
Фотосинтез. Освещенность и продолжительность. Фотосинтез для растений
Конспект урока "Фотосинтез растений"
Тема: Фотосинтез растений.
Цель: изучить сущность процесса фотосинтеза, показать его важность для жизни на Земле.
Задачи:
1.Выявить приспособления растений к процессу фотосинтеза и необходимые условия для его протекания, научиться определять продукты фотосинтеза, выяснить значимость фотосинтеза для жизни на Земле.
2.Продолжить формирование экспериментальных умений учащихся, навыков наблюдения, прогнозирования, умения делать выводы и объяснять результаты своих наблюдений.
3.Воспитывать уважение к труду других людей, культуру учебного труда, чувство патриотизма на примере трудов К.Тимирязева, развивать познавательный интерес к предмету.
Оборудование: табл. «Строение растительной клетки» и «Строение хлоропласта»; компьютер, проектор, экран, живые экземпляры растений, спирт, соляная кислота, пипетки, покровные стекла, иглы, ножницы, микроскоп, вода, йод, пробирки, ацетат свинца, фильтровальная бумага, пинцет, ступка с пестиком,
Ход урока
1.Орг.момент(постановка цели и задач урока)
«Дайте самому лучшему повару сколько угодно свежего воздуха, солнечного света и целую речку чистой воды и попросите, чтобы из всего этого он приготовил вам сахар, крахмал, жиры и зерно – он решит, что вы над ним смеётесь». – писал К.Тимирязев.
Как вы думаете, что он имел в виду?
Голландский естествоиспытатель Ян Баптист Ван-Гельмонт не только занимался врачебной практикой, но и экспериментировал с растениями. Он решил узнать, как питается растение.
«Как лишенное рта растение питается? Как поступают в его организм питательные вещества?»
Сегодня на уроке мы должны ответить на следующий вопрос: Можно ли считать фотосинтез гениальным изобретением природы?
Нам следует решить ряд ситуационных задач.
В чем сущность процесса фотосинтеза.
Какие приспособления имеют растения для протекания данного процесса?
Можно ли управлять процессом фотосинтеза?
Может ли человек извлечь пользу для себя из данного процесса?
Могут ли растения питаться другими способами?
Могут ли животные питаться с помощью фотосинтеза?
Какова роль фотосинтеза для жизни на Земле?
Работаем в группах. Каждой группе выдается кейс. Каждая группа получает свою задачу для решения.
кейс №1. Какие приспособления имеют растения для фотосинтеза?
Ответы учащихся
1. Лабораторная работа. Рассмотреть внешнее строение листьев нескольких комнатных растений.
Вывод: Растения имеют плоскую листовую пластинку, черешок для прикрепления листа к побегу, в основном имеют зеленую окраску.
2. Опыт. Рассмотреть положение листьев комнатного растения по направлению к окну. Развернуть растение в противоположную сторону.
Наблюдение. Через два дня листья растения были вновь направлены в строну окна.
Вывод: Черешок служит для поворачивания листовой пластинки к свету.
Лабораторная работа. Рассмотреть порядок расположения листьев на нескольких комнатных растениях.
Наблюдение. Растения имеют большое количество листьев, расположенных на побеге мозаично, не затеняя друг друга.
Вывод: Листья растения располагаются таким образом, чтобы как можно больше света попадало на листовые пластинки.
3.Лабораторная работа. Приготовить и рассмотреть под микроскопом микропрепарат кожицы листа лука, амариллиса и др.
Вывод: Плотное расположение клеток кожицы листа предохраняет лист от механических повреждений, прозрачные, неокрашенные клетки способствуют проникновению света, а наличие устьиц обеспечивает газообмен.
4.Лабораторная работа. Рассмотреть под микроскопом микропрепарат листа камелии.
Вывод: Клетки мякоти листа ярко – зеленые, т. к. содержат зеленые пластиды – хлоропласты; расположены рыхло, между ними есть пространства, заполненные воздухом.
5.Опыт. Измельчить листья зеленого растения, поместить в колбу, влить спирт и осторожно нагреть на спиртовке.
Наблюдение. Спирт окрасился в изумрудно –зеленый цвет.
Вывод: Листья растений содержат пигмент хлорофилл, который придает
зеленый цвет растениям.
Предположение: окраска хлорофилла
определяется наличием в нем атома магния
Опыт. В пробирку с вытяжкой хлорофилла добавим несколько капель соляной кислоты – атом водорода заместит атом магния и окраска измениться на оливково – бурую.В ту же пробирку добавим небольшое количество ацетата меди и подогреем содержимое на спиртовке – атом водорода заместится на атом меди и окраска вновь станет зеленой.
Вывод: Зеленая окраска хлорофилла определяется наличием в нем атома металла, вне зависимости от того, будет ли это магний или другой металл.
6.Опыт. С обильно политого растения пестролистной герани или бегонии, стоящей на свету срезают листочек,
опускают в кипящую воду, а затем в стакан с горячим спиртом для обесцвечивания. Обесцвеченный лист обливают раствором йода.
Наблюдение. Не весь лист окрасился в синий цвет.
Вывод: Только зеленые пластиды принимают участие в процессе фотосинтеза.
Какие приспособления имеют растения для фотосинтеза?
Выводы:
множество листьев с плоской поверхностью;
черешок для поворачивания листьев к свету;
мозаичное расположение листьев;
прозрачные, неокрашенные клетки кожицы листа для проникновения света;
устьица, обеспечивающие газообмен;
особые пластиды хлоропласты, содержащие зеленый пигмент хлорофилл, способный улавливать солнечный свет.
Как, в каком направлении шла эволюция растений?
В сторону образования черешковых листьев, зеленой окраски, улавливающих самый короткий спектр света.
Кейс №2. Какие условия необходимы для фотосинтеза?
Ответы учащихся:
1. Опыт. Два растения герани обильно поливают, одно из них ставят в темное место, а другое оставляют на свету. Через трое суток срезают по одному листочку с каждого растения, опускают в кипящую воду, а затем в стакан с горячим спиртом для обесцвечивания. Обесцвеченные листья и дольку картофеля обливают раствором йода.
Наблюдение. Лист с растения из темного места остался светлым, а другой и долька картофеля окрасились в синий цвет.
Вывод: При помощи солнечного света в клетках растений образуется крахмал.
2. Опыт. Одно из растений герани обильно поливают, а другое оставляют сухим, оставляют оба растения на свету. Через трое суток срезают по одному листочку с каждого растения, опускают в кипящую воду, а затем в стакан с
горячим спиртом для обесцвечивания. Обесцвеченные листья и дольку картофеля обливают раствором йода.
Наблюдение. Листья политого растения имеют более интенсивную синюю окраску.
Вывод: Вода необходима для образования крахмала и выделения кислорода.
Опыт. В две банки из светлого стекла помещают по 5-6 веточек герани, доливают немного воды, затем опускают зажженные свечи укрепленные на проволоке и закрывают. Когда свечи погаснут, их вынимают. Одну банку ставят на свет, а другую в темное место. На следующий день банки открывают и вносят зажженные свечи.
Наблюдение. В банке, стоящей на свету свеча горит, а в другой гаснет.
Вывод: растения на свету в процессе фотосинтеза поглощают углекислый газ и выделяют кислород.
Какие условия необходимы для фотосинтеза?
Выводы:
Углекислый газ, вода, солнечный свет.
Кейс №3. Какие продукты образуются в процессе фотосинтеза?
Ответы учащихся: крахмал и кислород.
Кейс №4 В чем же сущность процесса фотосинтеза?
Организация фотосинтетического аппарата листа. Активность первичных реакций фотосинтеза и их регулирование.
По современным представлениям процесс фотосинтеза включает следующие этапы: фотофизический, фотохимический (световой) и ферментативный (темновой).
Фотофизический этап фотосинтеза. Первоначальный акт фотосинтеза у всех фото синтезирующих организмов связан с поглощением квантов (фотонов) света. Свет имеет двойственную, корпускулярную и волновую природу, т.е. он представляет поток частиц с разной энергией (фотонов) и электромагнитных волн. У высших растений в поглощении света участвуют три группы пигментов — хлорофилл а и b и каротиноиды. Полагают, что хлорофилл включается в общую последовательность фотосинтетических реакций в двух участках и что в действительности существует две фотосистемы (ФС I и ФС II).
Фотохимический (световой) этап фотосинтеза.
Так как весь кислород фотосинтеза выделяется из воды, общее уравнение фотосинтеза имеет следующий вид:
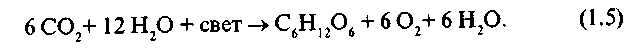 Фотосинтез представляет собой окислительно-восстановительный процесс, в котором вода окисляется до О2 ,а углекислый газ восстанавливается до углеводов.
Фотосинтез представляет собой окислительно-восстановительный процесс, в котором вода окисляется до О2 ,а углекислый газ восстанавливается до углеводов.
Сущность происходящих реакций фотосинтеза на свету была выяснена в 50-х годах американским физиологом и биохимиком растений Д.И. Арноном. Им было высказано предположение о том, что в процессе фотосинтеза происходит не фотолиз воды (разрыв молекулы Н-О-Н на два радикала Η и ОН), требующий большого количества энергии (110 ккал/моль), а фотоокисление, т.е. мобилизация электрона. В других опытах, проведенных в лаборатории Д.И. Арнона, было показано, что выделенные из листьев шпината хлоропласты под действием света способны восстанавливать НАДФ (никотинамидадениндинуклеотидфосфат окисленный) и фосфорилировать АДФ (аденозиндифосфат). Эти процессы протекают в тилакоидах хлоропластов. Полученный из стромы хлоропластов гомогенат, лишенный мембран, ассимилировал СО2, если в реакционной среде присутствовали НАДФ.Η (никотинамидадениндинук-леотидфосфат восстановленный) и АТФ (аденозинтрифосфат):

В этих опытах было открыто явление фотофосфорилирования и показано, что НАДФ.Н и АТФ — конечные продукты световой фазы фотосинтеза, а также установлено протекание световой фазы в тилакоидах хлоропласта, а темновой — в его строме.
Различают циклическое и нециклическое фотофосфорилирование, соответственно двум типам потока электронов. При циклическом потоке электроны, переданные от молекулы хлорофилла первичному акцептору, возвращаются к ней обратно, а при нециклическом — происходит фотоокисление воды и передача электрона от воды к НАДФ. Энергия, выделяемая в ходе этой окислительно-восстановительной реакции, частично используется на синтез АТФ.
Оба типа фотофосфорилирования принято включать в световую фазу фотосинтеза, однако свет необходим только на первых этапах этих процессов (перенос электрона по цепи переносчиков может происходить в темноте). В зависимости от состояния хлоропластов расположение и набор переносчиков в электрон-транспортной цепи (ЭТЦ) могут меняться (Якушкина, 1993).
Ферментативный (темповой) этап фотосинтеза. Сущность темновых реакций процесса фотосинтеза была раскрыта в исследованиях М. Кальвина. Он установил, что в присутствии СО2 рибулезодифосфат (РДФ) в темноте используется для образования фосфоглицериновой кислоты (ФГК), дальнейшее превращение которой требует света. На основании полученных данных предложена следующая схема процесса фотосинтеза:
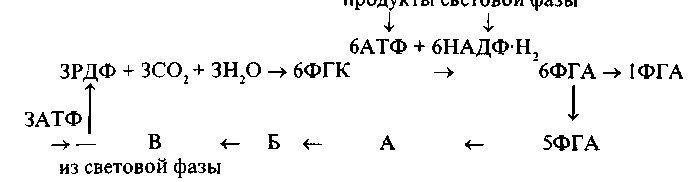
На схеме видно, что РДФ является акцептором, который присоединяет CO2 с образованием фосфоглицериновой кислоты. В отсутствие света РДФ быстро используется, при этом накапливается определенное количество ФГК. На свету при участии продуктов световой фазы ФГК восстанавливается до фосфоглицеринового альдегида (ФГА). Частично ФГА путем ряда превращений используется на регенерацию акцептора (РДФ). Для регенерации РДФ также используете АТФ, образующаяся на световой фазе, что обеспечивает поддержание количества РДФ на свету на постоянном уровне. Таким образом, в каждом цикле принимают участие 3 молекулы РДФ и образуется 6 молекул триозы (ФГА). Из них 5 молекул ФГА идет на регенерацию акцептора через ряд промежуточных продуктов, обозначенных на схеме А, Б, В. Как показано на схеме, каждая 6-я молекула ФГА выходит из цикла и используется для построения углеводов.
Следовательно, темновые реакции фотосинтеза представляют разветвленный цикл, включающий три взаимосвязанные фазы: карбоксилирование — превращение рибулезофосфата в рибулезодифосфат, являющийся акцептором СО , и образование двух молекул ФГК; восстановление ФГК в ФГА и регенерация РДФ. Экспериментально установлено, что характер превращений ФГК зависит от специфических особенностей обмена веществ отдельных видов растений, условий их корневого питания, интенсивности света и его спектрального состава и т.п. (Рубин, 1979).
Кейс №5 Можно ли управлять процессом фотосинтеза?
Домашний отложенный эксперимент.
1.Условия минерального питания
Опыт: В один ящик с богато удобренной почвой , а в другой с обедненной почвой высадим рассаду томатов и оставим на две недели. В течение этого срока будем поливать первый ящик обильно, а второй как обычно.
Наблюдение: Во втором ящике у томатов листовая пластинка уже и бледнее.
Вывод: Для ускорения процесса фотосинтеза необходимо хорошее минеральное питание и достаточная влажность
2. Опыт: Одно из растений томатов оставить в классе на подоконнике, а другое, внешне одинаковое, поместить под полиэтиленовый колпак и оставить на две недели. В течение этого времени периодически сжигать опилки под колпаком для увеличения концентрации СО2.
Наблюдение: Томаты, растущие под колпаком развиваются лучше.
Вывод: Для увеличения продуктивности растений в теплицах необходимо повышенное содержание углекислого газа.
Для решения этой проблемы на открытых полях—активизация деятельности почвенных микроорганизмов путем внесения органических удобрений.
3.Оптимизация посадки
Опыт: В один ящик высадим рассаду томатов густо, а в другой на оптимальном расстоянии и оставим на две недели.
Наблюдение: Во втором ящике растения развиваются лучше.
Вывод: Необходимо высаживать растения на оптимальном расстоянии для лучшего обеспечения солнечным светом.
Солнечный свет—состоит из нескольких волн, окрашенных в цвета радуги. Ученые провели исследование и выяснили в каких лучах спектра фотосинтез идет интенсивнее:
Вывод: В процессе эволюции растения приспособились к поглощению именно тех лучей солнечного спектра, энергия которых наиболее полно используется в ходе фотосинтеза.
Кейс №6 Какое значение имеет фотосинтез для Земли?
Нам всегда говорят, что зеленые растения очищают воздух. Но так ли это? Интересно, что различные породы деревьев способны поглощать различное количество углекислого газа:
Зная это, можно выбирать растения для озеленения городов. А что же с выделением кислорода? Ученые подсчитали: 1 га кукурузных посевов выделяет в год 15 тонн кислорода, что достаточно для дыхания 30 человек, а дерево средней величины обеспечивает трех человек. Однако , не следует забывать, что дыхание, процесс, обратный фотосинтезу. Таким образом, сколько кислорода выделилось при фотосинтезе, столько же его израсходовалось при дыхании. Кроме того, кислород расходуется и животными для дыхания, при горении и гниении. Оказывается ,основное возмещение кислорода происходит за счет деятельности фитопланктона, так как в глубоких водоемах отмершие
организмы оседают глубоко на дно и разлагаются там без доступа кислорода. Поэтому, в водоемах доля кислорода значительно больше, чем в атмосфере.
Сегодня много говорят и пишут о абиогенном происхождении кислорода атмосферы, исключающим участие живых организмов в этом процессе. Рассмотрим долю различных источников поступления кислорода в атмосферу Земли.
Известно, что кислород выделяется в атмосферу в процессе фотосинтеза. Но где же первые растения брали кислород для дыхания?
Оказывается, еще в очень древних геологических слоях Земли обнаружены сине -зеленые водоросли(сейчас их чаще называют цианобатериями) , которые не обладали способностью дышать, а механизм распада органических веществ у них напоминал процесс брожения. Таким образом, сине-зеленые водоросли имея способность к фотосинтезу и не используя кислород для дыхания, явились его накопителями. Накопление кислорода на нашей планете стало мощным стимулом для появления принципиально новых организмов, способных существовать в различных условиях.
Могут ли растения питаться другим способом?
Рассказ о растениях- хищниках.
Итоги урока:
1.Назовите приспособления растений к процессу фотосинтеза.
2.Укажите условия, необходимые для процесса фотосинтеза.
3.Назовите продукты фотосинтеза.
4.В чем заключается сущность процесса фотосинтеза?
5.Каково значение процесса фотосинтеза для Земли?
Необходимо составить каждой группе синквейн по итогам урока.
Рефлексия: О фотосинтезе можно говорить не только на уроках биологии и химии. Если по-настоящему любить природу, можно описать этот процесс красивым литературным языком. Послушайте выдержку из работы К.А. Тимирязева (предложить учащимся закрыть глаза и мысленно нарисовать картинку к тексту).
Когда-то, где-то на Землю упал луч Солнца, но он упал не на бесплодную почву, он упал на зеленую былинку пшеничного ростка, или, лучше сказать, на хлорофилловое зерно. Ударяясь о него, он потух, перестал быть светом, но не исчез. В той или другой форме он вошел в состав хлеба, который послужил нам пищей. Он преобразился в наши мускулы, в наши нервные клетки. Этот луч Солнца согревает нас. Он приводит нас в движение. Быть может, в эту минуту он играет в нашем мозгу. К. Тимирязев.
Домашнее задание:
1.Составить кроссворд
Список использованной литературы
1. В.Б.Захаров, С.Г. Мамонтов, Н.И. Сонин. Общая биология 10-11 кл. – М.: «Дрофа» 2008
2. Н.В. Чебышев . Биология.Пособие для поступающих в ВУЗы - М.: «Новая волна». 2010.
3.Л.А. Николаев . Химия жизни. – М.: «Просвещение» 1973.
4.В.И. Артамонов. Занимательная физиология растений. – М.: «Агропромиздат» 1991.
infourok.ru
Биология для студентов - 01. Современные представления о фотосинтезе растений. Фотосинтез и урожай
Фотосинтез - это биологический процесс, в ходе которого энергия электромагнитного излучения превращается в химическую энергию органических соединений. Сначала свет поглощается молекулами пигментов в светособирающей антенне, затем происходит перенос энергии возбуждения к реакционному центру (особому связанному с мембраной молекулярному комплексу), который содержит хлорофилл или бактериохлорофилл. Именно в реакционном центре происходит первичная фотохимическая реакция — перенос электрона от первичного донора, обозначаемого символами D или Р, к акцептору А. Энергию света, запасаемую при разделении зарядов, растение использует для осуществления реакций электронного транспорта, которые дают энергию для синтеза устойчивых высокоэнергетических соединений (АТФ, НАДФН+Н, углеводов).
В процессе фотосинтеза на Земле первично создаются органические вещества, используемые в дальнейшем в качестве пищи, кормов, горючего, промышленного сырья и т. д. Фотосинтез включен в глобальный газообмен на планете, обеспечивая необходимый для жизни уровень кислорода, а также необходимый для биосферы в целом уровень углекислого газа. Очевидно, что фотосинтез с момента своего возникновения играет ключевую роль в эволюции биосферы. Определяющими состояние биосферы параметрами являются количество запасенного органического вещества (валовая первичная продукция), количество выделившегося кислорода, балансовый уровень углекислого газа в атмосфере (глобальная температура, глобальный климат). Глобальная модель климата показывает, что после мелового периода уровень CO2 снизился от 2800 ppm до 330-360 ppm. Падение концентрации CO2 привело к снижению эффективности С3 фотосинтеза и усилению фотодыхания, в ходе которого может «теряться» до 50 % ассимилированного углерода. Это послужило эволюционным стимулом для создания CO2 - концентрирующего механизма. Первыми проблему увеличения концентрации CO2 в клеточном пространстве, решили цианобактерии и водоросли.
Хлоропласт. Эти органоиды содержатся в клетках листьев и других зеленых органов растений, а также у разнообразных водорослей. Размеры хлоропластов 4-6 мкм, наиболее часто они имеют овальную форму. Xлоропласт - основной органоид клеток растений, в котором происходит фотосинтез.

По строению хлоропласты сходны с митохондриями. От цитоплазмы хлоропласт отграничен двумя мембранами - наружной и внутренней. Наружная мембрана гладкая, без складок и выростов, а внутренняя образует много складчатых выростов, направленных внутрь хлоропласта. Поэтому внутри хлоропласта сосредоточено большое количество мембран, образующих особые структуры - граны. Они сложены наподобие стопки монет. В мембранах гран располагаются молекулы хлорофилла, потому именно здесь происходит фотосинтез. В хлоропластах синтезируется и АТФ. Между внутренними мембранами хлоропласта содержатся ДНК, РНК. и рибосомы. Следовательно, в хлоропластах, так же как и в митохондриях, происходит синтез белка, необходимого для деятельности этих органоидов. Хлоропласты размножаются делением.
Хлорофилл в хлоропластах появляется в процессе зеленения пластид. У высших растений и водорослей встречаются хлорофиллы, a, b, с, d, e. Только хлорофилл а присутствует во всех перечисленных группах. Эмпирическая формула хлорофилла а C55H7205N4Mg.

По химической природе молекула хлорофилла состоит из порфиринового кольца (тетрапиррола) в составе дикарбоновой кислоты - хлорофиллина, этерифицированной остатком метилового спирта и высокомолекулярного одноатомного спирта - фитола.
Ядро хлорофилла обладает гидрофильными свойствами, остаток фитола - гидрофобными свойствами. Это позволяет молекуле хлорофилла взаимодействовать как с белками, так и с липидами. Хлорофиллы легкорастворимы в ацетоне, серном эфире, этаноле, метаноле, сероуглероде, бензоле, плохо растворимы в петролейном эфире.
Представления о существовании фотосинтетических единиц сформировались на основе работ Эмерсона и Арнольда (1932). Ими впервые было показано, что в выделении одной молекулы кислорода в процессе фотосинтеза участвуют от 2000 до 2500 молекул хлорофилла. Эта величина и получила название «фотосинтетическая единица».
Фотосинтез начинается с того, что излучаемые солнцем фотоны попадают в особые пигментные молекулы, находящиеся в листе, — молекулы хлорофилла. Хлорофилл содержится в клетках листа, в мембранах клеточных органелл хлоропластов (именно они придают листу зеленую окраску). Процесс улавливания энергии состоит из двух этапов и осуществляется в раздельных кластерах молекул — эти кластеры принято называть Фотосистемой I и Фотосистемой II.

Номера кластеров отражают порядок, в котором эти процессы были открыты, и это одна из забавных научных странностей, поскольку в листе сначала происходят реакции в Фотосистеме II, и лишь затем — в Фотосистеме I. Когда фотон сталкивается с 250-400 молекулами Фотосистемы II, энергия скачкообразно возрастает и передается на молекулу хлорофилла.
В этот момент происходят две химические реакции:
- молекула хлорофилла теряет два электрона (которые принимает другая молекула, называемая акцептором электронов)
- расщепляется молекула воды.
Электроны двух атомов водорода, входивших в молекулу воды, возмещают два потерянных хлорофиллом электрона. После этого высокоэнергетический («быстрый») электрон перекидывают друг другу, как горячую картофелину, собранные в цепочку молекулярные переносчики. При этом часть энергии идет на образование молекулы аденозинтрифосфата (АТФ), одного из основных переносчиков энергии в клетке. Тем временем немного другая молекула хлорофилла Фотосистемы I поглощает энергию фотона и отдает электрон другой молекуле-акцептору. Этот электрон замещается в хлорофилле электроном, прибывшим по цепи переносчиков из Фотосистемы II. Энергия электрона из Фотосистемы I и ионы водорода, образовавшиеся ранее при расщеплении молекулы воды, идут на образование НАДФ-Н, другой молекулы-переносчика. В результате процесса улавливания света энергия двух фотонов запасается в молекулах, используемых клеткой для осуществления реакций, и дополнительно образуется одна молекула кислорода.

Первая стадия фотосинтеза протекает на свету. Световые кванты дают электронам энергию, необходимую для переноса их от хлорофилла или другого фотосинтезирующего пигмента. В ходе первой стадии из АДФ (аденозиндифосфата) и фосфата синтезируется АТФ, а НАДФ (никотинамидадениндинуклеотидфосфат) восстанавливается до НАДФ·h3. Синтез АТФ за счёт энергии световых квантов называется фотофосфорилированием. Этот процесс может быть циклическим (в реакции «работают» одни и те же электроны) и нециклическим (электроны в конце концов доходят до НАДФ и, взаимодействуя с ионами водорода, образуют НАДФ·h3). Кислород как побочный продукт реакции выделяется только во втором случае.
Для реакций второй стадии свет не нужен. Восстановление CO2 происходит за счет энергии АТФ и накопленного НАДФ·h3. Углекислый газ связывается с пятиуглеродным сахаром рибулозобисфосфатом, образуя две молекулы трёхуглеродной фосфоглицериновой кислоты (ФГК). Такой процесс получил название C3-фотосинтеза.
После того как солнечная энергия поглощена и запасена, наступает очередь образования углеводов. Основной механизм синтеза углеводов в растениях был открыт Мелвином Кальвином, проделавшим в 1940-е годы серию экспериментов, ставших уже классическими. Калвин и его сотрудники выращивали водоросль в присутствии углекислого газа, содержащего радиоактивный углерод-14. Им удалось установить химические реакции темновой фазы, прерывая фотосинтез на разных стадиях.
Цикл превращения солнечной энергии в углеводы — так называемый цикл Кальвина — сходен с циклом Кребса: он тоже состоит из серии химических реакций, которые начинаются с соединения входящей молекулы с молекулой-«помощником» с последующей инициацией других химических реакций. Эти реакции приводят к образованию конечного продукта и одновременно воспроизводят молекулу-«помощника», и цикл начинается вновь. В цикле Кальвина роль такой молекулы-«помощника» выполняет пятиуглеродный сахар рибулозодифосфат (РДФ).

Цикл Кальвина начинается с того, что молекулы углекислого газа соединяются с РДФ. За счет энергии солнечного света, запасенной в форме АТФ и НАДФ-H, сначала происходят химические реакции связывания углерода с образованием углеводов, а затем — реакции воссоздания рибулозодифосфата. На шести витках цикла шесть атомов углерода включаются в молекулы предшественников глюкозы и других углеводов. Этот цикл химических реакций будет продолжаться до тех пор, пока поступает энергия. Благодаря этому циклу энергия солнечного света становится доступной живым организмам. У некоторых растений (например, у кукурузы и сахарного тростника, а также у многих тропических трав, включая ползучий сорняк) цикл осуществляется по-другому.
Дело в том, что углекислый газ в норме проникает через отверстия в поверхности листа, называемые устьицами. При высоких температурах устьица закрываются, защищая растение от чрезмерной потери влаги. В C3-растения при закрытых устьицах прекращается и поступление углекислого газа, что приводит к замедлению фотосинтеза и изменению фотосинтетических реакций. В случае же кукурузы углекислый газ присоединяется к трехуглеродной молекуле на поверхности листа, затем переносится во внутренние участки листа, где углекислый газ высвобождается и начинается цикл Кальвина. Благодаря этому довольно сложному процессу фотосинтез у кукурузы осуществляется даже в очень жаркую, сухую погоду. Растения, в которых происходит такой процесс, мы называем C4-растениями, поскольку углекислый газ в начале цикла транспортируется в составе четырехуглеродной молекулы. C3-растения — это в основном растения умеренного климата, а C4-растения в основном произрастают в тропиках. САМ-метаболизм состоит в том, что в ночное время в цитоплазме происходит восстановительное карбоксилирование ФЕП с образованием малата, как у С4-растений. Малат накапливается и хранится в вакуоли. Для образования ФЕП расходуется крахмал. Днем малат из вакуоли переходит в цитоплазму, где декарбоксилируется одним из трех возможных механизмов: НАДФ-маликэнзима, НАД-маликэнзима и ФЕП-карбоксикиназы.
С экологической точки зрения, САМ-метаболизм дает преимущества в условиях водного дефицита. В настоящее время установлено, что к САМ- метаболизму могут переходить С3-растения при водном дефиците, засолении, т. е. это достаточно распространенное явление. На скорость фотосинтеза влияют многие факторы. Основными из них являются интенсивность света, концентрация кислорода и углекислого газа, температура окружающей среды.
Состояние, когда скорость выделения кислорода растением равна скорости его дыхания, называется точкой компенсации.
Кислород в процессе фотосинтеза может действовать как конкурентный ингибитор, взаимодействуя с рибулозодисфосфатом вместо углекислого газа. При этом образуется одна молекула ФГК и фосфогликолат, сразу расщепляющийся до гликолата. Чтобы вернуть хотя бы часть углерода, связанного в бесполезном гликолате, у растения имеется процесс, называемый фотодыханием. Это зависимое от света потребление кислорода с выделением углекислого газа, заметное лишь у C3-растений, не имеет ничего общего с обычным дыханием. Фотодыхание, в целом, идёт с поглощением энергии; в результате образуется фосфоглицерат, а 25 % углерода теряется в виде CO2. У C4-растений фотодыхания практически нет, что и является причиной их большей продуктивности.
В связи с энергетической проблемой учёные пытаются провести фотосинтетические процессы искусственно, особенно их первые этапы, когда вода под действием солнечной радиации расщепляется на кислород и водород. Сжигание водорода (с образованием воды) – экологически чистый процесс, который мог бы стать неплохой заменой современным источникам энергии. Фотосинтез – единственный процесс, на основе которого, используя солнечную энергию, можно получать органическое вещество для формирования урожаев и повышать плодородие почвы. В результате фотосинтеза создается 95% органического сухого вещества. Поэтому можно утверждать, что управление этим процессом один из наиболее эффективных путей воздействия на продуктивность растений.
vseobiology.ru
Фотосинтез. Освещенность и продолжительность
В прошлом статье мы рассмотрели элементарный состав растения. Оказалось что 95% всех строительных материалов растение получает из воды и воздуха. Поэтому, в этом видео мы поговорим о фотосинтезе, процессе в котором углекислый газ и вода, под действием солнечного света преобразуются в глюкозу, которая в организме растений является основным и универсальным источником энергии для обеспечения всех процессов.
Глюкоза — это углевод, есть и другие углеводы, такие как крахмал и целлюлоза. Вообще растение на 80% состоит из углеводов и поскольку углеводы образуются в процессе фотосинтеза, фотосинтез считается основным процессом протекающем в растении и влияющим на его рост.
Фотосинтез
Химическая формула фотосинтеза выглядит так:
Углекислый газ и вода, под действием солнечного света преобразуются в глюкозу, в качестве побочного эффекта растение выделяют кислород. В этой формуле, самая интересная его часть относится к солнечному свету. Вообще, что такое солнечный свет. В нашем обывательском представлении — солнце — это большая лампочка которая освещает эту землю. На самом деле, помимо световой энергии, в энергию солнца входит

Все эти виды солнечного излучения различаются между собой по длине волны. Волны в микроволновке имеют длину размером с муравья, а радиоволны будут размером с человека или даже здание.

Все виды волн несут в себе энергию в виде квантов. Запомните, квант — это частица, которая несет энергию. Энергия кванта зависит от частоты волны. Все мы знаем, что радиационное воздейтвие на растения и животных губительны, это связанно с высокой энергией кванта, которую несут такие волны, энергия настолько высокая, что она просто разрушает клетки! Ренгеновское излучение и низкочастотное ультрафиолетовое излучение также вредит живым организмам. На наше счастье, атмосфера земли задерживает большую часть радиационного, ренгеновского и ультрафиолетового излучения, защищая нас.

Подходящая энергия, которая не разрушает растение и достаточна для запуска фотосинтеза, находится практически в одном диапазоне с видимым светом. Для растений этому диапазону дали название — фотосинтетически активная радиацией, сокращенно ФАР, т.е. такое излучение, которое дает энергию подходящую для запуска фотосинтеза.
Перед тем, как мы пойдем дальше, давайте начнем представлять свет как поток падающих на растение шариков, которые несут энергию запускающую процесс фотосинтеза. Эти шарики мы будем называть кванты.
Характеристики света
Поговорим о свете подробнее. Наиболее важными характеристиками света являются:
Сейчас мы поговорим о первых двух характеристиках: освещенности и продолжительности.
Освещенность (облученность)
Логично предположить, что чем больше энергии получает растение, тем быстрее оно будет расти.
И действительно, скорость фотосинтеза, зависит от количества энергии полученной растением. Чем больше квантов падает на растение, тем выше фотосинтез,тем быстрее происходит рост.Количество энергии падающей на растение в области фотосинтетически активной радиации — называется освещенностью. При низкой освещенности, энергии квантов недостаточно, чтобы фотосинтез обеспечивал все потребности растения. В таких условиях энергия черпается из ранее запасенных питательных веществ. Этот процесс называется дыханием, представьте себе этот процесс со знаком минус, растение расходует ранее запасенную энергию. Для роста, необходимо, чтобы процессы фотосинтеза преобладали над процессами дыхания. А для этого нужно больше энергии!

Как видно из графика, есть участок, когда растение очень хорошо откликается на повышение освещенности. В какой-то момент эффективность падает, а после наступает плато насыщения. Если продолжать увеличивать освещенность, наступит световое ингибирование фотосинтеза. Получается, что как недостаток, так и избыточное количество энергии вредит растению. Как и во всем — все яд, все лекарство.
Таким образом, нам нужно знать меру, как много энергии давать растению. Поэтому, давайте определимся, в чем будем мерить освещенность.
Люмены и люксы
Очень часто в описании электрических лампочек, можно увидеть характеристику освещенности выраженную в люменах или люксах. Люмены и люксы — это величины связанные с восприятием света человеческим зрением, а глаз человека имеет максимум возприятия в зеленой области спектра. Таким образом люмены и люксы — субъективная величина, характеризующая освещенность с точки зрения человека.
Ваты на метр квадратный
Эта величина характеризует общую энергию упавших на поверхность квантов за одну секунду. Это объективная величина учитывающая энергию всех квантов. Как вы помните именно энергия квантов запускает процесс фотосинтеза. То есть ваты на м2, как раз то, что нам нужно для измерения освещенности!
Оптимальной освещенностью для выращивания рассады будет 40 Вт/м2, а для выращивание на продукцию, например огурцов и помидор, освещенность должна быть 100 Вт/м2 .
ПродолжительностьПомимо освещенности, важным условием для роста является продолжительность. Поэтому, когда обсуждают необходимые условия для роста, говорят, что для выращивания рассады нужна освещенность 40 Вт/м2, а продолжительность должна составлять 14 часов, а для выращивания на продукцию, освещенность должна быть 100 Вт/м2 с продолжительностью 16 часов.
Таким образом, говорят о общем количество энергии, которое должно получить растение за один день. Для рассады это 40Вт*м2 * 14 Часов = 560 Вт*Ч/м2 в области ФАР, для продукции 100 Вт*м2 * 16 Часов = 1600 Вт*Ч/м2 в области ФАР.Итак, если говорить о домашнем выращивании растений или о выращивании растений в теплице, где тепература позволяет выращивать круглогодично, то ограничением для роста будет именно общее количество солнечного света. Как известно, зимой солнечный день может длится всего 9 часов, что недостаточно даже для выращивания рассады.
Есть таблица, в которой показано с какого месяца можно выращивать рассаду и продукцию, с точки зрения достаточной для этого энергии.

Таким образом, если вы хотите выращивать растения зимой, то существенную часть года солнечный свет не может служить единственным источником освещения. И тут нам на помощь приходят лампы! Первые опыты по досвечиванию растений искусственными источниками начались еще в 1865 году, тогда растения пытались подсветить керосиновыми лампами. Уже тогда было установлено, что искусственный свет может положительно влиять на растение. В наше время есть современные источники света, например светодиоды или газоразрядные лампы, которые обладают высокой эффективностью. Такие лампы могут быть использованы как дополнительный источник света и даже полностью замещать солнечный свет.
Искусственное освещение активно изучается с целью выращивания растений в космосе. Но обычному садоводу тоже полезно знать о досветке растений, ведь многие из вас сталкивались с вытягиванием рассады. А как по мне, это само по себе очень интересно. Поэтому в следующей статье мы поговорим о искусственном освещении.
Выводы
Пожалуй самый важный момент из этого видео, в том, что скорость протекания фотосинтеза зависит от уровня освещенности. Для разных растений своя потребность в освещенности. Как избыток, так и недостаток света плохо сказывается на растении. Теперь вы знаете, почему так важно обращать внимание на рекомендации в освещенности.
botanik.me
Фотосинтез растений
Распускаются весной на деревьях почки, появляются зеленые листочки, пробивается первая зеленая травка. И у красных тюльпанов, и у желтых нарциссов, и у белой сирени — у всех листья и стебли зеленые. А кувшинки на воде покачиваются среди толстых круглых зеленых листьев, и зеленый камыш на ветру шумит, и цапля в зеленой тине себе на обед лягушек ловит. Даже пушистые голубые ели и те зеленые!
Почему и зачем?!
Все зеленые части растений окрашены одинаково потому, что в клетках, их образующих, содержится много хлорофилла. Он не только придает веселый цвет листьям и стеблям, но и участвует в важнейшем процессе — фотосинтезе.
Растение для развития и роста нуждается в питании. Его пищей являются органические вещества. Органические соединения — глюкоза и крахмал — образуются в процессе фотосинтеза. То есть фотосинтез для растения, по сути, способ приготовления пищи из воды и углекислого газа при использовании энергии солнца.
Химическое уравнение фотосинтеза выглядит так: 6CO2 + 6h3O (солнечный свет) = C6h22O6 + 6O2. Разобраться в нем без подготовки очень непросто, хотя на самом деле это всего лишь означает, что под действием солнечного света из углекислого газа и воды при наличии хлорофилла образуются сахар и кислород.
Сложные преобразования происходят в растительных клетках, содержащих хлорофилл. Этот пигмент, окрашивающий листья и стебли в зеленый цвет, играет едва ли не главную роль в процессе фотосинтеза. Вместе с водой и углекислым газом он является необходимым участником реакции, протекающей только при наличие солнечной энергии.
Кислород — побочный продукт фотосинтеза, но совершенно необходимый всем живым организмам для дыхания: он является одним из важнейших условий существования нашего мира. Без кислорода погибнут и животные, и растения, и высокоорганизованные, и простейшие. Так же как кислород и органические вещества, растению нужна вода. Любое растение почти наполовину состоит из нее. Она всасывается корнями из почвы и разносится по всему зеленому организму по особым каналам. Ежедневная потребность в воде у разных растений неодинаковая. Одним в сутки требуется всего пара литров, другим — несколько сотен. Обитатели засушливых районов, чтобы выжить, приспособились жидкость экономить и по возможности ею запасать.
Поделиться ссылкой
sitekid.ru
Факторы, лимитирующие фотосинтез растений — МегаЛекции
Из всех факторов одновременно влияющих на процесс фотосинтеза лимитирующимбудет тот, который ближе к минимальному уровню. Это установил Блэкман в 1905 году. Разные факторы могут быть лимитными, но один из них главный.
1. При низкой освещенности скорость фотосинтеза прямопропорциональна интенсивности света. Свет – лимитирующий фактор при низкой освещенности. При большой интенсивности света происходит обесцвечивание хлорофилла и фотосинтез замедляется. В таких условиях в природе растения обычно защищены (толстая кутикула, опушенные листья, чешуйки).
- Для темновых реакций фотосинтеза необходим углекислый газ, который включается в органические вещества, в полевых условиях является лимитирующим фактором. Концентрация СО2 варьирует в атмосфере в пределах от 0,03–0,04%, но если повысить ее, то можно увеличить скорость фотосинтеза. Некоторые тепличные культуры сейчас выращиваются при повышенном содержании СО2.
- Температурный фактор. Темновые и некоторые световые реакции фотосинтеза контролируются ферментами, а их действие зависит от температуры. Оптимальная температура для растений умеренного пояса составляет 25 °С. При каждом повышении температуры на 10 °С (вплоть до 35 °С) скорость реакций удваивается, но из-за влияния ряда иных факторов растения лучше растут при 25 °С.
- Вода – исходное вещество для фотосинтеза. Недостаток воды влияет на многие процессы в клетках. Но даже временное увядание приводит к серьезным потерям урожая. Причины: при увядании устьица растений закрываются, а это мешает свободному доступу СО2 для фотосинтеза; при нехватке воды в листьях некоторых растений накапливается абсцизовая кислота. Это гормон растений – ингибитор роста. В лабораторных условиях ее используют для изучения торможения ростового процесса.
- Концентрация хлорофилла. Количество хлорофилла может уменьшаться при заболеваниях мучнистой росой, ржавчиной, вирусными болезнями, недостатком минеральных веществ и возрастом (при нормальном старении). При пожелтении листьев наблюдаются хлоротичные явления или хлороз. Причиной может быть недостаток минеральных веществ. Для синтеза хлорофилла нужны Fe, Mg, N и К.
- Кислород. Высокая концентрация кислорода в атмосфере (21%) ингибирует фотосинтез. Кислород конкурирует с углекислым газом за активный центр фермента, участвующего в фиксации СО2, что снижает скорость фотосинтеза.
- Специфические ингибиторы. Лучший способ погубить растение – это подавить фотосинтез. Для этого ученые разработали ингибиторы – гербициды – диоксины. Например:ДХММ – дихлорфенилдиметилмочевина – подавляет световые реакции фотосинтеза. Успешно используют для изучения световых реакций фотосинтеза.
- Загрязнение окружающей среды. Газы промышленного происхождения, озон и сернистый газ, даже в малых концентрациях сильно повреждают листья у ряда растений. К сернистому газу очень чувствительны лишайники. Поэтому существует метод лихеноиндикации – определение загрязнения окружающей среды по лишайникам. Сажа забивает устьица и уменьшает прозрачность листовой эпидермы, что снижает скорость фотосинтеза.
6. Факторы жизни растений, тепло, свет, воздух, вода - Растения в течение всей своей жизни постоянно находятся во взаимодействии с внешней средой. Требования растений к факторам жизни определяются наследственностью растений, и они различны не только для каждого вида, но и для каждого сорта той или иной культуры. Вот почему глубокое знание этих требований дает возможность правильно устанавливать структуру посевных площадей, чередование культур, размещение севооборотов.Для нормальной жизнедеятельности растениям необходимы свет, тепло, вода, питательные вещества, включая углекислоту и воздух.Основным источником света для растений является солнечная радиация. Хотя этот источник находится вне влияния человека, степень использования световой энергии солнца для фотосинтеза зависит от уровня агротехники: способов посева (направление рядков с севера на юг или с востока на запад), дифференцированных норм высева, обработки почвы и др.Своевременное прореживание растений и уничтожение сорняков улучшают освещенность растений.Тепло в жизни растений, наряду со светом представляет основной фактор жизни растений и необходимое условие для биологических, химических и физических процессов в почве. Каждое растение на различных фазах и стадиях развития предъявляет определенные, но неодинаковые требования к теплу, изучение которых составляет одну из задач физиологии растений и научного земледелия. тепло в жизни растений влияет на скорость развития в каждой стадии роста. В задачу земледелия входит также изучение теплового режима почвы и способов его регулирования.Вода в жизни растений и питательные вещества, за исключением углекислоты, поступающей как из почвы, так и из атмосферы, представляют почвенные факторы жизни растений. Поэтому воду и питательные вещества называют элементами плодородия почвы.Воздух в жизни растений (атмосферный и почвенный) необходим как источник кислорода для дыхания растений и почвенных микроорганизмов, а также как источник углерода, который растение усваивает в процессе фотосинтеза. Кроме того, Воздух в жизни растений необходим для микробиологических процессов в почве, в результате которых органическое вещество почвы разлагается аэробными микроорганизмами с образованием растворимых минеральных соединений азота, фосфора, калия и других элементов питания растений.
7. Показатели фотосинтетической продуктивности посева
Урожай создается в процессе фотосинтеза, когда в зеленых рас тениях образуется органическое вещество из диоксида углерода, воды и минеральных веществ. Энергия солнечного луча переходит в энергию растительной биомассы. Эффективность этого процес са и в конечном счете урожай зависят от функционирования посе ва как фотосинтезирующей системы. В полевых условиях посев (ценоз) как совокупность растений на единице площади представляет собой сложную динамическую саморегулирующуюся фотосинтезирующую систему. Эта система включает в себя много компонентов, которые можно рассматри вать как подсистемы; она динамическая, так как постоянно меняет свои параметры во времени; саморегулирующаяся, так как, не смотря на разнообразные воздействия, посев изменяет свои пара метры определенным образом, поддерживая гомеостаз.
Показатели фотосинтетической деятельности посевов. Посев представляет собой оптическую систему, в которой листья погло щают ФАР. В начальный период развития растений ассимиляци онная поверхность невелика и значительная часть ФАР проходит мимо листьев, не улавливается ими. С повышением площади лис тьев увеличивается и поглощение ими энергии солнца. Когда ин декс листовой поверхности* составляет 4...5, т. е. площадь листьев в посеве 40...50 тыс. м 2 /га, поглощение ФАР листьями посева до стигает максимального значения — 75...80 % видимой, 40 % общей радиации. При дальнейшем увеличении площади листьев погло щение ФАР не повышается. В посевах, где ход формирования площади листьев оптималь ный, поглощение ФАР может составить в среднем за вегетацию 50...60 % падающей радиации. Поглощенная растительным по кровом ФАР — энергетическая основа для фотосинтеза. Однако в урожае аккумулируется только часть этой энергии. Коэффици ент использования ФАР обычно определяют по отношению к па дающей на растительный покров ФАР. Если в урожае биомассы в средней полосе России аккумулировано 2...3 % прихода на посев ФАР, то сухая масса всех органов растений составит 10... 15 т/га, а возможная урожайность — 4...6 т зерна с 1 га. В изреженных по севах коэффициент использования ФАР составляет всего 0,5...1,0%.
При рассмотрении посева как фотосинтезирующей системы урожай сухой биомассы, создаваемый за вегетационный период, или его прирост за определенный период зависит от величины средней площади листьев, продолжительности периода и чистой продуктивности фотосинтеза за этот период.
У = ФП • ЧПФ,
где У —урожайность сухой биомассы, т/га;
ФП— фотосинтетический потенциал, тыс. м 2 - дни/га;
ЧПФ —чистая продуктивность фотосинтеза, г/(м2 - дни).
Фотосинтетический потенциал рассчитывают по формуле
ФП = SCT,
где Sc — средняя за период площадь листьев, тыс. м 2 /га;
Т — продолжительность периода, дни.
Основные показатели для ценоза, как и урожайность, опреде ляют в расчете на единицу площади —1м 2 или 1 га. Так, площадь листьев измеряют в тыс. м 2 /га. Кроме того, пользуются таким по казателем, как индекс листовой поверхности. Основную часть ассимиляционной поверхности составляют листья, именно в них осуществляется фотосинтез. Фотосинтез мо жет происходить и в других зеленых частях растений — стеблях, остях, зеленых плодах и т. п., однако вклад этих органов в общий фотосинтез обычно небольшой. Принято сравнивать посевы меж ду собой, а также различные состояния одного посева в динамике по площади листьев, отождествляя ее с понятием «ассимиляцион ная поверхность». Динамика площади листьев в посеве подчиняется определен ной закономерности. После появления всходов площадь листьев медленно повышается, затем темпы нарастания увеличиваются. К моменту прекращения образования боковых побегов и роста ра стений в высоту площадь листьев достигает максимальной за веге тацию величины, затем начинает постепенно снижаться в связи с пожелтением и отмиранием нижних листьев. К концу вегетации в посевах многих культур (зерновые, зерновые бобовые) зеленые листья на растениях отсутствуют. Площадь листьев различных сельскохозяйственных растений может сильно варьировать в течение вегетации в зависимости от условий водоснабжения, питания, агротехнических приемов. Максимальная площадь листьев в засушливых условиях достигает всего 5... 10 тыс. м 2 /га, а при избыточных увлажнении и азотном питании она может превышать 70 тыс. м 2 /га. Считается, что при индексе листовой поверхности 4...5 посев как оптическая фото- синтезирующая система работает в оптимальном режиме, поглощая наибольшее количество ФАР. При меньшей площа ди листьев часть ФАР лис тья не улавливают. Если площадь листьев больше 50 тыс. м 2 /га, то верхние ли стья затеняют нижние, их доля в фотосинтезе резко снижается. Более того, вер хние листья «кормят» ниж ние, что невыгодно для формирования плодов, се мян, клубней и т. д. Динамика площади лис тьев показывает, что на разных этапах вегетации посев как фотосинтезиру- ющая система функциони рует неодинаково (рис. 3). Первые 20...30 дней вегетации, когда средняя площадь листьев составляет 3...7 тыс. м 2 /га, большая часть ФАР не улавливается листьями, и поэтому коэффициент использования ФАР не может быть высоким. Далее площадь ли стьев начинает быстро нарастать, достигая максимума. Как пра вило, это происходит у мятликовых в фазе молочного состояния зерна, у зерновых бобовых в фазе полного налива семян в сред нем ярусе, у многолетних трав в фазе цветения. Затем площадь листьев начинает быстро снижаться. В это время преобладают перераспределение и отток веществ из вегетативных органов в генеративные. На продолжительность этих периодов и их соотношение влияют различные факторы, в том числе агротехнические. С их помощью можно регулировать процесс нарастания площади листьев и продол жительность периодов. В засушливых условиях густоту растений, а следовательно, и площадь листьев намеренно снижают, так как при большой площади листьев усиливается транспирация, растения сильнее страдают от недостатка влаги, урожайность уменьшается.
Таким образом, высокие урожаи обеспечиваются определенным ходом фотосинтетической деятельности растений в посевах. Оптимальный ход нарастания площади листьев и биомассы может быть определен для каждой культуры и сорта в конкретных условиях выращивания.
megalektsii.ru
Фотосинтез | Акваловер
Фотосинтезом называется процесс образования из неорганических веществ (углекислого газа и воды) органических веществ, с использованием энергии световых лучей и с одновременным выделением кислорода. Это противоположный дыханию растений процесс.
По сути, фотосинтез — это уникальное явление в природе, благодаря которому, наша планета постоянно получает достаточное для жизнедеятельности животных организмов и человека количество кислорода.
Главным условием осуществления растениями фотосинтеза является световая энергия.
Фотосинтез растения происходит в листьях и зеленых стеблях, внутри клеточных структур, называемых хлоропластами. В каждой клетке хлоропласта содержатся сотни молекул хлорофилла, которые улавливают свет, а также каротиноиды, которые улавливают те участки спектра света, которые не улавливают молекулы хлорофилла. Хлорофилл поглощает красные и синие лучи, каротиноиды — зеленую часть спектра.
Молекулы хлорофилла активизируются световой волной. Фотосинтез достигает своей верхней точки при длине волны с 600 по 700 нанометров (оранжево-желтая часть спектра).
Красные и оранжевые лучи — основной вид энергии для фотосинтеза. Этот спектр очень полно поглощается хлорофиллом и увеличивает образование углеводов при фотосинтезе. Красно-оранжевый спектр имеет решающее значение для всех физиологических процессов в растениях.
Фиолетово-синий спектр тормозит рост стеблей, листовых черешков и пластинок, тo есть формируют более компактные растения с толстыми листьями. Эти лучи стимулируют образование белков и органический синтез растений. Сине-фиолетовые лучи почти полностью поглощается хлорофиллом, что максимально благоприятно для фотосинтеза.
Зелёные лучи не поглощаются листьями. Листья и стебли становятся очень тонкими. Для фотосинтеза эти лучи самые неблагоприятные. Хлорофилл отражает зеленый свет, что и придает растениям специфическую зеленую окраску.

Кроме видимой части света для растений имеет значение невидимый свет – ультрафиолет (UV), который в свою очередь, делится на короткие (UVC), средние (UVB) и длинные лучи (UVA).
Средние ультрафиолетовые лучи (длиной 280-315 нанометров) называют еще лучами пониженных температур, так как они «закаливают» растения и повышают их холодостойкость. На хлорофилл ультрафиолетовые лучи практически не действуют. Длинные ультрафиолетовые лучи (длиной 315-380 нанометров) необходимы для обмена веществ и роста растений. Они так же сдерживают вытягивание стеблей и повышают содержание витамина С.
В воздухе лучи рассеиваются незначительно. Однако, в толще воды они поглощаются, теряя энергию. Вода неодинаково поглощает световые лучи различных длин волн. Сильнее всего поглощаются красные лучи, потеря энергии на глубине 30 см составляет 50%! При этом потеря энергии синего луча на глубине в 1 метр составляет всего лишь 0,5%.
Но это не значит, что для аквариума нужно выбирать свет с преобладанием синего. Чем краснее цвет, тем ниже цветовая температура, чем синее цвет, тем цветовая температура выше. Цветовая температура измеряется в Кельвинах (К). При цветовой температуре ниже 5000К растения и вода приобретают нездоровый желтый оттенок, а при 10000К растения выглядят неестественно синими. Оптимальной цветовой температурой, при которой растения выглядят наиболее естественно, может считаться цветовая температура от 7000-8000 К.
Сам процесс фотосинтеза происходит, упрощенно, в два этапа. В первый этап для реакции важен свет. Хлоропласт улавливает световую энергию и трансформирует ее в химическую энергию. Второй этап не зависит от света — это реакция, когда полученная в ходе первого этапа молекула отдает атомы водорода для создания глюкозы. При разложении воды образуются кислород и водород. Кислород выделяется в воду, а водород связывается белком ферредоксином. На всех этапах фотосинтеза принимают участие фосфорилированные соединения.
Глюкоза — это топливо, образующееся в процессе фотосинтеза и использующееся растением для построения листьев, стеблей и т.п. Излишки глюкозы хранятся в корнях, стеблях и листьях растения. Глюкоза может быть трансформирована в целлюлозу, которая используется в качестве материала для построения клеточной структуры.
Наиболее оптимальным для фотосинтеза является промежуток pH от 6,6 до 7,3, с концентрацией CO2 от 25 до 30 ppm, а также наличие необходимых макро- и микроэлементов.
Активный фотосинтез отличает здоровое растение от растения, которое просто выживает в аквариуме. Для успешного фотосинтеза растений в аквариуме необходимы правильно подобранная световая гамма и достаточное количество СО2.

www.aqualover.ru
Фотосинтез | Наука | FANDOM powered by Wikia

лист растения
Фотоси́нтез — фотохимические реакции или Фотохимия — это процесс образования органического вещества из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактероихлорофилл и бактериородопсин у бактерий). В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества.
Различают оксигенный и аноксигенный типы фотосинтеза. Оксигенный гораздо более широко распространён, его осуществляют растения, цианобактерии и прохлорофиты. В данной статье описан только он, аноксигенному фотосинтезу пурпурных и зелёных бактерий, а также геликобактерий посвящена отдельная статья.
Выделяют три этапа фотосинтеза: фотофизический, фотохимический и химический. На первом этапе происходит поглощение квантов света пигментами, их переход в возбуждённое состояние и передача энергии к другим молекулам фотосистемы. На втором этапе происходит разделение зарядов в реакционном центре, перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом АТФ и НАДФН. Первые два этапа вместе называют светозависимой стадией фотосинтеза. Третий этап происходит уже без обязательного участия света и включает в себя биохимические реакции синтеза органических веществ с использованием энергии, накопленной на светозависимой стадии. Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюконеогенез, образование сахаров и крахмала из углекислого газа воздуха.
Пространственная локализация  Править
Править

Хлоропласты в клетках листа
Фотосинтез растений осуществляется в хлоропластах, обособленных двухмембранных органеллах клетки. Хлоропласты могут быть в клетках плодов, стеблей, однако основным органом фотосинтеза, анатомически приспособленным к его ведению, является лист. В листе наиболее богата хлоропластами ткань палисадной паренхимы. У некоторых суккулентов с вырожденными листьями (например, кактусы) основная фотосинтетическая активность связана со стеблем.
Свет для фотосинтеза захватывается более полно благодаря плоской форме листа, обеспечивающей большое отношение поверхности к объёму. Вода доставляется из корня по развитой сети сосудов (жилок листа). Углекислый газ поступает отчасти посредством диффузии через кутикулу и эпидермис, однако большая его часть диффундирует в лист через устьица и по листу по межклеточному пространству. Растения, осуществляющие С4 и CAM фотосинтез сформировали особые механизмы для активной ассимиляции углекислого газа.
Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые соединяясь друг с другом образуют тилакоиды, которые в свою очередь группируются в стопки, называемые граны. Внутритилакоидное пространство отделено и не сообщается с остальной стромой, предполагается также что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (70s типа), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их предшественниками свободные цианобактерии, вошедшие в состав эукариотической клетки в процессе симбиогенеза.
Цианобактерии, таким образом, как бы сами являются хлоропластом и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды, однако, не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе.
Световая (светозависимая) стадия Править
В ходе световой стадии фотосинтеза образуется высокоэнергетические продукты: АТФ, служащий в клетке источником энергии, и НАДФН, использующийся как восстановитель. В качестве побочного продукта выделяется кислород. В общем роль световых реакций фотосинтеза заключается в том, что в световую фазу синтезируются молекула АТФ и молекулы-переносчики протонов, т.е. НАДФ Н2.
Фотохимическая суть процесса Править
Хлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на спектре его поглощения): первый связан с переходом на более высокий энергетический уровень электрона системы сопряжённых двойных связей, второй — с возбуждением неспаренных электронов азота и кислорода порфиринового ядра. При неизменном спине электрона формируются синглетные первое и второе возбуждённое состояние, при изменённом — триплетное первое и второе.
Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно и хлорофилл за 10-12 сек переходит с него на первое, с потерей 100 кДж/моль энергии только в виде теплоты. Из первого синглетного и триплетного состояний молекула может переходить в основное с выделением энергии в виде света (флуоресценция) или тепла, с переносом энергии на другую молекулу, либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносов электрона на другое соединение.
Вторая возможность реализуется в светособирающих комплексах, первая — в реакционных центрах, где переходящий в возбужденное состояние под воздействием кванта света хлорофилл становится донором электрона (восстановителем) и передаёт его на первичный акцептор. Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передаёт его вторичному. Кроме того, время жизни полученных соединений выше чем у возбуждённой молекулы хлорофилла. Происходит стабилизация энергии и разделения зарядов. Для дальнейшей стабилизации вторичный донор электронов окисляет положительно заряженный хлорофилл, первичным донором же является в случае оксигенного фотосинтеза вода.
Проблемой, с которой сталкиваются при этом проводящие оксигенный фотосинтез организмы, является различие окислительно-восстановительных потенциалов воды (для полуреакции h30 → O2 E0=+0,82 В) и НАДФ+ (E0=-0,32 В). Хлорофилл при этом должен иметь в основном состоянии потенциал больший +0,82 В чтобы окислять воду, но при этом иметь в возбуждённом состоянии потенциал меньший чем −0,32 В чтобы восстанавливать НАДФ+. Одна молекула хлорофилла не может отвечать обоим требованиям. Поэтому сформировалось две фотосистемы и для полного проведения процесса необходимо два кванта света и два хлорофилла разных типов.
Светособирающие комплексы Править
Хлорофилл выполняет две функции: поглощения и передачу энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.
Передача энергии идёт резонансным путём (механизм Фёрстера) и занимает для одной пары молекул 10-10-10-12 сек., расстояние на которое осуществляется перенос составляет около 1 нм. Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каратиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины (с максимумом поглощения при 495—565 нм), за ними фикоцианины (550—615 нм) и аллофикоцианины (610—670 нм), последовательно передающие энергию на хлорофилл a (680—700 нм) реакционного центра.
Основные компоненты электронтранспортной цепи Править
Фотосистема II Править
Фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. Светособирающий комплекс II содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр фотосистемы II представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофилла a с максимумом поглощения при 680 нм (П680). На него в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула П680 становится сильным восстановителем (E0=-0,7 В).
П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС II и далее на пластохиноны, транспортируемые в восстановленной форме к b6f комплексу. Одна молекула пластохинона переносит 2 электрона и 2 протона, которые берутся из стромы.
Заполнение электронной вакансии в молекуле П680 происходит за счёт воды. В состав ФС II входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутритилакоидного пространства и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС II — это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве и 2 восстановленных пластохинонов в мембране.
b6f или b/f-комплекс Править
b6f комплекс является насосом, перекачивающим протоны из стромы во внутритилакоидное пространство и создающий градиент их концентрации за счёт выделяющейся в окислительно-восстановительных реакциях электронтранспортной цепи энергии. 2 пластохинона дают перекачку 4 протонов. В дальнейшем трансмембранный протонный градиент (pH стромы около 8, внутритилакоидного пространства — 5) используется для синтеза АТФ трансмембранным ферментом АТФ-синтазой.
Фотосистема I Править
Светособирающий комплекс I содержит примерно 200 молекул хлорофилла.
В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор — хлорофилл a, тот — вторичный (витамин K1), после чего электрон передаётся на филлохинон, от него на ферридоксин, который и восстанавливает НАДФ с помощью фермента ферридоксин-НАДФ-редуктазы.
Белок пластоцианин, восстановленный в b6f комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700.
Циклический и псевдоциклический транспорт электрона Править
Помимо полного нециклического пути электрона, описанного выше, обнаружены циклический и псевдоциклический.
Суть циклического пути заключается в том, что ферридоксин вместо НАДФ восстанавливает пластохинон, который переносит его назад на b6f комплекс. В результате образуется больший протонный градиент и больше АТФ, но не возникает НАДФН.
При псевдоциклическом пути ферридоксин восстанавливает кислород, который в дальнейшем превращается в воду и может быть использован в фотосистеме II. При этом также не образуется НАДФН.
Темновая стадия Править
В темновой стадии с участием АТФ и НАДФН происходит восстановление CO2 до глюкозы. Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
С3 — фотосинтез, цикл Кальвина Править
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий:
- Карбоксилирования
- Восстановления
- Регенерация акцептора CO2
На первой стадии к рибулозо-1,5-бифосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа (Rubisco). Этот белок составляет основную фракцию белков хлоропласта и предположительно наиболее распространённый фермент в природе. В результате образуется промежуточное неустойчивое соединение, распадающееся на две молекулы 3-фосфоглицериновой кислоты (ФГК).
Во второй стадии ФГК в два этапа восстанавливается. Сначала она фосфорилируется АТФ под действием фосфороглицерокиназы, затем НАДФН при воздействии триозофосфатдегидрогеназы её карбоксильная группа окисляется до альдегидной и она становится углеводом (ФГА).
В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 5-углеродных рибулозо-1,5-бифосфата, для чего необходимы 3АТФ.
Наконец, две ФГА необходимы для синтеза глюкозы. Для образования одной её молекулы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН и 18 АТФ.
С4 — фотосинтез Править
При низкой концентрации растворённого в строме CO2 Rubisco катализирует реакцию окисления рибулёзо-1,5-бифосфата и его распад на 3-фосфоглицериновую кислоту и фосфогликолевую кислоту, которая вынужденно используется в процессе фотодыхания.
Для увеличения концентрации CO2 растения С4 типа изменили анатомию листа. Цикл Кальвина у них локализуется в клетках обкладки проводящего пучка, в клетках мезофилла же под действием ФЕП-карбоксилазы фосфоенолпируват карбоксилируется с образованием щавелеуксусной кислоты, которая превращается в малат или аспартат и транспортируется в клетки обкладки, где декарбоксилируется с образованием пирувата, возвращаемого в клетки мезофилла.
С4 фотосинтез практические не сопровождается потерями рибулёзо-1,5-бифосфата из цикла Кальвина, поэтому более эффективен. Однако он требует не 18, а 30 АТФ на синтез 1 молекулы глюкозы. Это оправдывает себя в тропиках, где жаркий климат требует держать устьица закрытыми, что препятствует поступлению CO2 в лист, а также при рудеральной жизненной стратегии.
САМ фотосинтез Править
При CAM (Crassulaceae acid metabolism) фотосинтезе происходит разделение ассимиляции CO2 и цикла Кальвина не в пространстве как у С4, а во времени. Ночью в вакуолях клеток по аналогичному вышеописанному механизму при открытых устьицах накапливается малат, днём при закрытых устьицах идёт цикл Кальвина. Этот механизм позволяет максимально экономить воду, однако уступает в эффективности и С4, и С3. Он оправдан при стресстолерантной жизненной стратегии.
Значение фотосинтеза Править
Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф) также является запасённой в процессе фотосинтеза.
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь кислород атмосферы биогенного происхождения и является его побочным продуктом. Формирование окислительной атмосферы полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу.
Изучение фотосинтеза Править
Первые опыты по фотосинтезу были проведены Джозефом Пристли в 1770-1780-х годах, когда он обратил внимание на «порчу» воздуха в герметичном сосуде горящей свечой (воздух переставал быть способен поддерживать горение, помещённые в него животные задыхались) и «исправление» его растениями. Пристли сделал вывод что растения выделяют кислород, который необходим для дыхания и горения, однако не заметил что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз.
Позже было установлено что помимо выделения кислорода растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 Роберт Майер на основании закона сохранения энергии постулировал что растения преобразуют энергию солнечного света в энергию химических связей. В 1877 В. Пфеффер назвал этот процесс фотосинтезом.
Хлорофиллы был впервые выделены в 1818 П. Ж. Пелетье и Ж. Кавенту. Разделить пигменты и изучить их по отдельности удалось М. С. Цвету с помощью созданного им метода хроматографии. Спектры поглощения хлорофилла были изучены К. А. Тимирязевым, он же, развивая положения Майера, показал что именно поглощенные позволяют повысить энергию системы, создав вместо слабых связей С-О и О-Н высокоэнергетические С-С (до этого считалось что в фотосинтезе используются жёлтые лучи, не поглощаемые пигментами листа). Сделано это было благодаря созданному им методу учёта фотосинтеза по поглощённому CO2, в ходе экспериментов по освещению растения светом разных длин волн (разного цвета) оказалось что интенсивность фотосинтеза совпадает со спектром поглощения хлорофилла.
Окислительно-восстановительную сущность фотосинтеза (как оксигенного, так и аноксигенного) постулировал Корнелис ван Ниль. Это означало что кислород в фотосинтезе образуется полностью из воды, что экспериментально подтвердил в 1941 А. П. Виноградов в опытах с изотопной меткой. В 1937 Роберт Хилл установил что процесс окисления воды (и выделения кислорода), а также ассимиляции CO2 можно разобщить. В 1954-1958 Д. Арнон установил механизм световых стадий фотосинтеза, а сущность процесса ассимиляции CO2 была раскрыта Мельвином Кальвином с использованием изотопов углерода в конце 1940-х, за эту работу в 1961 ему была присуждена Нобелевская премия.
В 1955 была выделена и очищена Rubisco. С4 фотосинтез был описан Ю. С. Карпиловым в 1960 и М. Д. Хэтчем и К. Р. Слэком в 1966.
- Холл Д., Рао К. Фотосинтез: Пер. с англ. — М.: Мир, 1983.
ru.science.wikia.com








