хэтча/Слэка

Существуют и другие адаптивные процессы у различных типов растений, например, некоторые растения присоединяют диоксид углерода к первому соединению, известному как фосфоенолпировиноградная кислота или фосфоенолпируват (РЕР) с образованием соединения из четырех атомов углерода, щавелевоуксусной кислоты (промежуточное соединение в цикле Кребса). Среди других растений, которые имеют этот тип метаболизма, кукуруза и сорго. У них диоксид углерода, включенный в щавелевоуксусную кислоту, окончательно переносится в RuBP и входит в цикл Кальвина, но только после прохождения ряда реакций, которые транспортируют его в более глубокие участки внутри листа. Эти реакции протекают в клетках мезофилла, где хлоропласты характеризуются наличием обширной сети тилакоидов, организованных в хорошо развитые граны. Яблочная или аспарагиновая кислота, в зависимости от вида, транспортируются к клеткам обкладки сосудистого пучка, благодаря сети плазмодесм. Хлоропласты этих клеток, которые образуют плотные оболочки вокруг сосудистых пучков листа, имеют неразвитые граны и часто содержат граны крахмала. Растения, использующие этот путь, или путь Хэтча-Слэка, обычно называют C4-растениями, благодаря 4-углеродному соединению, которое служит в качестве акцептора СО2 (Cogua, 2011).
Кислотный метаболизм Толстянковых или растения MAC (CAM)

Существуют некоторые виды растений сухого климата, которые имеют толстые листы, с низким соотношением поверхности к объему, толстой кутикулой и низкой скоростью транспирации. Такие растения называются суккулентами. Как правило, фотосинтезирующие клетки листьев или стебля у них представлены в губчатом мезофилле. У некоторых суккулентов метаболизм CO2 необычен, первоначально исследован у членов семейства Толстянковых, поэтому его называют Кислотный метаболизм Толстянковых (Crasulaceae Acid Metabolism (CAM или MAC) по-английски).
Новый фотосинтетический путь, упомянутый выше, был найден у некоторых растений суккулентов или полусуккулентов из родов Бриофи́ллум, Каланхоэ, Седум, Клейния, Толстянка, Опунция (Salisbury и Ross, 1994). MAC или CAM растения поглощают диоксид углерода в течение ночи, фиксируя в органических кислотах, которые подкисляют среду и высвобождают его в течение дня, чтобы сразу же использовать в цикле Кальвина. Этот тип метаболизма уменьшает потери воды, связанные с открытием устьиц. (Cogua, 2011).
Факторы окружающей среды и фотосинтез
Подсчитано, что количество углерода, который фиксируется каждый год на планете, колеблется от 70000 до 120000 млн. тонн или эквивалентно около 170 – 290 гигатоннам сухого вещества, с минимальной формулой 0. Эта огромная производительность имеет место, несмотря на низкую концентрацию в атмосфере. Более 13% атмосферного углерода используется каждый год в процессе фотосинтеза, и почти равное количество обменивается при растворении в океанах. На фотосинтез оказывают влияние такие факторы, как вода, концентрация СО2, свет, питательные вещества и температура, а также возраст и генетика растений (Salisbury и Ross, 1994).
CO2
Количество СО2 является фактором, определяющим производительность, тем не менее некоторые реакции фотосинтеза могут быть осуществлены в его отсутствие. Однако, без этого газа не будет синтеза углеводов. Концентрация CO2 в атмосфере не является оптимальной для фотосинтеза, в сельскохозяйственной практике используется искусственное добавление газообразного СО2 в условиях постоянного освещения, для увеличения фотосинтетической скорости и урожайности при производстве биологического материала (Salisbury и Ross, 1994). CO2, как известно, производит немедленное увеличение скорости фотосинтеза, особенно у C3-растений. Однако, когда растения растут непрерывно с повышенным уровнем СО2, происходят биохимические изменения, которые снижают фотосинтетическую способность листьев, поэтому большой начальный рост фотосинтеза при высокой концентрации СО2 обычно не сохраняется таким же высоким, когда проходят несколько недель или месяцев. Это явление известно как акклиматизация фотосинтеза (Cogua, 2011).
Вода
Внеклеточно у высших растений вода транспортирует такие материалы, как соли, которые перемещаются от корней ко всем частям растения. Помимо того, что она является сырьем в процессе фотосинтеза, вода способствует в качестве реагента нескольким метаболическим реакциям. Ионы, образующие молекулу воды (Н) и (ОН) собираются и рекомбинируются в новые молекулы воды (Cogua, 2011).
Свет
Нет света, нет фотосинтеза, растения нуждаются в свете с точки зрения интенсивности и качества излучения. С увеличением интенсивности света увеличивается фотосинтетическая интенсивность вначале по линейной зависимости, а затем плавно уменьшается и, наконец, достигает постоянного значения, т.е. фотосинтетическая способность насыщается светом. Это значение насыщения достигается различными видами с разной скоростью. У светолюбивых растений это происходит после достижения высокой интенсивности излучения, а у тенелюбивых растений это насыщение достигается быстро, то есть требуется низкая интенсивность света (Cogua, 2011).
Температура
Зависимость фотосинтеза от факторов света и температуры позволяет сделать вывод, что фотосинтез не является непрерывным процессом, а состоит из множества фотохимических реакций, которые зависят от света и серии ферментативных реакций, зависящих от температуры. Последние очевидны в состоянии насыщения света, точке, в которой повышение температуры увеличивает фотосинтетическую интенсивность (Cogua, 2011). Транспорт, промышленность, вырубка лесов, сельское хозяйство и другие виды деятельности человека вызывают увеличение концентрации CO2 и других газов в атмосфере, таких как метан. Скопление этих газов имеет тенденцию к нагреву атмосферы, что может привести к региональным или глобальным изменениям, которые повлияют на такие параметры, как температура, осадки, влажность почвы и уровень моря, все они связаны с фотосинтезом и, следовательно, с самой жизнью на планете. (Cogua, 2011).
Инга Костенко, Mivena,Украина
Анна Устименко, Клуб Sirius Agro Plant
Universidad Nacional de Colombia
siriusap.com










