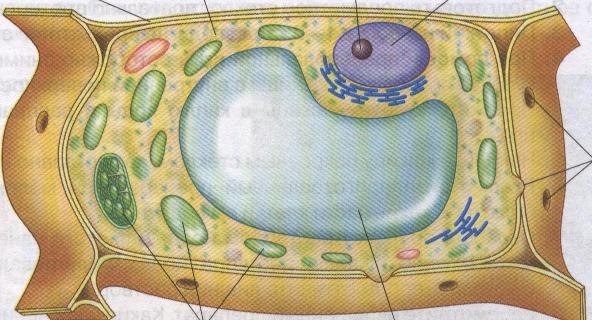
Корневая система как орган поглощения солей. Поступление воды в растение
Поступление воды из почвы в корень растения
Для нормального функционирования клетки растения должны быть насыщены водой. Состояние насыщения поддерживается с помощью двух взаимосвязанных процессов: поступления и выделения (расходования) воды, которые и составляют водный обмен растений. Соотношение между этими процессами называют водным балансом.
Растение выделяет воду в жидком и парообразном состояниях. Физиологический процесс испарения воды наземными органами растений получил, как уже отмечалось, название транспирации, выделение воды в капельножидком состоянии – гуттации.
Жизнь возникла в мировом океане. С выходом растений на сушу появилась опасность их гибели от высыхания. Почему? Концентрация водяных паров в воздухе, даже в условиях влажного климата, всегда меньше, чем в межклетниках листа, поэтому неизбежно происходит движение воды из тела растения в окружающую среду, при этом диффузия молекул воды от поверхности листа в воздух происходит в 1 500 раз быстрее, чем их поступление из почвы в корень.
Большая потеря воды растением обусловлена ее большой листовой поверхностью. Одно растение кукурузы расходует за вегетационный период 200 литров воды, а сахарный тростник – в два раза больше. Растения вынуждены формировать большую листовую поверхность, чтобы получать необходимое количество СО2 для фотосинтеза, несмотря на его малое содержание (0,045 %) в атмосфере.
Например, 8-летняя яблоня может ассимилировать за день до 50 г СО2 при интенсивном фотосинтезе. Это количество СО2 она поглощает из 300 000 литров воздуха.
Необходимость расходовать воду в больших количествах, связано еще с тем, что поглощение солнечных лучей растениями должно привести к повышению температуры, которая может вызвать коагуляцию белков. Испаряя воду, растение понижает температуру своего тела.
Для большинства растений подсыхание смертельно, поэтому расход воды должен соотноситься с приходом. Только мхи и лишайники могут выдерживать длительное отсутствие воды и переносить это время в состоянии высыхания.
Поэтому, как только растения вышли из воды на сушу, начался долгий процесс по выработке приспособлений для поддержания водного баланса: во-первых, для быстрого поступления, во-вторых, для уменьшения траты наземными органами; в-третьих, для экономически выгодного транспорта по стеблю.
При анализе соотношения между поступлением и расходованием воды возможны три случая: поступление больше расходования, равно или меньше. В последнем случае возникает водный дефицит. В полдень водный дефицит может достигать 5–10 и даже 25 %. Одно из условий нормального функционирования наземных растений – поддержание условий без длительного и глубокого водного дефицита. Для этого необходима хорошо развитая корневая система, обеспечивающая поступление воды с большой скоростью.
Поступление воды из почвы в корень
Поступление воды из почвы представляет собой более сложный процесс, чем простое всасывание воды растением, опущенным в какой-либо водный раствор корнями. В почве мы имеем ряд сил, противодействующих этому всасыванию, которые можно назвать водоудерживающими силами.
Почва – многофазная система, состоящая из четырех основных компонентов: твердых минеральных частиц, органического вещества (гумуса), почвенного раствора и почвенного воздуха.
Минеральные частицы и гумус образуют почвенную структуру, вода и воздух заполняют полости этой структуры.
Способность почвы удерживать воду зависит от ее состава и свойств. Определенное количество воды входит в состав минеральных компонентов почвы и недоступна растениям (гигроскопическая вода). Разнообразные глинистые минералы и гетерогенные гумусовые вещества, которые представляют собой коллоиды, могут также удерживать значительное количество гидратационной воды. Такая вода условно называется связанной (пленочная) и тоже труднодоступна. Вода, находящаяся в капиллярах почвы (капиллярная вода), легко всасывается корневыми волосками и поверхностными клетками сосущей зоны корня. Такая вода считается свободной.
Кроме того, в почве мы имеем дело с раствором, а не с чистой водой; поэтому сам раствор имеет сосущую силу. При равновесии концентраций в почве и в сосудах ксилемы в общем случае поступление воды должно остановиться.
Чтобы вычислить количество недоступной почвенной влаги поступают следующим образом. Растения выращивают в сосуде с непроницаемыми для воды стенками (стеклянный или металлический) и после того, как растения хорошо разовьются, почву перестают поливать и оставляют в притемненном месте до завядания растения. Завядание указывает, что доставка воды в корни прекратилась. То количество воды, которое остается в почве к моменту завядания, и будет недоступной для растения (мертвый запас). Количество недоступной воды получило название коэффициента завядания или влажности завядания. Следовательно, коэффициент завядания характеризует влажность, при которой устойчивое завядание только начинается.
В этот момент в почве имеется еще некоторое количество воды, хоть и малое, но в какой-то степени доступное для растения. В этих условиях даже сильно подвядшее растение оживает, если его начать поливать. Полная гибель наступает только при влажности значительно ниже, чем коэффициент завядания.
Таким образом, влажность завядания представляет собой нижнею границу того интервала влажности, при котором возможен рост растения.
При поступлении воды в сухую почву она впитывается очень быстро. Затем, скорость просачивания воды в нижние горизонты становится медленной. Когда скорость нисходящего тока воды резко понижается, влажность почвы достигает уровня, который называется полевой влагоемкостью.
Под доступной для растения почвенной влагой имеется ввиду количество воды, которое накапливается в почве от уровня влажности устойчивого завядания до полевой влагоемкости.
Говоря о перемещении воды по растению выделяют транспорт в тканях одного органа, который называют ближним (радиальный), и транспорт между отдельными органами, который называют дальним. Между ними есть существенная разница. Ближний транспорт идет по неспециализированным тканям, а для дальнего в растениях имеются специальные проводящие ткани.
biofile.ru
Поступление воды в растение. Двигатели водяного потока. - конспект лекций
Корневая система распространяется в почве в вертикальном и горизонтальном направлениях. Особенности распространения зависят от видовых особенностей растения. Так, у пустынных растений корневая система распространяются вглубь на десять (а отдельные виды и на большее количество) метров, а у теневыносливых растений, растущих в нижнем ярусе леса, корневая система в основном располагается в ярусе до 0.5 метра, но вширь может занимать несколько квадратных метров.Поступление воды в корневую систему растения и перемещение ее по тканям корня осуществляется путем пассивной диффузии. Поступление идет по градиенту концентрации, поэтому если в почве концентрация почвенного раствора выше, чем концентрация клеточного сока, то вода будет диффундировать не в растение, а из него, и наступит гибель растения. Такая ситуация может сложиться в результате передозировки минеральных удобрений, небрежного внесения минеральных удобрений, когда они рассыпаются неравномерно.Корневая система имеет поглощающую или всасывающую зону - это зона корневых волосков. Поступив в клетку корневого волоска вода становится частью живой системы - клетки растения - и подчиняется закономерностям, действующим в живой клетке. Передвижение по растению определяется двумя основными двигателями водного потока в растении:нижним двигателем водного потока или корневым давлением,верхним двигателем водного потока или присасывающим действием атмосферы.Корневое давление создается при переходе воды из коры корня в сосудистую систему корня при прохождении воды через пропускные клетки перицикла, из которых вода под давлением как бы впрыскивается в сосуды ксилемы. Доказательством этого служат явления гуттации и "плача растений".Гуттация - это выделение капельно-жидкой влаги листьями через гидатоды в условиях затрудненного испарения.Плач растения - это вытекание пасоки (воды с растворенными в ней минеральными веществами, находящейся в ксилеме) из стеблей растений со срезанными побегами. Механизм образования корневого давления по-видимому состоит из двух аспектов:переноса воды по законам осмоса,дополнительной сократительной деятельности актомиозиновых белков, находящихся в перицикле и паренхимных клетках корня.Присасывающее действие атмосферы определяется концентрацией водяных паров в атмосфере. Этот показатель в атмосфере почти всегда меньше, чем в листе растения, за исключением условий повышенной влажности воздуха, например, во время дождя, тумана.Определяющую роль в формировании верхнего двигателя водяного потока в растении играет водный потенциал Ґ (фэта).Водный потенциалYвыражает способность воды в данной системе, в том числе в почвенном растворе, или в клетке растения, или в атмосфере, совершить работу по сравнению с той работой, которую при тех же условиях совершила бы чистая вода.Водный потенциал, являясь фактически мерой активности воды, определяет термодинамически возможное направление ее транспорта. Молекулы воды всегда перемещаются от более высокого водного потенциала к более низкому, подобно тому, как вода течет вниз. Водный потенциал имеет размерность энергии, деленной на объем, поэтому его выражают в барах или паскалях (1 атмосфера = 1,013 бар = 105 Па.106Па равны 1 мегаПа)Химический потенциал воды - µw - это величина, производная от активности воды. Она выражает максимальное количество внутренней энергии молекул воды, которое может быть превращено в работу, измеряется в ДЖ. моль-1 и рассчитывается по уравнению:µw = µw 0 +RT ln aw, гдеµw 0 - химический потенциал чистой воды (принят равным нулю), R - газовая постоянная, T - абсолютная температура, aw - активность воды в системе.В системе "почвенный раствор - растение - атмосфера" водный потенциал изменяется от самого высокого значения в почвенном растворе до самого низкого в воздухе. Вода переходит из растения в окружающий воздух в парообразном состоянии. В мезофилле листа имеются обширные межклеточные пространства и каждая клетка мезофилла хотя бы одной стороной граничит с таким межклетником. Вследствие испарения воды с влажных клеточных стенок воздух в межклетниках насыщен водяными парами, часть которых через устьица выходит наружу.
2dip.su
Поступление воды в растение
Формы воды в растении.
Функции и формы воды в растении.
Передвижение воды у цветковых растении.
Поступление воды в растения.
Функции и формы воды в растении.
Водный режим растений
Л Е К Ц И Я 2
В тканях растений вода составляет 70-95% сырой массы. Все известные на Земле формы жизни не могут существовать без неё. При снижении содержания воды в клетках и тканях до критического уровня (например у спор, у семян при их полном созревании) живые структуры переходят в состояние анабиоза.
Вода в биологических объектах выполняет следующие основные функции:
1. Водная среда объединяет все части организма, начиная от молекул в клетках и кончая тканями и органами, в единое целое. В теле растения водная фаза представляет собой непрерывную среду на всём протяжении от влаги, извлекаемой корнями из почвы, до поверхности раздела жидкость-газ в листьях, где она испаряется.
2. Вода – важнейший растворитель и важнейшая среда для биохимических реакций.
3. Вода – метаболит и непосредственный компонент биохимических процессов. Так, при фотосинтезе вода является донором электронов. При дыхании, например в цикле Кребса, вода принимает участие в окислительных процессах. Вода необходима для гидролиза и для многих синтетических процессов.
4. Вода – главный компонент в транспортной системе высших растений – в сосудах ксилемы и в ситовидных трубках флоэмы.
5. Вода – терморегулирующий фактор. Она защищает ткани от резких колебаний температуры благодаря высокой теплоёмкости и большой удельной теплоте парообразования.
6. Вода – хороший амортизатор при механических воздействиях на организм.
7. Благодаря явлениям осмоса и тургора (напряжения) вода обеспечивает упругое состояние клеток и тканей растительных организмов.
8. Вода участвует в упорядочении структур в клетках. Она входит в состав белков, определяя их конформацию. Удаление воды из белков высаливанием или с помощью спирта приводит к их коагуляции и выпадению в осадок.
Имеющаяся в клетках вода может быть свободной и связанной.
Связанная вода удерживается силой притяжения гидрофильных коллоидов. В ходе приспособления растения к неблагоприятным условиям (засухе, понижениям температуры) количество связанной воды увеличивается.
Свободная вода служит средой, где идут процессы жизнедеятельности клетки, в которых она может участвовать. Она входит в состав клеточного сока, органических веществ, тратится на испарение. Потеря свободной воды может нарушить метаболизм в клетке.
Вода поступает в растение в основном через корневые волоски по законам осмоса. Различают диффузию и осмос .
Диффузия – это движение молекул или ионов из области с высокой концентрацией в область с более низкой концентрацией, иными словами как движение по градиенту концентрации. Диффузию жидкостей или газов можно также определить как смешивание их при непосредственном соприкосновении (например, распространение запаха духов или смешивание воды с другой жидкостью).
Если между двумя жидкостями поместить животную или растительную мембрану (перепонку), то растворы также будут проникать через неё, так как она мелкопориста. Такое явление получило название «осмоса».
Осмос – это переход молекул растворителя из области с более высокой концентрацией в область с более низкой концентрацией через полупроницаемую мембрану.
Во всех биологических системах растворителем служит вода.
Таким образом, осмос можно рассматривать как особый вид диффузии, при котором равновесие достигается за счёт перемещения одних только молекул растворителя.
Мембраны, пропускающие только молекулы растворителя и задерживающие все молекулы или ионы растворённого вещества, называют полупроницаемыми.
Избирательно проницаемыми являются мембраны живых клеток, которые пропускают определённые молекулы или ионы растворённых веществ.
Так, если раствор сахарозы в ячейке и избирательно проницаемого материала поместить в чистую воду, то вода устремится внутрь ячейки, а частицы сахара будут поступать из ячейки в воду. Проникновение частиц воды внутрь ячейки называется эндоосмосом, а поступление оттуда частиц сахара – экзоосмосом. Преобладание эндосмоса над экзосмосом вызывает в системе осмотическое давление.
Таким образом, если какой-либо раствор отделён избирательно проницаемой мембраной от чистой воды, то гидростатическое давление, которое необходимо приложить, чтобы предотвратить осмотическое поступление воды в раствор, называется осмотическим давлением этого раствора.
Величина осмотического давления зависит от разности концентраций растворов, а также от величины молекул, участвующих в осмосе. Частицы воды проникают через перепонки (растительные, животные) быстрее, чем частицы кристаллоидов, а те, в свою очередь, двигаются быстрее, чем более крупные коллоидные частицы. Последние иногда совсем не проникают в клетку.
Таким образом, растительные клетки обладают осмотическим давлением. Оно заключается в давлении вещества клеточного сока на цитоплазму и оболочку клетки. Этому давлению противопоставляется давление растянувшейся до предела клеточной оболочки на цитоплазму – тургорное давление.
studlib.info
Движение воды по растению
Основные двигатели водного тока
Поглощение воды корневой системой идет благодаря работе двух концевых двигателей водного тока: верхнегоконцевого двигателя, или присасывающей силы испарения (транспирации), и нижнего концевого двигателя, или корневого двигателя. Основной силой, вызывающей поступление и передвижение воды в растении, является присасывающая сила транспирации, в результате которого возникает градиент водного потенциала. Водный потенциал – это мера энергии, используемой водой для передвижения. Водный потенциал и сосущая сила одинаковы по абсолютному значению, но противоположны по знаку. Чем меньше насыщенность водой данной системы, тем меньше (более отрицателен) ее водный потенциал. При потере воды растением в процессе транспирации создается ненасыщенность клеток листа водой, как следствие, возникает сосущая сила (водный потенциал падает). поступление воды идет в сторону большей сосущей силы, или меньшего водного потенциала.
Таким образом, верхний концевой двигатель водного тока в растении – это присасывающая сила транспирации листьев, и его работа мало связана с жизнедеятельностью корневой системы. Действительно, опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется.
Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация.
Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, препятствующей испарению, выделяют капельно-жидкую воду с небольшим количеством растворенных веществ – гуттация. Выделение жидкости идет через специальные водные устьица – гидаторы. Выделяющаяся жидкость – гутта. Таким образом, процесс гуттации является результатом одностороннего тока воды, происходящего в отсутствие транспирации, и, следовательно, вызывается какой-то иной причиной.
К такому же выводу можно прийти и при рассмотрении явления плач растений. Если срезать побеги растения и к срезанному концу присоединить стеклянную трубку, то по ней будет подниматься жидкость. Анализ показывает, что это вода с растворенными веществами – пасока. В некоторых случаях, особенно в весенний период, плач наблюдается и при надрезе веток растений. Определения показали, что объем выделяющейся жидкости (пасоки) во много раз превышает объем корневой системы. Таким образом, плач – эт6о не просто вытекание жидкости в результате пореза. Все сказанное приводит к выводу, что плач, как и гуттация, связана с наличием одностороннего тока воды через корневые системы, не зависящего от транспирации. Силу, вызывающую односторонний ток воды по сосудам с растворенными веществами, не зависящую от процесса транспирации, называют корневым давлением. Наличие корневого давления позволяет говорить о нижнем концевом двигателе водного тока. Корневое давление можно измерить, присоединив манометр к концу, оставшемуся после срезания надземных органов растения, или поместив корневую систему в серию растворов различной концентрации и подобрав такую, при которой плач прекращается. Оказалось, что корневое давление равняется примерно 0,1 – 0,15 МПа (Д.А.Сабинин). Определения, проведенные советскими исследователями Л.В.Можаевой, В.Н.Жолкевичем, показали, что концентрация наружного раствора, останавливающего плач, значительно выше концентрации пасоки. Это позволило высказать мнение, что плач может идти против градиента концентрации. Было показано также, что плач осуществляется только в тех условиях, в которых нормально протекают все процессы жизнедеятельности клеток. Не только умерщвление клеток корня, но и снижение интенсивности их жизнедеятельности, в первую очередь интенсивность дыхания, прекращает плач. В отсутствии кислорода, под влиянием дыхательных ядов, при понижении температуры плач приостанавливается. Все сказанное позволило Д.А.Сабинину дать следующее определение: плач растений – это прижизненный односторонний ток воды и питательных веществ, зависящий от аэробной переработки ассимелятов. Д.А.Сабинин предложил схему, объясняющую механизм одностороннего тока воды в корне. Согласно этой гипотезе, клетки корня поляризованы в определенном направлении. Это проявляется в том, что в разных отсеках одной и той же клетки процессы обмена веществ различны. В одной части клетки идут усиленные процессы распада, в частности, крахмала на сахара, вследствие чего концентрация клеточного сока возрастает. На противоположном конце клетки преобладают процессы синтеза, благодаря чему концентрация растворенных веществ в этой части клетки уменьшается. Надо учитывать, что все эти механизмы будут работать только при достаточном количестве воды в среде и не нарушенном обмене веществ.
Согласно другой гипотезе, зависимость плача растений от интенсивности дыхания является косвенной. Энергия дыхания используется для поступления ионов в клетки коры, откуда они десорбируются в сосуды ксилемы. В результате концентрация солей в сосудах ксилемы повышается, что и вызывает поступление воды.
Передвижение воды по растению
Вода, поглощенная клетками корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации, а также силе корневого давления, передвигается до проводящих путей ксилемы. Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. Еще в 1932 г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно не зависимых друг от друга объемов, по которым передвигается вода, - апопласта и симпласта. Апопласт – это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт – это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт, по-видимому, не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая – по другую сторону клеток эндодермы, и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Для того чтоб попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану и главным образом по апопласту и лишь частично по симпласту. Однако в клетках эндодермы передвижение воды идет, по-видимому, по симпласту. Далее вода поступает в сосуды ксилемы. Затем передвижение воды идет по сосудистой системе корня, стебля и листа.
Из сосудов стебля вода движется через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой, имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. Вода передвигается от клетки к клетке благодаря градиенту сосущей силы.
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту значительно большую 10 м. сила сцепления увеличивается, так как молекулы воды обладают большим сродством друг к другу. Силы сцепления обладают и между водой и стенками сосудов.
Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды.
В том случае, если в отдельные членики сосудов попадает воздух, они, по-видимому, выключается из общего тока проведения воды. Таков путь передвижения воды по растению (рис. 1).

Рис. 1. Путь воды в растении.
Скорость перемещения воды по растению в течение суток изменяется. В дневные часы она на много больше. При этом разные виды растений различаются по скорости передвижения воды. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды. Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды.
Видео: Движение воды и органических веществ по стеблю. biofile.ru
Поступление воды в растение
1
поддержание структуры цитоплазмы путем гидратации ееколоидов и тургорного состояния;среда для протекания обмена веществ;среда для переноса веществ в пределах растительногоорганизма;регулятор температуры тела растения;вода обеспечивает непрерывность передвижения питательныхвеществ в растении.
Для нормальной физиологической деятельности клетка растениядолжна быть полностью насыщена водой (или близка к2насыщению).
Св ободнаяСв язанная
Свободнаяводавсоставляет 85-90 % всвязанная 10-15%.3
растениирастении,
Свободная вода легко передвигается по растению и испаряется.Она находится в основном в межклетниках т.е. в свободномпространстве.Связанная вода испаряется и передвигается с трудом. Онанаходится преимущественно внутри клетки – в цитоплазме ивакуоли. Эта форма воды в свою очередь разделяется наосмотически-связанную и коллоидно-связанную. Первая соединенас растворенными в ней веществами и находится в вакуоли. Этасвязь не очень прочная, и осмотически-связанная вода можетвыходить из клетки, например при плазмолизе. Свойстваколлоидно-связанной воды обусловлены наличием главнымобразом белков – коллоидов цитоплазмы, где она и находится.Такая связь большей частью очень прочная, и коллоидносвязанная вода выходит из клетки только при сильном4обезвоживании(продолжительной засухе).
Почвенная вода имеет большое значение, является одним изфакторов плодородия и урожайности растений. От содержания икачества воды в почве зависят произрастание растений идеятельность микроорганизмов.Основной источник влаги — атмосферные осадки, которыепроникают в почву и заполняют ее поры. В почве влага активновзаимодействует с твердой фазой (частью) почвы. Передвижениевлаги, ее доступность растениям зависят от состава и свойствпочвы.В естественных условиях почва обладает различной степеньювлажности. Понятие «влажность» характеризует содержание воды впочве, выраженное в процентах от массы сухой почвы (весоваявлажность) или от объема почвы (объемная влажность).
5
гравитационнаякапилярная
грунтовая
твердая
Формыпочвеннойвлаги
парообразная
кристализационная6
сорбированная
химическисвязанная
Непосредственно для питания растений имеет значение толькогравитационная и капиллярная вода, а остальные формы почвеннойвлаги, кроме небольшой части пленочной, растениям недоступны.Гравитационная вода заполняет капиллярные поры междуструктурными — отдельностями, по которым она передвигается подвлиянием силы тяжести (отсюда и ее название).Капиллярная вода заполняет капиллярные поры, главным образом,внутри структурных отдельностей. Она может передвигаться в почвево всех направлениях.
7
Сорбированная вода удерживается на поверхности почвенныхчастицсорбционнымисилами,тоестьмолекулыводыпритягиваются к твердым частицам почвы и прочно удерживаютсяими. Эту форму воды подразделяют на два вида: пленочную игигроскопическую.Пленочная вода окружает твердые частицы почвы в виде пленки,притягиваясь к ним под действием поверхностной энергии. Онапередвигается только под влиянием молекулярных сил в разныхнаправлениях, но всегда от более толстых пленок к тонким.Пленочная вода определяет смачивание почвы, но растениям почтинедоступна, так как притягивается к поверхности частиц твердойфазы почвы с силой в несколько тысяч атмосфер (от 6 до 10 тыс.).8
Гигроскопическая влага представляет собой молекулы водяногопара, удерживаемые поверхностным притяжением почвенных частицподобно тому, как удерживается пленочная вода. Поэтомугигроскопическая влага не принимает участия в газовом давленииокружающей среды и не способна передвигаться. Для растений онанедоступна, полностью удаляется при высушивании почвы втечение нескольких часов при температуре 100—105 °С.Свободная парообразная влага входит в состав почвенноговоздуха в виде отдельных молекул водяного пара и поэтомупринимает участие в газовом давлении и передвигается из мест сбольшей упругостью пара в места с меньшей упругостью. Онанедоступна для растений, но при переходе в капельно-жидкую можетусваиваться ими.9
Грунтовая вода — это влага водоносного слоя почвы, лежащегонижепочвеннойтолщи,удерживаемаяслоемводоупора.Использование грунтовой воды растениями возможно, но приблизком залегании и поднятии до корнеобитаемого слоя.Твердая вода (лед) — переход влаги из жидкого состояния втвердое происходит у свободных форм влаги при температуре ниже0 °С.Химически связанная и кристаллизационная вода входит в составмолекул минералов в виде ионов. Кристаллизационная воданаходится в составе кристаллических веществ в виде молекул.Растениям эти формы воды недоступны.10
Следовательно в почве всегда есть часть воды, которую растения немогут использовать. Эта вода носит название мертвого запаса, авлажность почвы, содержащей неусвояемую воду, называетсякоэффициентом завядания. Величина его колеблется от 0,9 до 16,2 %на сухую массу почвы и зависит в основном от ее свойств: легкие,бедные гумусом почвы имеют низкий коэфициент завядания, атяжелые, богатые гумусом – высокий.Для нормальной жизни растения в почве должно быть определенноесоотношение между водой и воздухом, поэтому она должна бытьполностью насыщена влагой. Оптимальной считается влажность 6080% от полной влагоемкости. Влажность почвы 30-60% уженедостаточна, а ниже 30 % означает сильную засуху и вредна длярастений.11
У сосудистых растений передвижение веществ осуществляется подвум системам: ксилеме (вода и минеральные соли) и флоэме(органические вещества). Передвижение веществ по ксилеменаправлено от корней к надземным частям растения; по флоэмепитательные вещества движутся от листьев.Растение может поглощать воду всеми своими органами, но большаячасть ее усваивается корнями. Поэтому корневую систему (а именнорастущую часть корня с корневыми волосками) считают органомпоглощения воды, которое может осуществляться двумя способами.
12
13
Всасывание — пассивный осмотический процесс, которыйосуществляется за счет сосущих сил клеток корня. Для этогонеобходимо, чтобы поглощенная вода тотчас передвигалась порастению. Поэтому всасывание происходит при недостатке воды врастении. Хотя процесс и считается пассивным, растение может внекоторых пределах регулировать (увеличивать) всасывание. Этодостигается повышением осмотического потенциала и сосущих силклеток корня за счет активного закачивания ионов натрия.Нагнетание воды — активный процесс поглощения, которыйпроисходит главным образом при достаточном или высокомсодержании воды в растении. Основой нагнетания служит корневоедавление, т. е. активное выдавливание воды корнем в направленииот периферии к центру. Наличие корневого давления подтверждаютявления гуттация и плач растений.14
Плач растений — вытекание жидкости изперерезанного у корневой шейки стебля.Если на пенек растения надеть трубку, товытекающая из среза жидкость — пасока— поднимется по ней на определеннуювысоту, которая обусловлена величинойкорневого давления. У разных видоврастений она неодинакова, но в среднемравна 0,15 МПа (1,5 атм).
15
Гуттация — выделение капельножидкой воды, которое наблюдается налистьях растений утром во влажнуюпогоду при хорошем водоснабжении ислабом испарении. В виде капельвыделяетсявода,котораяактивнонагнетается растением, но не успеваетиспаряться.
Спонижениемтемпературыскорость поступления воды резкосокращается. Это может оказатьзаметноевлияниенажизньрастительного организма, особеннов осенний период, когда испарениеидет еще достаточно интенсивно, апоступление воды задерживаетсяиз-запониженнойтемпературыпочвы.Врезультатерастения завядают и даже могутпогибнуть от обезвоживания.16
Причин, по которым понижение температуры вызывает уменьшениепоступления воды несколько:повышается вязкость воды и, как следствие, снижается ееподвижность;уменьшается проницаемость протоплазмы для воды;тормозится рост корней;уменьшается скорость всех метаболических процессов.Последнее обстоятельство, по - видимому, должно сказатьсякосвенно, через уменьшение поступления солей и, как следствие,торможение работы нижнего концевого двигателя (корневогодавления).17
Снижение аэрации почвы также тормозит поступление воды. Этосвязано с тем, что все условия, снижающие метаболизм, такие, какнедостаток кислорода, избыток СО2, дыхательные яды, снижаютпоступление ионов и, как следствие, уменьшают поступление воды.18
Большое значение имеет содержание водыв почве, а также концентрация почвенногораствора. Естественно, вода поступает вкорень только тогда, когда водныйпотенциал корня ниже, т. е. болееотрицателен, водного потенциала почвы.В том случае, если почвенный растворимеет более отрицательный осмотическийпотенциал, вода не только не будетпоступать в корень, но будет выделятьсяиз него. Всякое уменьшение влажностипочвы снижает поступление воды. Чемменьше воды в почве, тем с большей силойона удерживается и тем меньше ее водныйпотенциал. 19
freedocs.xyz
Корневая система как орган поглощения солей
На заре эволюции способность к поглощению воды и питательных веществ была присуща всем клеткам растительного организма. По-видимому, разделение функций между клетками отдельных органов осуществлялось постепенно. Это подтверждается тем, что клетки листа сохранили в определенной мере способность к поглощению не только воды, но и питательных солей. Сформировавшаяся корневая система — сложный специализированный орган. Основной зоной поглощения питательных веществ, снабжающей и наземные органы растения, является зона растяжения клеток и зона корневых волосков. Подсчеты показывают, что на 1 мм2 поверхности корня развивается от 200 до 400 корневых волосков. Таким образом, корневые волоски увеличивают поверхность корня в сотни раз. Они обладают и повышенной способностью к поглощению. Было установлено, что и клетки меристематической зоны корня обладают способностью к поглощению ионов, однако поглощенные ионы используются в этих же клетках. Это связано как с физиологическими особенностями меристематических клеток, так и с особенностями анатомического строения корня. В меристематической зоне еще нет дифференцированной сосудистой системы. При этом флоэма дифференцируется раньше и лишь несколько выше по длине корня образуется ксилема. Именно по ксилеме происходит передвижение воды с растворенными питательными веществами. Таким образом, в корне различают зону, участвующую в поглощении питательных веществ (меристематическая зона), и зону, участвующую как в поглощении, так и в снабжении питательными веществами надземных органов растений (зона корневых волосков). Поверхность корня, участвующую в адсорбции питательных веществ, называют общей адсорбирующей поверхностью. Поверхность корня, которая участвует не только в адсорбции, но и в передаче питательных веществ в сосуды ксилемы и дальше в надземные органы, называют рабочей адсорбирующей поверхностью.
Особенности поступления солей в корневую систему
Растительный организм обладает способностью к избирательному накоплению питательных элементов. Для выяснения механизма поступления минеральных веществ в растение большое значение имели исследования по влиянию на этот процесс различных условий.
Влияние внешних условий на поступление солей. При температуре, близкой к 0°С, поглощение солей идет очень медленно. Затем в пределах до 40°С поглощение усиливается. Увеличение температуры на 10°С может вызвать возрастание поглощения в два и даже три раза.
В темноте поглощение солей замедляется и постепенно прекращается, а под влиянием освещения ускоряется. Так, при освещении поглощение фосфора усиливается уже через 2—5 мин. Быстрота реакции указывает на прямое действие света. Вместе с тем свет может оказывать и косвенное влияние. На свету в процессе фотосинтеза образуются углеводы, которые необходимы для дыхания. Зависимость поглощения от интенсивности дыхания проявляется четко. При длительном выдерживании растений в темноте, после того как запас дыхательных субстратов исчерпан, поглощение солей не только прекращается, но может даже наблюдаться их выделение.
Интенсивность поступления солей остается на одном уровне при уменьшении содержания кислорода до 2—3%. Лишь снижение концентрации кислорода ниже 3% вызывает падение поглощения примерно в два раза. Необходимо учитывать, что и интенсивность дыхания сохраняется на высоком уровне в широком диапазоне концентраций кислорода. Падение интенсивности дыхания наблюдается при той же концентрации кислорода, при которой поглощение солей уменьшается.
Концентрация ионов водорода (рН) сказывается на поглощении различными путями. Было показано, что при подкислении раствора поступление катионов задерживается. Это происходит, по всей вероятности, в силу конкуренции, которая наблюдается между одинаково заряженными ионами, за возможность вступить в реакцию с переносчиками. Вместе с тем подкисление улучшает доступность ионов фосфорной кислоты. Наоборот, подщелачивание внешнего раствора снижает поступление фосфора благодаря переходу одновалентного иона Н2РО4- в двухвалентный НРО42- и трехвалентный Р043-, которые являются менее доступными для растения. Резкое изменение рН может также оказать влияние из-за повреждения мембран клетки.
Поглощение одного иона зависит от присутствия других ионов. Так, в присутствии легко поглощаемого аниона катионы той же соли поступают быстрее. Ионы с одинаковым зарядом обычно конкурируют между собой. Однако в некоторых случаях наблюдается противоположная закономерность. Так, при наличии в среде иона фосфора (РО43-) поглощение нитратов (NО3-) ускоряется. Это может явиться косвенным результатом увеличения использования нитратов в присутствии фосфорной кислоты.
Влияние внутренних факторов на поступление солей. Зависимость поступления солей от интенсивности дыхания является установленным фактом. При замене кислорода азотом не только прекращается поступление, но наблюдается выделение питательных ионов из корня. Ингибиторы процесса дыхания (в частности, цианистый калий) резко тормозят поступление солей. Процесс дыхания может оказывать влияние на поступление солей в нескольких направлениях. Так, в процессе дыхания выделяющаяся углекислота в водной среде диссоциирует на ионы Н+ и НСО3-. Адсорбируясь на поверхности корня, эти ионы служат обменным фондом для поступающих катионов и анионов. В процессе переноса ионов через мембрану участвуют специфические белки-акцепторы, синтез которых находится в зависимости от интенсивности дыхательного процесса. Наконец, энергия, выделяемая в процессе дыхания, непосредственно используется для поступления солей. В этой связи особенно важно, что вещества, нарушающие накопление энергии дыхания в макроэргических фосфорных связях (динитрофенол), также тормозят поступление солей.
Поступление воды и солей во многих случаях идет независимо друг от друга. Сопоставление количества испаренной воды в процессе транспирации и количества поступивших солей показывает, что прямой зависимости между этими процессами обычно нет. Так, при выращивании растений в условиях высокой влажности воздуха транспирация резко падает, а поступление солей идет с достаточной интенсивностью. Все же в некоторых случаях увеличение интенсивности транспирации может сказаться положительно на поглощении солей. Опыты показали, что в молодом возрасте (до фазы выхода в трубку) растения пшеницы поглощают фосфора в 14 раз больше, а в фазу колошения поглощение фосфора в 2 раза меньше, чем это следовало бы, исходя из того количества, которое должно было бы содержаться в испаренной воде. Известно также, что усиление транспирации приводит к ускорению передвижения восходящего тока воды с растворенными солями, что способствует быстрейшему освобождению от них клеток корня, а, следовательно, косвенно ускоряет поглощение. Надо иметь в виду и трудность разграничения влияния транспирации от влияния фотосинтеза. Открытие устьиц вызывает усиление как того, так и другого процесса. Увеличение интенсивности фотосинтеза приводит к возрастанию содержания углеводов и, как следствие, к увеличению интенсивности дыхания и поступления солей. Как правило, ускорение темпов роста увеличивает использование питательных веществ и тем самым усиливает их поступление. Наряду с этим быстрый рост корневой системы оказывает прямое влияние на поглощение благодаря увеличению поверхности, соприкасающейся с почвой.
Механизм и пути поступления минеральных солей через корневую систему.
Способность растения к избирательному концентрированию веществ, относительная независимость поступления воды и солей, зависимость поступления от дыхания и фотосинтеза, ускорение поступления под влиянием температуры и света показывают, что поступление питательных солей в корневую систему носит частично активный, связанный с метаболизмом характер. Для разграничения пассивного и активного поступления солей в корневую систему важное значение имели опыты, в которых было показано, что, если корневые системы растений перенести из слабого раствора в концентрированный, скорость поглощения на короткое время усиливается, а затем выравнивается. Эта первая фаза поглощения из более концентрированного раствора не зависит от температуры и не тормозится в присутствии ингибиторов. Если растение снова перенести в более слабый раствор, то некоторое количество ионов выделяется во внешнюю среду, так как часть тканей корня открыта для свободной диффузии ионов. Проникновение веществ через мембрану идет с помощью переносчиков и требует обычно затраты энергии (активное поступление). Установлено понятие свободного пространства корня, т. е. той части тканей, в которую вода с растворенными веществами может поступать путем свободной диффузии. Свободное пространство можно определить по количеству ионов, диффундирующих из ткани в воду. Однако надо иметь в виду, что часть солей, поступившая путем свободной диффузии, концентрируется (адсорбируется) на клеточных оболочках. Благодаря адсорбции обратная диффузия солей, естественно, уменьшается. Поскольку именно этот показатель служит для определения величины свободного пространства, то она оказывается преуменьшенной потому что, определяется не истинное, а кажущееся свободное пространство. Это тот объем, в котором идет свободная диффузия солей и который поддается определению. Большинство исследователей считает, что объем свободного пространства составляет 5—10% от всего объема корневой системы. Дополнительно к свободному пространству, входящему в объем клетки, в свободное пространство корня входит система межклетников. Таким образом, свободное пространство тканей является внешним по отношению к протопласту клеток и внутренним по отношению к органу в целом пли к окружающей среде.
В зоне поглощения каждого корня ионы адсорбируются на поверхности клеточных оболочек ризодермы. Из адсорбированного состояния ионы могут по коре корня передвигаться двумя путями: по апопласту и симпласту. Апопласт — это свободное пространство ткани. Симпласт — это совокупность протопластов всех клеток корпя, соединенных плазмодесмами. При поступлении в симпласт ионы должны проникнуть через мембрану и далее передвигаться по плазмодесмам к сосудам ксилемы. Ток воды с растворенными веществами, движущийся по свободному пространству (апопласту), как бы омывает все клетки коры. На всем этом пути может наблюдаться активное поступление ионов в клетки через соответствующие мембраны и включение их в обмен веществ. Необходимо учесть, что па пути движения по апопласту имеется преграда в виде клеток эндодермы с поясками Каспари. Передвижение через клетки эндодермы возможно, по-видимому, только через цитоплазму. Даже если признать, что в стенках клеток эндодермы остаются промежутки для свободной диффузии, то они настолько малы, что вещества не могут через них проникнуть. В связи с этим перенос ионов через мембраны клеток эндодермы необходим и осуществляется с помощью переносчиков.
Вопрос, что является движущей силой, заставляющей клетки корпя секретировать ионы в сосуды ксилемы, является спорным. Имеется мнение, что клетки, расположенные около сосудов ксилемы, обладают более низкой метаболической активностью по сравнению с более удаленными благодаря меньшему содержанию в них кислорода. Эти клетки отдают соли в сосуды ксилемы. Затем соли вместе с водой благодаря присасывающей силе транспирации поднимаются по сосудам вверх.
studfiles.net
Физиология растений - страница 5
| Транслосомы | небольшие вакуольки | накапливают продукты метаболизма и транспортируют их в вакуоль |
| Аппарат Гольджи | система из диктиосом (цистерн), маленьких вакуолей (везикул) | организация плазмалеммы, участие в образовании и росте клеточной стенки, участие в делении клетки |
| Пероксисомы | небольшие вакуоли, содержащие окислительно-восстановительные ферменты | участие в фотодыхании |
Лекция 6-9
Водный обмен у растений.
Дополнительная литература по теме:
Вопросы к разделу:
Формы воды в почве и их доступность для растений.
Поступление воды в растение. Двигатели водяного потока.
Передвижение воды по тканям корня.
Передвижение воды по растению.
Транспирация.
Водный баланс в растении.
Формы воды в почве и их доступность для растений.
Растения как основные автотрофы в природе, продуценты биомассы Земли, находятся в особенных условиях по отношению к окружающей неживой природе. Вода поступает в растение из почвенного раствора через корневую систему и испаряется из растения через листья. Собственно весь водный обмен в растении состоит из трех основных этапов:
поглощения воды из почвы,
передачи воды из корня ко всем органам растения,
испарение воды из листьев.
Рассматривая комплекс вопросов по механизмам водного обмена, необходимо прежде всего разобраться в вопросе о формах воды в почве и образовании собственно почвенного раствора.
В почве имеются водоудерживающие силы, которые определяют притяжение воды к почвенным частицам, поэтому далеко не вся вода, находящаяся в почве доступна растениям.
Почвенный раствор обладает собственной сосущей силой, поэтому механизм поступления воды в растение прежде всего обуславливается разницей между осмотическим давлением корневого волоска и почвенного раствора. Концентрация почвенного раствора зависит от количества солей в почве, механического состава почвы, соотношения минеральных и коллоидных частиц в почве. Вода, находящаяся в почве, в зависимости от своего состояния может находиться в одной из следующих форм:
Гравитационная - это вода, заполняющая большие почвенные капилляры, попадающая в почву при дожде или поливе, быстро двигающаяся вниз в глубокие слои почвы под действием силы тяжести собственного веса. Для растений существенного значения не имеет, так как хотя и поглощается ими, но быстро уходит из зоны почвы, где располагается корневая система.
Капиллярная - это вода, заполняющая узкие капилляры и удерживающаяся силами поверхностного натяжения менисков. Она находится в почве длительное время, незначительно притягивается к почвенным частицам, является наиболее доступной для растений формой.
Пленочная - это вода, покрывающая непосредственно почвенные частицы, удерживающаяся на их поверхности силами молекулярного притяжения или адсорбционными силами почвенных частиц. Эта вода труднодоступна для растений, поглощается в основном растениями, приспособленными к засушливым условиям, имеющими очень высокую концентрацию клеточного сока.
Гигроскопическая - это вода, находящаяся в воздушно-сухой почве, удерживаемая внутри почвенных частиц силой свыше 100000 килопаскаль. Ее количество колеблется от 5% в песчаной почве до 14% в глинистой почве. Для растений эта вода недоступна.
Имбибиционная - это вода, находящаяся внутри коллоидных частиц почвы, вызывающая их набухание, при этом в набухшей коллоидной частице создаются значительные водоудерживающие силы. Эта форма воды характерна для торфяников. Для растений она также практически недоступна.
Очень важным моментом является соотношение скорости поглощения воды из почвы и скорости испарения воды растением. При испарении воды из листьев корневая система поглощает воду в доступной зоне, в результате чего в близлежащей почве образуется зона иссушения. Корневая система, разрастаясь вширь и вглубь, поглощает воду из более дальних участков почвы, но этот процесс не бесконечен и не всегда достаточно быстро происходит. Поэтому если испарение происходит со значительной скоростью, то корневая система слишком быстро поглощает воду и оказывается полностью в зоне иссушения. В этом случае наличие в почве запасов воды не обеспечивает поглощение воды растением. Статически доступная вода оказывается динамически недоступной.
Вода в почве будет находиться в равновесном статическом и динамическом состоянии при следующих условиях:
Будет наблюдаться очень значительное насыщение почвы корнями, так что благодаря малым расстояниям между ближайшими корнями станет невозможным местное иссушение почвы. Вот почему так важно обеспечить полив растениям на первых фазах развития, когда корневая система недостаточно развита.
Будет наблюдаться медленный ток воды через растение, когда скорость поглощения воды корнями из почвы окажется равной скорости восстановления исходного содержания воды в местах иссушения. Вот почему значительную роль играет влажность воздуха, поэтому освежительные поливы в виде дождевания часто рекомендуются в качестве элемента технологии при возделывании культур в южном засушливом климате.
Для различных видов растений (засухоустойчивых или влаголюбивых) оптимальное значение влажности почвы может варьировать в достаточно широких пределах. Кроме того, для одного и того же вида растения в разные фазы его развития этот показатель также может различаться. Более того, семена растений обладают настолько большой сосущей силой, что способны при прорастании даже использовать недоступную гигроскопическую форму воды.
Наиболее важным показателем, характеризующим почву, является влагоемкость почвы. Влагоемкость почвы - это величина, количественно характеризующая водоудерживающую способность почвы.
Водоудерживающая способность почвы - это свойство почвы удерживать в себе то или иное количество влаги от стекания действием капиллярных и сорбционных сил.
Различают следующие разновидности влагоемкости:
общую,
полную,
капиллярную или относительную,
полевую или предельную или наименьшую,
максимальную молекулярную.
Для определения необходимости полива чаще всего используют понятие предельной полевой влагоемкости (ППВ). Поливы назначают при показателе влажности почвы равном 70-75% от предельной полевой влагоемкости.
Поэтому мы дадим определение именно этой разновидности влагоемкости, о подробнее о других разновидностях влагоемкости информацию можно получить из курса почвоведения.
Полевая или наименьшая или предельная влагоемкость - это наибольшее возможное содержание подвешенной влаги в данном слое почвы в ее естественном сложении при отсутствии слоистости и подпирающего действия грунтовых вод, после стекания всей гравитационной влаги.
Вторым существенным показателем для характеристики влажности почвы является коэффициент завядания. Коэффициент завядания для данной почвы - это такая величина влажности почвы при которой в специально поставленных опытах наступает длительное завядание растения. Этот показатель зависит только от типа почвы. Чем легче почва (песчаные, супесчаные), тем полнее используется растениями имеющаяся в ней вода, собственная влагоемкость почвы при этом меньше, т.е. меньше воды находится в виде мертвого запаса, недоступного растениям. Наоборот, влагоемкость тяжелых глинистых почв выше, значит и мертвый запас воды в ней больше.
Поступление воды в растение. Двигатели водяного потока.
Корневая система распространяется в почве в вертикальном и горизонтальном направлениях. Особенности распространения зависят от видовых особенностей растения. Так, у пустынных растений корневая система распространяются вглубь на десять (а отдельные виды и на большее количество) метров, а у теневыносливых растений, растущих в нижнем ярусе леса, корневая система в основном располагается в ярусе до 0.5 метра, но вширь может занимать несколько квадратных метров.
Поступление воды в корневую систему растения и перемещение ее по тканям корня осуществляется путем пассивной диффузии. Поступление идет по градиенту концентрации, поэтому если в почве концентрация почвенного раствора выше, чем концентрация клеточного сока, то вода будет диффундировать не в растение, а из него, и наступит гибель растения. Такая ситуация может сложиться в результате передозировки минеральных удобрений, небрежного внесения минеральных удобрений, когда они рассыпаются неравномерно.
Корневая система имеет поглощающую или всасывающую зону - это зона корневых волосков. Поступив в клетку корневого волоска вода становится частью живой системы - клетки растения - и подчиняется закономерностям, действующим в живой клетке. Передвижение по растению определяется двумя основными двигателями водного потока в растении:
нижним двигателем водного потока или корневым давлением,
верхним двигателем водного потока или присасывающим действием атмосферы.
Корневое давление создается при переходе воды из коры корня в сосудистую систему корня при прохождении воды через пропускные клетки перицикла, из которых вода под давлением как бы впрыскивается в сосуды ксилемы. Доказательством этого служат явления гуттации и "плача растений".
Гуттация - это выделение капельно-жидкой влаги листьями через гидатоды в условиях затрудненного испарения.
Плач растения - это вытекание пасоки (воды с растворенными в ней минеральными веществами, находящейся в ксилеме) из стеблей растений со срезанными побегами. Механизм образования корневого давления по-видимому состоит из двух аспектов:
переноса воды по законам осмоса,
дополнительной сократительной деятельности актомиозиновых белков, находящихся в перицикле и паренхимных клетках корня.
Присасывающее действие атмосферы определяется концентрацией водяных паров в атмосфере. Этот показатель в атмосфере почти всегда меньше, чем в листе растения, за исключением условий повышенной влажности воздуха, например, во время дождя, тумана.
Определяющую роль в формировании верхнего двигателя водяного потока в растении играет водный потенциал ¥ (фэта).
Водный потенциалYвыражает способность воды в данной системе, в том числе в почвенном растворе, или в клетке растения, или в атмосфере, совершить работу по сравнению с той работой, которую при тех же условиях совершила бы чистая вода.
Водный потенциал, являясь фактически мерой активности воды, определяет термодинамически возможное направление ее транспорта. Молекулы воды всегда перемещаются от более высокого водного потенциала к более низкому, подобно тому, как вода течет вниз. Водный потенциал имеет размерность энергии, деленной на объем, поэтому его выражают в барах или паскалях (1 атмосфера = 1,013 бар = 105 Па.106Па равны 1 мегаПа)
Химический потенциал воды - µw - это величина, производная от активности воды. Она выражает максимальное количество внутренней энергии молекул воды, которое может быть превращено в работу, измеряется в ДЖ. моль-1 и рассчитывается по уравнению:
µw = µw 0 + RT ln aw, где
µw 0 - химический потенциал чистой воды (принят равным нулю), R - газовая постоянная, T - абсолютная температура, aw - активность воды в системе.
продолжениеwww.coolreferat.com