Экология СПРАВОЧНИК. Биомасса животных меньше чем биомасса растений
Биомасса потребителей водных и наземных биоценозов
Почему в наземных биоценозах биомасса потребителей обычно меньше биомассы фотосинтезирующих организмов, а в некоторых водных биоценозах наоборот?
Общая закономерность, состоящая в том, что биомасса продуцентов (фотосинтезирующих организмов) превышает биомассу потребителей (растительноядных организмов), которая, в свою очередь, должна быть больше биомассы хищных животных, известна под названием «правило пирамиды биомасс». На первый взгляд кажется, что это правило не терпит никаких исключений. На самом деле к приведенной выше формулировке требуется сделать очень существенную поправку. Для того чтобы «правило пирамиды биомасс» было справедливо, следует говорить не о биомассах, а о продукциях. Продукция есть биомасса, произведенная за некоторую единицу времени. Тогда все становится на свои места. Действительно, растительноядные организмы никак не могут произвести больше биомассы, чем они потребили растительной пищи, и потребить больше, чем было за некоторый промежуток времени произведено растительной продукции. На самом деле продукция растительноядных животных обычно бывает раз в десять меньше, чем продукция растений, ими потребленная, поскольку не вся пища усваивается, а из усвоенной пищи некоторая часть обязательно расходуется в качестве источ-ника энергии для движения, продукции тепла, процессов биосинтеза, и поэтому не может быть использована на построение тела животного, то есть на продукцию. В том случае, если продукция фотосинтезирующих организмов высока, но вся она постоянно потребляется растительноядными животными, мы, измеряя в каждый момент времени биомассу, обнаружим, что она невелика и на первый взгляд недостаточна для прокорма растительноядных организмов. Именно такая картина наблюдается во многих водных сообществах, где основными продуцентами являются микроскопические планктонные водоросли. Для них, как и для всех мелких организмов, характерна высокая скорость размножения, а следовательно, у них высока и продукция. В наземных сообществах ситуация несколько иная. Здесь основную массу продуцентов составляют высшие растения. Обычно это крупные растения, а следовательно, для них характерна меньшая, чем для одноклеточных водорослей, относительная скорость размножения. Кроме того, лишь небольшая часть их биомассы потребляется животными, в основном семена, плоды, молодые побеги, стебли и листья травянистых растений. Большой запас накопленной биомассы (листва деревьев и кустарников, живые части стволов и корней и т. д.) обычно практически не потребляется растительноядными организмами. Естественно, что в наземных сообществах не только продукция, но и биомасса растений больше, чем продукция животных. Однако высокая продукция и быстрое ее потребление — далеко не единственная возможная причина относительно небольшой биомассы фотосинтезирующих организмов. Во многих морских сообществах это явление может объясняться тем, что значительную часть продукции органических веществ дают симбиотические водоросли, обитающие внутри различных беспозвоночных — коралловых полипов, двустворчатых моллюсков тридакн и др. Они, конечно, тоже относятся к фотосинтезирующим организмам, но обычными методами мы не сможем их обнаружить. Кроме того, органическое вещество может создаваться не только в результате фотосинтеза, но и в результате хемосинтеза, при котором источником энергии для синтеза органических веществ служит не свет, а окислительно-восстановительные реакции. Существуют целые сообщества, например сообщества так называемых «черных курильщиков», или гидротермов, — глубоководных выходов горячих вод, богатых сероводородом, в которых вообще все органическое вещество продуцируется только в процессе хемосинтеза. Продуцентами являются серобактерии, окисляющие сероводород и служащие пищей разнообразным червям, моллюскам и ракообразным. Не так давно обнаружено подобное сообщество и на небольшой глубине в районе сероводородных источников на одном из Курильских островов. Наконец, многие водные биоценозы, прежде всего глубоководные, являются зависимыми и не могут существовать без притока органики извне. На больших океанических глубинах всегда темно и фотосинтез невозможен, и тем не менее там обитают многочисленные животные, потребляющие органические остатки, оседающие на дно из верхних слоев воды.
biootvet.ru
Биомасса мелких организмов
У мелких растений и животных — водорослей, бактерий, простейших— удельный метаболизм (на 1 г биомассы) значительно выше, чем у крупных организмов, например деревьев и позвоночных. Это относится и к фотосинтезу, и к дыханию. Во многих случаях самыми важными для метаболизма сообществами оказываются не малочисленные крупные, выделяющиеся своим размером организмы, а многочисленные мельчайшие организмы, часто неразличимые простым глазом. Так, микроскопические водоросли (фитопланктон), которых в озере в каждый данный момент наберется всего несколько килограммов на гектар, могут иметь такой же метаболизм, как намного большая биомасса деревьев в лесу или травы на лугу. Точно так же несколько килограммов мелких рачков (зоопланктон), «пасущихся» на водорослях, могут иметь общее дыхание, равное дыханию многих килограммов пасущейся на лугу коровы.[ ...]
Пирамиды биомассы представляют более фундаментальный интерес, так как в них устранен «физический» фактор и четко показаны количественные соотношения биомасс. Если организмы не слишком сильно различаются по размеру, то, обозначив на трофических уровнях общую массу особей, можно получить ступенчатую пирамиду. Но если организмы низших уровней в среднем мельче организмов высших уровней, то имеет место обращенная пирамида биомассы. Например, в экосистемах с очень мелкими продуцентами и крупными консументами общая масса последних может быть в любой данный момент выше общей массы продуцентов. Для пирамид биомассы можно сделать несколько обобщений.[ ...]
В общем пирамида биомасс лучше показывает соотношения урожаев на корню для экологических групп в целом. Обозначив на последовательных трофических уровнях общую массу особей, обычно можно получить ступенчатую пирамиду, если только организмы не слишком различаются по размерам. Но если организмы низших уровней в среднем значительно меньше организмов высших уровней, то может получиться обращенная пирамида биомассы. Например, в системах с очень мелкими продуцентами и крупными консументами общая масса последних может быть в любой данный момент выше. В таких случаях, хотя через трофический уровень продуцентов проходит больше энергии, чем через уровни консументов (это всегда остается законом), интенсивный обмен и быстрый оборот мелких организмов-продуцентов обусловливают в результате большую продукцию, но малый урожай на корню. Обращенные пирамиды биомассы чаще всего характерны для озер и морей. Растения (фитопланктон) в сумме весят больше своих потребителей (зоопланктона) в периоды высокой первичной продуктивности, например в период весеннего «цветения», но в другие периоды, зимой например, может наблюдаться обратная ситуация. В озерах и в море вторичные и третичные кон-сументы, например рыбы и моллюски, в большинстве случаев крупны и в сумме весят больше продуцентов — фитопланктона.[ ...]
Бентическими называются организмы, обитающие на дне водоема. В Ладожском озере фитобентос (донные водоросли, мхи, высшие водные растения) развит слабо, преимущественно в заливах и узкой полосе литорали открытого озера. Зообентос — обитающие на дне животные: простейшие (протозоа), черви, губки, ракообразные, моллюски, личинки насекомых. В последнее время зообентос принято делить на мейо- и макробентос. В состав озерного мейо-бентоса входят мелкие организмы с размером тела от 0.3 до 4 мм и индивидуальным весом до 1—2 мг. К Ладожском озере в составе мейобентоса наиболее важную роль играют нематоды и низшие ракообразные, а из факультативно мейобентических — личинки хирономид и олигохет. Размеры озерного макробентоса — от нескольких миллиметров до нескольких сантиметров. В Ладожском озере на глубинах более 100 м преобладают олигохеты. На меньших глубинах отмечена разнообразная фауна олигохет, гаммарид, хирономид и моллюсков с преобладанием по биомассе гаммарид.[ ...]
Крупные виды являются лучшими индикаторами, чем мелкие, потому что при том же энергетическом потоке может поддерживаться большая биомасса. Скорость оборота органического вещества у мелких организмов бывает так велика, что отдельный вид, зарегистрированный в момент исследования, может и не быть особенно полезным экологическим индикатором.[ ...]
Рассмотрим подробнее признаки 14 и 15. На начальной стадии сукцессии организмы обычно бывают мелкими, для них характерны простые жизненные циклы и высокие скорости размножения. Изменения размеров особей происходят вследствие перемещения биогенных веществ из неорганической в органическую фазу. Малые размеры дают преимущество, особенно автотрофам, в среде, богатой минеральными веществами, поскольку возрастает отношение площади поверхности тела к его объему. Однако по мере развития экосистемы неорганические биогенные вещества становятся все более связанными в биомассе, так что преимущество переходит к организмам с большим объемом, т. е. к более крупным - либо к более крупным особям данного вида, либо к видам с более крупными особями, либо к тем и другим, так или иначе обладающим большими возможностями для накопления веществ и более сложными жизненными циклами.[ ...]
При неизменном энергетическом потоке в пищевой сети, или цепи, более мелкие наземные организмы с высоким удельным метаболизмом создают относительно меньшую биомассу, чем крупные1. Значительная часть энергии уходит на поддержание обмена веществ. Это правило «метаболизм и размеры особей», или правило Ю. Одума, обычно не реализуется в водных биоценозах при учете реальных условий обитания в них (в идеальных условиях оно имеет всеобщее значение). Связано это с тем, что мелкие водные организмы в значительной мере поддерживают свой обмен веществ за счет внешней энергии непосредственно окружающей их среды.[ ...]
ПРАВИЛО ОДУМА — при неизменном потоке энергии через пищевую сеть более мелкие наземные организмы с более высоким удельным метаболизмом создают меньшую биомассу, чем более крупные.[ ...]
Как неоднократно подчеркивалось на протяжении всей книги, ни численность организмов, ни биомасса не могут служить показателем того, что делают мелкие организмы или как быстро они это делают. В продуктивных наземных экосистемах численность микроорганизмов-редуцентов достигает 1012—1015 на 1 м2, а биомасса—10—100 г на 1 м2 (табл. 2) (само собой разумеется, что в пустынях или других лимитированных средах эти величины много меньше). Дело не только в том, что измерение биомассы сопряжено с серьезными трудностями, но и в том, что такие измерения весьма мало информативны. Значительно целесообразнее производить количественную оценку функций— интенсивности дыхания, скорости разложения субстрата; некоторые аспекты этого будут рассмотрены в этой главе и в гл. 19.[ ...]
Энергетика природных биоценозов построена таким образом, что микроорганизмы (мелкие грибки и бактерии) потребляют примерно 90% энергии растительной биомассы, мелкие беспозвоночные животные — еще около 10%, а крупные животные (в том числе позвоночные) — всего 1 %. Эти организмы имеют малый коэффициент полезного действия, и их роль в биоте заключается в тонкой настройке функционирования сообществ. Следовательно, человек в естественных границах биосферы должен потреблять не более 1% добытой энергии, т. е. тратить только 1% на свои нужды, а 99% — на поддержание биоты.[ ...]
Метаболизм и размеры особей. При неизменном энергетическом потоке в пищевой цепи более мелкие организмы имеют более высокую интенсивность обмена, более высокий удельный метаболизм (метаболизм в пересчете на 1 кг массы), чем крупные организмы. При этом мелкие организмы создают относительно меньшую биомассу, чем крупные. Так, биомасса бактерий, имеющихся в данный момент в экосистеме, гораздо ниже биомассы млекопитающих. Эта закономерность получила название правила Одума. Это правило заслуживает особого внимания, поскольку из-за антропогенного нарушения природы происходит измельчание организмов, которое неминуемо должно привести к общему снижению продуктивности и к разладу в экосистемах.[ ...]
Данные, приведенные в табл. 15, — еще одна иллюстрация того факта, что общая численность или биомасса (в данный момент) может почти совсем не отражать активности редуцентов и других мелких организмов. Заметим, что благодаря внесению органики рассеиваемая энергия возросла в 15 раз, а численность бактерий и грибов — всего в 2 раза. Иными словами, увеличилась активность этих мелких организмов, они начали быстрее «проворачивать» энергию, но при этом не произошло пропорционального увеличения их биомассы на корню, как бывает у крупных организмов. Простейшие крупнее бактерий, и их численность возросла заметнее.[ ...]
Все живые компоненты экосистемы — продуценты, кон-сументы и редуценты — составляют общую биомассу («живой вес») сообщества в целом или тех или иных групп организмов. Биомассу обычно выражают в г/см3 в сыром или сухом виде, или в энергетических единицах — в калориях, джоулях и т.п. На образование биомассы расходуется не вся энергия, а только та, которая создает первичную продукцию. Если скорость изъятия биомассы консументами отстает от скорости прироста растений, то это ведет к постепенному приросту биомассы продуцентов и к избытку мертвого органического вещества. Последнее приводит к заторфовыванию болот и зарастанию мелких водоемов.[ ...]
Зоопланктон представлен 25 видами (Rotifera - 11, Cladocera - 5, Copepoda - 9), численность изменялась от 0.01 до 14170 экз./л, биомасса - от 0.002 до 551.1 мг/л. Численность макрозообентоса (отмечено 7 форм) изменялась от 450 до 55800 экз./кв.м, биомасса - от 4,2 до 110 г/кв.м. Для ванн с наибольшей соленостью (23%о) характерны практически полное отсутствие зоопланктона и высокое обилие зообентоса (биомасса до 109 г/кв.м), обусловленные доминированием Littorina obtusata. В водоеме с соленостью 13%о в планктоне доминируют каляниды Eurytemora hirundoides, основу зообентоса составляют личинки Chironomidae. В водоемах с соленостью 5-9 %о в планктоне обычно доминируют коловратки Нех-arthra (численность до 10000 экз./л) и Ascomorpha, что, возможно, связано с отсутствием ракообразных, механически повреждающих мелкие организмы при активном движении, в планктоне по биомассе доминируют хар-пактициды Idyaea furcata. В зообентосе преобладают амфиподы рода Gammarus. В пресноводных наскальных ваннах (менее 1%о) в планктоне преобладают ветвистоусые ракообразные Bosmina obtusirostris, Chydorus sphaericus и Daphnia magna. Здесь нередко отмечается резкое увеличение численности (до 14000 экз./л) - т.н. «эффект сгущения» при частичном пересыхании водоема. Большую часть биомассы и численности донных организмов составляют личинки комаров сем.[ ...]
При сильном стрессе Я возрастает. Величина Р значительна в активных популяциях мелких организмов, например бактерий и водорослей, а также в системах, получающих энергию извне.[ ...]
Важными характеристиками различных экосистем являются также продуктивные коэффициенты — отношения продукции к биомассе: Р%/В и Р„/В. Они всегда велики в сообществах мелких организмов с короткой продолжительностью жизни. Так, для планктона годовая продукция может во много раз превосходить среднегодовую биомассу (Р„/В>> 1). Коэффициент близок к единице в фитоценозах однолетних трав, но всегда мал в лесных сообществах, в биоценозах крупных организмов с относительно большой продолжительностью жизни.[ ...]
Избыточный активный ил отбирают из вторичных отстойников при влажности примерно 99% с содержанием в 1 м3 жидкости около 160 г биомассы. В илоуплотнителях влажность снижается примерно до 98%. Активный ил состоит из живых организмов и твердого субстрата (до 40%). Живые организмы — это одиночные бактериальные клетки, скопления бактерий, образующих зооглеи, простейшие, черви и грибы. Встречаются также личинки насекомых, рачки и другие мелкие животные. Твердый субстрат— это отмершая часть биомассы. Состав биомассы и количество различных микроорганизмов в ней зависят от состава примесей в очищаемых стоках. При соответствующей обработке и уплотнении избыточного активного ила из него можно получить концентрат для подкормки сельскохозяйственных животных. Создание технологии получения высококачественного концентрата также является решением проблемы утилизации избыточного активного ила, количество которого составляет около 1 % объема очищаемой воды.[ ...]
К концу первой декады июня в фауне защищенных участков прибрежья наступили значительные качественные изменения: ко-пеподитно-коловратный комплекс организмов заменился рачко-вым. Снижение численности коловраток произошло за счет исчезновения мелких мирных форм, тогда как крупная коловратка Asplanchna, имеющая относительно большую собственную массу, увеличила численность и дала до 50 % общей биомассы коловраток, которая в целом увеличилась до 0,2 г/м3. Число веслоногих рачков и их молоди в этот период и в дальнейшем продолжало оставаться низким, и в конце второй декады июня они исчезли.[ ...]
Приведенные рассуждения позволяют заметить, что на энергетические характеристики экосистемы определенное влияние оказывают размеры особей. Чем мельче организм, тем выше его удельный метаболизм (на единицу массы) и, следовательно, меньше биомасса, которая может сохраняться на данном трофическом уровне. И наоборот, чем крупнее организм, тем больше биомасса на корню. Так, «урожай» бактерий в данный момент будет гораздо ниже «урожая» рыбы или млекопитающих, хотя эти группы использовали одинаковое количество энергии. Иначе обстоит дело с продуктивностью. Поскольку продуктивность - это скорость прироста биомассы, то преимуществами здесь обладают мелкие организмы, которые благодаря более высокому уровню метаболизма имеют более высокие темпы размножения и обновления биомассы, т. е. более высокую продуктивность.[ ...]
Сезонная динамика зоопланктона в заливе Липовая курья (рис. 45, 46) имеет совершенно иной характер. Два небольших пика численности сообщества приходятся на июнь и август, тогда как биомасса резко возрастает в августе, а затем стремительно снижается. Таким образом, в начале лета в планктоне преобладают мелкие организмы, в августе наблюдается перестройка сообщества и появляются крупные формы.[ ...]
Концепция потока энергии не только позволяет сравнивать экосистемы между собой, но и дает средство для оценки относительной роли популяций в них. В табл. 14 приведены оценки плотности, биомассы и скорости потока энергии для 6 популяций, различающихся по размеру особей и по местообитанию. Численности в этом ряду варьируют на 17 порядков величины (в 1017 раз), биомасса — примерно на 5 порядков (в 10° раз), а поток энергии — лишь примерно в 5 раз Это сравнительное единообразие потоков энергии свидетельствует о том, что все 6 популяций относятся в своих сообществах к одному трофическому уровню (первичные консументы), хотя ни по численности, ни по биомассе этого предположить нельзя. Можно сформулировать некое «экологическое правило»: данные по численности приводят к преувеличению значения мелких организмов, а данные по биомассе — к преувеличению роли крупных организмов; следовательно, эти критерии непригодны для сравнения функциональной роли популяций, сильно различающихся по отношению интенсивности метаболизма к размеру особей, хотя, как правило, биомасса все же более надежный критерий, нежели численность. В то же время поток энергии (т. е. Р- -Й) служит более подходящим показателем для сравнения любого компонента с другим и всех компонентов экосистемы между собой.[ ...]
Отношения P/R и B/R широко варьируют. В целом часть энергии, идущая на дыхание, т. е. на поддержание структуры, велика в популяциях крупных организмов (люди, деревья и др.) и в сообществах с большой биомассой на корню. При стрессовых воздействиях на систему R возрастает. Величина Р, напротив, сравнительно велика в активных популяциях мелких организмов, например бактерий или водорослей, в молодых, быстро растущих сообществах и в системах, получающих энергетические дотации.[ ...]
Это утверждение, по-видимому, особенно справедливо по отношению к океанической эвфотической зоне, однако наннопланктон может играть решающую роль и в метаболизме прибрежных вод (Иентш и Райтер, 1959). Мельчайшие жгутиковые (размером около 5 мкм) встречаются также в изобилии в афотической зоне на глубине 1000 м и более. Большинство из них бесцветны, но некоторые содержат хлорофилл. Это заставляет предполагать, что они питаются гетеротрофно по крайней мере в течение большей части времени, используя растворенное органическое вещество, которое было синтезировано в фотической зоне. Эти жгутиковые, возможно, образуют одно из ключевых звеньев пищевой цепи от первичной продукции в фотической зоне к зоопланктону и бентосу афотической зоны (другое звено может быть представлено опускающимися вниз агрегатами, сформированными из растворенных органических веществ; вопрос этот будет обсуждаться ниже). Работами Помроя и Иоханнеса (1966) было показано, что на долю наннопланкто-на может приходиться большая часть дыхания и фотосинтеза планктона. Они нашли, что в общем дыхании планктона из вод Гольфстрима и Саргассова моря доля жгутиковых, слишком мелких, чтобы быть пойманными планктонной сетью, составляет от 94 до 99%.[ ...]
Благодаря массивной структуре наземных растений они образуют большое количество стойкого волокнистого детрита (листовой .опад, древесные остатки и т. д.), скапливающегося в гетеротрофном ярусе. В фитопланктонной системе, наоборот, «дождь детрита» состоит из мелких частиц, которые легче разлагаются и потребляются мелкими животными. Поэтому следует ожидать, что население сапротрофных микроорганизмов в почве будет более обильным, чем в донных отложениях под открытой водой (табл. 2). Однако, как мы уже подчеркивали, численность и биомасса мелких организмов не обязательно соответствуют их активности; интенсивность метаболизма и оборот грамма бактерий могут в зависимости от условий изменяться во много раз. В противоположность тому, что отмечается для продуцентов и микроконсумен-тов, число и вес макроконсументов в водных и наземных экосистемах более сопоставимы, если системы получают одинаковое количество энергии. Если включить в расчеты крупных наземных пастбищных животных, то численность и биомасса крупных подвижных консументов, или «пермеантов» (номадов), получится в обеих системах почти одинаковой (табл. 2).[ ...]
В том случае, если по данным о численности и биомассе можно вычислить скорость изменения и поток энергии, удается получить более надежную оценку значения популяции в ее сообществе. Это относительно простой случай: Огске-Нтит — насекомые строго растительноядные, ведут оседлый образ жизни и имеют только одну генерацию в год; травостой состоит только из одного вида растения, служащего насекомым единственным источником пищи и убежищем. Численность и биомасса особей на 1 м2 определялись через интервалы времени в 3—4 дня. С помощью этих данных •определяли рост популяции, или продуктивность, для чего к приросту веса живых особей прибавляли вес тех особей, которые погибли за период учета численности. Продуктивность выражали в ккал/м2 в день. Затем в лаборатории определяли потребление кислорода (дыхание) взрослых особей и нимф разного размера в зависимости от температуры. На основе этих данных для каждого промежутка времени вычисляли дыхание популяции по отношению к средней сырой биомассе и приводили эти данные к фактической температуре среды. С помощью соответствующего коэффициента (Ивлев, 1934) потребление кислорода переводили в калории. Общую скорость ассимиляции в популяции получали, складывая величины продукции и дыхания.[ ...]
Опыт проведенной работы показывает, что для выполнения спектрального анализа одрой пробы какого-либо биосубстрата требуется от 30 до 60 мг золы. Исходя из этого необходимо первоначально иметь достаточное количество сырого вещества изучаемого органа или ткани. При проведении экспериментов с очень мелкими организмами (фитопланктон, кормовые беспозвоночные, личинки и молодь рыб и др.) нужную для получения требуемых порций золы биомассу можно получить за счет увеличения количества подопытных организмов. В зависимости от целей и задач исследования метод спектрального анализа может быть применен при проведении самых разнообразных био-лоТических экспериментов на любых этапах проведения опыта.[ ...]
Описанные отношения означают существование крутого ступенеобразного снижения продуктивности при переходе от низших к высшим трофическим уровням, последовательность которых формирует пирамиду продуктивности (рис. 5-7). Две другие пирамиды являются естественным следствием первой. Число индивидуумов в последовательности трофических уровней должно в целом убывать и формировать пирамиду чисел, на что обратил внимание первым Чарльз Элтон (рис. 5-7, С). Пирамида чисел, однако, может иметь противоположное значение в случаях, когда много мелких организмов питаются одним большим организмом более низкого трофического уровня: тысячи насекомых могут питаться одним деревом или сотни паразитических червей — одним хозяином. В ряду трофических уровней также убывает биомасса (рис. 5-7,5). Пирамида биомассы значительно реже изменяется в обратном направлении, однако в сообществах планктона животные иногда по массе превосходят растения. Пирамида продуктивности является наиболее важной, пирамиды чисел и биомассы имеют подчиненное значение и в меньшей степени отражают последовательность трофических уровней.[ ...]
Теперь планктонные сети делают обычно из прочного шелка или нейлона; их пряди прочно скрепляют, скручивая и связывая. В таких сетях бывает от 18 до 200 ячеек на дюйм. При количественных исследованиях применяются замыкающиеся сети, позволяющие вылавливать животных на определенных глубинах, при этом в улов не попадаются животные, встречающиеся в то время, когда сеть опускается или поднимается вверх. Замыкающаяся сеть, снабженная устройством, измеряющим объем профильтрованной воды, называется планктоночерпателем Кларка—Бампуса (названа по именам двух биоголов, занимавшихся изучением моря). Даже тончайшая шелковая сеть улавливает только часть биомассы планктона (сетной планктон). Сетной планктон и нанно-планктон называют еще (это, по-видимому, более подходящие названия) макро- и микропланктон соответственно.[ ...]
В качестве примера контроля первого типа можно указать на непрерывную регистрацию загрязнения воздуха над большими городами Калифорнии. Сейчас это осуществляется при помощи датчиков, смонтированных на самолете, которые ежедневно измеряют и регистрируют концентрации БОг, N02, СО и других загрязнителей над большой областью. Во многих городах показатели загрязнения воздуха входят в сводки погоды. Уже отмечалась необходимость контроля содержания в атмосфере двуокиси углерода в глобальном масштабе. Биологические показатели широко используются для контроля загрязнения воды. Очень часто сообщество может дать больше «информации» о тотальных эффектах загрязнения, чем измерение отдельных факторов. Важная экологическая задача — найти способы быстрого «прочтения» этой информации.[ ...]
ru-ecology.info
биомасса
биома́ссаобщая масса особей одного вида, группы видов или сообщества в целом, приходящаяся на единицу площади или объёма местообитания. Выражается в массе сырого или сухого вещества (г/м², кг/га, г/м³ и т. д.). Биомасса растений называется фитомассой, животных – зоомассой. Наибольшая биомасса наблюдается во влажных тропических лесах (в горных влажных лесах Бразилии 350–700 т/га), в широколиственных лесах умеренного пояса (400–500 т/га), большая часть её приходится на надземные части растений (древесину). Несмотря на обилие животных, зоомасса здесь менее 1 % общей биомассы. При этом более половины зоомассы (в широколиственных лесах до 90 %) составляют представители почвенной фауны, в осн. дождевые черви. Наименьшая биомасса суши – в пустынях. В водной среде наибольшая биомасса в сообществах бентоса на литорали и в сублиторали морей (прикреплённые водоросли и донные животные), она достигает нескольких кг/м² и снижается с увеличением глубины, составляя на большей части дна океана в ср. десятые и сотые доли г/м². Биомасса фито– и зоопланктона в малопродуктивных морских водах не превышает в ср. нескольких десятков мг/м². В высокопродуктивных р-нах, напр. в Азовском море, макс. биомасса планктона в период цветения достигает 200 г/м², а ценозов бентоса 500 г/м² и более. Сильно различаются по биомассе планктона и бентоса озёра: в ср. биомасса планктона колеблется от 1 до нескольких десятков г/м², при этом у зообентоса она часто меньше, чем у зоопланктона. В более продуктивных озёрах она достигает 10–30 г/м² (100–300 кг/га). Биомасса рыб в озёрах ср. и высокой продуктивности – 75–150 кг/га. Биомасса всех форм жизни на Земле составляет, по разным подсчётам, 2,5–6,5 млрд. т; биомасса океана в 1000 раз меньше биомассы суши. На суше фитомасса в 1000 раз превышает зоомассу, в океане её в 30 раз меньше. Если всё живое вещество распределить по поверхности Земли, оно образует плёнку толщиной 5 мм. Однако его роль сопоставима с геологическими процессами. Суммарная биомасса, которая была произведена на Земле в течение 1 млрд. лет, превышает массу земной коры. Важной дополняющей характеристикой биоценоза является продукция – количество органического вещества, создаваемого в единицу времени на единицу площади, т. е. скорость его прироста. Вследствие интенсивного обмена веществ годовая продукция фитопланктона в наиболее продуктивных водах соизмерима с годовой продукцией лесов, биомасса которых в тысячи раз больше. Продукция луговых степей больше, чем продукция хвойных лесов, хотя их биомасса в 10 раз меньше.
Поделитесь на страничкеslovar.wikireading.ru
Биомасса поверхности суши и океана
На суше Земли, начиная от полюсов к экватору, биомасса постепенно увеличивается [44]. Вместе с тем возрастает и количество видов растений. Тундра с лишайниками и мхами (до 500 видов) сменяется хвойными и широколиственными лесами, затем степями (до 2000 видов) и субтропической растительностью (свыше 3000 видов). Наибольшее сгущение и многообразие растений имеет место во влажных тропических лесах (свыше 8000 видов). Высота деревьев достигает 110-120 м. Растения растут в несколько ярусов, эпифиты покрывают деревья. Количество и разнообразие видов животных зависят от растительной массы и тоже увеличиваются к экватору. В лесах животные расселены в различных ярусах. Наивысшей плотности жизнь достигает нри большом разнообразии строения организмов, т. е. при различной приспособленности видов к условиям совместного существования. Поэтому наибольшая плотность жизни наблюдается в биогеоценозах, где виды связаны цепями питания. Цепи питания, переплетаясь, образуют сложную сеть передачи химических элементов и энергии от одного звена к другому. Между организмами идет жесточайшее состязание за обладание пространством, пищей, светом, кислородом. Большое влияние на биомассу суши оказывает человек. Под его воздействием сокращаются площади, производящие биомассу. Это требует рационального использования земель и водоемов для промышленных и сельскохозяйственных нужд.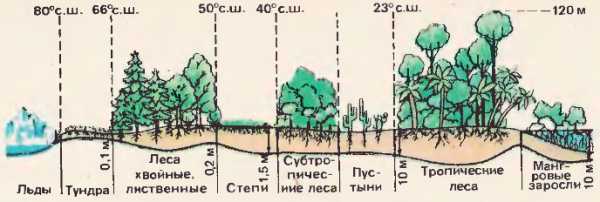 Рис. 44. Распространение биомассы по поверхности сушиБиомасса почвы. Своеобразные биогеоценозы почв покрывают почти всю поверхность суши. Почва не только среда, необходимая для жизни растений, но и биогеоценоз с разнообразными мельчайшими живыми организмами. Почва – рыхлый поверхностный слой земной коры, изменяемый атмосферой и организмами и постоянно пополняемый органическими остатками. Образование живого органического вещества происходит на земной поверхности; разложение органических веществ, их минерализация осуществляются главным образом в почве. Почва образовалась под воздействием организмов и физико-химических факторов. Мощность почвы наряду с поверхностной биомассой и под влиянием ее увеличивается от полюсов к экватору. В северных широтах особое значение имеет перегной, мощность которого в подзолистых почвах примерно 5-10 см, а в черноземных – 1-1,5 м В разных почвах существуют своеобразные биоценозы. Их составляют корни деревьев, кустарников, травянистых растений, расположенные в почве и в нижележащих слоях подпочвы ярусами. Скопления насекомых и их личинок, долбящих, роющих, сверлящих почву, производят огромную работу. По наблюдению Ч. Дарвина, дождевые черви, пропуская почву через кишечник, выносят ее на поверхность, ежегодно образуя слой толщиной 0,5 см, массой 25 т на 1 га.
Рис. 44. Распространение биомассы по поверхности сушиБиомасса почвы. Своеобразные биогеоценозы почв покрывают почти всю поверхность суши. Почва не только среда, необходимая для жизни растений, но и биогеоценоз с разнообразными мельчайшими живыми организмами. Почва – рыхлый поверхностный слой земной коры, изменяемый атмосферой и организмами и постоянно пополняемый органическими остатками. Образование живого органического вещества происходит на земной поверхности; разложение органических веществ, их минерализация осуществляются главным образом в почве. Почва образовалась под воздействием организмов и физико-химических факторов. Мощность почвы наряду с поверхностной биомассой и под влиянием ее увеличивается от полюсов к экватору. В северных широтах особое значение имеет перегной, мощность которого в подзолистых почвах примерно 5-10 см, а в черноземных – 1-1,5 м В разных почвах существуют своеобразные биоценозы. Их составляют корни деревьев, кустарников, травянистых растений, расположенные в почве и в нижележащих слоях подпочвы ярусами. Скопления насекомых и их личинок, долбящих, роющих, сверлящих почву, производят огромную работу. По наблюдению Ч. Дарвина, дождевые черви, пропуская почву через кишечник, выносят ее на поверхность, ежегодно образуя слой толщиной 0,5 см, массой 25 т на 1 га.
Почва плотно заселена живыми организмами. Биомасса одних дождевых червей в суглинистых почвах достигает 1,2 т на 1 га, или 2,5 млн. особей. Количество бактерий в 1 г почвы измеряется сотнями миллионов. Вода от дождей, тающих снегов обогащает ее кислородом и растворяет минеральные соли. Часть растворов удерживается в почве, часть выносится в реки и океан. Почва испаряет поднимающуюся по капиллярам грунтовую воду. Происходит движение растворов и выпадение солей в разных почвенных горизонтах.
В почве происходит и газообмен. Ночью при охлаждении и сжатии газов в нее проникает некоторое количество воздуха. Кислород воздуха поглощается животными и растениями и входит в состав химических соединений. Проникший в почву с воздухом азот улавливается некоторыми бактериями. Днем при нагревании почвы выделяются газы: углекислый, сероводород, аммиак. Все процессы, происходящие в почве, входят в круговорот веществ биосферы.
Некоторые виды хозяйственной деятельности человека (химизация сельскохозяйственного производства, переработка нефтепродуктов и др.) вызывают массовую гибель почвенных организмов, играющих важную роль в биосфере. Необходимо бережное отношение к почве, рациональное ее использование и защита от загрязнения.
Биомасса Мирового океана. Гидросфера Земли, или Мировой океан, занимает более 2/3 поверхности планеты. Объем воды в Мировом океане в 15 раз больше возвышающейся над уровнем моря суши.
Вода обладает особыми свойствами, важными для жизни организмов. Ее высокая теплоемкость делает более равномерной температуру океанов и морей, смягчая крайние изменения температуры зимой и летом. Теплопроводность воды больше теплопроводности воздуха в 20 раз. Океан замерзает только у полюсов, но и подо льдом существуют живые организмы
Вода – хороший растворитель. В состав воды океана входят минеральные соли, содержащие около 60 химических элементов. И, что особенно важно для жизни растений и животных, в ней растворяются поступающие из воздуха кислород и углекислый газ. Водные животные также выделяют при дыхании углекислый газ, а водоросли в процессе фотосинтеза обогащают воду кислородом.
Физические свойства и химический состав вод океана весьма постоянны и создают среду, благоприятную для жизни. Фотосинтез водорослей происходит главным образом в верхнем слое воды – до 100 м. Поверхность океана в этой толще заполнена микроскопическими одноклеточными водорослями, образующими микропланктон (греч. «планктос» -- блуждающий).
На океан приходится около 1/3 фотосинтеза, происходящего на всей планете. Водоросли поверхностного слоя океана – трансформаторы энергии солнечного излучения, превращающей ее в энергию химических реакций.
В питании животных океана преимущественное значение имеет планктон. Водорослями и простейшими питаются веслоногие рачки. Рачков поедают сельди и другие рыбы. Сельди идут в пищу хищным рыбам и чайкам. Исключительно планктоном питаются усатые киты.
В океане, кроме планктона и свободноплавающих животных, много организмов, прикрепленных ко дну и ползающих по нему. Население дна носит название бентоса (греч. «бентос» – глубинный).
В океане наблюдаются сгущения организмов: планктонное, прибрежное, донное. К живым сгущениям относятся и колонии кораллов, образующие рифы и острова. В основном в океане биомасса рассеяна. В громадной толще воды плавают рыбы, морские млекопитающие, кальмары.
В океане, особенно на дне его, распространены бактерии, превращающие органические остатки в неорганические вещества. Отмершие организмы медленно оседают на дно океана. Многие из них покрыты кремневыми или известковыми оболочками, а также известковыми раковинами. На дне океана они образуют осадочные породы. Так, на месте моря, покрывавшего 100 млн. лет назад Центральную Европу, находят в земле известняки, мел. В нем можно рассмотреть микроскопические раковины древнейших животных (корненожки и др.).
В Мировом океане живой биомассы в 1000 раз меньше, чем на суще. Использование энергии солнечного излучения на площади океана – 0,04%, на суше – 0,1%. Океан не так богат жизнью, как недавно еще предполагали.
В настоящее время в ряде стран решается проблема добычи из океана пресной воды, металлов и более полного использования его пищевых ресурсов с охраной наиболее ценных животных.
Гидросфера оказывает мощное влияние на всю биосферу. Суточные и сезонные колебания нагревания поверхности суши н океана вызывают циркуляцию тепла и влаги в атмосфере и влияют на климат и круговороты веществ во всей биосфере.
Добыча нефти в морях, перевозка ее в танкерах и другие виды деятельности человека приводят к загрязнению Мирового океана и сокращению его биомассы. Необходимо соблюдение мер охраны вод от загрязнений.
Воздействие человека на биомассу планеты. Человечество представляет собой небольшую биомассу в биосфере, но оно оказывает на нее грандиозное воздействие. Масштабы деятельности человечества расширяются благодаря постоянно растущей численности людей, стремительному ускорению научно-технического воздействия на природную среду.
В процессе природопользования человечество ежегодно перемещает на нашей планете более 4 трлн. т вещества, создает тысячи новых химических соединений, большинство из которых не включается в круговорот веществ и в конечном счете накапливается в биосфере, вызывая ее загрязнение. В результате промышленной деятельности происходит загрязнение природной среды, сокращение уровня солнечной радиации над крупными географическими регионами.
В развитии биосферы наступил такой период, когда человек должен планировать свою хозяйственную деятельность таким образом, чтобы она не нарушала сложившиеся в этой гигантской экосистеме закономерности, не способствовала сокращению биомассы.
1. Охарактеризуйте биомассу суши. 2. Что составляет биомассу почвы? 3. Как распределена биомасса в Мировом океане? 4. Какое влияние оказывает деятельность человечества на биомассу Земли?
blgy.ru