Виды естественного отбора. Среди каких растений и животных происходит естественный отбор
|
Естественный отбор – это дифференциальное выживание и размножение особей, которые отличаются друг от друга генетически детерминированными признаками. Более приспособленные к данным условиям среды особи оставляют больше потомков, чем менее приспособленные. Мы можем измерить относительную приспособленность особи долей её потомков среди особей следующего поколения и, следовательно, частотой ее аллелей, которые вошли в генофонд следующего поколения.
Для того, чтобы оценить приспособленность каждого конкретного зайца нам не нужно измерять его скорость бега, его хитрость в ускользании от волка, лисицы и прочих врагов, его способность находить и использовать пищу, его привлекательность для зайчих, нам нужно только знать, сколько он оставил потомков. Из двух зайцев более приспособлен тот, кто оставил больше потомков, кто внес больше своих генов в генофонд следующего поколения зайцев. Таким образом, частота аллелей, которые определяют более высокую приспособленность их носителей, увеличивается в генофонде популяции. Генетический состав популяции меняется. Поскольку мы определили эволюцию, как изменение генетического состава популяции, то мы можем рассматривать естественный отбор как фактор эволюции.
Естественный отбор принципиально отличается от других уже известных вам факторов эволюции – мутационного процесса и дрейфа генов. Мутации возникают случайно. Они не являются и не могут быть изначально адаптивными. При дрейфе генов частоты аллелей в популяциях меняются случайно, безотносительно к тому, насколько хорошо приспособлены носители того или иного аллеля. Естественный отбор определяет жесткую связь между приспособительной ценностью аллелей и их частотой в популяциях. Это единственный фактор, который обеспечивает приспособление всех живых организмов к постоянно меняющимся условиям внешней среды и регулирует гармоничные взаимодействия между генами внутри каждого организма.
Естественный отбор происходит автоматически. Все живые организмы из поколения в поколение проходят суровую проверку по всем мельчайшим деталям их строения, функционирования всех их систем в разнообразных условиях. Только те, кто выдержал эту проверку, оказываются отобранными и дают начало следующему поколению. Дарвин писал: «Естественный отбор ежедневно и ежечасно расследует по всему свету мельчайшие вариации, отбрасывая дурные, сохраняя и слагая хорошие, работая неслышно и незаметно, где бы и когда бы, ни представился к тому случай, над усовершенствованием каждого органического существа по отношению к условиям его жизни, органическим и неорганическим. Мы ничего не замечаем в этих медленных переменах в развитии, пока рука времени не отметит истекших веков».
Эффективность естественного отбора зависит от его интенсивности и запаса наследственной изменчивости, накопленного в популяции. Интенсивность отбора определяется тем, какая доля особей доживает до половой зрелости и участвует в размножении. Чем меньше эта доля, тем больше интенсивность отбора. Если, например, из 10 000 особей в каждом поколении выживают и размножаются только 100 самых крупных, то средний размер особей в этой популяции растет гораздо быстрее, чем в случае более мягкого отбора, когда, например, половина всех особей в популяции участвует в размножении.
Интенсивность отбора варьирует в широких пределах в зависимости от вида и экологической ситуации. Однако она никогда не бывает нулевой. Даже в человеческих популяциях, которые благодаря достижениям современной цивилизации в сильной степени защищены от действия отбора, его интенсивность весьма значительна. Около 30% всех беременностей заканчиваются выкидышами на самых ранних стадиях; 5% новорожденных гибнет при рождении, еще около 3% не доживает до зрелости; 20% людей, доживших до зрелости, не вступают в брак, и 10% вступивших в брак не имеют детей. В природных популяциях микроорганизмов, растений и животных интенсивность естественного отбора гораздо выше.
Однако даже самый интенсивный отбор будет неэффективен, если изменчивость особей в популяции незначительна или она носит ненаследственный характер. Если все особи в популяции совершенно одинаковы, то даже в условиях жесткой борьбы за существование все они имеют равные шансы уцелеть или погибнуть. Жизнь и размножение каждой такой особи зависит от случая. Из поколения в поколение генетический состав этой популяции будет оставаться неизменным.
Изменения не будут накапливаться и в том случае, если изменчивость особей в популяции родителей обусловлена не генетической изменчивостью, а разнообразием средовых условий. Тогда среднее значение признака и потомков особей с крайне высоким и крайне низким проявлением признака будет одинаковым. Такую картину мы всегда наблюдаем в так называемых чистых линиях (гомозиготных по всем генам) растений и животных и в клонах бесполых организмов. Поэтому отбор в чистых линиях и клонах оказывается неэффективным. Сколько бы поколений мы ни отбирали представителей чистых линий с наибольшим или наименьшим проявлением признака, популяционная средняя в последующих поколениях остается неизменной. В генетически гетерогенных популяциях отбор ведет к постепенному смещению средней из поколения в поколение.
Важнейшим свойством естественного отбора является его способность накапливать и совмещать полезные аллели. Отбор действует в каждом поколении, и в каждом поколении он начинается не с нуля, не с «чистого листа», а того состояния популяции, в которое она была приведена в результате предыдущего цикла отбора. Если в какой-то популяции из года в год высокие растения получают преимущество в выживании и размножении, то поколение за поколением в этой популяции растет частота высоких особей, и увеличивается средних размер особей. Результаты действия естественного отбора аккумулируются, накапливаются из поколения в поколение. Все новые и новые аллели возникают в популяции за счет мутационного процесса. Каждая мутация случайна, но ее носители регулярно отбираются, и размножаются в популяции, если она хоть чуть-чуть повышает приспособленность ее носителей. Если критерием приспособленности в данной популяции является высота растения, то отбор подхватывает все аллели, которые тем или иным способом работают на этом признак – увеличивают скорость роста, эффективность использования солнечного света, воды и других ресурсов.
Способностью отбора накапливать и совмещать полезные аллели и удалять вредные обусловлена и постепенность эволюции, и возникновение изумительно сложных органов, форм и функций. Самые сложные приспособления в строении, функциях и поведении живых организмов сформировались постепенно путем систематического отбора случайно возникающих мутаций.
Объектами отбора всегда являются отдельные особи, а не популяции и не виды. Ни одно приспособление, сколь угодно полезное для вида, не сможет быть подхвачено естественным отбором и распространиться в популяции, если оно вредно для конкретных особей.
Вернемся к проблеме взаимопомощи между животными, которую мы обсуждали выше. Долгое время оставалось загадкой, как это свойство могло возникнуть и закрепиться. Представим себе популяцию, состоящую из одних «эгоистов», которые сосредоточены только на своих собственных интересах и не склонны жертвовать собой ради других. Допустим, что в этой популяции появляются редкие мутанты-«альтруисты», которые всегда готова прийти на помощь другим. При этом они ставят под угрозу свою собственную жизнь. В такой ситуации победа в борьбе за жизнь достанется эгоистам, которые пользуются помощью альтруистов, но сами ничего не дают взамен. Гены «альтруистического поведения» будут довольно быстро утрачены. К такому же результату мы придем, если начнем наше рассмотрение с полностью альтруистической популяции. Допустим, что в такой популяции появляется эгоистичный мутант. Он охотно принимает помощь, но сам никому не помогает. Выгоды такого поведения для него и его эгоистичных потомков очевидны. Они получают незаслуженные преимущества в борьбе за жизнь, успешно размножаются и, в конечном счете, вытесняют гены альтруистов из генофонда популяции.
Ключ к разрешению этого парадокса был найден в расширении понятия приспособленности. В современной эволюционной биологии выделяется два компонента приспособленности: непосредственная приспособленность и расширенная. Под прямой приспособленностью особи понимается вклад ее собственных аллелей в генофонд популяции. Чем выше жизнеспособность и плодовитость особи, чем лучше она обеспечивает выживание своих потомков, тем выше ее непосредственная приспособленность. Расширенная приспособленность особи тем выше, чем больше аллелей, идентичных ее аллелям, передается следующему поколению. Особь может повысить свою совокупную приспособленность, повышая своими действиями непосредственную приспособленность своих родственников, которые содержат в своих генотипах те же аллели, что и она. Мать, защищая своих потомков, защищает по существу свои гены, переданные этим потомкам. Каждый из них имеет половину генов, полученных от матери, половину – от отца. Следовательно, коэффициент родства между родителем и потомком равен ?. Отсюда следует, что если ценой своей жизни, мать спасет жизнь двум своим детям, то ее совокупная приспособленность не снизится. Но ведь точно такая же степень родства связывает между собой братьев и сестер. Следовательно, по генетико-эволюционным соображениям братья и сестры должны проявлять друг к другу такую же степень заботы, как и мать к своим детям.
Такое понимание приспособленности объясняет возникновение и распространение альтруизма и взаимопомощи в популяциях животных. Помогая родственникам, животные способствуют распространению своих генов, в том числе и генов, ответственных за формирование альтруистического поведения. Не следует думать, однако, что животные действительно скрупулезно подсчитывают коэффициент родства, который связывает их с той или иной особью в популяции и на основании этих подсчетов принимают решение как им поступать – эгоистично или альтруистично – в каждом конкретном случае. Когда возникает угроза жизни ребенка, мать инстинктивно бросается на его защиту. Те матери, которые так не поступали, остались без потомков, аллели этих матерей были бесследно утеряны и не оставили следов в эволюции. Возникновение и распространение взаимопомощи между родственниками является, таким образом, автоматическим следствием естественного отбора.
|
www.evoluts.ru
Естественный отбор - «Энциклопедия»
ЕСТЕСТВЕННЫЙ ОТБОР, процесс избирательного выживания и дифференциального размножения организмов, основной движущий фактор их эволюции. Идеи о существовании естественного отбора высказывались с начала 19 века разными английскими натуралистами (в том числе А. Уоллесом). Но только Ч. Дарвин (1842, 1859) оценил его как главный фактор эволюции. По Дарвину, естественный отбор является результатом борьбы за существование; даже незначительные наследуемые различия между особями одного вида могут дать преимущества в этой борьбе, которая обусловлена тенденцией организмов к высокой интенсивности размножения (в геометрической прогрессии) и невозможности сохранения всего потомства вследствие ограниченности природных ресурсов. Гибель подавляющего числа особей в каждом поколении неизбежно ведёт к естественному отбору - «выживанию наиболее приспособленного» к данным условиям. В результате суммирования полезных изменений в течение многих поколений формируются новые адаптации и в конечном счёте возникают новые виды. Рассуждения о действии естественного отбора Дарвин строил преимущественно на обобщении опыта доместикации животных и растений по аналогии с искусственным отбором, подчёркивая, однако, что в отличие от проводимой человеком селекции естественный отбор определяется взаимодействием организмов с условиями окружающей среды и не имеет определённой цели.
Реклама
Систематическое исследование естественного отбора, расширение и совершенствование методик его изучения началось с конца 19 века. Использование методов биометрии позволило установить статистически значимые различия между выжившими и погибшими организмами при изменении условий среды. Благодаря разработкам Р. Фишера, Дж. Холдейна, С. Райта и С. С. Четверикова, осуществивших синтез классического дарвинизма и генетики, появилась возможность приступить к экспериментальному изучению генетических основ естественного отбора. Обследованные природные популяции оказались буквально насыщены мутациями, многие из которых становились полезными при изменениях условий существования или при комбинации с другими мутациями. Было установлено, что мутационный процесс и свободное скрещивание (панмиксия) обеспечивают генетическую разнородность популяций и уникальность особей, имеющих разные шансы на выживание; это обусловливает высокую интенсивность и эффективность естественного отбора. Кроме того, стало очевидным, что естественный отбор имеет дело не с единичными признаками, а с целыми организмами и что генетическая сущность естественного отбора заключается в неслучайном (дифференцированном) сохранении в популяции определённых генотипов, избирательно передающихся следующим поколениям. Естественный отбор носит вероятностный характер, действует на основе мутационного процесса и существующего генофонда, влияет на частоту распространения генов и их комбинаций, способствует уменьшению негативного действия мутаций и образованию механизмов защиты от их вредного действия, определяя тем самым темпы и направления эволюции. Под контролем естественного отбора находятся не только разнообразные признаки, но и сами факторы эволюции, например интенсивность и характер мутабельности, аппарат наследственности (отсюда понятие «эволюция эволюции»). При отсутствии же естественного отбора происходит снижение или утрата приспособленности организмов из-за накопления нежелательных мутаций, что проявляется в возрастании генетические груза, в том числе в популяциях современного человека.
Выделяют более 30 форм естественного отбора; ни одна из них не существует в чистом виде, а скорее характеризует тенденцию действия отбора в конкретной экологической ситуации. Так, движущий отбор способствует сохранению определённого отклонения от прежней нормы и приводит к выработке новых приспособлений через направленную перестройку всего генофонда популяций, а также генотипов и фенотипов особей. Он может вести к доминированию одной (или нескольких) ранее существовавшей формы над другими. Классическим примером его действия стало преобладание в промышленных районах темноокрашенных форм бабочки берёзовая пяденица, незаметных для птиц на загрязнённых копотью стволах деревьев (до середины 19 века встречалась только светлая форма, имитировавшая пятна лишайников на светлых стволах берёзы). Быстрое привыкание к ядам различных видов насекомых и грызунов, возникновение резистентности микроорганизмов к антибиотикам свидетельствуют о том, что давления движущего отбора в природных популяциях достаточно для того, чтобы обеспечить быстрый адаптивный ответ на резкие изменения среды. Как правило, селекция по одному признаку влечёт за собой целый ряд преобразований. Например, длительный отбор на содержание белка или масла в зёрнах кукурузы сопровождается изменениями форм зёрен, размеров початков, их расположения над уровнем почвы и др.
Результатом действия движущего отбора в филогенезе крупных таксонов является ортоселекция, примером которой служит установленная В. О. Ковалевским направленная эволюция конечности предков лошади (от пятипалости к однопалости), протекавшая миллионы лет и обеспечившая увеличение скорости и экономичности бега.
Дизруптивный, или разрывающий, отбор благоприятствует сохранению крайних отклонений и ведёт к увеличению полиморфизма. Он проявляется в тех случаях, когда ни одна из внутривидовых форм с разными генотипами не получает абсолютного преимущества в борьбе за существование из-за разнообразия условий, одновременно встречающихся на одной территории; при этом прежде всего элиминируются особи со средним или промежуточным характером признаков. Ещё в начале 20 века российский ботаник Н. В. Цингер показал, что большой погремок (Alectoroleophus major), цветущий и плодоносящий на некошеных лугах в течение всего лета, на скашиваемых лугах образует две расы: ранневесеннюю, успевающую принести семена до начала покоса, и позднеосеннюю - низкие растения, не повреждаемые при покосе, а затем быстро зацветающие и успевающие дать семена до начала морозов. Другим примером полиморфизма служит различие в окраске раковин у земляной улитки (Capacea nemoralis), являющейся кормом для птиц: в густых буковых лесах, где в течение всего года сохраняется подстилка из красно-бурого опада, обычны особи с коричневой и розовой окраской; на лугах с жёлтой подстилкой преобладают улитки с жёлтой окраской. В смешанных же лиственных лесах, где характер фона меняется с наступлением нового сезона, ранней весной доминируют улитки с бурой и розовой окраской, а летом - с жёлтой. Дарвиновы вьюрки (Geospizinae) на островах Галапагос (классический пример адаптивной радиации) - конечный результат длительного дизруптивного отбора, приведшего к образованию десятков близкородственных видов.
Если указанные формы естественного отбора приводят к изменению и фенотипической, и генетической структуры популяций, то впервые описанный И. И. Шмалъгаузеном (1938) стабилизирующий отбор сохраняет в популяции среднее значение признаков (норму) и не пропускает в следующее поколение геномы особей, наиболее отклоняющихся от этой нормы. Он направлен на поддержание и повышение устойчивости в популяции среднего, ранее сложившегося фенотипа. Известно, например, что во время снежных бурь выживают птицы, которые по многим признакам (длина крыла, клюва, масса тела и т.д.) приближаются к средней норме, а отклонившиеся от этой нормы особи погибают. Размеры и форма цветков у растений, опыляемых насекомыми, более устойчивы, чем у растений, опыляемых ветром, что обусловлено сопряжённой эволюцией растений и их опылителей, «выбраковкой» уклонившихся от нормы форм (например, шмель не может проникнуть в слишком узкий венчик цветка, а хоботок бабочки не касается слишком коротких тычинок у растений с длинным венчиком). Благодаря стабилизирующему отбору при внешнем неизменном фенотипе могут идти существенные генетические изменения, обеспечивающие независимость развития адаптаций от колеблющихся условий среды. Одним из результатов действия стабилизирующего отбора можно считать «биохимическую универсальность» жизни на Земле.
Дестабилизирующий отбор (название предложено Д. К. Беляевым, 1970) ведёт к резкому нарушению систем регуляции онтогенеза, вскрытию мобилизационного резерва и росту фенотипической изменчивости при интенсивном отборе в каком-либо определённом направлении. Например, отбор на снижение агрессивности хищных зверей в неволе через перестройку нейрогуморальной системы ведёт к дестабилизации цикла размножения, сдвигам в сроках линьки, изменениям в положении хвоста, ушей, в окраске и др.
Обнаружены гены, которые могут быть летальными или снижать жизнеспособность организмов в гомозиготном состоянии, а в гетерозиготном, напротив, повышать экологическую пластичность и другие показатели. В этом случае можно говорить о так называемом сбалансированном отборе, обеспечивающем поддержание генетического разнообразия с определённым соотношением частот аллелей. Примером его действия может служить повышение устойчивости у больных серповидно-клеточной анемией (гетерозиготных по гену гемоглобина S) к заражению различными штаммами малярийного плазмодия (смотри Гемоглобины).
Важным шагом в преодолении стремления все признаки организмов объяснять действием естественного отбора стала концепция нейтральной эволюции, согласно которой часть изменений на уровне белков и нуклеиновых кислот происходит путём фиксации адаптивно нейтральных или почти нейтральных мутаций. Возможен отбор видов, возникающих в периферийных популяциях «внезапно» с геохронологической точки зрения. Ещё раньше было доказано, что катастрофический отбор, при котором в период резких изменений среды выживает небольшое число особей и даже единственный организм, может стать основой формирования нового вида за счёт хромосомной перестройки и смены экологической ниши. Так, образование ксерофитного, эндемичного вида Clarkia lingulata в горах Сьерра-Невада в Калифорнии объясняют сильной засухой, вызвавшей массовую гибель растений, принявшую катастрофический характер в периферийных популяциях.
Естественный отбор, затрагивающий вторичные половые признаки особей, называют половым (например, яркая брачная окраска самцов у многих видов рыб и птиц, зазывающие крики, специфические запахи, сильно развитые орудия для турнирного боя у млекопитающих). Эти признаки полезны, так как повышают возможность участия их носителей в воспроизведении потомства. В половом отборе наибольшую активность проявляют самцы, что выгодно для вида в целом, т.к. самки остаются в большей безопасности в период размножения.
Выделяют также групповой отбор, способствующий сохранению признаков, полезных семье, стае, колонии. Его частным случаем у колониальных насекомых является отбор сородичей, при котором стерильные касты (рабочие, солдаты и др.) обеспечивают (нередко ценой собственной жизни) выживание плодовитых особей (маток) и личинок и тем самым сохранение всей колонии. Альтруистическое поведение родителей, притворяющихся ранеными, чтобы увести хищника от своих детей, грозит гибелью имитатору, но в целом повышает шансы на выживание его потомства.
Хотя представления о ведущей роли естественного отбора в эволюции получили подтверждение во множестве экспериментов, до сих пор они подвергаются критике, исходящей из представления о невозможности образования организмов в результате случайной комбинации мутаций. При этом игнорируется тот факт, что каждый акт естественного отбора совершается на базе предыдущих результатов его же действия, которые, в свою очередь, предопределяют формы, интенсивность и направления естественного отбора, а значит, пути и закономерности эволюции.
Лит.: Шмальгаузен И. И. Факторы эволюции. 2-е изд. М., 1968; Майр Э. Зоологический вид и эволюция. М., 1968; Шеппард Ф. М. Естественный отбор и наследственность. М., 1970; Левонтин Р. Генетические основы эволюции. М., 1978; Wilson D. S. The natural selection of populations and communities. Menlo Park, 1980; Галл Я. М. Исследования по естественному отбору // Развитие эволюционной теории в СССР. Л., 1983; Гаузе Г. Ф. Экология и некоторые проблемы происхождения видов // Экология и эволюционная теория. Л., 1984; Ратнер В. А. Краткий очерк теории молекулярной эволюции. Новосиб., 1992; Докинз Р. Эгоистичный генерал М., 1993; Sober Е. The nature of selection: evolutionary theory in philosophical focus. Chi., 1993; Дарвин Ч. Происхождение видов... 2-е изд. СПб., 2001; Coyne J., Orr Н. А. Speciation. Sunderland, 2004; Gavrilets S. Fitness landscapes and the origin of species. Princeton, 2004; Яблоков А. В., Юсуфов А. Г. Эволюционное учение. 5-е изд. М., 2004; Северцов А. С. Теория эволюции. М., 2005; Колчинский Э. И. Э. Майр и современный эволюционный синтез. М., 2006.
Э. И. Колчинский.
knowledge.su
Естественный отбор и его роль в эволюции
Естественный отбор — природный процесс, при котором из всех живых организмов сохраняются во времени только те, которые обладают качествами, способствующими успешному воспроизведению себе подобных. По представлениям синтетической теории эволюции, естественный отбор является одним из важнейших факторов эволюции.
Механизм естественного отбораИдею о том, что в живой природе действует механизм, подобный искусственному отбору, впервые высказали английские учёные Чарльз Дарвин и Альфред Уоллес. Суть их идеи состоит в том, что для появления удачных созданий, природе вовсе не обязательно понимать и анализировать ситуацию, а можно действовать наугад. Достаточно создавать широкий спектр разнообразных особей — и, в конечном счёте, выживут наиболее приспособленные.
1. Сначала появляется особь с новыми, совершенно случайными, свойствами
2. Потом она оказывается или не оказывается способной оставить потомство, в зависимости от этих свойств
3. Наконец, если исход предыдущего этапа оказывается положительным, то она оставляет потомство и её потомки наследуют новоприобретённые свойства
В настоящее время, отчасти наивные взгляды самого Дарвина оказались частично переработаны. Так, Дарвин представлял, что изменения должны происходить очень плавно, а спектр изменчивости является непрерывным. Сегодня, однако, механизмы естественного отбора объясняются при помощи генетики, которая вносит некоторое своеобразие в эту картину. Мутации в генах, которые работают на первом этапе описанного выше процесса, являются существенно дискретными. Ясно, однако, что основная суть идеи Дарвина осталась без изменений.
Формы естественного отбора
Движущий отбор — форма естественного отбора, когда условия среды способствуют определённому направлению изменения какого-либо признака или группы признаков. При этом иные возможности изменения признака подвергаются отрицательному отбору. В результате в популяции от поколения к поколению происходит сдвиг средней величины признака в определённом направлении. При этом давление движущего отбора должно отвечать приспособительным возможностям популяции и скорости мутационных изменений (в ином случае давление среды может привести к вымиранию).
Современным случаем движущего отбора является «индустриальный меланизм английских бабочек». «Индустриальный меланизм» представляет собой резкое повышение доли меланистических (имеющих тёмную окраску) особей в тех популяциях бабочек, которые обитают в промышленных районах. Из-за промышленного воздействия стволы деревьев значительно потемнели, а также погибли светлые лишайники, из-за чего светлые бабочки стали лучше видны для птиц, а тёмные — хуже. В XX веке в ряде районов доля тёмноокрашенных бабочек достигла 95 %, в то время как впервые тёмная бабочка (Morfa carbonaria) была отловлена в 1848 году.
Движущий отбор осуществляется при изменении окружающей среды или приспособлении к новым условиям при расширении ареала. Он сохраняет наследственные изменения в определенном направлении, перемещая соответственно и норму реакции. Например, при освоении почвы, как среды обитания у различных неродственных групп животных конечности превратились в роющие.
Стабилизирующий отбор — форма естественного отбора, при котором действие направлено против особей, имеющих крайние отклонения от средней нормы, в пользу особей со средней выраженностью признака.
Описано множество примеров действия стабилизующего отбора в природе. Например, на первый взгляд кажется, что наибольший вклад в генофонд следующего поколения должны вносить особи с максимальной плодовитостью. Однако наблюдения над природными популяциями птиц и млекопитающих показывают, что это не так. Чем больше птенцов или детенышей в гнезде, тем труднее их выкормить, тем каждый из них меньше и слабее. В результате наиболее приспособленными оказываются особи со средней плодовитостью.
Отбор в пользу средних значений был обнаружен по множеству признаков. У млекопитающих новорожденные с очень низким и очень высоким весом чаще погибают при рождении или в первые недели жизни, чем новорожденные со средним весом. Учет размера крыльев у птиц, погибших после бури, показал, что большинство из них имели слишком маленькие или слишком большие крылья. И в этом случае наиболее приспособленными оказались средние особи.
Дизруптивный (разрывающий) отбор — форма естественного отбора, при котором условия благоприятствуют двум или нескольким крайним вариантам (направлениям) изменчивости, но не благоприятствуют промежуточному, среднему состоянию признака. В результате может появиться несколько новых форм из одной исходной. Дизруптивный отбор способствует возникновению и поддержанию полиморфизма популяций, а в некоторых случаях может служить причиной видообразования.
Одна из возможных в природе ситуаций, в которой, вступает в действие дизруптивный отбор, — когда полиморфная популяция занимает неоднородное местообитание. При этом разные формы приспосабливаются к различным экологическим нишам или субнишам.
Примером дизруптивного отбора является образование двух рас у погремка лугового на сенокосных лугах. В нормальных условиях сроки цветения и созревания семян у этого растения покрывают всё лето. Но на сенокосных лугах семена дают преимущественно те растения, которые успевают отцвести и созреть либо до периода покоса, либо цветут в конце лета, после покоса. В результате образуются две расы погремка — ранне- и позднецветущая.
Дизруптивный отбор осуществлялся искусственно в экспериментах с дрозофилами. Отбор проводился по числу щетинок, оставлялись лишь особи с малым и большим количеством щетинок. В результате примерно с 30-го поколения две линии разошлись очень сильно, несмотря на то, что мухи продолжали скрещиваться между собой, осуществляя обмен генами. В ряде других экспериментов (с растениями) интенсивное скрещивание препятствовало эффективному действию дизруптивного отбора.
Отсекающий отбор — форма естественного отбора. Его действие противоположно положительному отбору. Отсекающий отбор выбраковывает из популяции подавляющее большинство особей, несущих признаки, резко снижающие жизнеспособность при данных условиях среды. С помощью отсекающего отбора из популяции удаляются сильно вредные аллели. Также отсекающему отбору могут подвергаться особи с хромосомными перестройками и набором хромосом, резко нарушающими нормальную работу генетического аппарата.
Положительный отбор — форма естественного отбора. Его действие противоположно отсекающему отбору. Положительный отбор увеличивает в популяции число особей, обладающих полезными признаками, повышающими жизнеспособность вида в целом. С помощью положительного отбора и отсекающего отбора совершается изменение видов (а не только посредством уничтожения ненужных особей, тогда любое развитие должно остановиться, но этого не происходит). Среди примеров положительного отбора: чучело археоптерикса можно использовать как планер, а чучело ласточки или чайки нельзя. Но первые птицы летали лучше археоптерикса.
Другой пример положительного отбора — появление хищников, превосходящих своими «умственными способностями» многих других теплокровных. Или появление таких рептилий, как крокодилы, обладающих четырехкамерным сердцем и способных жить как на земле, так и в воде.
Палеонтолог Иван Ефремов утверждал, что человек прошел не только отбор на лучшую приспособленность к условиям окружающей среды, но и «отбор на социальность» — выживали те сообщества, члены которых лучше поддерживали друг друга. Это еще один из примеров положительного отбора.
Частные направления естественного отбора· Выживание наиболее приспособленных к условиям обитания видов и популяций, например видов, обладающих жабрами в воде, поскольку приспособленность позволяет выигрывать борьбу за выживание.
· Выживание физически здоровых организмов.
· Выживание физически сильнейших организмов, поскольку физическая борьба за ресурсы является неотъемлемой частью жизни. Имеет значение во внутривидовой борьбе.
· Выживание наиболее сексуально успешных организмов, поскольку половое размножение является доминирующим способом размножения. В данном случае в дело вступает половой отбор.
Однако все эти случаи являются частными, а главным остаётся успешное сохранение во времени. Поэтому иногда эти направления нарушаются ради следования главной цели.
Роль естественного отбора в эволюцииЧ. Дарвин полагал естественный отбор основополагающим фактором эволюции живого (селекционизм в биологии). Накопление в конце XIX - начале XX века сведений по генетике, в частности обнаружение дискретного характера наследования фенотипических признаков, подтолкнуло многих исследователей к пересмотру указанного тезиса Дарвина: в качестве чрезвычайно важных факторов эволюции стали рассматриваться мутации генотипа (мутационизм Г. де Фриза, сальтационизм Р. Гольдшмитда и др.). С другой стороны, открытие известных корреляций среди признаков родственных видов (закон гомологических рядов) Н. И. Вавилова привело к формулировке гипотез об эволюции на основе закономерностей, а не случайной изменчивости (номогенез Л. С. Берга, батмогенез Э. Д. Копа и др.). В 1920-1940-е г. г. в эволюционной биологии интерес к селекционистским теориям возродился благодаря синтезу классической генетики и теории естественного отбора.
Разработанная в результате этого синтетическая теория эволюции (СТЭ), часто называемая неодарвинизмом, опирается на количественный анализ частоты аллелей в популяциях, изменяющейся под влиянием естественного отбора. Тем не менее, открытия последних десятилетий в различных областях научного знания — от молекулярной биологии с её теорией нейтральных мутаций М. Кимуры и палеонтологии с её теорией прерывистого равновесия С. Дж. Гоулда и Н. Элдриджа (в которой вид понимается как относительно статическая фаза эволюционного процесса) до математики с её теорией бифуркаций и фазовых переходов — свидетельствуют о недостаточности классической СТЭ для адекватного описания всех аспектов биологической эволюции. Дискуссия о роли различных факторов в эволюции продолжается и сегодня, и эволюционная биология подошла к необходимости своего очередного, третьего синтеза.
Возникновение адаптаций в результате естественного отбора
Адаптациями называются свойства и признаки организмов, которые обеспечивают приспособление к той среде, в которой эти организмы живут. Адаптацией также называют процесс возникновения приспособлений. Выше мы рассмотрели, как некоторые адаптации возникают в результате естественного отбора. Популяции березовой пяденицы приспособились к изменившимся внешним условиям благодаря накоплению мутаций темной окраски. В популяциях человека, населяющих малярийные районы, адаптация возникла благодаря распространению мутации серповидно-клеточной анемии. И в том, и в другом случае, адаптация достигается за счет действия естественного отбора.
При этом материалом для отбора служит наследственная изменчивость, накопленная в популяциях. Поскольку разные популяции отличаются друг от друга по набору накопленных мутаций, то к одним и тем же факторам внешней среды они приспосабливаются по-разному. Так, африканские популяции адаптировались к жизни в малярийных районах за счет накопления мутаций серповидно-клеточной анемии HbS, а в популяциях, населяющих юго-восточную Азию устойчивость к малярии сформировалась на основе накопления ряда других мутаций, которые в гомозиготном состоянии также вызывают болезни крови, а в гетерозиготном - обеспечивают защиту от малярии.
Эти примеры иллюстрируют роль естественного отбора в формировании адаптаций. Нужно, однако, ясно понимать, что это – частные случаи относительно простых адаптаций, возникающих за счет селективного размножения носителей единичных «полезных» мутаций. Маловероятно, что большинство адаптаций возникло таким путем.
Покровительственная, предостерегающая и подражательная окраска. Рассмотрим, например, такие широко распространенные адаптации, как покровительственная, предостерегающая и подражательная окраска (мимикрия). Покровительственная окраска позволяет животным становится незаметными, сливаясь с субстратом. Одни насекомые поразительно сходны с листьями деревьев, на которых они обитают, другие напоминают засохшие веточки или шипы на стволах деревьев. Эти морфологические адаптации дополняются поведенческими приспособлениями. Насекомые выбирают для укрытия именно те места, где они менее заметны.
Несъедобные насекомые и ядовитые животные - змеи и лягушки, имеют яркую, предостерегающую окраску. Хищник, раз столкнувшись с таким животным, надолго ассоциирует этот тип окраски с опасностью. Этим пользуются некоторые неядовитые животные. Они приобретают поразительное сходство с ядовитыми, и тем самым снижают опасность со стороны хищников. Уж имитирует окраску гадюки, муха подражает пчеле. Это явление называется мимикрией.
Как возникли все эти удивительные приспособления? Маловероятно, чтобы единичная мутация могла обеспечивать такое точное соответствие между крылом насекомого и живым листом, между мухой и пчелой. Невероятно, чтобы единственная мутация заставляла покровительственно окрашенное насекомое прятаться именно на тех листьях, на которые оно похоже. Очевидно, что такие приспособления как покровительственная и предостерегающая окраски и мимикрия возникали путем постепенного отбора всех тех мелких уклонений в форме тела, в распределении определенных пигментов, во врожденном поведении, которые существовали в популяциях предков этих животных. Одной из важнейших характеристик естественного отбора является его кумулятивность – его способность накапливать и усиливать эти уклонения в ряду поколений, слагая изменения отдельных генов и контролируемых ими систем организмов.
Самая интересная и трудная проблема – это начальные стадии возникновения адаптаций. Понятно, какие преимущества дает почти идеальное сходство богомола с сухим сучком. Но какие преимущества могли быть у его далекого предка, который лишь отдаленно напоминал сучок? Неужели хищники так глупы, что их можно так легко обмануть? Нет, хищники отнюдь не глупы, и естественные отбор из поколения в поколение «учит» их все лучше и лучше распознавать уловки их добычи. Даже идеальное сходство современного богомола с сучком не дает ему 100-процентной гарантии, что ни одна птица его никогда не заметит. Однако его шансы ускользнуть от хищника выше, чем у насекомого с менее совершенной покровительственной окраски. Точно также, у его далекого предка, лишь чуть-чуть похожего на сучок, шансы на жизнь были несколько выше, чем у его родственника вовсе на сучок не похожего. Конечно, птица, которая сидит рядом с ним, в ясный день легко его заметит. Но если день туманный, если птица не сидит рядом, а пролетает мимо и решает не тратить времени на то, что может быть богомолом, а может быть и сучком, тогда и минимальное сходство сохраняет жизнь носителю этого едва заметного сходства. Его потомки, которые унаследуют это минимальное сходство, будут более многочисленны. Их доля в популяции станет больше. Это осложнит жизнь птицам. Среди них станут более успешными те, кто точнее будет распознавать замаскированную добычу. Вступает в действие тот самый принцип Красной Королевы, который мы обсуждали в параграфе, посвященном борьбе за существование. Чтобы сохранить преимущество в борьбе за жизнь, достигнутое за счет минимального сходства, виду-жертве приходится меняться.
Естественный отбор подхватывает все те мельчайшие изменения, которые усиливают сходство в окраске и форме с субстратом, сходство между съедобным видом и тем несъедобным видом, которому он подражает. Следует учитывать, что разные виды хищников пользуются разными методами поиска добычи. Одни обращают внимание на форму, другие на окраску, одни обладают цветным зрением, другие нет. Поэтому естественный отбор автоматически усиливает, насколько это возможно, сходство между имитатором и моделью и приводит к тем изумительным адаптациям, которые мы наблюдаем в живой природе.
Возникновение сложных адаптаций
Многие адаптации производят впечатление тщательно продуманных и направленно спланированных устройств. Как такая сложнейшая структура как глаз человека могла возникнуть путем естественного отбора случайно возникавших мутаций?
Ученые предполагают, что эволюция глаза началась с небольших групп светочувствительных клеток на поверхности тела наших очень далеких предков, живших около 550 млн. лет назад. Способность различать свет и тьму была, безусловно, полезна для них, повышала их шансы на жизнь по сравнению с их абсолютно слепыми сородичами. Случайно возникшее искривление «зрительной» поверхности улучшило зрение, это позволяло определить направление на источник света. Возник глазной бокал. Вновь возникающие мутации могли вести к сужению и расширению отверстия глазного бокала. Сужение постепенно улучшало зрение – свет стал проходить через узкую диафрагму. Как видите, каждый шаг повышал приспособленность тех особей, которые менялись в «правильном» направлении. Светочувствительные клетки формировали сетчатку. Со временем в передней части глазного яблока сформировался хрусталик, выполняющий функцию линзы. Он возник, по-видимому, как прозрачная двухслойная структура, наполненная жидкостью.
Ученые попытались смоделировать этот процесс на компьютере. Они показали, что глаз, подобный сложному глазу моллюска, мог возникнуть из слоя фоточувствительных клеток при относительно мягком отборе всего за 364 000 поколений. Иными словами, животные, у которых смена поколений происходит каждый год, могли сформировать полностью развитый и оптически совершенный глаз в менее, чем за полмиллиона лет. Эта очень короткий срок для эволюции, если учесть что средний возраст вида у моллюсков равняется нескольким миллионам лет.
Все предполагаемые стадии эволюции глаза человека мы можем обнаружить среди ныне живущих животных. Эволюция глаза шла разными путями в разных типах животных. Благодаря естественному отбору независимо возникло множество разных форм глаза, и человеческий глаз – только один из них, причем не самый совершенный
Если внимательно рассмотреть конструкцию глаза человека и других позвоночных животных, можно обнаружить целый ряд странных несообразностей. Когда свет попадает в глаз человека он проходит через хрусталик и попадает на светочувствительные клетки сетчатки. Свет вынужден пробиваться через густую сеть капилляров и нейронов, чтобы попасть на фоторецепторный слой. Как это ни удивительно, но нервные окончания подходят к светочувствительным клеткам не сзади, а спереди! Более того, нервные окончания собираются в оптический нерв, который отходит от центра сетчатки, и создает тем самым слепое пятно. Чтобы компенсировать затенение фоторецепторов нейронами и капиллярами и избавится от слепого пятна, наш глаз постоянно движется, посылая в мозг серию разных проекций одного и того же изображения. Наш мозг производит сложнейшие операции, складывая эти изображения, вычитая тени, и вычисляя реальную картину. Всех этих сложностей можно было бы избежать, если бы нервные окончания подходили к нейронам не спереди, а сзади как, например, у осьминога.
Само несовершенство глаза позвоночных проливает свет на механизмы эволюции путем естественного отбора. Мы уже не раз говорили о том, что отбор всегда действует «здесь и сейчас». Он сортирует разные варианты уже существующих структур, выбирая и слагая вместе лучшие из них: лучшие «здесь и сейчас», безотносительно к тому, во что эти структуры могут превратиться в далеком будущем. Поэтому ключ к объяснению и совершенств и несовершенств современных структур следует искать в прошлом. Ученые считают, что все современные позвоночные произошли от животных подобных ланцетнику. У ланцетника светочувствительные нейроны располагаются на переднем конце нервной трубки. Перед ними расположены нервные и пигментные клетки, которые прикрывают фоторецепторы от света попадающего спереди. Ланцетник принимает световые сигналы, приходящие с боков его прозрачного тела. Можно думать, что у общего предка позвоночных глаз был устроен сходным образом. Затем эта плоская структура стала преобразовываться в глазной бокал. Передняя часть нервной трубки впячивалась внутрь, и нейроны, находившиеся впереди рецепторных клеток, оказались поверх них. Процесс развития глаза у эмбрионов современных позвоночных в известном смысле воспроизводит последовательность событий, происходивших в далеком прошлом.
Эволюция не создает новых конструкций «с чистого листа», она меняет (часто неузнаваемо меняет) старые конструкции, так чтобы каждый этап этих изменений был приспособительным. Любое изменение должно повышать приспособленность его носителей или, хотя бы, не снижать ее. Эта особенность эволюции ведет к неуклонному совершенствованию различных структур. Она же является причиной несовершенства многих адаптаций, странных несообразностей в строении живых организмов.
Следует помнить, однако, что все приспособления, сколь бы совершенны они ни были, носят относительный характер. Понятно, что развитие способности к полету не очень хорошо совмещается с возможностью быстро бегать. Поэтому птицы, обладающие наилучшими способностями к полету, — плохие бегуны. Напротив, страусы, которые не способны летать, прекрасно бегают. Приспособление к определенным условиям может быть бесполезно или даже вредно при появлении новых условий. Однако условия обитания меняются постоянно и иногда очень резко. В этих случаях накопленные ранее адаптации могут затруднить формирование новых, что может вести к вымиранию больших групп организмов, как это случилось более 60—70 млн лет назад с некогда очень многочисленными и разнообразными динозаврами.
biofile.ru
Естественный отбор
Формы естественного отбора Выживание наиболее приспособленных особей Дарвин называл естественным отбором. Его не следует понимать как какой-то выбор, так как здесь мы имеем лишь естественное следствие гибели менее приспособленных. Естественный отбор реализуется через действие естественных факторов среды (температура, влажность, свет, паразиты, конкуренты, враги, трудности добывания пищи и т. п.), действует через сохранение и накопление мелких наследственных изменений. Например, даже незначительное удлинение хоботка у пчелы делает ее более способной добывать нектар из цветков, недоступных для других пчел. Пчелы с удлиненным хоботком и будут иметь преимущество по сравнению с теми, у которых сохраняется.в поколении обычная длина этого органа. Отбор происходит непрерывно на протяжении бесконечного ряда следующих друг за другом поколений и сохраняет главным образом те формы, которые в большей мере соответствуют данным условиям. Естественный отбор и элиминация части особей вида неразрывно связаны между собой и являются необходимым условием эволюции видов в природе. Схема действия естественного отбора в системе вида по Дарвину сводится к следующему: 1) Изменчивость свойственна любой группе животных и растений, и организмы отличаются друг от друга во многих отношениях; 2) Число организмов каждого вида, рождающихся на свет, превышает число тех, которые могут найти пропитание и выжить. Тем не менее, поскольку численность каждого вида в естественных условиях постоянна, следует предполагать, что большая часть потомства гибнет. Если бы все потомки какого-либо вида выживали и размножались, то весьма скоро они вытеснили бы все другие виды на земном шаре; 3) Поскольку рождается больше особей, чем может выжить, происходит борьба за существование, конкуренция за пищу и место обитания. Это может быть активная борьба не на жизнь, а на смерть, или менее явная, но не менее действенная конкуренция, как, например, для растений в период засухи или холода; 4) Среди множества изменений, наблюдающихся у живых существ, одни облегчают выживание в борьбе за существование, другие же приводят к тому, что их обладатели гибнут. Концепция «выживания наиболее приспособленных» представляет собой ядро теории естественного отбора; 5) Выживающие особи дают начало следующему поколению, и таким образом «удачные» изменения передаются последующим поколениям. В результате каждое следующее поколение оказывается более приспособленным к среде обитания; по мере изменения среды возникают дальнейшие приспособления. Если естественный отбор действует на протяжении многих лет, то последние отпрыски могут оказаться настолько несхожими со своими предками, что их целесообразно будет выделить в самостоятельный вид. Может также случиться, что некоторые члены данной группы особей приобретут одни изменения и окажутся приспособленными к окружающей среде одним способом, тогда как другие ее члены, обладающие другим комплексом изменений, окажутся приспособленными иначе; таким путем от одного предкового вида при условии изоляции подобных групп может возникнуть два и более видов. Движущий отбор Естественный отбор всегда ведет к увеличению средней приспособленности популяций. Изменение внешних условий может приводить к изменению приспособленности отдельных генотипов. В ответ на эти изменения, естественный отбор, используя огромный запас генетического разнообразия по множеству разных признаков, ведет к значительным сдвигам в генетической структуре популяции. Если внешняя среда меняется постоянно в определенном направлении, то естественный отбор меняет генетическую структуру популяции таким образом, чтобы ее приспособленность в этих меняющихся условиях оставалась максимальной. При этом меняются частоты отдельных аллелей в популяции. Меняется и средние значения приспособительных признаков в популяциях. В ряду поколений прослеживается их постепенное смещение в определенном направлении. Такую форму отбора называют движущим отбором. Классическим примером движущего отбора является эволюция окраски у березовой пяденицы. Окраска крыльев этой бабочки имитирует окраску покрытой лишайниками коры деревьев, на которых она проводит светлое время суток. Очевидно, такая покровительственная окраска сформировалась за многие поколения предшествующей эволюции. Однако с началом индустриальной революции в Англии это приспособление стало терять свое значение. Загрязнение атмосферы привело к массовой гибели лишайников и потемнению стволов деревьев. Светлые бабочки на темном фоне стали легко заметны для птиц. Начиная с середины XIX века, в популяциях березовой пяденицы стали появляться мутантные темные (меланистические) формы бабочек. Частота их быстро возрастала. К концу XIX века некоторые городские популяции березовой пяденицы почти целиком состояли из темных форм, в то время как в сельских популяциях по-прежнему преобладали светлые формы. Это явление было названо индустриальным меланизмом. Ученые обнаружили, что в загрязненных районах птицы чаще поедают светлые формы, а в чистых – темные. Введение ограничений на загрязнение атмосферы в 1950-х годах привело к тому, что естественный отбор вновь изменил направление, и частота темных форм в городских популяциях начала снижаться. В наше время они почти так же редки, как и до начала индустриальной революции. Движущий отбор приводит генетический состав популяций в соответствие изменениям во внешней среде так, чтобы средняя приспособленность популяций была максимальной. На острове Тринидад рыбки гуппи обитают в разных водоемах. Множество тех, что живут в низовьях речек и в прудах гибнет в зубах хищных рыб. В верховьях жизнь для гуппи гораздо спокойней – там мало хищников. Эти различия во внешних условиях привели к тому, что «верховые» и «низовые» гуппи эволюционировали в разных направлениях. «Низовые», находящиеся под постоянной угрозой истребления, начинают размножаться в более раннем возрасте и производят множество очень мелких мальков. Шанс на выживание каждого из них очень невелик, но их очень много и некоторые из них успевают размножиться. «Верховые» достигают половой зрелости позднее, их плодовитость ниже, но потомки крупнее. Когда исследователи переносили «низовых» гуппи в незаселенные водоемы в верховьях речек, они наблюдали постепенное изменение типа развития рыбок. Через 11 лет после перемещения они стали значительно крупнее, вступали в размножение позже и производили меньшее количество, но более крупных потомков. Скорость изменения частот аллелей в популяции и средних значений признаков при действии отбора зависит не только от интенсивности отбора, но и от генетической структуры признаков, по которым идет обор. Отбор против рецессивных мутаций оказывается значительно менее эффективным, чем против доминантных. В гетерозиготе рецессивный аллель не проявляется в фенотипе и поэтому ускользает от отбора. Используя уравнение Харди-Вейнберга можно оценить скорость изменения частоты рецессивного аллеля в популяции в зависимости от интенсивности отбора и начального соотношения частот. Чем ниже частота аллеля, тем медленнее происходит его элиминация. Для того чтобы снизить частоту рецессивной летали от 0,1 до 0,05 нужно всего 10 поколений; 100 поколений - чтобы уменьшить ее от 0,01 до 0,005 и 1000 поколений - от 0,001 до 0,0005. Движущая форма естественного отбора играет решающую роль в приспособлении живых организмов к меняющимся во времени внешним условиям. Она же обеспечивает широкое распространение жизни, ее проникновение во все возможные экологические ниши. Ошибочно думать, однако, что в стабильных условиях существования естественный отбор прекращается. В таких условиях он продолжает действовать в форме стабилизирующего отбора. Стабилизирующий отбор Стабилизирующий отбор сохраняет то состояние популяции, которое обеспечивает ее максимальную приспособленность в постоянных условиях существования. В каждом поколении удаляются особи, отклоняющиеся от среднего оптимального значения по приспособительным признакам. Описано множество примеров действия стабилизующего отбора в природе. Например, на первый взгляд кажется, что наибольший вклад в генофонд следующего поколения должны вносить особи с максимальной плодовитостью. Однако наблюдения над природными популяциями птиц и млекопитающих показывают, что это не так. Чем больше птенцов или детенышей в гнезде, тем труднее их выкормить, тем каждый из них меньше и слабее. В результате наиболее приспособленными оказываются особи со средней плодовитостью. Отбор в пользу средних значений был обнаружен по множеству признаков. У млекопитающих новорожденные с очень низким и очень высоким весом чаше погибают при рождении или в первые недели жизни, чем новорожденные со средним весом. Учет размера крыльев у птиц, погибших после бури, показал, что большинство из них имели слишком маленькие или слишком большие крылья. И в этом случае наиболее приспособленными оказались средние особи. В чем причина постоянного появления малоприспособленных форм в постоянных условиях существования? Почему естественный отбор не способен раз и навсегда очистить популяцию от нежелательных уклоняющихся форм? Причина не только и не столько в постоянном возникновении все новых и новых мутаций. Причина в том, что часто наиболее приспособленными оказываются гетерозиготные генотипы. При скрещивании они постоянно дают расщепление и в их потомстве появляются гомозиготные потомки со сниженной приспособленностью. Это явление получило название сбалансированный полиморфизм. Половой отбор У самцов многих видов обнаруживаются явно выраженные вторичные половые признаки, которые на первый взгляд кажутся неадаптивными: хвост павлина, яркие перья райских птиц и попугаев, алые гребни петухов, феерические цвета тропических рыбок, песни птиц и лягушек, и т.п. Многие из этих особенностей осложняют жизнь их носителей, делают их легко заметными для хищников. Казалось бы, эти признаки не дают никаких преимуществ их носителям в борьбе за существование, и тем не менее они очень широко распространены в природе. Какую роль в их возникновении и распространении сыграл естественный отбор? Известно, что выживание организмов является важным, но не единственным компонентом естественного отбора. Другим важнейшим компонентом является привлекательность для особей противоположного пола. Ч. Дарвин назвал это явление половым отбором. Впервые он упомянул эту форму отбора в «Происхождении видов», а затем подробно проанализировал ее в книге «Происхождение человека и половой отбор». Он считал, что «эта форма отбора определяется не борьбой за существование в отношениях органических существ между собою или с внешними условиями, но соперничеством между особями одного пола, обычно самцами, за обладание особями другого пола». Половой отбор - это естественный отбор на успех в размножении. Признаки, которые снижают жизнеспособность их носителей, могут возникать и распространяться, если преимущества, которые они дают в успехе размножения значительно выше, чем их недостатки для выживания. Самец, который живет недолго, но нравится самкам и поэтому производит много потомков, имеет гораздо более высокую совокупную приспособленность, чем тот, что живет долго, но оставляет мало потомков. У многих видов животных подавляющее большинство самцов вовсе не участвует в размножении. В каждом поколении между самцами возникает жесточайшая конкуренция за самок. Эта конкуренция может быть прямой, и проявляться в виде борьбы за территории или турнирных боев. Она может происходить и в косвенной форме и быть обусловленной выбором самок. В тех случаях, когда самки выбирают самцов, конкуренция самцов проявляется в демонстрации их яркого внешнего вида или сложного поведения ухаживания. Самки выбирают тех самцов, которые им больше всего нравятся. Как правило, это наиболее яркие самцы. Но почему самкам нравятся яркие самцы? Приспособленность самки зависит о того, насколько объективно она способна оценить потенциальную приспособленность будущего отца своих детей. Она должна выбрать такого самца, сыновья которого будут обладать высокой приспособленностью и привлекательностью для самок. Было предложено две основные гипотезы о механизмах полового отбора. Согласно гипотезе «хороших генов» самка «рассуждает» следующим образом: «Если этот самец, несмотря на его яркое оперение и длинный хвост, каким-то образом умудрился не погибнуть в лапах хищника и дожить до половой зрелости, то, следовательно, он обладает хорошими генами, которые позволили ему это сделать. Значит, его стоит выбрать в качестве отца для своих детей: он передаст им свои хорошие гены». Яркость окраски перьев и гребней у птиц, чешуи и хвоста у рыб является надежной характеристикой здоровья. Так, на рыбке корюшке было показано, что чем ярче окраска самца, тем более он привлекателен для самок и тем менее он заражен паразитами. Яркость гребня у петухов напрямую зависит от содержания тестостерона в крови и от степени зараженности паразитами. Чем выше уровень тестостерона, тем ярче гребень, чем больше паразитов, тем он бледнее. Известно, также, что тестостерон угнетает иммунную систему. Следовательно, самцы с ярким гребнем обладают такой эффективной иммунной системой, которая обеспечивает их устойчивость к паразитам, несмотря на высокий уровень тестостерона. Выбирая ярких самцов, самки выбирают хорошие гены для своих потомков. Согласно гипотезе «привлекательных сыновей» логика выбора самок несколько иная. Если яркие самцы, по каким бы то ни было причинам, являются привлекательными для самок, то стоит выбирать яркого отца для своих будущих сыновей, потому что его сыновья унаследуют гены яркой окраски и будут привлекательными для самок в следующем поколении. Таким образом, возникает положительная обратная связь, которая приводит к тому, что из поколения в поколение яркость оперения самцов все более и более усиливается. Процесс идет по нарастающей до тех пор, пока не достигнет предела жизнеспособности. Представим себе ситуацию, когда самки выбирают самцов с более длинным хвостом. Длиннохвостые производят больше потомков, чем самцы с короткими и средними хвостами. Из поколения в поколение длина хвоста увеличивается, потому что самки выбирают самцов не с определенным размером хвоста, но с большим, чем в среднем размером. В конце концов, хвост достигает такой длины, когда его вред для жизнеспособности самца уравновешивается его привлекательностью в глазах самок. Объясняя эти гипотезы, мы старались понять логику действия самок птиц. Может создаться впечатление, что мы слишком много от них ожидаем, что такие сложные расчеты приспособленности им вряд ли доступны. На самом деле, в выборе самцов самки не более и не менее логичны, чем во всем остальном их поведении. Когда животное чувствует жажду, оно не рассуждает, что ему следует попить воды, для того чтобы восстановить водно-солевой баланс в организме – оно идет на водопой, потому что чувствует жажду. Когда рабочая пчела жалит хищника, напавшего на улей, она не вычисляет, насколько этим своим самопожертвованием она повышает совокупную приспособленность своих сестер – она следует инстинкту. Точно так же и самки, выбирая ярких самцов, следуют своим инстинктами – им нравятся яркие хвосты. Все те, кому инстинкт подсказывал иное поведение, все они не оставили потомства. Таким образом, мы обсуждали не логику самок, а логику борьбы за существование и естественного отбора – слепого и автоматического процесса, который, действуя постоянно из поколения в поколение, сформировал все то удивительное разнообразие форм, окрасок и инстинктов, которое мы наблюдаем в мире живой природы.
|
www.biokan.ru
Естественный отбор - Психологос
Естественный отбор – основной движущий фактор эволюции живых организмов.
К мысли о существовании естественного отбора пришли независимо друг от друга и почти одновременно несколько английских натуралистов: В. Уеллс (1813г.), П. Мэтью (1831г.), Э. Блайт (1835, 1837гг.), А. Уоллес (1858г.), Ч. Дарвин (1858-1859гг.).
Но только Дарвин сумел вскрыть значение этого явления как главного фактора эволюции и создал теорию естественного отбора.
Теория Дарвина о происхождении видов в животном мире
Главные положения: виды не являются неизменными сотворенными формами, а представляют собою результат многовекового процесса приспособления животных в борьбе за существование. Особенности организации, благоприятные для добывания пищи, борьбы с соперниками, защиты от врагов, наследственно передаваясь потомству, укрепляются и накапливаются. Таким образом, лучше организованные особи имеют больше шансов пережить более слабых и оставить потомство.
Примеры действия естественного отбора
Пример волка, питающегося различными животными и одолевающего одних силой, других хитростью, третьих - быстротой.
Представим, что самая быстрая добыча, олени, например увеличились в числе вследствие каких-нибудь перемен, происшедших в данной местности, или, наоборот другая добыча уменьшилась в числе, как раз в то время года, когда волки наиболее терпят от недостатка в пище. При таких обстоятельствах самые быстрые и поджарые волки будут иметь больше шансов выжить и, таким образом, сохраниться или быть отобранными.
Обстоятельства, благоприятствующие образованию новых форм посредством естественного отбора
Значительная часть изменчивости и индивидуальные различия, будут, очевидно, являться обстоятельством благоприятствующим. Большое количество особей, увеличивая шансы появления в данный период полезных изменений, может компенсировать меньшую степень изменчивости у отдельной особи и является важным элементом успеха.
Скрещивание играет важную роль в природе, так как поддерживает однообразие и постоянство признаков у особей одного и того же вида или одной и той же разновидности. Оно, очевидно, будет влиять всего действительнее на животных, спаривающихся для каждого рождения, но скрещиванию подвергаются, время от времени все растения и все животные. Даже если это будет происходить через длинные промежутки времени, то происшедшая от этого скрещивания молодь будет настолько превосходить силой и плодовитостью потомство, полученное от продолжительного самооплодотворения, что будет иметь более шансов на выживание и размножение; и таким образом, в конце концов, влияние скрещивания, даже через долгие промежутки времени, окажутся весьма важными.
Продолжительность времени сама по себе не содействует и не препятствует естественному отбору. Продолжительность времени имеет значение лишь настолько (в этом смысле её значение велико), насколько она увеличивает шансы появления благоприятных изменений, их отбора, накопления и закрепления.
Изоляция является также важным элементом в процессе изменения видов посредством естественного отбора. В ограниченном или изолированном ареале, если он не очень велик, органические и неорганические условия жизни будут обычно почти однородными, так что естественный отбор будет стремиться изменить все изменяющиеся особи того же вида в одном и том же направлении. Скрещивание с обитателями окружающих областей будет тем самым также устранено. Хотя изоляция имеет большое значение в образовании новых видов, тем не менее, обширность ареала ещё важнее, особенно в процессе образования видов, которые могли бы сохраниться на долгое время и распространиться. На большом и открытом пространстве не только увеличиваются шансы появления благоприятных изменений благодаря многочисленности особей того же вида, которых может прокормить эта площадь, но и самые условия существования гораздо более сложны вследствие многочисленности уже существующих видов; а если некоторые из этих многочисленных видов изменяться и улучшаться в соответствующей степени, иначе они будут истреблены.
Вымирание, вызываемое естественным отбором
Естественный отбор действует только посредством сохранения изменений, в каком-нибудь смысле полезных и, следовательно, укореняющихся. Вследствие быстрого возрастания численности всех органических существ геометрической прогрессии каждый ареал уже до предела заполнен обитателями, а из этого вытекает, что, так как благоприятствуемые формы будут увеличиваться в числе, то менее благоприятствуемые будут обычно уменьшаться в числе и становиться редкими. Редкость формы - предвестник вымирания. Всякая форма представленная малым числом особей, имеет большие шансы на окончательное исчезновение, вследствие ли значительных климатических колебаний на протяжении года или вследствие временного увеличения числа ее врагов. Виды, наиболее богатые особями, обладают наибольшими шансами для появления во всякий данный период благоприятных изменений. Отсюда виды редки будут во всякий данный период изменятся, и совершенствоваться медленнее и вследствие этого будут побеждены в жизненной борьбе изменившимися и улучшившимися потомками более обыкновенных видов. Из этого вытекает, что, так как с течением времени деятельностью естественного отбора образуются новые виды, то другие должны становится все более редкими и, наконец, исчезать. Формы, всего сильнее конкурирующие с теми, которые изменяются и совершенствуются, пострадают более всего. Каждая новая разновидность или новый вид будет в процессе своего образования сильнее всего подавлять своих ближних родственников и стремиться их истребить. Тот же процесс истребления происходит и у домашних рас вследствие отбора наиболее усовершенствованных форм человеком.

www.psychologos.ru
Виды естественного отбора. Рабочие материалы
Дополнительные сочиненияИз этого урока вы узнаете, что такое естественный обор, каковы его виды. Как естественный отбор влияет на популяции живых организмов? В чем сходство и отличие естественного и искусственного отбора? Что именно отбирается в процессе естественного отбора и как происходит этот процесс? Вы познакомитесь со стабилизирующим, движущим и разрывающим (дизруптивным) отбором, выясните природу полового отбора, открытого Ч. Дарвиным. Возможно, этот урок поможет вам в вашей личной борьбе за существование. Вы узнаете, как влияет естественный отбор на современного человека.
Тема: Эволюционное учение
Урок: Виды естественного отбора
1. Естественный отбор и его виды
Естественный отбор – это основная движущая сила эволюции.
Представление о естественном отборе сильно углубились благодаря современным представлениям генетики и трудам отечественных ученых И. И. Шмальгаузена и С. С. Четверикова (Рис. 1), а также многих их зарубежных коллег.
Рис. 1.
Согласно современным представлениям о естественном отборе, можно выделить три его формы.
2. Движущий отбор
Первая форма естественного отбора – это движущий отбор. Он возникает при смене условий среды и приводит к сдвигу среднего значения проявления признака в популяции под действием факторов среды (рис. 2). Новый признак или его значение должны лучше подходить к изменившимся условиям, чем старые.
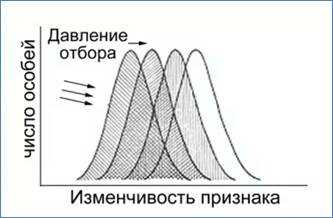
Рис. 2. Схема влияния движущего отбора на значение представленности признака в популяции
Например, при похолодании климата происходит отбор особей с более теплой шерстью.
Классическим примером движущего отбора является эволюция окраски у берёзовой пяденицы. Окраска крыльев этой бабочки имитирует окраску покрытых серой корой деревьев. Загрязнение атмосферы, связанное с выбросами заводов и фабрик, привело к потемнению стволов деревьев. Светлые бабочки на темном фоне стали легко заметны для птиц. С середины XVII века, в популяциях березовой пяденицы стали появляться мутантные темные формы бабочек. Частота этой аллели быстро возрастала, и уже к концу XIX века некоторые городские популяции берёзовой пяденицы почти целиком состояли из темных форм. В то время как в сельских популяциях, где уровень загрязнения был ниже, по-прежнему преобладали светлые формы.
Изменение признака может происходить как в сторону его усиления, так и в сторону ослабления, вплоть до полной редукции. Так, например, произошло исчезновение зрительных органов у кротов и прочих роющих животных, или редукция крыльев у нелетающих птиц и насекомых (см. Рис. 3).


Рис. 3. Примеры длительного воздействия движущего отбора: отсутствие глаз у крота (слева) и крыльев у страуса (справа)
3. Дизруптивный отбор
Второй вид отбора – это дизруптивный (разрывающий) отбор. В этом случае оставляют потомство особи с несколькими крайними вариантами признака, а особи со средним значением признака элиминируются (Рис. 4).
Рис. 4. Схема влияния дизруптивного (разрывающего) отбора на представленность признака среди особей в популяции
Дарвин считал, что дизруптивный отбор ведет к дивергенции, т. е. к расхождению признаков, и служит для поддержания полиморфизма популяции. В ходе дизруптивного отбора из общего светло-жёлтого предка появились две формы бабочек: белая и желтая. Разная окраска приводит к разному нагреву крыльев. Белым бабочкам удобно летать в полдень, а желтым – утром. Светло-жёлтым бабочкам неудобно летать и днем, и утром, поэтому отбор действует как раз против среднего значения признака.
4. Стабилизирующий отбор
Третья форма естественного отбора – это стабилизирующий отбор. Он действует при постоянстве условий внешней среды, путем выбраковывания особей со значительными отклонениями признака (Рис. 5).
Рис. 5. Схема стабилизирующего отбора
Он направлен на сохранение и закрепление среднего значения признака. Например, цветы растений, которые опыляются насекомыми, очень консервативны, то есть их форма редко меняется. Это связано с тем, что насекомые-опылители не могут проникнуть в венчик цветка слишком глубокий, либо слишком узкий (см. видео).
Поэтому гены, которые приводят к таким изменениям строения цветов, не передаются дальше и вытесняются из генофонда.
Благодаря стабилизирующему отбору до наших дней сохранились т. н. живые ископаемые.
6. Живые ископаемые
До наших дней в неизменном виде дожили некоторые виды живых существ, миллионы лет назад бывшие типичными представителями флоры и фауны давно минувшей эпохи.
Например, мечехвосты (см. Рис. 6), древние членистоногие, жившие полмиллиарда лет назад, успешно существуют и сейчас благодаря стабилизирующему отбору. Этот вид почти вдвое старше вымерших уже динозавров.

Рис. 6.
Кистеперая рыба латимерия, предки которой были широко распространены в палеозойскую эру, наглядно показывает, как могло происходить превращение плавников рыб в лапы будущих амфибий.
Стабилизирующий отбор прекратил дальнейшую эволюцию ее конечностей в связи с переходом этих рыб к жизни в глубинах океана (см. видео).
Голосеменное растение – гинкго (см. Рис. 7), широко распространенное в мезозойскую эру, имеет множество переходных признаков, например пластинчатые листья, характерные для древних покрытосеменных.

Рис. 7.
5. Половой отбор
Существует ещё понятие половой отбор. Он не имеет отношения к приведенной классификации, и представляет собой борьбу самцов или самок за возможность оставить потомство. Т. е. это пример внутривидовой борьбы за существование.
Чаще всего, особь просто выбирает себе наиболее сильного и жизнеспособного партнера. Половая конкуренция приводит к возникновению сложных поведенческих механизмов: пению, демонстративному поведению, ухаживанию (см. видео). Нередко между самцами возникают драки, которые могут закончиться увечьем или смертью участников.
Характерные кошачьи вопли по ночам обычно сопровождают именно такие схватки конкурирующих самцов.
Половой отбор способствует половому диморфизму, т. е. различию во внешнем строении самцов и самок. Можно вспомнить, как отличаются петухи и куры, утки и селезни, самцы и самки оленей и моржей (см. видео).
В результате полового отбора, потомство оставляют наиболее сильные, жизнеспособные и здоровые особи. Остальные отстраняются от размножения, и их гены исчезают из генофонда популяции.
Домашнее задание:
1. Что такое естественный отбор? Почему он происходит?
2. Чем отличаются естественный и искусственный отбор?
3. Чем отличаются движущий и стабилизирующий отбор?
4. Что такое разрывающий отбор?
5. Куда направлен естественный отбор?
6. Что такое половой отбор?
7. Какие виды естественного отбора действуют в популяциях человека?
8. Приведите примеры влияния разных видов естественного отбора на популяции живых существ. Можно ли наблюдать действие естественного отбора в природе?
9. Какие эксперименты могут подтвердить или опровергнуть существование естественного отбора?
Дополнительные рекомендованные ссылки на ресурсы сети Интернет
1. Laboratory of Protein Ptysics .
2. Xvatit. com .
3. Afonin-59-bio. narod. ru .
Список литературы
1. Каменский А. А., Криксунов Е. А., Пасечник В. В. Общая биология 10-11 класс Дрофа, 2005.
2. Беляев Д. К. Биология 10-11 класс. Общая биология. Базовый уровень. – 11-е изд., стереотип. – М.: Просвещение, 2012. – 304 с.
3. Биология 11 класс. Общая биология. Профильный уровень / В. Б. Захаров, С. Г. Мамонтов, Н. И. Сонин и др. – 5-е изд., стереотип. – Дрофа, 2010. – 388 с.
4. Агафонова И. Б., Захарова Е. Т., Сивоглазов В. И. Биология 10-11 класс. Общая биология. Базовый уровень. – 6-е изд., доп. – Дрофа, 2010. – 384 с.
dp-adilet.kz
Естественный отбор – направляющий фактор эволюции
Естественный отбор – направляющий фактор эволюции
Естественный отбор – это дифференциальное выживание и размножение особей, которые отличаются друг от друга генетически детерминированными признаками. Более приспособленные к данным условиям среды особи оставляют больше потомков, чем менее приспособленные. Мы можем измерить относительную приспособленность особи долей её потомков среди особей следующего поколения и, следовательно, частотой ее аллелей, которые вошли в генофонд следующего поколения.
Для того, чтобы оценить приспособленность каждого конкретного зайца нам не нужно измерять его скорость бега, его хитрость в ускользании от волка, лисицы и прочих врагов, его способность находить и использовать пищу, его привлекательность для зайчих, нам нужно только знать, сколько он оставил потомков. Из двух зайцев более приспособлен тот, кто оставил больше потомков, кто внес больше своих генов в генофонд следующего поколения зайцев. Таким образом, частота аллелей, которые определяют более высокую приспособленность их носителей, увеличивается в генофонде популяции. Генетический состав популяции меняется. Поскольку мы определили эволюцию, как изменение генетического состава популяции, то мы можем рассматривать естественный отбор как фактор эволюции.
Естественный отбор принципиально отличается от других уже известных вам факторов эволюции – мутационного процесса и дрейфа генов. Мутации возникают случайно. Они не являются и не могут быть изначально адаптивными. При дрейфе генов частоты аллелей в популяциях меняются случайно, безотносительно к тому, насколько хорошо приспособлены носители того или иного аллеля. Естественный отбор определяет жесткую связь между приспособительной ценностью аллелей и их частотой в популяциях. Это единственный фактор, который обеспечивает приспособление всех живых организмов к постоянно меняющимся условиям внешней среды и регулирует гармоничные взаимодействия между генами внутри каждого организма.
Естественный отбор происходит автоматически. Все живые организмы из поколения в поколение проходят суровую проверку по всем мельчайшим деталям их строения, функционирования всех их систем в разнообразных условиях. Только те, кто выдержал эту проверку, оказываются отобранными и дают начало следующему поколению. Дарвин писал: «Естественный отбор ежедневно и ежечасно расследует по всему свету мельчайшие вариации, отбрасывая дурные, сохраняя и слагая хорошие, работая неслышно и незаметно, где бы и когда бы, ни представился к тому случай, над усовершенствованием каждого органического существа по отношению к условиям его жизни, органическим и неорганическим. Мы ничего не замечаем в этих медленных переменах в развитии, пока рука времени не отметит истекших веков».
Эффективность естественного отбора зависит от его интенсивности и запаса наследственной изменчивости, накопленного в популяции. Интенсивность отбора определяется тем, какая доля особей доживает до половой зрелости и участвует в размножении. Чем меньше эта доля, тем больше интенсивность отбора. Если, например, из 10 000 особей в каждом поколении выживают и размножаются только 100 самых крупных, то средний размер особей в этой популяции растет гораздо быстрее, чем в случае более мягкого отбора, когда, например, половина всех особей в популяции участвует в размножении.
Интенсивность отбора варьирует в широких пределах в зависимости от вида и экологической ситуации. Однако она никогда не бывает нулевой. Даже в человеческих популяциях, которые благодаря достижениям современной цивилизации в сильной степени защищены от действия отбора, его интенсивность весьма значительна. Около 30% всех беременностей заканчиваются выкидышами на самых ранних стадиях; 5% новорожденных гибнет при рождении, еще около 3% не доживает до зрелости; 20% людей, доживших до зрелости, не вступают в брак, и 10% вступивших в брак не имеют детей. В природных популяциях микроорганизмов, растений и животных интенсивность естественного отбора гораздо выше.
Однако даже самый интенсивный отбор будет неэффективен, если изменчивость особей в популяции незначительна или она носит ненаследственный характер. Если все особи в популяции совершенно одинаковы, то даже в условиях жесткой борьбы за существование все они имеют равные шансы уцелеть или погибнуть. Жизнь и размножение каждой такой особи зависит от случая. Из поколения в поколение генетический состав этой популяции будет оставаться неизменным.
Изменения не будут накапливаться и в том случае, если изменчивость особей в популяции родителей обусловлена не генетической изменчивостью, а разнообразием средовых условий. Тогда среднее значение признака и потомков особей с крайне высоким и крайне низким проявлением признака будет одинаковым. Такую картину мы всегда наблюдаем в так называемых чистых линиях (гомозиготных по всем генам) растений и животных и в клонах бесполых организмов. Поэтому отбор в чистых линиях и клонах оказывается неэффективным. Сколько бы поколений мы ни отбирали представителей чистых линий с наибольшим или наименьшим проявлением признака, популяционная средняя в последующих поколениях остается неизменной. В генетически гетерогенных популяциях отбор ведет к постепенному смещению средней из поколения в поколение.
Важнейшим свойством естественного отбора является его способность накапливать и совмещать полезные аллели. Отбор действует в каждом поколении, и в каждом поколении он начинается не с нуля, не с «чистого листа», а того состояния популяции, в которое она была приведена в результате предыдущего цикла отбора. Если в какой-то популяции из года в год высокие растения получают преимущество в выживании и размножении, то поколение за поколением в этой популяции растет частота высоких особей, и увеличивается средних размер особей. Результаты действия естественного отбора аккумулируются, накапливаются из поколения в поколение. Все новые и новые аллели возникают в популяции за счет мутационного процесса. Каждая мутация случайна, но ее носители регулярно отбираются, и размножаются в популяции, если она хоть чуть-чуть повышает приспособленность ее носителей. Если критерием приспособленности в данной популяции является высота растения, то отбор подхватывает все аллели, которые тем или иным способом работают на этом признак – увеличивают скорость роста, эффективность использования солнечного света, воды и других ресурсов.
Способностью отбора накапливать и совмещать полезные аллели и удалять вредные обусловлена и постепенность эволюции, и возникновение изумительно сложных органов, форм и функций. Самые сложные приспособления в строении, функциях и поведении живых организмов сформировались постепенно путем систематического отбора случайно возникающих мутаций.
Объектами отбора всегда являются отдельные особи, а не популяции и не виды. Ни одно приспособление, сколь угодно полезное для вида, не сможет быть подхвачено естественным отбором и распространиться в популяции, если оно вредно для конкретных особей.
Вернемся к проблеме взаимопомощи между животными, которую мы обсуждали выше. Долгое время оставалось загадкой, как это свойство могло возникнуть и закрепиться. Представим себе популяцию, состоящую из одних «эгоистов», которые сосредоточены только на своих собственных интересах и не склонны жертвовать собой ради других. Допустим, что в этой популяции появляются редкие мутанты-«альтруисты», которые всегда готова прийти на помощь другим. При этом они ставят под угрозу свою собственную жизнь. В такой ситуации победа в борьбе за жизнь достанется эгоистам, которые пользуются помощью альтруистов, но сами ничего не дают взамен. Гены «альтруистического поведения» будут довольно быстро утрачены. К такому же результату мы придем, если начнем наше рассмотрение с полностью альтруистической популяции. Допустим, что в такой популяции появляется эгоистичный мутант. Он охотно принимает помощь, но сам никому не помогает. Выгоды такого поведения для него и его эгоистичных потомков очевидны. Они получают незаслуженные преимущества в борьбе за жизнь, успешно размножаются и, в конечном счете, вытесняют гены альтруистов из генофонда популяции.
Ключ к разрешению этого парадокса был найден в расширении понятия приспособленности. В современной эволюционной биологии выделяется два компонента приспособленности: непосредственная приспособленность и расширенная. Под прямой приспособленностью особи понимается вклад ее собственных аллелей в генофонд популяции. Чем выше жизнеспособность и плодовитость особи, чем лучше она обеспечивает выживание своих потомков, тем выше ее непосредственная приспособленность. Расширенная приспособленность особи тем выше, чем больше аллелей, идентичных ее аллелям, передается следующему поколению. Особь может повысить свою совокупную приспособленность, повышая своими действиями непосредственную приспособленность своих родственников, которые содержат в своих генотипах те же аллели, что и она. Мать, защищая своих потомков, защищает по существу свои гены, переданные этим потомкам. Каждый из них имеет половину генов, полученных от матери, половину – от отца. Следовательно, коэффициент родства между родителем и потомком равен ½. Отсюда следует, что если ценой своей жизни, мать спасет жизнь двум своим детям, то ее совокупная приспособленность не снизится. Но ведь точно такая же степень родства связывает между собой братьев и сестер. Следовательно, по генетико-эволюционным соображениям братья и сестры должны проявлять друг к другу такую же степень заботы, как и мать к своим детям.
Такое понимание приспособленности объясняет возникновение и распространение альтруизма и взаимопомощи в популяциях животных. Помогая родственникам, животные способствуют распространению своих генов, в том числе и генов, ответственных за формирование альтруистического поведения. Не следует думать, однако, что животные действительно скрупулезно подсчитывают коэффициент родства, который связывает их с той или иной особью в популяции и на основании этих подсчетов принимают решение как им поступать – эгоистично или альтруистично – в каждом конкретном случае. Когда возникает угроза жизни ребенка, мать инстинктивно бросается на его защиту. Те матери, которые так не поступали, остались без потомков, аллели этих матерей были бесследно утеряны и не оставили следов в эволюции. Возникновение и распространение взаимопомощи между родственниками является, таким образом, автоматическим следствием естественного отбора.
1. Что такое приспособленность особи? Как можно оценить относительную приспособленность особей в природе и в лабораторных экспериментах?
2. От чего зависит эффективность отбора? Где больше сходство между родителями и потомками – в чистых гомозиготных линиях или в гетерогенных популяциях? Почему отбор в чистых линиях не приводит к смещению среднего значения признака в ряду поколений?
3. Одно из возражений против Дарвиновской теории эволюции заключается в том, что отбор только отсекает заведомо неприспособленные отклонения и поэтому не может создавать ничего нового. Опровергните это возражение.
4. Как естественный отбор может способствовать возникновению и развитию альтруистического поведения и взаимопомощи у животных?
evoldar.com