Цитокинез растительной и животной клетки. Как происходит цитокинез в клетках растений и животных
Цитокинез растительной и животной клетки
В телофазе перед цитокинезом увеличивается биосинтез мембран, которые необходимы для того, чтобы покрыть обе дочерние клетки. Вновь синтезированные мембраны до момента разделения клетки образуют на ее поверхности пузырьки, которые затем встраиваются в плазмалеммы дочерних клеток. Перетяжка становится все более глубокой, и в результате в конце концов одна клетка разделяется на две (цитокинез). Обе дочерние клетки диплоидны. Однако не всегда деление ядра сопровождается разделением клетки. Поэтому помимо телофазы (при полном делении клетки) и выделяют цитокинез.
После митоза в течение нескольких часов дочерние клетки связаны между собой небольшим остаточным тельцем, образованным непрерывными микротрубочками и электроноплотным материалом матрикса. Остаточное тельце покрыто плазмалеммой. Есть все основания считать, что сила, необходимая для разделения клеток, возникает в результате взаимного скольжения актиновых и миозиновых филаментов.
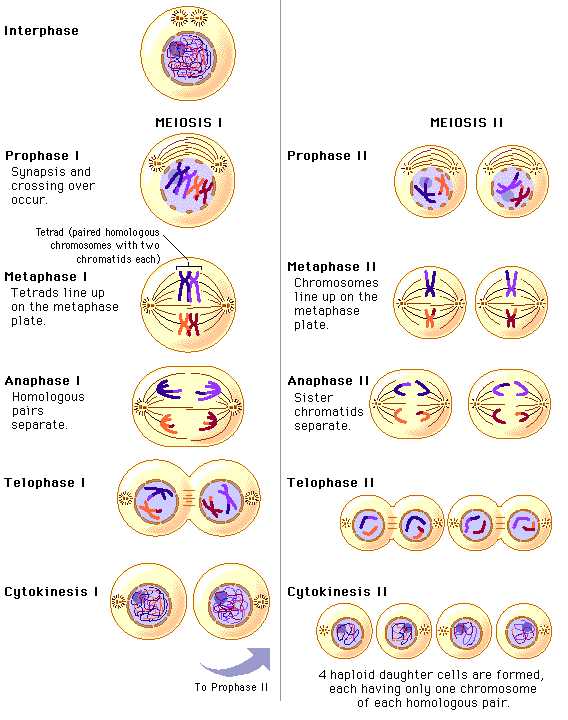
Фазы мейоза
У организмов, размножающихся половым путем, имеется две категории клеток: диплоидные и гаплоидные. К первым относятся соматические и предшественницы половых клеток, ко вторым — зрелые половые (гаметы). Уменьшение количества хромосом в два раза достигается благодаря мейозу. Он включает в себя два последовательных деления. После слияния гамет возникает новый одноклеточный диплоидный организм (зигота), который несет не просто сумму признаков своих родителей, а является индивидуумом с присущими только ему свойствами.
При дальнейшем митотическом делении зиготы образуются диплоидные же клетки, содержащие по два экземпляра каждой хромосомы, которые называются гомологичными. Гомологичные хромосомы, имеющие одинаковую длину и одинаковое расположение центромер, содержат одинаковое количество генов, а эти гены имеют одну и ту же линейную последовательность. Каждая из пары гомологичных хромосом диплоидного организма происходит либо из ядра спермия, либо из ядра яйцеклетки.
При образовании гамет в зрелом организме в результате мейоза в каждую дочернюю клетку от всех пар гомологичных хромосом попадает лишь по одной из них. Это становится возможным потому, что при мейозе происходит лишь одна репликация ДНК, за которой следуют два последовательных деления ядер (мейоз I и II) без повторного синтеза ДНК. В результате из одной диплоидной образуются четыре гаплоидные клетки.
Напомним, что перед началом мейоза в интерфазе клетка прошла обычные фазы Gx, S и G2, так что стала тетраплоидной. Иначе говоря, произошла репликация ДНК и бел-ков-гистонов хромосом, а сестринские хроматиды при этом остались связанными своими центромерами, так что в ядре имеется по четыре набора каждой хромосомы. Увеличена масса клетки и ее органелл.
Каждое из двух делений мейоза (деления I и II) имеет свои отличительные черты.
Особенность деления I состоит в необычном и сложном прохождении профазы (профаза I). Она подразделяется на несколько стадий: пролептонему, леп-тонему, зигонему, пахинему, диплонему и диакинез.
Во время пролептонемы (от греч. pro — период, leptos — тонкий, пета — нить) происходит значительная, но не полная спирализация хромосом. Ядерная оболочка сохраняется, ядрышко не распадается. Поэтому во время профазы мейоза возможны синтезы некоторых РНК и белков. За счет этих синтезов в половых клетках (особенно в женской) создаются запасы веществ, которые будут необходимы для оплодотворения и ранних стадий развития зародыша.
Во время лептонемы хромосомы еще больше спирализуются, и в ядре становятся видными тонкие нитевидные d-хромосомы (их 46, т. е. два набора). Подчеркнем, что каждая гомологичная хромосома уже реплицировна и состоит из двух сестринских хроматид. Каждая хромосома представляет собой тонкую фибриллу, состоящую из осевой белковой нити, к которой прикрепляется хроматин сестринских хроматид (петли ДНК). Хромосомы с помощью белковых скоплений — прикрепительных дисков — закреплены обоими своими концами на внутренней мембране ядерной оболочки (ядерная оболочка сохраняется, ядрышко хорошо видно).
Во время зигонемы (от греч. zygon — парный) гомологичные диплоидные хромосомы выстраиваются рядом, обвивают друг друга, укорачиваются и сцепляются между собой (конъюгация). Образуются так называемые тетраплоидные биваленты (от лат. Ы — двойной, valens — сильный). Напомним, что каждая диплоидная хромосома из одного бивалента происходит либо от отца, либо от матери. Половые хромосомы располагаются около внутренней ядерной мембраны. Область, занятая ими, называется половым пузырьком.
В зигонеме гомологичные d-хромосомы выстраиваются рядом, сближаются, между ними образуются специализированные синаптонемальные комплексы (от греч. synapsis — связь, соединение), которые представляют собой белковые структуры. При небольшом электронно-микроскопическом увеличении синаптонемальный комплекс выглядит в виде двух электронно-плотных полос, разделенных светлой полосой. При большом увеличении в комплексе видны две параллельные боковые белковые нити длиной 120—150 нм и толщиной 10 нм каждая, соединенные тонкими поперечными полосами размерами около 7 нм, по обе стороны от них лежат d-хромосомы. Их ДНК формирует множество петель.
В центре комплекса проходит осевой элемент толщиной 20—40 нм. Синаптонемальный комплекс удачно сравнивают с веревочной лестницей, стороны которой образованы гомологичными хромосомами. Именно в результате этого гомологичные хромосомы сцепляются между собой и образуют биваленты, 46 d-хромосом образуют 23 бивалента. Каждый бивалент состоит из двух d-хромосом, т. е. из четырех хроматид.
К концу зиготены каждая пара гомологичных хромосом связана между собой с помощью синаптонемальных комплексов. Лишь половые хромосомы X и Y конъюгируют неполностью, т. к. они неполностью гомологичны.
Пахинема (от греч. pahys — толстый) продолжается не менее, чем несколько суток. Процессы развертываются постепенно. Хромосомы несколько укорачиваются и утолщаются. Между хроматидами материнского и отцовского происхождения в нескольких местах возникают соединения — хиазмы (от греч. chiasma — перекрест), или рекомбинантные узелки. Они представляют собой белковые комплексы размерами около 90 нм. В области каждой хиазмы происходит обмен соответствующих участков гомологичных хромосом — от отцовской к материнской и наоборот. Этот процесс называют кроссинговером (от англ. crossing-over — перекрест). Таким образом, кроссинговер обеспечивает многочисленные генетические рекомбинации. В каждом биваленте человека в профазе I кроссинговер происходит в среднем в двух-трех участках. Количество рекомбинантных узелков равно количеству перекрестков.
По окончании кроссинговера хроматиды разъединяются, но остаются связанными в области хиазм. Наступает стадия диплонемы.
В диплонеме (от греч. diploos — двойной) синаптонемальные комплексы распадаются, конъюгировавшие хромосомы раздвигаются, и гомологичные хромосомы каждого бивалента отодвигаются друг от друга, но связь между ними сохраняется в зонах хиазм.
Между диплонемой и диакинезом нет четкой морфологической границы, равно как и разграничений во времени. В диакинезе продолжается конденсация хромосом, они отделяются от нуклеолеммы, но гомологичные d-хромосомы продолжают еще оставаться связанными между собой хиазмами, а сестринские хроматиды каждой d-хромосомы — центромерами. Благодаря наличию нескольких хиазм биваленты образуют петли. В это время разрушаются ядерная оболочка и ядрышки. Реплицированные центриоли направляются к полюсам, образуется веретено деления.
Вследствие сильно затянутой диплонемы профаза мейоза очень длительна. При развитии спермиев она может длиться несколько суток, а при развитии яйцеклеток — в течение многих лет. Половые клетки в профазе мейоза называются гаметоцитами первого порядка (первичными га-метоцитами, гаметоцитами I).
Метафаза I напоминает аналогичную стадию митоза. Хромосомы устанавливаются в экваториальной плоскости, образуя метафазную пластинку.
В анафазе I хиазмы распадаются, гомологичные хромосомы отделяются друг от друга и расходятся к полюсам. Центромеры этих хромосом не реплицируются- сестринские хромосомы не расходятся
В телофазе I формируется ядерная оболочка и ядрышко, образуется и углубляется борозда деления, происходит карикенез.
Интерфаза II очень короткая. ДНК не реплицируется.
Профаза II не длительная, и коньюгация хромосом при этом не наступает.
В метафазе II 23 хромосомы выстраиваются в области экватора.
В анафазе II ДНК в области центромеры реплицируется, хромосомы расходятся к полюсам клетки.
В телофазе II образуются две дочерние клетки.
Итак, в результате двух последовательных делений мейоза образуются 4 клетки, каждая из которых несет гаплоидный набор хромосом.
Как известно, клетки не возникают сами по себе, а образуются только при делении других. После деления во вновь образованной клетке не всегда сразу существуют все системы, обеспечивающие ее специфическую функцию. Должно пройти некоторое время, чтобы сформировались все органеллы и были бы синтезированы все необходимые ферменты. Этот отрезок времени называется созреванием. Созревание клетки осуществляется на основе уже сложившейся ее полной детерминации.
Большинство клеток, однако, погибает тогда, когда проявляются особые естественные генетические механизмы. Генетически запрограммированную клеточную гибель называют апоптозом. Механизм возникновения апоптоза весьма сложен. Каждая клетка несет в хромосомах гены, которые могут запускать синтез ферментов, стимулирующих ее к делению. Есть также гены, которые обеспечивают синтез ферментов, препятствующих делению. Пока клетка функционирует, эти синтезы уравновешены.
Для поддержания жизненного равновесия клетка должна также получать сигналы от других клеток, нередко другого вида. Обычно в качестве сигнальных выступают специфические молекулы олигопептидов. Поскольку они поддерживают жизнь клеток, их назвали цитокинами.
В жизненном цикле многих видов клеток наступает момент, когда функциональные их возможности исчерпываются. У таких клеток нарушается чувствительность к цитокинам и изменяется соотношение активности генов, обеспечивающих внутреннее равновесие. Гены, обеспечивающие размножение клетки, блокируются. Напротив, гены, обеспечивающие синтез литических ферментов, стимулируются. Последние поступают в ядро и лизируют хроматин.
Хромосомы распадаются, синтезы в клетке прекращаются. Внешние проявления такой гибели клеток разнообразны и известны давно. Их называли пикнозом (сморщивание ядра), хроматолизисом (снижение окрашиваемости ядра), кариорексисом (распад ядра на части). Лишь недавно было показано, что это лишь частные проявления апоптоза. Вслед за гибелью ядра разрушается и цитоплазма. Остатки фагоцитируются макрофагами. Материал погибших клеток перерабатывается макрофагами и может выводиться ими на поверхность. В таком случае этот материал может опять использоваться другими клетками. Вокруг клеток, подвергшихся апоптозу, воспалительный процесс не возникает, и жизнедеятельность ткани, часть которой составляли погибшие клетки, продолжается без нарушений.
studfiles.net
это процесс деления клетки. А что вы о нем знаете?
Цитокинез – это процесс деления эукариотической клетки. Цитокинез был одним из первых событий клеточного цикла, наблюдаемых с помощью простых клеточных биологических методов, однако молекулярная характеристика цитокинеза замедлилась благодаря особой устойчивости к биохимическим подходам in vitro. В то время как результат цитокинеза одинаков во всех делящихся клетках, механизм деления варьируется в разных крупных эукариотических царствах. Например, дрожжи и животные используют сократительное кольцо, которое проникает в середину клетки, чтобы разделить ее, в то время как клетки строят новую клеточную стенку наружу, к коре. Как и следовало ожидать, существуют некоторые сходства в молекулах, участвующих в цитокинезе одноклеточных и клетках животных, но на первый взгляд цитокинез в растительной и животной клетке кажется совершенно разным. Однако в последние годы стало ясно, что базовые процессы совершенно одинаковы у клеток растений, простейших и животных.
Один из видов цитокинеза – это митоз, который подразделяется на несколько стадий: профаза, метафаза, анафаза и телофаза. Ниже приведены особенности каждой фазы цитокинеза.
Профаза
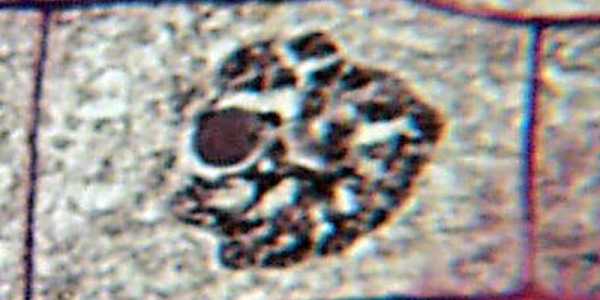
Профаза знаменуется бурными биохимическими изменениями, по результатам которых клетка приходит в состояние, после которого начинается непосредственно деление. Во время профазы хромосомы собираются в центре клетки, а затем удваиваются, предоставляя генетический материал для обеих новообразованных дочерних клеток. Обычно их не видно в микроскоп, однако в тот момент они становятся хорошо различимыми в оптический микроскоп. Также в это время пропадает ядрышко. К середине профазы полностью останавливается транскрипционная активность. Особенности строения клеток таковы, что на ранних стадиях цитокинеза у клеток с крупными хромосомами он замедляется и может растягиваться на несколько часов, в то время как в клетках существ с мелкими хромосомами (например, млекопитающих) она длится около 15 минут. По прошествии этого времени начинается деление тела эукариотической клетки.
Метафаза
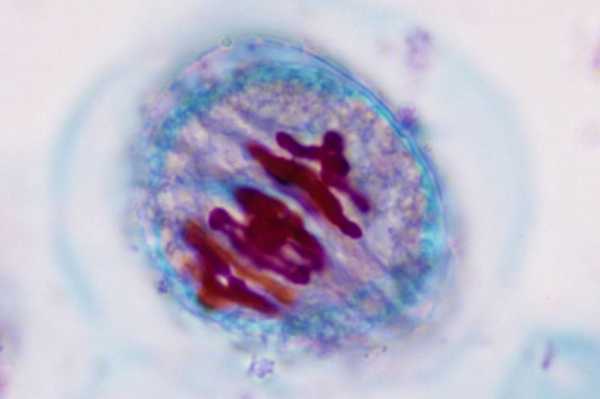
Метафаза цитокинеза – это стадия деления клетки, в которой хромосомы выходят в экваториальную плоскость клетки. Микротрубочки на этой фазе обновляются особенно активно. Хромосомы в клетке располагаются так, чтобы ранее упомянутым микротрубочкам было удобно к ним прикрепиться. Сестринские хроматиды разделяются, но не расходятся, останавливаемые центромерами. Из-за особенностей строения клетки метафаза может завершиться только после того, как клетке пошлет сигнал комплекс стимуляции анафазы. Таким образом, если разрушить веретено деления, хромосомы не смогут перейти к анафазе до тех пор, пока вредоносное воздействие не будет полностью устранено. Такой метод исследования часто употребляется генетиками для того, чтобы создать клетки, часами находящиеся в метафазе, которые потом используют для исследований. Молекулярные механизмы этого действия пока остаются загадкой, но на данный момент над раскрытием их тайн успешно работают ученые.
Анафаза
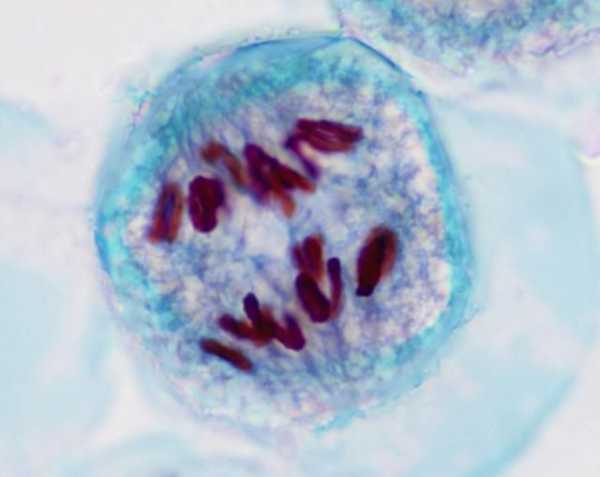
За метафазой следует анафаза. Для цитокинеза это одновременно самая ключевая и самая короткая стадия, во время которой сестринские хроматиды расходятся к краям клетки, образуя дочерние хромосомы. Несмотря на то, что анафаза является самой короткой стадией, она подразделяется на множество фаз. Эти стадии контролирует упомянутый ранее комплекс стимуляции анафазы. Во время анафазы хромосомы расходятся в две новые клетки. Хроматиды каждой хромосомы разгоняются на противоположные стороны клеток, образуя две новые дочерние клетки. Каждая сторона клетки начинает обладать полным набором хромосом. Анафаза важна, потому что именно она помогает ДНК разделить на две части, идя в обе стороны клетки. Она гарантирует, что следующий ген сможет выполнить свою функцию. Если этого не произойдет, тогда не будет двух разных ДНК для следующего процесса
Телофаза
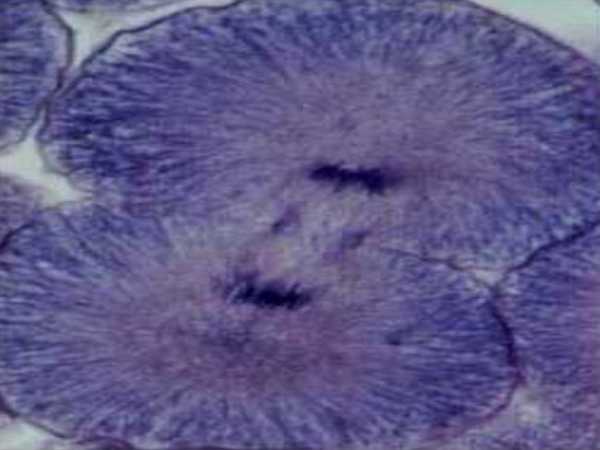
Телофаза – завершающая часть деления клетки. Ее название происходит от латинского слова telos, что означает конец. На этом этапе сестринские хроматиды достигают противоположных полюсов. Маленькие ядерные везикулы в клетке начинают перестраиваться вокруг группы хромосом на каждом конце. По мере того, как ядерная оболочка реформируется путем связывания с хромосомами, в одной клетке создаются два ядра. Телофаза также отмечается растворением микротрубочек кинетохора и продолжением удлинения полярных микротрубочек. По мере преобразования ядерных оболочек хромосомы начинают разлагаться и становиться более рассеянными. После того, как все процессы завершены, две новые клетки начинают функционировать, как ни в чем не бывало.
Как мы увидели, цитокинез является сложным, но в то же время понятным и завораживающим процессом. Ученые до сих пор изучают особенности строения клетки.
fb.ru
Биология для студентов - 15. Цитотомия бактериальных, растительных и животных клеток
Цитотомия (цитокинез) – деление цитоплазмы. Оно обычно следует за телофазой и ведёт к периоду G1 интерфазы. При подготовке к цитокинезу клеточные органеллы вместе с хромосомами равномерно распределяются по двум полюсам телофазной клетки; при этом увеличивается биосинтез фосфолипидов для мембран, которые необходимы, чтобы покрыть обе дочерние клетки.
В животной клетке под плазмалеммой кольцом на том уровне, на котором прежде располагался экватор веретена, активируются элементы цитоскелета – актиновые микрофиламенты. Рядом с ними полимеризуется миозин. Актино-миозиновое кольцо сжимается, и возникает перетяжка плазмалеммы – непрерывная борозда, опоясывающая клетку по экватору. В конце концов клеточные мембраны в области борозды смыкаются, полностью разделяя две клетки.
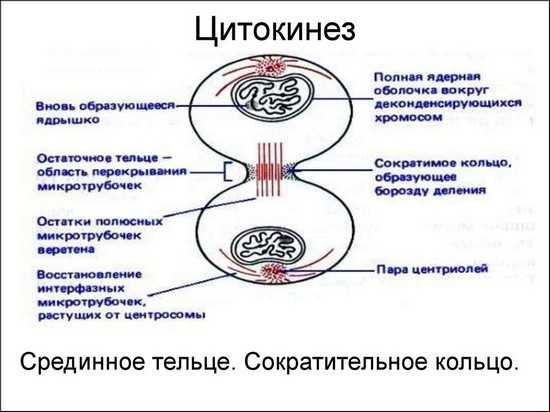
В растительных клетках нити веретена во время телофазы сохраняются только в области экватора, где они сдвигаются к периферии клетки. Их число увеличивается, и они образуют боченковидное тельце – фрагмопласт. В эту область перемещаются также микротрубочки, рибосомы, митохондрии, ЭПС и АГ. АГ образует множество мелких пузырьков. Пузырьки появляются сначала в центре клетки, а затем, направляемые микротрубочками, перемещаются и сливаются друг с другом, образуя клеточную пластинку, расположенную в плоскости экватора. Содержимое пузырьков участвует в построении новой срединной пластинки и стенок дочерних клеток, а из их мембран образуются новые наружные клеточные мембраны. Клеточная пластинка, разрастаясь, в конце концов сливается со стенкой родительской клетки и полностью разделяет две дочерние клетки. Новообразованные клеточные стенки называются первичными; в дальнейшем они могут дополнительно утолщаться за счёт отложения целлюлозы и других веществ (напр., лигнина и суберина), образуя вторичную клеточную стенку. В определённых участках клетки пузырьки клеточной пластинки не сливаются, так что между соседними дочерними клетками сохраняется контакт. Эти цитоплазматические каналы выстланы клеточной мембраной и образуют плазмодесмы.
vseobiology.ru
Митоз — урок. Биология, Общие биологические закономерности (9–11 класс).
Митоз — процесс непрямого деления соматических клеток эукариот, в результате которого из одной диплоидной материнской клетки образуются две дочерние с таким же набором хромосом.
Подготовка клетки к митозу происходит в интерфазу: удваивается ДНК, накапливается АТФ, синтезируются белки веретена деления.
Митоз ключает в себя два процесса: кариокинез (деление ядра) и цитокинез (деление цитоплазмы).
Выделяют четыре фазы митоза: профазу, метафазу, анафазу и телофазу.
Обрати внимание!
В схемах деления гаплоидный набор хромосом обозначают буквой n, а молекул ДНК (т.е. хроматид) — буквой с. Перед буквами указывают число гаплоидных наборов:
1n2с — гаплоидный набор удвоенных хромосом,
2n2с — диплоидный набор одиночных хромосом,
2n4с — диплоидный набор удвоенных хромосом.
Пример:
В клетках человека гаплоидный набор составляют \(23\) хромосомы. Значит, запись 2n2с обозначает \(46\) хромосом и \(46\) хроматид, а 2n4с — 46 хромосом и 92 хроматиды и т.д.
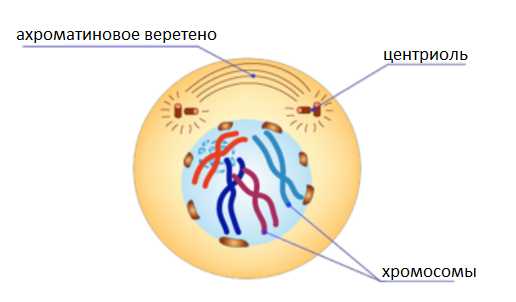
Профаза.
В ядре молекулы ДНК укорачиваются и скручиваются (спирализуются), образуя компактные хромосомы.
Каждая хромосома состоит из двух молекул ДНК (двух хроматид), соединённых центромерой.
Ядерная оболочка распадается.
Хромосомы неупорядоченно располагаются в цитоплазме.
Растворяются ядрышки.
Начинает формироваться веретено деления, часть нитей которого прикрепляется к центромерам хромосом.
В животной клетке центриоли удваиваются и начинают расходиться.
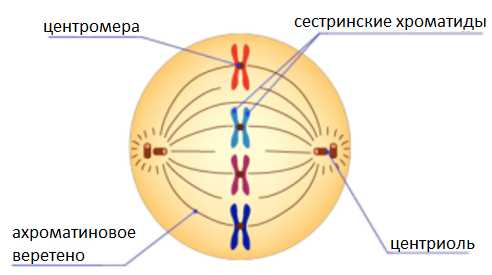
Метафаза.
Хромосомы располагаются на экваторе клетки, образуя метафазную пластинку.
Хроматиды соединены в области первичной перетяжки с нитями веретена деления.
Центриоли располагаются у полюсов клетки.
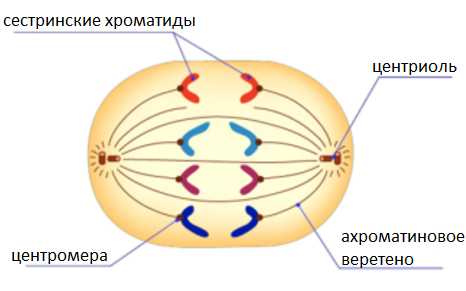
Анафаза.
Каждая хромосома, состоящая из двух хроматид, разделяется на две идентичные дочерние хромосомы.
Дочерние хромосомы растягиваются нитями веретена деления к полюсам клетки.
У каждого полюса оказывается одинаковый генетический материал.
Телофаза.
Хромосомы раскручиваются.
Вокруг хромосом начинают формироваться ядерные оболочки.
В ядрах появляются ядрышки.
Нити веретена деления разрушаются.
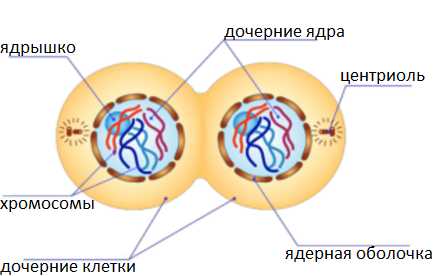
На этом кариокинез завершается. Происходит цитокинез — разделение цитоплазмы
Цитокинез животной клетки
Митоз у растений:
1 — профаза, 2 — метафаза, 3 — анафаза, 4 — телофаза.
Биологическое значение митоза.
В результате митоза образуются генетически одинаковые дочерние клетки с тем же набором хромосом, что был у материнской клетки. Сохраняется преемственность в ряду клеточных поколений.
Источники:
Каменский А.А., Криксунов Е.А., Пасечник Е.В. Биология 10-11класс М.: Дрофа.2005. с.77.
Каменский А.А., Криксунов Е.А., Пасечник Е.В. Биология 9класс М.: Дрофа.
www.yaklass.ru
Клеточное деление цитокинез - Справочник химика 21
| Рис. 13-1. Четыре последовательные фазы клеточного цикла типичной эукариотической клетки. После фазы М, которая состоит в делении ядра (митоз) и цитоплазмы (цитокинез) дочерние клетки вступают в интерфазу нового цикла. Интерфаза начинается с фазы С1, в которой возобновляются интенсивные биосинтетические процессы, резко замедленные во время митоза. Фаза 8-это период синтеза ДНК она заканчивается, когда содержание ДНК в ядре удвоится и хромосомы полностью реплицируются (теперь каждая хромосома состоит из двух идентичных сестринских хроматид ). Затем клетка вступает в фазу Сг, которая продолжается до начала митоза, т.е. фазы М. В фазе М удвоившиеся хромосомы конденсируются и становятся хорошо видимыми в световой микроскоп. Ядерная оболочка разрушается (исключение составляют одноклеточные эукариоты, например дрожжи, - у них она остается интактной) сестринские хроматиды расходятся и формируют два новых ядра, а цитоплазма делится с образованием двух дочерних клеток, имеющих по одному ядру. Процесс цитокинеза завершает фазу М, и начинается интерфаза следующего клеточного цикла. На рисунке представлен типичный 24-часовой цикл, однако длительность клеточного цикла у высших эукариот сильно варьирует она может быть короче 8 часов, а у взрослых животных - больше года, причем различия в основном зависят от |  |
Большая часть компонентов клетки синтезируется на протяжении всего интерфазного периода между последовательными митозами. Это затрудняет вьщеление различных стадий в интерфазе растущих клеток. Исключение составляет лишь синтез ДНК, поскольку ДНК клеточного ядра реплицируется в определенный период, составляющий лишь часть интерфазы. Этот период был назван фазой S (от слова synthesis) клеточного цикла. Вторая четко выраженная стадия цикла-это, конечно, фаза клеточного деления, включающая деление ядра (митоз), а затем и цитоплазмы (цитокинез) фазу клеточного деления назвали фазой М (от слова mitosis). Период между фазой М и началом синтеза ДНК обозначают как фазу Gj (от англ. gap-промежуток), а период между завершением синтеза ДНК и последующей фазой М-как фазу Gj. Таким образом, интерфаза включает фазы Gj, S и Ог. Она обычно занимает не менее 90% времени всего клеточного цикла. Например, у быстро делящихся [c.139]
Телофаза. Два набора хромосом группируются у противоположных полюсов веретена. Здесь они начинают раскручиваться и удлиняться, приобретая форму интерфазных хромосом. Вокруг каждого набора хромосом образуется ядерная мембрана, и вновь возникают ядрышки. Клеточное деление (цитокинез) также полностью завершается на этой стадии. [c.26]
Хотя клетки проводят большую часть своей жизни в интерфазе и только изредка находятся в фазе деления, большинство ранних работ по клеточному циклу было посвящено анализу именно этих коротких периодов клеточного деления (митоза и цитокинеза)-в основном потому, что они доступны для прямого микроскопического исследования. Относительно недавно благодаря использованию более тонких непрямых методов мы сравнительно многое узнали и об интерфазном периоде. В этой главе мы опишем некоторые методы, применяемые в настоящее время для исследования клеточного цикла, рассмотрим регуляцию этого цикла и познакомимся с важнейшими событиями, происходящими в каждой из его фаз. Хотя наши знания о молекулярных основах клеточного цикла довольно фрагментарны, там, где это возможно, мы постараемся обсудить и вероятные механизмы интересующих нас процессов. [c.139]
Прежде чем обсуждать механизмы клеточного деления, необходимо дать самое общее представление о различных стадиях митоза и цитокинеза в типичной клетке высшего организма. [c.175]
Во время митоза микротрубочки и пучки актиновых микрофиламентов, составляющие цитоскелет, распадаются на субъединицы, которые, вероятно, используются для построения машин митоза и цитокинеза соответственно. Остающиеся в клетках промежуточные филаменты не используются для клеточного деления и не распадаются. Вместо этого в большинстве клеток сеть промежуточных филаментов, окружающих интерфазное ядро, в митозе удлиняется, чтобы охватить оба дочерних ядра, после чего перетяжка делит ее на две части (рис. 11-60). [c.190]
Еще до начала разделения материнской клетки на клеточную пластинку начинают откладываться микрофибриллы целлюлозы. При цитокинезе каждый протопласт выделяет новую первичную оболочку, окружая себя ею. По окончании цикла деления клетка переходит в состояние, называемое рост растяжением . [c.28]
Косвенным свидетельством того, что органеллы не могут образовываться без предсуществующих копий, служат особенности клеточного деления у некоторых водорослей, имеющих только один хлоропласт или только один аппарат Гольджи. Органелла, присутствующая в единственном экземпляре, перед цитокинезом расщепляется у них пополам, и по одной половинке старой органеллы попадает затем в каждую дочернюю клетку (см. рис. 9-63). Сходный механизм действует при дупликации и сегрегации центриолей в животных клетках (см. рис. 11-19). По-видимому, при надобности может найтись способ упорядоченного наследования любого клеточного компонента. [c.195]
Наблюдение за цитологическими изменениями, происходящими в культуре, позволяет предсказать тот момент, когда большая часть популяции вступит в определенную фазу клеточного цикла. Например, во время первого клеточного деления важно определить, когда большая часть популяции вступает в стадию цитокинеза и S-фазу, и, следовательно, клетки становятся компетентными для трансформации агробактериями. [c.150]
Цитокинез-процесс собственно клеточного деления-привлекает внимание ученых на протяжении более чем ста лет. Как уже отмечалось, предложены всевозможные объяснения цитокинеза задача состоит в том, чтобы решить, какое из них является верным. Рассмотрим следующие три гипотезы цитокинеза. [c.257]Мы мало знаем о том, с какими изменениями иа молекулярном уровне связана детерминация. Представляется вероятным, что мы здесь имеем дело со стабильными состояниями генной экспрессии, которые не изменяются даже после клеточного деления. Трудно представить себе, каким образом различия в активности генов могут сохраняться в ходе репликации ДНК и Последующего цитокинеза, ио передача дифференцированного состояния через повторные клеточные деления хорошо установлена для животных клеток. [c.477]
ВИЯХ, было выделено и идентифицировано как 6-фурфуриладе-нин. Из-за его высокой активности при индуцировании клеточного деления (цитокинеза) в сердцевине табака (в присутствии ауксина) это вещество было названо кинетином. [c.295]
Мужской пронуклеус сливается с женским пронуклеусом. В этом и состоит подлинный акт оплодотворения. Обрузую-щееся в результате новое ядро содержит два набора хромосом — один от яйцеклетки и один от спермия. Клетка теперь стала диплоидной и ее назьшают зиготой. Новое ядро немедленно делится митотически. А затем зигота претерпевает цитокинез (клеточное деление) с образованием двух диплоидных клеток. [c.87]
Мы начнем главу с обсуждения координации и регулирования этих двух взаимосвязанных циклов. Мы рассмотрим механизмы, благодаря которым в период между двумя клеточными делениями вся ядерная ДНК обязательно удваивается, причем только один раз, и увидим, как события хромосомного цикла скоординированы с событиями цитоплазматического цикла. Затем речь пойдет о регуляции клеточного деления у многоклеточных животных факторами внеклеточной среды этот вопрос сушественно прояснился в результате последних успехов в изучении проблемы рака. И наконец, мы обсудим молекулярные механизмы, ответственные за митоз и цитокинез. Для осуществления этих двух процессов необходимо, чтобы центросома (разд. 13.5.2) надежно наследовалась и точно удваивалась для формирования двух полюсов митотического веретена этот центросомныи цикл можно рассматривать как гретий компонент клеточного цикла. [c.394]
Основная стратегия деления клеток у зукариотических организмов удивительно постоянна. Первые пять стадий фазы М составляет митоз, шестой является цитокинез. Эти шесть стадий образуют динамическую последовательность, сложность и красоту которой трудно оценить по описаниям или по серии статических изображений. Описание митоза основано на наблюдениях двоякого рода на результатах световой микроскопии живой клетки (нередко в сочетании с микрокиносъемкой) и на данных световой и электронной микроскопии фиксированных и окрашенных клеток. Различные стадии клеточного деления кратко описаны на схеме 13-1. Пять стадий митоза - профаза, прометафаза, метафаза, анафаза и телофаза - осуществляются в строго определенном порядке цитокинез начинается во время анафазы и продолжается до конца митотического цикла (рис. 13-43). Световые микрофотографии деления типичной животной и типичной растительной клеток приведены на рис. 13-44 и 13-45 соответственно. [c.439]
Процесс клеточного деления состоит из деления ядра (митоз) и следующего за ним деления цитоплазмы (цитокинез). Митоз характеризуется образованием высокоупорядоченного биполярного веретена, состоящего из микротрубочек и связанных с ними белков микротрубочки организуются двумя митотическими центрами на противоположных полюсах веретена. Хромосомы конденсируются во время профазы. Затем появляются кинетохорные нити они начинают взаимодействовать с полюсными нитя.ыи веретена, после того как ядерная оболочка растворяется в прометафазе. В метафазе в результате действия сил, тянущих кинетохорные нити к полюсаы, хромосомы выстрой- [c.196]
Большинство клеток высших растений окружено жесткой клеточной стенкой, и поэтому механизм цитокинеза существенно отличается от только что описанного для животных клеток. Вместо образования двух дочерних клеток путем их отшпуровки с помощью сократимого кольца, лежащего под поверхностью клетки, цитоплазма разделяется здесь в результате образования новой стенки на границе между дочерними клетками. Образующаяся перегородка точно определяет относительное положение двух новых клеток в растении. Из этого следует, что ориентация плоскостей клеточного деления и увеличение размеров клеток определяют форму растения (см. гл. 20). [c.460]
Процесс клеточного деления состоит из деления ядра (митоз) и слеоующего за ним деления цитоплазмы (цитокинез). Митоз начинается с профазы - переходного периода, когда расщепление центросомы приводит к образованию двух полюсов веретена, организующего в дальнейшем распределение ядерного материала. В это же время начало фазы М сопровождается заметным усилением фосфорилирования определенных белков. Видимо, в результате этого в митотической клетке создается необычайно динамичная система микротрубочек. После разрушения ядерной оболочки в прометафазе кинетохоры конденсированных хромосом могут захватываться и стабилизироваться группами микротрубочек, в большом числе отходящих от обоих полюсов веретена. Эти микротрубочки тянут кинетохоры к противоположным полюсам, и в результате хромосомы располагаются во время метафазы по экватору веретена. В анафазе это натяжение внезапно ослабевает, когда сестринские хроматиды отделяются друг от друга и расходятся к разным полюсам. В добавление к этому часто раздвигаются и оба полюса. В конечной стадии митоза, телофазе, вокруг каждой группы разделившихся хромосом вновь формируется ядерная оболочка, когда белки, фосфорилированные в начале фазы М, вновь дефосфорилируются [c.467]
Клеточное деление заканчивается разделением цитоплазмы (цитокинезом) хромосомы деконденсируются. и на них возобновляется синтез РНК. По-видимому, цитокинез у столь различных эукариотических организмов, как животные, растения и грибы, направляется организованными пучками актиновых филаментов. Крупные органеллы, ограниченные мембранами, такие как аппарат Гольджи и эндоплазматический ретикулум, во время фазы М разделяются на более мелкие фрагменты и пузырьки, что обеспечивает их равномерное распределение между дочерними клетками. Однако при цитокинезе может происходить и запрограммированное асимметричное распределение материала. Например, клетка может делиться с образованием неравных по величине дочерних клеток или же какой-то компонент цитоплазмы может перед цитокинезом скапливаться на одной стороне клетки и передаваться только одной из двух в остальном одинаковых дочерних клеток. [c.467]
Можно предположить, что роль такого механизма играет клеточный цикл. Однако факты не нодтвержают это нредноложение дифференцировка ранних эмбриональных клеток следует установленной схеме и при искусственном ограничении клеточных делений под влиянием химических веществ, ингибирующих цитокинез или синтез ДНК. Клеточные деления не следует уподоблять периоду колебаний маятника биохимических часов, определяющих темп развития скорее ситуация обратная и именно биохимические часы контролируют темп клеточных делений и продолжительность клеточного цикла у множества видов животных. Изменение химического состояния клетки одновременно влияет на принятие решений о делении клеток, а также на время и нанравление дифференцировки. Молекулярные механизмы контроля клеточных делений в эмбриогенезе практически не изучены и представляют собой одну из центральных проблем современной биологии развития. Генеалогические мутанты нематод могут сыграть ключевую роль в решении этой проблемы. [c.91]
Стволовые клетки нужны в любом месте, где постоянно возникает потребность в новых дифференгр1рованных клетках, которые, однако, сами делиться не могут. В ряде тканей конечное состояние дифференгр1ровки явно несовместимо с клеточным делением. Например, ядра клеток могут разрушаться, как это происходт в наружных слоях эпидермиса, или выталкиваться из клеток, как при созревании эритроцитов > млекопитающих. Иногда осуществлению митоза и цитокинеза препятствует то, что цитоплазма плотно заполнена таким материалом, как, например, миофибриллы мышечных клеток. В других терминально дифференцированных клетках невозможность деления может быть обусловлена какими-то более тонкими биохимическими причинами. В любом таком случае обновление будет зависеть от стволовых клеток. [c.169]
Сам процесс клеточного деления очень легко наблюдать в микроскоп он включает две последовательные стадии-деление ядра (митоз) и деление цитоплазмы (цитокинез). Однако, прежде чем клетка сможет разделиться, она должна удвоить свою массу и редуплицировать все свои компоненты. Только в этом случае обе дочерние клетки будут иметь все необходимое для того, чтобы начать свой собственный цикл развития, который закончится делением. Большая часть работы по подготовке к делению совершается незаметно для глаза во время фазы роста, получившей весьма неудачное название интерфазы. [c.139]
Первые пять стадий деления составляют митоз, а шестой является цитокинез. In vivo эти шесть стадий образуют непрерывную динамическую последовательность, сложность и красоту которой трудно оценить по описанию или по серии статических изображений. Описание клеточного деления базируется на данных световой микроскопии живых клеток (часто в сочетании с микрокиносъемкой) и на результатах световой и электронной микроскопии фиксированных и окрашенных клеток. На рис. 11-40 и 11-41 представлены схемы различных фаз клеточного деления, а на рис. 11-42 и И-43-микрофотографии картин деления типичных животных и растительных клеток. [c.179]
События, происходящие в период от одного клеточного деления до другого, называются клеточным ииклом (рис. L3). Фаза митоза (М-фаза) цикла охватывает период деления и хромосом, и клеток. После расхождения клеток (цитокшеза) каждая дочерняя клетка вступает в период повы-щенной биосинтетической активности—в так называемую Gj-фазу (от англ. g ). Gj-фаза заканчивается перед началом удвоения хромосом, или, в молекулярных терминах, с началом дупликации хромосомной ДНК период репликации генома называется фазой шнтеза (5-фаз(й). С момента заверщения S-фазы в клетках инициируются события, хфактерные для митотической профазы,—части цикла, называемой G -фазой. В конце концов опять начинаются митоз и цитокинез, и цикл повторяется. Как правило, Gj-, S- и О -периоды, вместе составляющие интерфазу, занимают около 90% времени клеточного цикла, а М-фаза—менее 10%. Полное время прохождения клеточного цикла в клетках разного типа сильно варьирует (от минут до суток) в зависимости от условий роста. Основным показателем продолжительности всего цикла является продолжительность Gj-фазы. Например, покоя- [c.16]
Клеточный цикл мтоз и цитокинез (клеточное деление) составляют М-фазу цикла, кульминацией которой является образование двух дочерних клеток. Каждая дочерняя клетка вступает в С,-период интерфазы и может начать новый клеточный цикл. За периодом О, следует 8-фаза, во время которой ДНК и хромосомы дуплицируются, и далее-фаза О ,. Начало митоза означает конец интерфазы. Покоящиеся клетки задерживаются в фазе С, и, как говорят, находятся в фазе Обычно эукариотические клетки, которые не остановились в фазе Од, завершают цикл за 24 ч. [c.16]
Образование новой оболочки при делении клетки — цитокинезе — начинается с появления в экваториальной плоскости фраг-мопласта клеточной пластинки, которая, разрастаясь, разделяет материнскую клетку на две дочерние. На месте будущей пластинки образуются иузырьки, отделенные от структур Гольджи и трубочек веретена, сливающиеся по краям фрагмопласта. В результате внедрения в оболочку содержимого пузырьков сначала формируется клеточная пластинка, а затем и аморфный матрикс первичной и, иозже, вторичной оболочек [20, с. 36 8, с. 10], [c.28]
Удвоение многих компонентов клетки не требует точного контроля. Если в клетке имеется много молекул или органелл определенного типа, то достаточно того, чтобы число их приблизительно удвоилось за один цикл и они затем примерно поровну распределились между двумя дочерними клетками. Однако существует по крайней мере одно очевидное исключение в случае ДНК такое удвоение и распределение должно быть совершенно точным, а для этого нужен специальный механизм. Поэтому при рассмотрении клеточного цикла иногда удобно бывает различать хромосомный цикл и параллельный ему цитоплазматический цикл. В хромосомном цикле репликация ядерной ДНК (синтез ДНК) чередуется с митозом, в котором разделяются реплицированные копии генома В цитонлазматическом никле рост клетки, при котором удваиваются в числе другие клеточные компоненты, чередуется с цитокинезом-делением всей клетки на две. [c.394]
Если повысить температуру до рестриктивного уровня, у большинства мутантов сс1с клеточный цикл останавливается на той стадии, на которой действует продукт гена сб/с. Как правило, клетка теряет способность переходить к следующей стадии цикла, и это означает, что начало каждого процесса находится в зависимости от завершения предыдущего процесса Таким образом, у дрожжей, как и у млекопитающих, большинство этапов клеточного цикла, но-видимому, связаны между собой как звенья единой цепи. Эта связь была более тщательно проанализирована в экспериментах с клетками, содержащими разные комбинации различных мутаций сс1с. Как показали результаты, события хромосомного цикла образуют ряд зависимых друх от друга этапов, который не связан жестко с событиями цитоплазматического цикла (рис. 13-19). Например, хотя цитокинеза не произойдет, если предотвратить деление ядра, тем не менее мутанты сс1с, не способные пройти цитокинез из-за дефектов в механизме формирования почки, все же осуществляют повторные циклы синтеза ДНК и деления ядра. Но-видимому, общим правилом не только для дрожжей, но и для клеток млекопитающих, насекомых и многих других организмов является то, что хромосомный цикл может продолжаться, даже если цитокинез предотвращен. В самом [c.410]
chem21.info
Прошу! Помогите! На этот сайт последняя надежда! Сравнение процессов развития половых клеток у растений и животных?
<a rel="nofollow" href="http://umal.me/0sj" target="_blank">Влад Вот тут есть страница 252</a>
Длинное, но поможет. Продолжительность интерфазы в клетках растений и животных составляет в среднем 10- 20 часов, тогда как митоз занимает около 1-2 часов. В ходе последовательных делений в многоклеточных организмах дочерние клетки становятся все более разнообразными, поскольку в них происходит считывание информации со все большего числа генов. Некоторые клетки со временем перестают делиться и погибают, что может быть связано с завершением выполнения определенных функций, как в случае клеток эпидермиса кожи и клеток крови или с повреждением этих клеток факторами окружающей среды, в частности возбудителями болезней. Генетически запрограммированная смерть клетки называется апоптозом, тогда как случайная гибель — некрозом. Митоз — деление соматических клеток. Фазы митоза Митоз — способ непрямого деления соматических клеток. Во время митоза клетка проходит ряд последовательных фаз, в результате которых каждая дочерняя клетка получает такой же набор хромосом, как и в материнской клетке (табл. 2.5). Митоз делится на четыре основные фазы: профазу, метафазу, анафазу и телофазу (рис. 2.52). Профаза — наиболее длительная стадия митоза, в процессе которой происходит конденсация хроматина, в результате чего становятся видны Х-образные хромосомы, состоящие из двух хроматид (дочерних хромосом). При этом исчезает ядрышко, центриоли расходятся к полюсам клетки, и начинает формироваться ахроматиновое веретено (веретено деления) из микротрубочек. В конце профазы ядерная оболочка распадается на отдельные пузырьки. В метафазе хромосомы выстраиваются по экватору клетки своими центромерами, к которым прикрепляются микротрубочки полностью сформированного веретена деления. На этой стадии деления хромосомы наиболее уплотнены и имеют характерную форму, что позволяет изучить кариотип. В анафазе происходит быстрая репликация ДНК в центромерах, вследствие которой хромосомы расщепляются и хроматиды расходятся к полюсам клетки, растягиваемые микротрубочками. Распределение хроматид должно быть абсолютно равным, поскольку именно этот процесс обеспечивает поддержание постоянства числа хромосом в клетках организма. На стадии телофазы дочерние хромосомы собираются на полюсах, деспирализуются, вокруг них из пузырьков формируются ядерные оболочки, а во вновь образовавшихся ядрах возникают ядрышки. После деления ядра происходит деление цитоплазмы — цитокинез, в ходе которого и происходит более или менее равномерное распределение всех органоидов материнской клетки. Таким образом, в результате митоза из одной материнской клетки образуется две дочерних, каждая из которых является генетической копией материнской (2п2с). В больных, поврежденных, стареющих клетках и специализированных тканях организма может происходить несколько иной процесс деления — амитоз. Амитозом называют прямое деление эукариотических клеток, при котором не происходит образования генетически равноценных клеток, так как клеточные компоненты распределяются неравномерно. Он встречается у растений в эндосперме, а у животных — в печени, хрящах и роговице глаза. - Мейоз. Фазы мейоза Мейоз — это способ непрямого деления первичных половых клеток (2п2с), в результате которого образуются гаплоидные клетки (lnlc), чаще всего половые. В отличие от митоза, мейоз состоит из двух последовательных делений клетки, каждому из которых предшествует интерфаза (рис. 2.53). Первое деление мейоза (мейоз I) называется редукционным, так как при этом количество хромосом уменьшается вдвое, а второе деление (мейоз II) — эквационным, так как в его процессе количество хромосом сохраняется (см. табл. 2.5). Интерфаза I протекает подобно интерфазе митоза. Мейоз I делится на четыре фазы: профазу I, метафазу I, анафазу I и телофазу I. В профазе I происходят два важнейших процесса — конъюгация и кроссинговер. Конъюгация
Половые клетки животных образуются в половых органах - семенниках и яичниках. Половые клетки (мужские) имеют жгутики, у растений таких нет. У растенйий они образуются в пестиках и тычинках. У животных половые клетки с гаплоидным набором хромосом, при оплодотворении - зигота диплоидна, а у растений половые клетки 3 n набор, т. к. образуется зигота - диплоидная и плюс ещё эндосперм.
touch.otvet.mail.ru
Биология для студентов - 20. Судьба органелл при митозе
Пластиды — самовоспроизводящиеся органеллы клетки. В отличие от хромосом ядра при распределении между дочерними клетками не подчиняются строгим законам митоза и мейоза. Аппарат, управляющий распределением пластид, в настоящее время неизвестен и считается, что они попадают в дочерние клетки случайно при делении цитоплазмы, благодаря тому, что содержатся в клетке во множестве экземпляров (до нескольких сотен).
Во время деления клетки происходит репликация различных органелл, в том числе пластид и митохондрий. Простейшим типом пластиды является пропластида, из которой развиваются все типы пластид, включая хлоропласты. Пластиды представляют собой полуавторюмные органеллы, способные к удвоению путем деления или почкования. В клетках высших растений может содержаться от нескольких до большого числа пластид, и клетки различных типов значительно отличаются друг от друга по содержанию в них пластид. Число пластид в клетках какого-либо одного типа обычно остается приблизительно постоянным, и это наводит на мысль, что репликация пластид происходит одновременно с делением клетки. Однако распределение пластид материнской клетки между дочерними происходит, по-видимому, случайным образом.
Как же при делении клеток высших эукариот разделяются различные органеллы, окруженные мембраной (за исключением ядра) В большинстве случаев число этих органелл достаточно велико, чтобы и при случайном распределении их в процессе цитокинеза каждая дочерняя клетка получала их более или менее представительный набор. Таким образом, хотя клетка млекопитающего не выживет, не получив, например, ни одной митохондрии, вполне возможно, что для надежной передачи их дочерним клеткам не требуется никакого специального механизма. Органеллы, присутствующие в клетках в большом количестве, будут всегда успешно наследоваться, если в среднем их число будет удваиваться в каждом клеточном поколении.
Другие органеллы, такие как аппарат Гольджи и эндоплазматический ретикулум во время митоза распадаются на более мелкие фрагменты и пузырьки. Такое раздробление, вероятно, способствует их равному распределению между дочерними клетками.
Если в клетке имеется много органелл определенного типа, то достаточно того, чтобы число их приблизительно удвоилось за один цикл, и они затем примерно поровну распределились между двумя дочерними клетками. Однако существует по крайней мере одно очевидное исключение. В случае ДНК такое удвоение и распределение должно быть совершенно точным, а для этого нужен специальный механизм. Поэтому при рассмотрении клеточного цикла иногда удобно бывает различать хромосомный цикл и параллельный ему цитоплазматический цикл. В хромосомном цикле репликация ядерной ДНК (синтез ДНК) чередуется с митозом, в котором разделяются реплицированные копии генома. В цитоплазматическом цикле рост клетки, при котором удваиваются в числе другие клеточные компоненты, чередуется с цитокинезом-делением всей клетки на две.
Клеточное деление заканчивается разделением цитоплазмы (цитокинезом) хромосомы деконденсируются и на них возобновляется синтез РНК. По-видимому, цитокинез у столь различных эукариотических организмов, как животные, растения и грибы, направляется организованными пучками актиновых филаментов. Крупные органеллы, ограниченные мембранами, такие как аппарат Гольджи и эндоплазматический ретикулум, во время фазы митоза разделяются на более мелкие фрагменты и пузырьки, что обеспечивает их равномерное распределение между дочерними клетками. Однако при цитокинезе может происходить и запрограммированное асимметричное распределение материала. Например, клетка может делиться с образованием неравных по величине дочерних клеток или же какой-то компонент цитоплазмы может перед цитокинезом скапливаться на одной стороне клетки и передаваться только одной из двух в остальном одинаковых дочерних клеток.
Так какова же действительная роль центриолей. Согласно одной из крайних точек зрения, центриоли-это органеллы, единственная функция которых состоит в образовании ресничек или жгутиков. Их физическая ассоциация с клеточным центром и с полюсами веретена нужна лишь для равномерного распределения материала, способного к образованию базальных телец, между дочерними клетками. Однако митотическое веретено с центриолями на обоих полюсах и множеством расходящихся от них микротрубочек представляет собой четко локализованную структуру, тогда как в клетках, лишенных центриолей, например, у растений, нити веретена в гораздо меньшей степени сфокусированы у полюсов. Кроме того, ряд косвенных данных указывает на то, что в животных клетках центриоли играют роль главных организующих элементов, от которых зависит точное положение перицентриолярного материала, а тем самым и структурная полярность всей клетки.
vseobiology.ru