Экология СПРАВОЧНИК. У растений гаметы образуются
Где образуются женские гаметы у растений. Размножение цветковых растений
Цветок - это генеративный орган растения, то есть орган, который служит для полового размножения. По своему происхождению - это укороченный видоизмененный побег, листья которого превратились в отдельные части цветка и приспособились к опылению и оплодотворению.
Тычинки и пестики определяют пол цветка. Мужские гаметы образуются в пыльниках тычинок, а женские гаметы - в семязачатках пестиков. Поэтому цветки, которые содержат только тычинки, называют мужскими цветками, а цветки, содержащие только пестики, - женскими. Цветки, в которых есть и тычинки, и пестики, называются обоеполые. Бесполые цветки не содержат ни пестиков, ни тычинок и не принимают прямого участия в размножении, их назначение заключается только в привлечении насекомых-опылителей.
Растения, на которых развиваются однополые цветки обоих видов - и мужские, и женские, называются однодомными. К однодомным растениям относятся кукуруза, береза, орешник- лещина, дуб, каштан. Если мужские цветки развиваются на одних растениях, а женские – на других, то такие растения называют двудомными. Двудомные растения - это крапива, тополь, осина, ива, конопля, щавель.
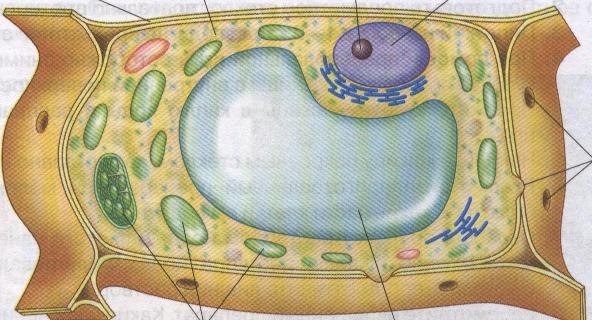
Сразу после венчика в цветке находится круг тычинок. Количество тычинок в цветке может быть разным - от одной тычинки до нескольких сотен, но обычно их 3-10. Тычинка состоит из тычиночной нити и пыльника. В пыльнике, как правило, содержится 4 полости, в которых созревает пыльца (рис. 158). По сосудам и ситовидным трубкам проводящих пучков, которые проходят внутри тычиночных нитей, к зреющей пыльце подводится питание. Снаружи пыльник покрыт эпидермисом. Под эпидермисом располагается слой клеток механической ткани. Внутри каждой половинки пыльника находится две полости. Сначала они заполнены клетками образовательной ткани. Клетки образовательной ткани делятся мейозом, поэтому получающиеся из них клетки имеют гаплоидный (одинарный) набор хромосом. Каждая вновь возникающая гаплоидная клетка делится еще один раз, но уже митозом (рис. 159).
В результате этого деления образуются две дочерние гаплоидные клетки - одна побольше и другая поменьше. Они окружают себя толстой и прочной оболочкой и получается пыльца, или точнее пыльцевое зерно (рис. 160). Таким образом, каждое пыльцевое зерно состоит из двух гаплоидных клеток, заключенных в общую оболочку. Та клетка, которая поменьше, называется генеративной клеткой, а другая клетка содержит питательные вещества (поэтому она крупнее) и называется вегетатив- ной клеткой. Почему они так называются, вам станет ясно чуть позже, когда вы узнаете, что происходит с пыльцевым зерном дальше. Постепенно вся образовательная ткань внутр. пыльника заменяется созревшими пыльцевыми зернами. Когда пыльца созревает, слой механической ткани, располагающийся под эпидермисом, начинает подсыхать, и пыльник трескается. Созревшая пыльца готова высыпаться через образовавшуюся трещину. Количество пыльцевых зерен, созревающих в одном пыльнике, может быть у разных растений от нескольких десятков до нескольких тысяч. Например, у осины их около 1000.
Размеры пыльцевых зерен тоже бывают разными - от видимых глазом (0,2 мм, как у тыквы) до мельчайших в 0,008 мм (у фикуса). Форма их - шаровидная, овальная, кубическая или даже нитевидная. Оболочка пыльцы обычно окрашена в желтоватый, бурый, синеватый или белый цвет и имеет различные неровности на поверхности в виде бугорков, шипиков, сеточки (рис. 160). Форма, окраска и поверхность пыльцы постоянна для каждой разновидности растений. Это свойство пыльцы используется, когда исследуются образцы пород из толщи земли. Пыльцевые зерна прекрасно сохраняются в ископаемом состоянии и по составу пыльцы можно узнать, какая растительность произрастала в древние времена.
В самом центре цветка находится один или несколько пестиков. В пестике различают самую нижнюю вздутую часть - завязь, отходящий от верхушки завязи столбик и самую верхнюю часть - рыльце (рис. 161). Из завязи впоследствии формируется плод. Внутри завязи находится полость, в которой находятся один или несколько семязачатков. После оплодотворения из них развиваются семена. Поэтому от количества семязачатков в завязи зависит, сколько семян будет внутри плода. Роль рыльца заключается в восприятии пыльцы, а столбик поднимает рыльце вверх, чтобы пыльце было легче попасть на рыльце. В то время, когда в пыльниках тычинок созревает пыльца, в завязи пестиков формируются зародышевые мешки, которые содержат женские гаметы - яйцеклетки.
В полости завязи пестика располагаются один или несколько семязачатков (рис. 161). Семязачатки прикрепляются к внутренним стенкам завязи посредством семяножек. Внутри каждого семязачатка есть зародышевый мешок (рис. 162). Внутри семяножки проходит проводящий пучок. По сосудам и ситовидным трубкам проводящих пучков семяножек к созревающим зародышевым мешкам подводится питание. Снаружи зародышевый мешок покрыт покровами - несколькими слоями покровных тканей. Покровы семязачатка не сплошные, их края образуют узкий канал - пыльцевход. Сам зародышевый мешок состоит из 7 клеток. Самая крупная клетка зародышевого мешка расположена в его центре и поэтому так и называется центральная клетка. Рядом с пыльцевходом находится самая главная клетка зародышевого мешка - яйцеклетка. По обеим сторонам от нее располагаются две вспомогательные клетки, а на противоположной стороне мешка (то есть напротив яйцеклетки) находятся еще три клетки, которые из-за своего расположения называются антиподами (рис. 162).
Зародышевый мешок образуется из одной единственной клетки семязачатка в результате нескольких последовательных делений этой клетки (рис. 163). Сначала она делится мейозом и как всегда в ходе мейоза образуются 4 клетки, имеющие гаплоидный (одинарный) набор хромосом. Но из этих четырех гаплоидных клеток три отмирают и оста- ется только одна. Ядро оставшейся клетки делится митозом, но перегородка между получившимися дочерними ядрами не возникает. Эти ядра расходятся к полюсам сильно удлиняющейся клетки, а между ними образуется крупная вакуоль. Затем каждое из этих ядер делится митозом еще дважды, в результате чего возникают по 4 гаплоидных ядра у каждого полюса.
В получившейся 8-ядерной клетке от каждого полюса отходит по одному ядру в центр зародышевого мешка. Они сливаются, образуя центральное ядро. Между оставшимися у полюсов клетки ядер образуются клеточные перегородки, и ядра вместе с облегающей их цитоплазмой превращаются в клетки, плотно прилегающие друг к другу. На одном полюсе зародышевого мешка - яйцеклетка и две вспомогательные клетки, а на другом - три клетки-антиподы. Центральное ядро вместе с окружающей его цитоплазмой образует центральную клетку. Таким образом, получающийся зародышевый мешок состоит из семи клеток. Из них 6 имеют одинарный набор хромосом (яйцеклетка, две вспомогательные клетки и три клетки-антиподы). В отличие от них центральное ядро, образовавшееся при слиянии двух гаплоидных ядер, обладает двойным набором хромосом.
Развитие половых клеток животных , или гаметогенез , протекает в несколько этапов. В период размножения первичные половые клетки размножаются путем митоза. В период роста каждая из них растет, достигая определенных размеров. После этого начинается процесс созревания. В результате из одной первичной мужской половой клетки образуются четыре одинаковых сперматозоида. В отличие от этого, из одной первичной женской половой клетки получается только одна яйцеклетка. Образующиеся в процессе деления три направительных тельца погибают.
У многоклеточных животных Г. происходит в специальных органах - половых железах, или гонадах (яичниках, семенниках, гермафродитных половых железах), и складывается из трёх основных этапов: 1) размножение первичных половых клеток - гаметогониев (сперматогониев и оогониев) путём ряда последовательных митозов, 2) рост и созревание этих клеток, называют теперь гаметоцитами (сперматоцитами и ооцитами), которые, как и гаметогонии, обладают полным (большей частью диплоидным) набором хромосом. В это время совершается основное событие гаметогенеза у животных - деление гаметоцитов путём мейоза, приводящее к редукции (уменьшению вдвое) числа хромосом в этих клетках и превращению их в гаплоидные клетки- сперматиды и оотиды; 3) формирование сперматозоидов (либо спермиев) и яйцеклеток; при этом яйцеклетки одеваются рядом зародышевых оболочек, а сперматозоиды приобретают жгутики, обеспечивающие их подвижность. У самок многих видов животных мейоз и формирование яйца завершаются после проникновения сперматозоида в цитоплазму ооцита, но до слияния ядер сперматозоида и яйцеклетки.
У растений гаметогенез отделен от мейоза и начинается в гаплоидных клетках - в спорах (у высших растений - микроспоры и мегаспоры). Из спор развивается половое поколение растения - гаплоидный гаметофит, в половых органах которого - гаметангиях (мужских - антеридиях, женских - архегониях) путём митозов происходит Г. Исключение составляют голосеменные и покрытосеменные растения , у которых сперматогенез идёт непосредственно в прорастающей микроспоре - пыльцевой клетке. У всех низших и высших споровых растений Г. в антеридиях - это многократное деление клеток, в результате которого образуется большое число мелких подвижных сперматозоидов. Г. в архегониях - формирование одной, двух или нескольких яйцеклеток. У голосеменных и покрытосеменных растений мужской Г. состоит из деления (путём митоза) ядра пыльцевой клетки на генеративное и вегетативное и дальнейшего деления (также путём митоза) генеративного ядра на два спермия. Это деление происходит в прорастающей пыльцевой трубке. Женский Г. у покрытосеменных растений - обособление путём митоза одной яйцеклетки внутри 8-ядерного зародышевого мешка. Основное различие Г. у животных и растений: у животных он совмещает в себе превращение клеток из диплоидных в гаплоидные и формирование гаплоидных гамет; у растений Г. сводится к формированию гамет из гаплоидных клеток.
Размножение половое семенных растений - размножение семенами нормального (не апомиктного) происхождения. Образующиеся новые особи имеют генотипы, отличные от родительских организмов.
У растений наблюдается регулярная смена ядерных фаз (гаплоидной и диплоидной). Особого внимания заслуживают цветковые растения – самые распространенные на Земле. В жизненном цикле высших растений выделяют смену двух поколений: гаметофита и спорофита. Гаметофит – небольшое растеньице полового поколения, на котором формируются половые органы, продуцирующие гаметы. На нем развиваются как женские, так и мужские гаметы. У семенных растений гаметофиты практически утратили способность к самостоятельному существованию. Преобладающим поколением являетсяспорофит (большинство клеток диплоидны), обычно представляющий собой крупное листостебельное растение, существующее достаточно длительный срок. Спорофит образуется после слияния мужских и женских гаплоидных гамет.
Цветок – основной орган размножения покрытосеменных цветковых растений. Цветок можно считать как спорофитом, органом бесполого размножения (так как он производит микроспоры и мегаспоры), так и гаметофитом – органом полового размножения (так как из микроспор развиваются мужские гаметы– спермии, а из мегаспор – женские – яйцеклетки).
Развитие пыльцевых зерен происходит в пыльцевых гнездах – микроспорангиях пыльников – в два этапа.
Этап первый – микроспорогенез спорогенной ткани делятся митозом, образуя клетки микроспор –микроспороциты (2n). Микроспороциты делятся мейозом, образуямикроспоры (n). Каждая материнская клетка дает четыре микроспоры (тетрада микроспор).
Этап второй – микрогаметогенез– развитие микрогаметофита. Каждая микроспора (n) делится митозом, образуямикрогаметофит – мужской гаметофит, илипыльцевое зерно . Сначала осуществляется процесс бесполого размножения спорофита, для чего и используются мелкие споры. Затем внутри пыльцевого мешка из прорастающей (делящейся) споры формируется микроскопический мужской гаметофит, являющийся уже новым половым поколением.
Развитие зародышевого мешка происходит в семязачатке (мегаспорангии) в два этапа. Первый этап – мегаспорогенез– развитие мегаспор. Спорогенные клетки (2n) делятся митозом, образуя клетки мегаспор –мегаспороциты (2n). Мегаспороциты делятся мейозом, образуямегаспоры (n). Каждая материнская клетка дает четыре мегаспоры. В мегагаметофите развивается только одна из микроспор (обычно нижняя), остальные дегенерируют. Второй этап –метагаметогенез– развитие мегагаметофита (зародышевого мешка). Оставшаяся из четырех однамегаспора (n) последовательно делится тремя митозами без цитокинеза (делятся только ядра). Образуется по четыре ядра на полюсах зародышевого мешка –восьмиядерный зародышевый мешок .
Два ядра от полюсов отходят к центру и сливаются вместе, образуя центральные (вторичные) ядра (2n). Остающиеся на полюсах ядра превращаются в клетки: антиподы (n),яйцеклетку (n),синергиды (n). Формируется мегагаметофит (зародышевый мешок).
Необходимо обратить внимание на тот факт, что у высших растений (в отличие от животных) процесс образования половых клеток осуществляется с помощью митоза. У всех многоклеточных животных и человека для этого используется мейоз. Мужской гаметофит у цветковых растений состоит из 3 клеток, при этом один спермий оплодотворяет яйцеклетку зародышевого мешка, а другой – центральную яйцеклетку. Происходит «двойное оплодотворение ».
Результатом полового размножения гаметофита цветкового растения является образование диплоидной зиготы и крупной триплоидной клетки. Их деление путем митоза в конечном итогеприводит к формированию зародыша и эндосперма семени (запасы питательных веществ). Семя – это важный этап в развитии нового поколения спорофита.
У позвоночных животных женские половые клетки образуются в половых железах – яичниках, а мужские – в семенниках. Именно в половых железах из исходных диплоидных клеток образуются гаплоидные гаметы.
Формирование зрелых сперматозоидов в организме млекопитающих начинается с наступлением половой зрелости, а яйцеклеток – во внутриутробном периоде развития женского организма.
В развитии половых клеток выделяют несколько стадий. Первую стадию развития половых клеток называют размножением. Для этой стадии характерно деление диплоидных клеток путем митоза. При этом из каждой материнской клетки образуются две дочерние диплоидные клетки. За счет митозов увеличивается число клеток.
Затем наступает стадия роста. В этот период размеры клеток увеличиваются. Клетки находятся в состоянии интерфазы. В них синтезируются белки, углеводы, липиды, АТФ, удваиваются хромосомы.
На стадии созревания клетки делятся путем мейоза. Число хромосом уменьшается вдвое, и из каждой диплоидной клетки образуются четыр 1000 е гаплоидные дочерние клетки.
У особей мужского пола все образовавшиеся в результате мейоза клетки одинаковые, полноценные. У особей женского пола только в одной клетке – яйцеклетке – накапливается большой запас питательных веществ, необходимых для развития будущего зародыша, остальные три маленькие клетки в дальнейшем погибают.
Развитие половых клеток завершается периодом формирования, во время которого образуются гаметы – сперматозоиды и яйцеклетка.
Своеобразно происходит формирование половых клеток у покрытосеменных растений. Гаметы образуются в тычинках и пестиках. В пыльниках тычинки содержится много диплоидных клеток, каждая из которых делится путем мейоза В результате из каждой диплоидной клетки образуются четыре гаплоидные, превращающиеся в пыльцевые зерна На этом процесс формирования пыльцы не заканчивается. Гаплоидное ядро каждого пыльцевого зерна делится путем митоза Так образуются две гаплоидные клетки – генеративная и вегетативная. Генеративная клетка еще раз делится путем митоза, в результате чего образуются два гаплоидных спермия. Спермии – мужские гаметы Они неподвижны, так как лишены жгутиков и доставляются к семязачатку через пыльцевую трубку
Таким образом, зрелое пыльцевое зерно содержит три клетки: вегетативную, или клетку пыльцевой трубки, и два спермия.
В завязи расположен семязачаток, в котором формируется женская половая клетка. В семязачатке из одной диплоидной клетки в результате мейоза образуются четыре гаплоидные клетки. Три клетки погибают, а одна оставшаяся трижды делится путем митоза. Так возникает восемь гаплоидных клеток, которые образуют зародышевый мешок. Одна из них превращается в яйцеклетку, две клетки сливаются и образуют диплоидную клетку – вторичное ядро зародышевого мешка. Оставшиеся пять клеток играют вспомогательную роль, образуя стенку зародышевого мешка.
У человека зрелая половая клетка (гамета) - это сперматозоид у мужчины, яйцеклетка (яйцо) у женщины. Перед слиянием гамет с образованием зиготы эти половые клетки должны сформироваться, созреть и затем встретиться. Половые клетки человека по структуре сходны с гаметами большинства животных. Принципиальное отличие гамет от остальных клеток организма, называемых соматическими, заключается в том, что гамета содержит только половину от числа хромосом соматической клетки. В половых клетках человека их 23. В процессе оплодотворения каждая половая клетка привносит в зиготу свои 23 хромосомы, и таким образом зигота имеет 46 хромосом, т.е. двойной их набор, как это присуще всем соматическим клеткам человека. Будучи сходны по главным структурным признакам с соматическими клетками, сперматозоид и яйцеклетка в то же время высоко специализированы для своей роли в репродукции. Сперматозоид - небольшая и очень подвижная клетка. Яйцеклетка, напротив, неподвижна и гораздо крупнее (почти в 100 000 раз), чем сперматозоид. Большую часть ее объема составляет цитоплазма, содержащая запасы питательных веществ, необходимые эмбриону в начальный период развития. Для оплодотворения необходимо, чтобы яйцеклетка и сперматозоид достигли стадии зрелости. Более того, яйцеклетка должна быть оплодотворена в течение 12 часов после выхода из яичника, в противном случае она погибает. Человеческий сперматозоид живет дольше, около суток. Быстро двигаясь с помощью своего кнутообразного хвоста, сперматозоид достигает соединенного с маткой протока - маточной (фаллопиевой) трубы, куда попадает из яичника и яйцеклетка. Обычно это занимает менее часа после совокупления. Считается, что оплодотворение происходит в верхней трети маточной трубы. Несмотря на то, что в норме эякулят содержит миллионы сперматозоидов, только один проникает в яйцеклетку, активируя цепочку процессов, приводящих к развитию эмбриона. В силу того, что сперматозоид весь целиком проникает в яйцеклетку, мужчина привносит потомку, помимо ядерного, и некоторое количество цитоплазматического материала, в том числе центросому - небольшую структуру, необходимую для клеточного деления зиготы. Сперматозоид определяет и пол потомка. Кульминацией оплодотворения считается момент слияния ядра сперматозоида с ядром яйцеклетки.
Оплодотворению у покрытосеменных предшествует микро- и мегаспорогенез, а также опыление .
Микроспорогенез протекает в пыльниках тычинок. При этом диплоидные клетки образовательной ткани пыльника в результате мейоза превращаются в 4 гаплоидные микроспоры. Через некоторое время микроспора приступает к митотическому делению и преобразуется в мужской гаметофит – пыльцевое зерно. Пыльцевое зерно снаружи покрыто двумя оболочками: экзиной и интиной.
Экзина – верхняя оболочка более толстая и пропитана спороленнином – жироподобным веществом. Это позволяет пыльце выдерживать существенные температурные и химические воздействия. В экзине находятся проростковые поры, до опыления закрытые «пробочками».
Интина содержит целлюлозу и эластична. В пыльцевом зерне имеются две клетки: вегетативная и генеративная.
Мегаспорогенез осуществляется в семязачатке . Из материнской клетки нуцеллуса в результате мейоза образуются 4 мегаспоры, из которых в результате остается только одна. Эта мегаспора сильно разрастается и оттесняет ткани нуцеллуса к интегументам, формируя зародышевый мешок. Ядро зародышевого мешка делится 3 раза митозом. После первого деления два дочерних ядра расходятся к разным полюсам: халазальному и микропилярному (находится ближе к пыльцевой трубке) , и там делятся два раза. Таким образом, на каждом полюсе находится по четыре ядра. Три ядра у каждого полюса обособляются в отдельные клетки, а два оставшихся перемещаются в центр и сливаются, образуя вторичное диплоидное ядро. На микропилярном полюсе находятся две синергиды и одна более крупная клетка – яйцеклетка. На халазальном полюсе располагаются антиподы. Таким образом, зрелый зародышевыый мешок содержит 8 клеток
Опыление заключается в переносе пыльцы с тычинок на рыльце пестика.
Оплодотворение. Пыльцевые зерна, тем или иным образом попавшие на рыльце, - прорастают. Прорастание пыльцы начинается с разбухания зерна и образования пыльцевой трубки из вегетативной клетки. Пыльцевая трубка прорывает оболочку в более тонком ее месте – так называемой апертуре. Кончик пыльцевой трубки выделяет специальные вещества, размягчающие ткани рыльца и столбика. По мере роста пыльцевой трубки в нее переходит ядро вегетативной клетки и генеративной, которая делится и образует два спермия . Через микропиле семязачатка пыльцевая трубка проникает в зародышевый мешок, там она разрывается, и ее содержимое изливается внутрь.
Один из спермиев сливается с яйцеклеткой, образуя зиготу , которая затем дает начало зародышу семени. Второй спермий сливается с центральным ядром, что приводит к образованию триплоидного ядра , развивающегося затем в триплоидный эндосперм.
Таким образом, эндосперм у покрытосеменных триплоидный и вторичный, т.к. образуется после оплодотворения.
Весь этот процесс получил название двойного оплодотворения. Он был впервые описан русским ученым С.Г.Навашиным. (1898 г.).
Оплодотворение у цветковых растений.1. половое размножение растений осуществляется
А - спорами В – почками
Б - семенами Г – вегетативными органами
2. оплодотворение у цветковых растений называется двойным, потому что
А – оно происходит подряд два раза
Б – в результате оплодотворения образуется два зародыша
В – в нем участвуют две гаметы
Г - в нем два спермия оплодотворяют две различные клетки (яйцеклетку и
Центральную клетку)
3. оплодотворением называют
А – перенос пыльцы с тычинок на рыльце пестика
Б – образование плодов и семян
В – слияние двух гамет
Г - прорастание пыльцевого зерна через рыльце и столбик пестика
4. гаметами называют:
А – женские половые клетки В – оплодотворенные яйцеклетки
Б – мужские половые клетки Г – мужские и женские половые клетки
5. женскими половыми клетками (гаметами) у цветковых растений являются:
А - яйцеклетки В – семядоли
Б - семязачатки Г - спермии
6. мужскими половыми клетками (гаметами) у цветковых растений являются:
А - пыльцевые зерна В – сперматозоиды
Б - спермии Г - семязачатки
7. семязачатки в цветках растений развиваются в
А - рыльце пестика В – завязи пестика
Б - столбике пестика Г – у разных растений поразному
8. половые клетки цветковых растений (гаметы) имеют
А – диплоидный набор хромосом
Б – гаплоидный набор хромосом
В – не имеют хромосом
Г - у разных растений поразному
9. при половом размножении цветковых растений дочерний организм чаще всего
А – имеет все признаки материнского организма
Б – имеет все признаки отцовского организма
В – имеет признаки как отцовского, так и материнского организма
Г - не имеет признаков ни отцовского, ни материнского организма
10. из яйцеклетки, слившейся с одним из спермиев, впоследствии образуется
11. из центральной клетки, слившейся с одним из спермиев, впоследствии образуется
А - зародыш семени В –семенная кожура
Б - эндосперм Г - околоплодник
12. из покровов семязачатка у цветковых растений впоследствии развивается
А - зародыш семени В –семенная кожура
Б - эндосперм Г – околоплодник
13. из семязачатков впоследствии образуются
А - семена В – гаметы
Б - яйцеклетки Г - зигота
14. эндосперм образуется в результате
А – слияния одного из спермиев с яйцеклеткой
Б – слияния одного из спермиев с центральной клеткой
В – опыления
Г - разрастания стенок завязи
15. околоплодник чаще всего образуется за счет
А – разрастания стенок завязи
Б – разрастания семязачатка
В – многократного деления оплодотворенной центральной клетки
Г - всех перечисленных факторов
16. процесс двойного опыления был открыт
А - С.Г.Навашиным В – Антонии Ванн Левенгуком
Б - К.А.Тимирязевым Г – Робертом Гуком
17. если в завязи находится несколько семязачатков, то
А – в результате образуется несколько плодов
Б – оплодотворяется только один из них и образуется один плод
В – образуется один плод с несколькими семенами
Г - оплодотворение не происходит
18. в некоторых случаях при отсутствии оплодотворения образуются плоды
А – с удвоенным количеством семян
Б – семена, которых имеют гаплоидный набор хромосом
Г - семена которых не содержат эндосперма
19. из перечисленных растений в завязи пестика развивается по одному семязачатку:
А - сливы и яблока В – вишни и абрикоса
Б - груши и огурца Г – томата и хлопчатника
У растений процесс формирования половых клеток подразделяется на два этапа:
- 1-й этап-спорогенез, который завершается образованием вегетативных гаплоидных спор,
- 2-й этап-собственно гаметогенез, в течение которого образуются зрелые гаметы.
Процесс образования мужских половых клеток складывается из:
- микроспорогенеза (образования микроспор),
- микрогаметогенеза -созревания спермиев внутри пыльцевого зерна.
Процесс образования женских половых клеток складывается из:
- мегаспорогенеза (или макро-спорогенеза),
- мегагаметогенеза -формирования зрелого зародышевого мешка, в котором образуется яйцеклетка
Процесс микроспорогенеза у цветковых растений протекает в пыльниках , причем еще в то время, когда бутон не раскрылся или, когда колос у злаковых еще в трубке, у многолетних растений все это происходит осенью. В основе микроспорогенеза лежит мейоз , в результате которого образуются четыре споры (гаплоидные вегетативные клетки). Образование пыльцевого зерна (микрогаметогенез) начинается с митотического деления ядра споры. Ядро делится митозом на два, и образуются две клетки-вегетативная и генеративная . Вегетативная клетка обладает запасом питательных веществ, которые необходимы для обеспечения роста пыльцевой трубки при прорастании пыльцы на рыльце пестика и для деления генеративной клетки на две, которые и представляют собой собственно мужские половые клетки растений. Они не обладают способностью к самостоятельному движению и называются спермиями .
Мегаспорогенез протекает в семяпочке цветковых растении, когда одна из клеток субэпидермального слоя (археспориальная клетка) обособляется, растет и превращается в материнскую клетку мегаспоры.

Этот процесс протекает во время цветения растений. В материнской клетке происходит мейоз , в результате которого образуются четыре клетки-мегаспоры . У большинства высших растений только одна развивается дальше, остальные три гибнут (моноспорический тип развития ).
После образования мегаспоры, имеющей одно гаплоидное ядро, начинается процесс формирования зародышевого мешка , в котором и образуется собственно яйцеклетка, т. е. начинается процесс мегагаметогенеза . Мегаспора растет, и у большинства растений ядро ее претерпевает три митотических деления. Крупная клетка с восьмью одинаковыми ядрами называется зародышевым мешком . Вокруг ядер обособляется плазма, поэтому эти образования иногда называют зародышевыми клетками . Две клетки, находящиеся у микропиле-входа в зародышевый мешок, называются синергидами . Они играют вспомогательную роль при оплодотворении и скоро погибают. Одна клетка-это собственно яйцеклетка. Две клетки называются полярными , при перемещении их в центр зародышевого мешка они сливаются, образуя одно диплоидное ядро. Оно называется центральным ядром зародышевого мешка . Последние три клетки чаще всего находятся на стороне, противоположной микропиле, и называются антиподами . Они играют вспомогательную роль в начале развития зиготы и затем погибают.
Процесс формирования половых клеток у растений подразделяется на два этапа: 1-й этап - спорогенез - завершается образованием гаплоидных клеток - спор, в ходе 2-го этапа - гаметогенеза - происходит ряд делений гаплоидных клеток, прежде чем образуются зрелые гаметы.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом , а процесс образования мегаспор (или макроспор) - мега- или макроспорогенезом . Микроспорогенез протекает аналогично делению созревания у животных мужских половых клеток до стадии сперматиды, а мегаспорогенез - соответственно до стадии незрелой яйцеклетки - ооцита II.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У животных после двух мейотических делений формируются гаметы, и никаких дополнительных клеточных делений не происходит. У растений в результате двух мейотических делений возникает гаплоидная спора, из которой развивается гаметофит, представляющий собой у низших растений (грибов, печеночников, мхов, ряда водорослей) целый организм и наиболее продолжительную стадию цикла существования. У высших растений гаплоидная фаза редуцирована, однако ядра мужской и женской спор претерпевают ряд митотических делений, прежде чем образуются гаметы.
Микроспорогенез и микрогаметогенез
Мы рассмотрим микроспорогенез и микрогаметогенез на примере покрытосеменных растений как наиболее общем. В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием . Каждая первичная археспориальная клетка после ряда делении становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами .
У однодольных растений каждое деление ядра в мейозе, как правило, сопровождается цитокинезом; у двудольных оба деления клетки наступают одновременно по окончании мейоза.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, кутинизированная, поверхность ее либо гладкая, либо шероховатая; приспособленная для переноса пыльцы и прилипания ее к рыльцу пестика. Этим заканчивается микроспорогенез вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, который в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые в отличие от сперматозоидов животных называются спермиоклетками, или спермиями .
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: Два из них - спермии и одно - вегетативное. При образовании пыльцевой трубки это вегетативное ядро в полужидком диффузном состоянии переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в пыльцевой трубке были впервые подробно изучены С. Г. Цвашиным в 1910 г. на лилейных растениях.
Мегаспорогенез и мегагаметогенез
У покрытосеменных растений женский гаметофит - это зародышевый мешок, который закладывается и развивается внутри семяпочки.

Развитию женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез. В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна клетка археспория растет, превращаясь в материнскую клетку мегаспоры. В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады по числу хромосом является гаплоидной. Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития), судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти и затем ее ядро претерпевает ряд эквационных делений. При этом сама клетка не делится, делится только ядро.
У разных систематических групп растений число эквационных делений ядра мегаспоры может варьировать от одного до трех. У большинства растений (70% видов покрытосеменных) этих делений, как правило, в результате возникает восемь наследственно одинаковых ядер, вовремя этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других - в противоположном конце зародышевого мешка, называемого халазальным. Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные количества цитоплазмы.
Из четырех клеток, располагающихся у микропиле, три клетки - яйцеклетка, и две так называемые синергиды образуют яйцевой аппарат. Однако из этих трех клеток после оплодотворения развивается только одна, а две другие разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца. Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное - вторичное или центральное, ядро зародышевого мешка. Это ядро с цитоплазмой зародышевого мешка называют обычно центральной клеткой зародышевого мешка. Однако часто полярные ядра, передвинувшиеся к центру, не сливаются до оплодотворения. Оставшиеся у халазального конца зародышевого мешка три ядра также обособляются в клетки; они называются антиподами .
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 наследственно одинаковых гаплоидных ядер, из которых только одно дает яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно. В одних случаях, как мы только что рассмотрели, развитие зародышевого мешка начинается из одной гаплоидной споры (моноспорический тип развития), в других - из двух (биспорический тип) и четырех спор (тетраспорический тип).

Как мы указывали, при моноспорическом типе развивается лишь одна мегаспора из четырех, а остальные три разрушаются подобно тому, что имеет место с редукционными тельцами у животных. При других типах развития зародышевого мешка сохраняется разное количество мегаспор, возникших в результате мейоза и готовых к дальнейшим митотическим делениям.

Изучая гаметогенез, нельзя не поражаться тому параллелизму, который наблюдается при созревании половых клеток у животных и растений, несмотря на то, что их расхождение (дивергенция) в филогенезе произошло на очень раннем этапе возникновения клеточной организации. Это указывает на однотипность принципов построения ряда приспособительных механизмов как в растительном, так и животном мире.
Итак, изучение развития половых клеток у животных и у растений показало, что формирование гамет является сложным процессом. Прежде чем яйцеклетка и спермий объединятся в процессе оплодотворения, они претерпевают ряд превращений. Однако половые клетки так же, как и клетки любой другой ткани, происходят из соматических. Поэтому их нельзя рассматривать как нечто обособленное от тела организма. Вместе с тем половые клетки имеют и свои особенности. Основными характерными моментами, отличающими их от соматических клеток, являются следующие:
1. У разных животных и растений на разных стадиях дифференциации тканей зародыша происходит обособление половых клеток. Процесс закладки и дифференциации, половых клеток у животных называется зачатковым путем .
2. В процессе развития половых клеток особое значение имеет мейоз с характерными для него стадиями деления ядра, а именно профазой I, во время которой конъюгируют гомологичные хромосомы, метафазой I и анафазой I, когда осуществляется редукция числа хромосом и расхождение гомологичных хромосом к различным полюсам.
3. Главным свойством половых клеток является способность их при оплодотворении сливаться в одну с образованием зиготы, которая претерпевает затем дробление и развитие. Соматические клетки этой способностью, как правило, не обладают.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
oaogpk.ru
Гаплоидные гаметы, образование - Справочник химика 21
У животных мейоз происходит непосредственно перед образованием гамет. Иными словами, в клетках каждой особи данного вида на протяжении большей части жизненного цикла содержится 2N хромосом. Такие организмы называют диплоидными. Однако это отнюдь не всеобщее правило у эукариот, размножающихся половым путем. У многих протистов мейоз происходит сразу после образования зиготы, так что эти организмы на протяжении большей части жизненного цикла содержат N хромосом. Такие организмы называют гаплоидными. У многих водорослей и растений, а также у некоторых грибов и простейших происходит чередование гаплоидных и диплоидных поколений. При таком типе жизненного цикла из диплоидной зиготы возникает диплоидная особь, образующая путем мейоза гаплоидные клетки, предназначенные для бесполого размножения. Из каждой такой гаплоидной клетки возникает гаплоид- [c.50]РИС. 15-27. Мейоз. Деление клетки, приводящее к образованию гаплоидных гамет. [c.266]
| Рис. 1.17. Жизненный цикл и образование гамет у растения. У диплоида в результате мейоза происходит образование спор. Диплоидное растение называется спорофитом гаплоидная фаза, включающая стадию созревания гамет, называется гаметофитом. Гаплоидная фаза может существовать в форме |  |
Процесс размножения делится на три основные стадии гаметогенез, оплодотворение и развитие зародыша. Гаметогенезом называют образование гамет спермиев в процессе сперматогенеза и яйцеклеток — в процессе оогенеза. Как тот, так и другой процессы происходят в гонадах в семенниках у мужчин и в яичниках у женщин. Оба процесса включают мейоз — деление ядра, в результате которого число хромосом в клетке, соответствовавшее двум наборам (диплоидное состояние), уменьшается вдвое, т. е.становится равным одному набору (гаплоидное состояние). Клетки, претерпевающие мейоз, называют материнскими клетками. Материнские клетки спермиев называют сперматоцитами, а материнские [c.76]
Мейоз представляет собой процесс деления клеточного ядра с образованием дочерних ядер, каждое из которых содержит вдвое меньше хромосом, чем исходное ядро. Мейоз называют также редукционным делением, так как при этом число хромосом в клетке уменьшается от диплоидного (2л) до гаплоидного (л). Значение мейоза состоит в том, что у видов с половым размножением он обеспечивает сохранение постоянного числа хромосом в ряду поколений. Мейоз происходит при образовании гамет у животных и спор у растений. В результате слияния гаплоидных гамет при оплодотворении восстанавливается диплоидное число хромосом. [c.145]
В гаплоидных, диплоидных и полиплоидных клетках При образовании соматических клеток и некоторых спор При образовании гамет у растений [c.158]
Сегрегация — разделение пары аллелей диплоидных организмов (в общем случае — независимое разделение различных пар гомологичных хромосом) при образовании гаплоидных гамет в процессе мейоза. [c.507]
Анеуплоидия, или гетероплоидия, возникает вследствие изменения числа хромосом, не кратного гаплоидному набору. Об этом уже упоминалось в гл. 5, где обсуждалось нерасхождение и потери Х-хромосом у D. melanogaster. В результате нерасхождения хромосом при гаметогенезе могут возникать половые клетки с лишними хромосомами, и тогда при последующем слиянии с нормальными гаплоидными гаметами они образуют зиготы 2п- -1, или трисомики, по определенной хромосоме. Если в гамете оказалось меньше на одну хромосому, то последующее оплодотворение приведет к образованию зиготы 2п— 1, или моносомика, по какой-либо из хромосом. Полисомия и моносомия могут иметь самостоятельное фенотипическое проявление вследствие изменения соотношений доз некоторых генов или нарушения генного баланса. Так, А. Блексли и Дж. Бел-линг в 20-х годах показали, что создание трисомиков по каждой из 12 хромосом у дурмана (Datura stramonium) приводит к появлению характерного, отличного от других типа растения. В частности, это выражалось в специфическом изменении формы семенной коробочки. [c.362]
Иной цели служит процесс образования половых клеток (гамет, т. е. яйцеклеток или сперматозоидов), который происходит при другом типе деления. Этот путь деления изображен на рис. 1.6. В процессе мейоза образуются клетки, содержащие гаплоидное число (и) хромосом. Этот процесс включает в себя два последовательных деления. И в этом случае хромосомы сначала удваиваются, так что клетка приступает к делению в состоянии 4и. [c.11]Теперь образование гаплоидных ядер гамет может очень просто происходить в результате второго деления мейоза, при котором хромосомы выстраиваются на экваторе нового веретена и без дальнейшей [c.15]
Образованием микроспор и мегаспор у растений заканчивается диплоидная фаза спорофита и начинается гаплоидная фаза развития гаметофита, которая, в свою очередь, завершается образованием пыльцевых зерен в пыльниках и зародышевых мешков в завязях. Формирование мужских гамет (спермиев) в пыльцевых [c.42]
Большинство высших растений размножается половым путем. Половое размножение связано с образованием гаплоидных клеток и с последующим слиянием мужских и женских гамет. Для образования гаплоидных клеток Должен существовать механизм, при помощи которого число хромосом уменьшается вдвое, т. е. клетка из диплоидного состояния переходит в гаплоидное. Без этого невозможно сохранение у видов в последующих поколениях свойственного им кариотипа. Такой переход осуществляется во время особого клеточного деления — мейоза (греч. мейозис — [c.189]
Независимое распределение геиов (Independent assortment) Распределение генов, локализованных на разных хромосомах, по гаплоидным гаметам с образованием всех возможных комбинаций генов. Лежит в основе закона Менделя о независимом распределении признаков. [c.554]
У ряда растений при переопылении тетраплоидов с диплоидами образуются не триплоидные, а гораздо чаше тетраплоидные или диплоидные гибриды. Возникновение таких гибридов обусловлено мейотическими нарушениями в процессе гаметообразования у диплоидных и тетраплоид— ных форм. Предмейотические отклонения, отсутствие редукционного деления хромосом в мейозе у диплоидных форм либо эндо-удвоение их после мейоза приводит к образованию гамет тетраплоидного типа Дополнительное редукционное деление и расхождение омосом в мейозе тетраплоидов вызывает формирование гаплоидных гамет. Появление нередунированных гамет наблюдалось у диплоидных растений кукурузы [И], сорго [12], примулы [13], ежи сборной [14], картофеля [15,16], а гамет диплоидного типа - у тетраплоидных растений капусты [17], картофеля [18], кунжута [19], петунии [20]. [c.126]
Анэуплоидия может выражаться в появлении в дочерних клетках добавочной хромосомы (и + 1), (2п + 1) и т. д. или в нехватке какой-либо хромосомы (и — 1), (2л — 1) и т. д. Анэуплоидия может возникнуть, если в анафазе I мейоза гомологичные хромосомы одной или нескольких пар не разойдутся. В этом случае оба члена пары направляются к одному и тому же полюсу клетки, и тогда разделение гомологичных хромосом в анафазе II может привести к образованию гамет, содержащих на одну или несколько хромосом больще или меньще, чем в норме (рис. 24.31). Это явление известно под названием нерасхождения. Когда гамета с недостающей или лищней хромосомой сливается с нормальной гаплоидной гаметой, образуется зигота с нечетным числом хромосом вместо каких-либо двух гомологов в такой зиготе их может быть три или только один. [c.210]
Двойной набор хромосом в одном клеточном ядре (табл. 1.2). При оплодотворении две гаплоидные гаметы (сперматозоид и яйцеклетка) сливаются с образованием дигиюид-ной зиготы. [c.200]
Образование гаплоидных гамет при мейозе и слияние двух гамет с образованием диплоидной клетки при оплодотворении. Обратите внимание на то, что у О. melanogaster, рассмотренной здесь в качестве примера, как и у других организмов, включая млекопитающих, две половые хромосомы у самца (X и V) не гомологичны друг другу. При мейозе формируются два типа [c.18]
Путем последовательных митотических делений из одной оплодотворенной яйцеклетки формируется взрослый организм. Для формирования организма человека достаточно всего 40—50 последовательных митозов. Однако образование гамет (половых клеток), имеющих гаплоидный набор хромосом, осуществляется путем мейоза — специального процесса, в ходе которого число хромосом делится надвое. При мейозе одна хромосома из каждой гомологичной пары, содержащейся в диплоидной клетке, переходит в одну из образующихся гамет. В организме, подобном As aris, который содержит единственную пару хромосом, гамета получает хромосому либо от отцовского организма, либо от материнского, но не от обоих сразу. В организмах, имеющих несколько пар хромосом, хромосомы при мейозе распределяются случайным образом, так что в каждой гамете имеются как материнские, так и отцовские хромосомы. [c.40]
Теперь образование гаплоидных ядер гамет может очень просто происходить в результате второго делеппя мейоза, при котором хромосомы выстраиваются на экваторе нового веретена и без дальнейшей репликации ДНК сестринские хроматиды отделяются друг от друга, как при обычном мнтозе, образуя клетки с гаплоидщ1м набором ДНК. Таким образом, мейоз состоит из двух клеточных делений, следующих за единственной фазой удвоения хромосом, так что из каждой клетки, вступающей в мейоз, получаются в итоге четыре гаплоидные клетки. [c.17]
У всех высших растений и животных в процессе полового размножения происходит смена ядерных фаз. При оплодотворении половые клетки (гаметы) и их ядра сливаются, образуя зиготу. Отцовское и материнское ядра вносят при оплодотворении одинаковое число хромосом (п) таким образом, ядро зиготы содержит двойной хромосомный набор (2п). Иными словами, гаметы-гаплоидные клетки (т.е. клетки с одним набором хромосом), а соматические клетки-диплоидные (с двумя наборами). Поэтому при образовании гамет следующего поколения число хромосом в клетке (2и) должно уменьшиться вдвое (2и/2 = и). Совокупность процессов, приводящих к уменьшению числа хромосом, называют мейозом или редукционным делением (рис. 2.3). Мейоз - важнейший процесс у организмов, размножающихся половым путем он приводит к двум результатам 1) к перекомбинированию отцовских и материнских наследственных факторов (генов) и 2) к уменьшению числа хромосом. Мейоз начинается с конъюгации хромосом-каждая хромосома соединяется с соответствующей (гомологичной) хромосомой, происходящей от дфугого родителя. Во время конъюгации путем разрыва и перекрестного воссоединения (кроссинговера) может происходить обмен фрагментами одинаковой длины между гомологичными хромосомами. Затем следует двукратное разделение спаренных расщепившихся хромосом, и в результате образуются четыре клетки, каждая из которых имеет гаплоидное ядро. Таким образом, в процессе мейоза не только происходит перетасовка хромосом материнского и отцовского происхождения, но может произойти и обмен сегментами между гомологичными хромосомами. Оба процесса приводят к новым сочетаниям генов (к их рекомбинации). [c.24]
Поздняя гибель зародыша характерна для реципрокных транслокаций у гаплоидных организмов и, возможно, также у диплоидных. При реципрокных транслокациях хромосомных мостов не образуется, хотя имеются ядра с нехваткой некоторых блоков генов. Поздно действующие летальные мутации предположительно вызывают гибель, когда недостающие блоки генов становятся необходимы для дифференциации. Однако доминантные летальные мутации обычно приводят к гибели организмов на ранних фазах их развития. Во время дробления образование дицентрических хромосом и продолжающееся образование мостов приводят к не-компенсируемой потере генов в ядре, которое становится все более несбалансированным по генетическому составу. Следовательно, недостающие блоки генов, обнаруживаемые в ядрах зародышей, образовавшихся из гаметы, несущей доминантную летальную мутацию, нельзя сравнивать с недостающими блоками генов среди продуктов деления реципрокной транслокации. Гибель зародыша вследствие доминантных летальных мутаций, ве- [c.122]
Полиплоидизация может быть двух типов митотическая к мейотическая. Митотическая полиплоидизация происходит в соматической ткани и сразу приводит к возникновению клетки с удвоенным набором хромосом. При мейотической полиплоидиза-ции имеет место блока да движения хромосом в первом или во втором делении мейоза, что приводит к образованию половых клеток с нередуцированным числом хромосом. Соедивение двух таких гамет дает зиготу с удвоенным числом хромосом, из которой развивается тетраплоидный организм. Однако гораздо чаще нередуцированные гаметы сливаются в процессе оплодотворения с нормальными гаплоидными, давая начало триплоид-ному организму. [c.52]
На нижней поверхности каждого спорофилла мужской шишки находятся два микроспорангия, или пыльцевых мешка. Внутри каждого пыльцевого мешка происходит мейотическое деление, приводяшее к образованию гаплоидных пыльцевых зерен, или микроспор. В них находятся мужские гаметы. У пыльцевых зерен имеется по два больших воздушных мешка, облегчающих их распространение ветром. В мае шишки становятся совсем желтыми из-за пыльцы, которая целым облаком вылетает из них. В конце лета они увядают и отпадают. [c.65]
Половым размножением называют процесс получения потомства в результате слияния двух гамет с образованием диплоидной зиготы, из которой развивается новый зрелый организм. Процесс слияния гамет называют оплодотворением. При оплодотворении сливаются ядра гамет, что приводит к объединению двух наборов хромосом, по одному от каждой родительской особи. Клетку, содержащую два набора хромосом, называют диплоидной. Гаметы гаплоидны, т. е. кажцая гамета содержит один набор хромосом. [c.55]
Менделевский закон независимого распределения тоже можно объяснить особенностями перемещения хромосом во время мейоза. При образовании гамет распределение между ними аллелей из данной пары гомологичных хромосом происходит совершенно независимо от распределения аллелей из других пар (рис. 24.7). Именно случайное расположение гомологичных хромосом на экваторе веретена в метафазе I мейоза и их последующее расхождение в анафазе I ведет к разнообразию рекомбинаций аллелей в гаметах. Число возможных сочетаний аллелей в мужсыгх или женских гаметах можно определить по общей формуле 2", где п — гаплоидное число хромосом. У человека и = 23, а возможное число различных сочетаний составляет 223 = 8 338 608. [c.190]
Перед растениями, вышедшими на сушу, встает задача обеспечения встречи мужской и женской гамет, произведенных желательно разными особями. Эта задача решалась по-разному. Отметим поэтому лишь конвергенцию, связанную с одним из возможных решений. Хорошо известно, что цветки большинства покрытосеменных опыляются насекомыми. Исходно насекомых привлекала сама пыльца, а затем растения научились выделять особую сладкую жидкость — нектар. Для улавливания переносимой насекомыми пыльцы образовался специальный орган—пестик (точнее его рыльце) или несколько стило-диев. Оказывается аналогичные образования возникли и у некоторых ржавчинных грибов (дальнейшее относится к виду Пукциния граминис). В жизненном цикле этого гриба чередуются две формы. Одиа из них, назовем ее первой, диплоидна (дикарионна) и живет на хлебных злаках. Другая, вторая форма, гаплоидна и живет иа листьях барбариса. Вторая форма развивается из гаплоидных спор, образуемых первой формой в результате редукционного деления. Первая форма в свою очередь развивается из диплоидных спор, возникающих в ходе жизни второй формы. Для формирования диплоидных спор у второй формы необходимо оплодотворение. Оно может происходить при встрече мицелиев разных полов (точнее разных знаков —полы здесь ие различима морфологически). Такая встреча возможна, если мицелин разных знаков вы- [c.238]
В результате полового процесса образуется зигота, содержащая одно купуляционное диплоидное ядро — продукт слияния двух гаплоидных ядер, происходящих из той и другой гаметы. Обычно зигота окружается толстой оболочкой, переполняется запасными продуктами и растворимым в жирах пигментом кирпично-красного цвета — астаксантин ом. Зигота прорастает или непосредственно после образования, или после более или менее продолжительного периода покоя. [c.52]
У других водорослей, наоборот, вся вегетативная жизнь осуществляется в диплоидном состоянии, а гаплоидная фаза представлена лишь гаметами, перед образованием которых и происходит редукционное деление ядра гаметическая редукция). Зиго та без редукционного деления ядра прорастает в диплоидный таллом. Эти водоросли—зеленые водоросли, имеющие сифоновое строение, все диатомовые, из бурых — представители порядка фукальные. [c.52]
Бесполое размножение — зооспорами. Они развиваются на диплоидных растениях в одногнездных спорангиях, представляющих конечные клетки коротких боковых ветвей. Содержимое их распадается на ряд голых зооспор, выходящих через разрыв оболочки спорангия. Образованию зооспор предшествует редукционное деление ядра и гаплоидные, таким образом, зооспоры вырастают в гаплоидные растения, размножающиеся половым путем. Гаметы образуются в многогнездных спорангиях. Хотя гаметы внешне одинаковы и по ювой процесс у эктокарпуса изогамный, по поведению можно различить женские и мужские гаметы. Первые быстро теряют подвижность, вторые могут оставаться подвижными до 24 ча- [c.142]
Слияние гаплоидных клеток соответствует половому процессу, свойственному высшим растениям и животным. В семенниках мужчин специализированные диплоидные клетки делятся мейотически и формируют гаплоидные спермин, а в яичниках у женщин — также в результате мейотических делений — образуются гаплоидные яйцеклетки. Последующее слияние гамет приводит к образованию ди-сглоидной зиготы, из которой в конце концов возникает многоклеточный организм (подробнее см. гл. 10). [c.115]
Мужской гаметофит (пыльцевое зерно) образуется из спорогенной ткани молодого пыльника тычинки редукционным делением ее клеток и образованием тетрады клеток, ядра которых имеют гаплоидный набор хромосом. Затем клетки тетрад становятся пыльцевыми зернами. Молодые клетки вначале одноядерные, с гаплоидным набором хромосом. В результате эква-циоииого делення содержимое пыльцевого зерна распределяется в две неравные по размеру голые клетки — вегетативную (большего размера) и генеративную (меньшего размера). После второго эквационного деления у многих растений генеративная клетка образует две новые голые клетки, которые называются спермиями, или мужскими гаметами. Следовательно, у многих растений пыльцевое зерно, покрытое двойной общей оболочкой, содержит три голые клетки вегетативную и два спермия (рис, 82). [c.487]
Диплоидный тип отличается тем, что вся вегетативная жизнь водорослей осуществляется в диплоидном состоянии, а гаплоидная фаза представлена только гаметами. Перед их образованием происходит редукционное деление ядра (гаметическая редукция). Зигота без деления ядра прорастает в диплоидный таллом. Эти растения являются диплонтами. Такой тип развития характерен для зеленых водорослей, имеющих сифоновую структуру, всех диатомовых и некоторых представителей бурых (порядок Фукусовые). [c.11]
chem21.info
Гаметы растений I III - Справочник химика 21
У человека и высших животных в результате мейоза образуются гаметы— яйцеклетка и сперматозоиды. При их слиянии возникает снова диплоидное ядро, из которого путем последовательных митозов развивается взрослый организм. Стадия мейоза характерна для жизненного цикла всех эукариот, однако отнюдь не всегда этот процесс протекает в период, аналогичный соответствующему моменту жизненного цикла человека. Так, клетки многих простейших и грибов обычно гаплоидны. После слияния двух гаплоидных ядер с образованием диплоидной клетки быстро наступает мейотическое деление, в результате которого вновь возникают гаплоидные особи. Чередование гаплоидных и диплоидных фаз жизненного цикла часто встречается у низших растений и примитивных животных. Например, гаметы папоротника падают на почву и [c.42] У животных мейоз происходит непосредственно перед образованием гамет. Иными словами, в клетках каждой особи данного вида на протяжении большей части жизненного цикла содержится 2N хромосом. Такие организмы называют диплоидными. Однако это отнюдь не всеобщее правило у эукариот, размножающихся половым путем. У многих протистов мейоз происходит сразу после образования зиготы, так что эти организмы на протяжении большей части жизненного цикла содержат N хромосом. Такие организмы называют гаплоидными. У многих водорослей и растений, а также у некоторых грибов и простейших происходит чередование гаплоидных и диплоидных поколений. При таком типе жизненного цикла из диплоидной зиготы возникает диплоидная особь, образующая путем мейоза гаплоидные клетки, предназначенные для бесполого размножения. Из каждой такой гаплоидной клетки возникает гаплоид- [c.50]Естественное или искусственное скрещивание растений и животных, когда происходит оплодотворение женской гаметы мужской, по сути своей касается переноса и слияния хромосом с последующим возникновением жизнеспособных гибридов Межвидовое скрещивание как правило сопровождается худшими результатами — получаемые гибриды обладают сниженной плодовитостью или могут быть совершенно бесплодными [c.184]
Гаплоидные клетки, приспособленные для полового слияния, называются гаметами. В типичном случае образуются гаметы двух типов крупные неподвижные яйцеклетки (или яйца) и мелкие, способные передвигаться спермин (или сперматозоиды) (рис. 14-4). Во время диплоидной фазы, начинающейся сразу после слияния гамет, клетки размножаются и специализируются, образуя сложный многоклеточный организм. У большинства животных (но не растений) полезно различать клетки зачаткового пут, от которых берет начало следующее поколение гамет, и соматические клетки, образующие весь остальной организм и не оставляющие потомства. В некотором смысле соматические клетки нужны только для того, чтобы способствовать выживанию и размножению клеток зачаткового пути. [c.8]
Это относится к мужским и женским гаметам у растений, животных и человека, несмотря на то, что с женскими гаметами дело обстоит сложнее, поскольку лишь одна из четырех клеток, образующихся в процессе мейоза из исходной материнской клетки, способна функционировать. Легко понять, что функционирующая яйцеклетка будет с одинаковой частотой нести гены Л и а, поскольку хромосомы, несущие эти гены, располагаются в метафазе мейоза случайно. И в самом деле у млекопитающих частота случаев, в которых аллель Л попадает в полярное тельце, и случаев, когда он остается в яйцеклетке, одинакова. В принципе то же самое происходит и при делении материнской клетки мегаспоры у цветковых растений. [c.44]
Мендель ввел буквенные обозначения для пар аллелей, причем доминантные аллели обозначались большими буквами, а рецессивные — маленькими. Так, например, если сорт гороха с окрашенными цветками обозначить как АА, то сорт гороха с белыми цветками будет иметь обозначение аа. Согласно Менделю, половые клетки содержат только по одному представителю из каждой пары аллелей. Поэтому у сорта гороха с окрашенными цветками гаметы должны соответственно нести аллель А, а у сорта с белыми цветками — аллель а. При оплодотворении внутри данного сорта (самоопыление или скрещивание между особями, принадлежащими к одному и тому же сорту) мы должны получить в одном случае А + А = АА, а в другом случае а + а = аа. Если скрещивают два сорта, т. е. пыльцу растения одного сорта наносят на пестик растения другого сорта, то получается гибридная особь, в которой объединение доминантного аллеля Л с рецессивным аллелем а дает конституцию Аа. Так как аллель Л доминирует над аллелем а, то у всех гибридных особей проявляется только доминантный признак, т. е. в данном примере все они имеют окрашенные цветки. Однако ген, определяющий рецессивный признак, присутствует во всех гибридах Р], не проявляясь вплоть до получения р2, в котором в среднем Д всех особей имеет белую окраску, т. е. проявляет рецессивный признак. [c.49]
Детальные исследования определения пола и относительной сексуальности были выполнены М. Гартманом и его сотрудниками на водорослях и других споровых растениях. Между прочим они показали, что не только двудомные, но и однодомные водоросли обычно образуют гаметы двух типов, которые также можно обозначить как плюс- и минус-гаметы. Только плюс- и минус-гаметы могут копулировать друг с другом и образовывать зиготы. Иногда плюс-гаметы крупнее, чем минус-гаметы, но часто они одного размера и различить их невозможно. Однако с помощью специальных методов удалось показать, что эти гаметы физиологически различны. [c.139]
Один из главных результатов исследований группы Гартмана заключается в признании того, что водоросли и грибы обладают биполярной бисексуальностью, т. е. образуют гаметы двух различных типов. Лишь полярно-противоположные гаметы могут оплодотворять друг друга. Наличие относительной сексуальности показывает, что каждый организм потенциально бисексуален, т. е. каждое растение, даже принадлежащее к двудомному виду, способно образовывать гаметы, которые в зависимости от условий могут функционировать либо как мужские, либо как женские. [c.140]
Если же превращение гена А в ген а произошло в гамете и последняя соединилась с нормальной гаметой А, то получается целиком гетерозиготное растение Аа. В следующем поколении подобное растение даст типичное моногибридное расщепление. [c.191]
Однако обычно стерильность сильнее выражена у пыльцы, чем у яйцеклеток, и по крайней мере часть женских гамет с отклоняющимся от нормы числом хромосом способна функционировать. Это дает начало анеуплоидии, т. е. появляются растения с такими хромосомными числами, которые не вмещаются в строгий полиплоидный ряд. [c.320]
Однако довольно часто образуется небольшое число функционирующих гамет, причем у этих гамет число хромосом не редуцировано они возникают благодаря образованию реституционных ядер и способны функционировать, так как содержат все хромосомы. Таким образом, они имеют ту же наследственную конституцию, как и гаметы, нормально образуемые соответствующим диплоидом. Так, нормальный диплоид дурмана имеет 24 хромосомы, а соответствующий гаплоид—12 хромосом. Все эти хромосомы качественно различны и поэтому могут функционировать лишь нередуцированные гаметы, содержащие все хромосомы. Если от подобных гаплоидов удается получить потомство, то некоторые из развившихся растений будут диплоидными и, таким образом, будет восстановлено число хромосом, типичное для данного вида. [c.331]
Однако у животных вместе с прогрессом непрерывности зародышевой плазмы носители гаплоидного состояния — гаметы несут крайне редуцированный отпечаток смены поколений. Вследствие этого у животных передается по наследству несравненно больший вес леталей и полулеталей среди индуцированных мутаций, чем у растений, где летали не выживают в гаметофите. [c.23]
Рентгеновские лучи вызвали делецию в одной из гамет растения с генотипом АА. Это привело к псевдодоминантности у части потомства. [c.295]
ЭТИМ животные клетки. Диплоидной является только зигота. С другой стороны, клетки на редкость красивой спирогиры (Spirogyra) (рис. 1-9) неподвижны, а амебоидная мужская гамета продвигается по трубочке, соединяющей две спаривающиеся клетки. Такая особенность размножения указывает на связь спирогиры с высшими зелеными растениями. [c.49]
Хемотаксис происходит и в нематодах [5] — червях, нервная система которых состоит из 300 нервных клеток. В такой простой системе можно исследовать поведение, и, используя мутантные организмы, определить его клеточную и молекулярную основу. Рассмотрим растения в качестве доказательства того, что универсальные принципы рецепции стимулов и обработки сигналов были заложены еще на ранних стадиях эволюции хемотаксис наблюдается и на гаметах бурых водорослей [6], которые узнают половые аттрактанты в морской воде и плывут к ним. Здесь же следует упомянуть слизистые грибы Si tyostelium (118Со1йеит, рост колоний которых регулируется сАМР, высвобождающегося в среду. [c.359]
У всех высших растений и животных в процессе полового размножения происходит смена ядерных фаз. При оплодотворении половые клетки (гаметы) и их ядра сливаются, образуя зиготу. Отцовское и материнское ядра вносят при оплодотворении одинаковое число хромосом (п) таким образом, ядро зиготы содержит двойной хромосомный набор (2п). Иными словами, гаметы-гаплоидные клетки (т.е. клетки с одним набором хромосом), а соматические клетки-диплоидные (с двумя наборами). Поэтому при образовании гамет следующего поколения число хромосом в клетке (2и) должно уменьшиться вдвое (2и/2 = и). Совокупность процессов, приводящих к уменьшению числа хромосом, называют мейозом или редукционным делением (рис. 2.3). Мейоз - важнейший процесс у организмов, размножающихся половым путем он приводит к двум результатам 1) к перекомбинированию отцовских и материнских наследственных факторов (генов) и 2) к уменьшению числа хромосом. Мейоз начинается с конъюгации хромосом-каждая хромосома соединяется с соответствующей (гомологичной) хромосомой, происходящей от дфугого родителя. Во время конъюгации путем разрыва и перекрестного воссоединения (кроссинговера) может происходить обмен фрагментами одинаковой длины между гомологичными хромосомами. Затем следует двукратное разделение спаренных расщепившихся хромосом, и в результате образуются четыре клетки, каждая из которых имеет гаплоидное ядро. Таким образом, в процессе мейоза не только происходит перетасовка хромосом материнского и отцовского происхождения, но может произойти и обмен сегментами между гомологичными хромосомами. Оба процесса приводят к новым сочетаниям генов (к их рекомбинации). [c.24]
Относительное развитие гаплофазы и диплофазы у разных организмов очень различно. У большинства животных гаплофаза сильно редуцирована и практически сведена к половым клеткам. У многих низших растений наблюдается противоположная картина диплофаза представлена лишь оплодотворенной женской гаметой (зиготой) в ней происходит мейоз и образуются споры, из которых затем развивается гаплофаза. [c.34]
В только что приведенном примере дигибридного расщепления одна из родительских форм несла два доминантных гена (ААВВ), а другая была рецессивна по обои.м этим генам (ааЬЬ). Точно такое же расщепление произойдет в том случае, если каждая из родительских форм имеет только один доминантный ген ААЬЬ, ааВВ) при этом родительские формы будут иметь соответственно следующие признаки красные цветки, морщинистые семена и белые цветки, гладкие семена. Соединение при скрещивании гамет АЬ и аВ приведет к образованию в р1 особей, которые, как и в первом случае, будут иметь конституцию АаВЬ. Внешними признаками этих потомков будут красные цветки и гладкие семена, и, следовательно, в этом случае особи Р1 будут отличаться от обеих родительских форм. Однако расщепление во втором поколении будет такое же, как и при скрещивании ААВВ X ааЬЬ. Самая распространенная категория растений в Рг (соответствующая [c.53]
Самооплодотворяющиеся организмы практически всегда гомозиготны и образуют только один род гамет. Гомозигот-ность в этом случае — прямое следствие способа размножения. Это явствует из того, что гетерозиготная особь Аа после самооплодотворения образует потомство, лишь наполовину состоящее из гетерозигот, тогда как другая половина потомства будет го.мозиготной расщепление в данном случае идет в уже известном нам отношении АА 2Аа аа. Если эти комбинации в свою очередь будут размножаться самооплодотворением, то потомство особей АА и аа останется гомозиготным и неизменным, тогда как в потомстве особей Аа снова будет 50% гомо- и 50% гетерозигот. Таким образом, каждое данное поколение будет соде ржать вдвое меньше гетерозигот, чем предыдущее. Если размножение самооплодотворением продолжается достаточно долго, то гетерозиготность полностью исчезнет. Именно это и произошло у спонтанно самооплодо-творяющихся высших растений. [c.76]
У плодоьой мушки частоту спонтанных мутаций можно определить значительно точнее, чем у большинства растений. Для этого разработаны специальные методы. Вычислено, что для каждого отдельного гена частота мутирования за одно поколение равна в среднем 1 1 ООО ООО поэтому какой-либо определенный ген мутирует у, плодовой мушки в рреднем лишь один раз в 40 000 лет. Но так как число генов велико и, по-видимому, равно нескольким тысячам, то нередко гаметы несут новую мутацию. Если учесть все типы мутаций, в том числе и такие, которые не дают видимых эффектов, но влияют на жизнеспособность ц плодовитость, то примерно каждая [c.195]
В жизненном цикле цветковых растений преобладает диплофаза, а гаплофаза обычно ограничена пыльцевыми зернами и зародышевым мешком. Однако интересно отметить, что в исключительных случаях могут возникать целиком гаплоидные растения. Хотя эти гаплоилы имеют такое же число хромосом, как и гаметы диплоидного растения, они развиваются [c.329]
В 1925 г. два американских исследователя Гудспид и Клаузен сообщили о получении нового константного вида табака с 72 хромосомами в результате гибридизации между 48- и 24-хромосомными видами (фиг. 161). Позднее было описано много сходных случаев. Оказалось, однако, что такие новые звенья в полиплоидном ряду возникают не только непосредственно в благодаря удвоению хромосом в оплодотворенной яйцеклетке или в молодом зародыще, как это представлял себе Винге, но и другим путем, а именно в результате образования гибридом р1 нередуцированных гамет, дающих начало аллополиплоидным растениям в Рг. [c.334]
Гетерогамность — наличие у цветковых растений генотипически различных мужских и женских гамет (например, у некоторых видов энотеры). [c.454]
В противоположность этому у покрытосеменных растений семяпочка до оплодотворения. развивается в незначительной степени и дальнейшее развитие фактически зависит от оплодотворения. Семяпочка заключена внутри завязи, которая по созревании превраш,ается в плод. Опыление происходит при участии специального рецепторного органа — рыльца. В зародышевом мешке присутствует ядро одной оосферы, эндосперм развивается в зародышевом мешке во время роста зародыша. Развитие ткани эндосперма начинается со слияния двух полярных ядер с одной из мужских гамет, так [c.466]
Подобные половые клетки, или гаметы (от греческого гамео — вступаю в брак), внешне могут в точности повторять друг друга это так называемые изогаметы (от греческого изос — равный). Однако и у таких одинаковых по внешнему виду клеток имеются различия в наследственном материале, поэтому говорят о плюс- и минус-гаметах. У других организмов гаметы уже и внешне отличаются друг от друга одни из них крупнее макрогаметы, их называют также женскими), другие мельче микрогаметы, или мужские гаметы). Но чаще всего гаметы резко различаются между собой. Одни из них очень велики и неподвижны — это яйцеклетки (которые на сей раз по праву называются женскими гаметами), другие же сохраняют подвижность — это истинные мужские гаметы, или сперматозоиды. Большинство цветковых растений и некоторые грибы пошли еще дальше по пути упразднения способности половых клеток к самостоятельному движению — яйцеклетка у них оплодотворяется не подвижной мужской гаметой, а просто ядром (мужским). Это, собственно, ничего или почти ничего не меняет, поскольку достигается главная цель — слияние ядер. Продукт слияния гамет — оплодотворенная яйцеклетка, или зигота, содержит теперь одно ядро, образовавшееся путем слияния ее собственного ядра с проникшим в нее мужским ядром. [c.117]
Однако подобный жизненный цикл ни в коем случае не является правилом. Многие растения, почти все животные и, конечно, человек — диплобионты у них оплодотворенная яйцеклетка претерпевает митотическое деление и, следовательно, все клетки развившегося из нее организма будут диплоидными, т. е. будут содержать двойной набор хромосом. Лишь при формировании половых клеток происходит, наконец, мейоз, так что гаметы (и только они), т. е. неонлодотворенные яйцеклетки и сперматозоиды, гаплоидны уже продукты их слияния снова диплоидны. Таким образом, здесь мейоз и слияние гамет чередуются как раз в обратном порядке (рис. 52). [c.136]
Как известно, нарушения в мейозе у автотетраплоицных растений сахарной свеклы сопровождаются образованием гамет с отклоняющимися числами хромосом. Анеуплоицные гаметы, с одной стороны, снижают фертильность тетраплоидных растений, а с другой, - приводят к появлению в их потомстве значительного числа анеуплоидов, присутствие которых отрицательно отражается на продуктивности тетраплоидных форм сахарной свеклы [1-61 [c.14]
Данные распределения хромосом в анафазах мейоза у растений в исходной популяции и в линиях (табл. 4, 5) показывают, что у тетра-плоидной сахарной свеклы клетки с неравным распределением хромосом возникают с высокой частотой и в преобладающем большинстве случаев составляют более трети всех материнских клеток пыльцы. Следует сказать, что в то время как подсчет хромосом в диадах не представлял особых трудностей, их подсчет в четырех ядрах, образующихся в результате второго деления, хотя и возможен у свеклы, но довольно затруднителен. В некоторых случаях это могло отразиться на точности анализа распределения хромосом в анафазе П. Несмотря на это, полученные данные достаточно четко показывают, что у тетраплоидной сахарной свеклы несбалансированные гаметы в основном образуются уже в первом делении мейоза и в дальнейшем их частота не претерпевает сколько-нибудь значительных изменений, В среднем количество анеуплоицных гамет образовавшихся в первом и во втором делении, в исходной популяции оказалось одинаковым (см. табл. 4) > а у лкнейных растений - очень близким (см табл, 3). [c.18]
Среди растений исходной популяции у двух был относительно высокий процент стерильной пыльцы, коррелирующий с высоким процентом неправильных тетрад, у всех же остальных растений в исходной популяции содержание стерильной пыльцы было низким, составляя в среднем 4,8%. Такой невысокий процент стерильной пыльцы в сопоставлении со значительно более высоким процентом образующихся анеуплоидных гамет свидетельствует о том, что анеуплоидные гаметы с хромосомными числами, близкими к эуплоидному состоянию, частично жизнеспособны и, участвуя в оплодотворении, могут давать начало анеуплоидным зиготам. [c.24]
chem21.info
Палитра знаний: Гаметогенез и оплодотворение
Что такое оплодотворение? Какое биологическое значение имеет оплодотворение?
Гаметогенез. У большинства организмов гаметы (половые клетки) образуются в специальных органах. У покрытосеменных растений спермии развиваются в пыльцевых зернах, а яйцеклетки формируются в семязачатках завязи пестика.
У большинства животных гаметы формируются в половых железах – гонадах (от греч. gone – порождающее). В семенниках образуются сперматозоиды, в яичниках – яйцеклетки.
Процесс возникновения половых клеток называют гаметогенезом (от греч. gametes – супруг, genesis – происхождение).
Процесс образования мужских половых клеток называют сперматогенезом (от греч. sperma – семя). У животных он происходит в три этапа: размножение, рост, созревание (рис. 24).
 Рис. 24. Схема гаметогенеза
Рис. 24. Схема гаметогенезаНа первом этапе (размножение) диплоидные клетки семенников многократно делятся митозом, образуя диплоидные сперматогонии – недифференцированные первичные половые клетки. На стадии роста сперматогонии увеличиваются в размерах и превращаются в сперматоциты первого порядка, которые в результате первого деления мейоза на этапе созревания образуют гаплоидные сперматоциты второго порядка. Они проходят второе деление мейоза и превращаются в сперматиды – гаплоидные половые клетки последнего периода сперматогенеза. Из сперматид постепенно формируются сперматозоиды.
Форма сперматозоидов у разных животных различна, однако строение однотипно (см. рис. 24). Каждый сперматозоид имеет головку, в которой содержится ядро с гаплоидным числом хромосом и акросома – особая структура, содержащая ферменты, способствующие проникновению сперматозоида в яйцеклетку. Средняя часть сперматозоида – шейка. В шейке находятся центриоль и митохондрии. Двигаются сперматозоиды с помощью жгутика.
Процесс образования женских половых клеток называют овогенезом (от лат. ovum – яйцо). Он происходит так же, как и сперматогенез, в три этапа.
На этапе размножения клетки зачаткового эпителия яичников делятся митозом с образованием диплоидных овогониев. Затем овогонии, пройдя стадию роста, превращаются в овоциты первого зарядка. На этапе созревания каждый овоцит первого порядка в результате первого деления мейоза образует один овоцит второго порядка, крупную по размерам клетку, и одну клетку мелких размеров – редукционную клетку. Затем следует второе мейотическое деление. Из овоцита второго порядка образуются одна гаплоидная клетка (овотида) в еще одна редукционная клетка. Овотида превращается в зрелую яйцеклетку (см. рис. 24).
Механизм овогенеза и сперматогенеза сходен, однако между этими процессами имеются и различия. Так, стадия роста в овогенезе более продолжительна, а в яйцеклетку на стадии созревания превращается лишь одна из четырех гаплоидных клеток, три гаплоидные клетки погибают.
Неравномерное деление цитоплазмы при мейозе обеспечивает будущую яйцеклетку запасом питательных веществ, что и обусловливает ее более крупные размеры по сравнению со сперматозоидом. Например, яйцеклетки млекопитающих имеют диаметр от 60 до 2000 мкм, у лососевых рыб – 6–9 мм, у страуса – несколько сантиметров. У большинства животных яйцеклетка окружена одной или несколькими оболочками, выполняющими защитную функцию.
В 1875 г. немецкий зоолог О. Гертвиг открыл очень важный процесс: слияние ядра яйца с ядром сперматозоида – оплодотворение.
Оплодотворение – это слияние гамет и образование первой клетки нового организма – зиготы (от греч. zygotos – соединенная в пару).
Половое размножение связано с оплодотворением и передачей наследственной информации от родителей потомкам. Биологический смысл оплодотворения состоит в образовании нового организма, несущего признаки обеих родительских особей.
Оплодотворение у животных. Большинство животных, обитающих в водной среде (морские кольчатые черви, пластинчатожаберные моллюски, иглокожие, рыбы, большинство видов земноводных), свои яйца и сперму выделяют в воду, половые клетки – гаметы – соединяются друг с другом случайно. Такое наружное оплодотворение – примитивный и ненадежный способ соединения гамет. Для повышения вероятности оплодотворения у многих из названных организмов в ходе эволюции выработалось приспособление производить гаметы в очень больших количествах. Например, луна-рыба откладывает до 300 млн икринок! Однако данный способ оплодотворения требует от организма огромных физических и энергетических затрат.
Более прогрессивно в эволюционном плане внутреннее оплодотворение. При внутреннем оплодотворении слияние гамет происходит в теле материнской особи. Вероятность встречи гамет при внутреннем оплодотворении достаточно высока, поэтому количество вырабатываемых яйцеклеток уменьшается, количество же сперматозоидов остается большим.
Внутреннее оплодотворение характерно для многих водных беспозвоночных, насекомых, живородящих рыб, некоторых земноводных и для всех пресмыкающихся, птиц и млекопитающих. Появление внутреннего оплодотворения в ходе эволюции способствовало утрате зависимости организмов от водной среды и освоению ими наземно-воздушной среды.
Оплодотворение у животных происходит поэтапно (рис. 25).

Рис. 25. Оплодотворение у животных
А - проникновение сперматозоида внутрь яйцеклетки; Б - образование оболочки оплодотворения и слияние гаплоидных наборов хромосом гамет
Первый этап – проникновение сперматозоида в яйцеклетку. У большинства животных яйцеклетки сохраняют способность к оплодотворению в течение нескольких часов. Если оплодотворения не происходит, половые клетки саморазрушаются и рассасываются благодаря действию ферментов лизосом.
Сперматозоиды у разных видов животных вне мужской половой системы живут от нескольких десятков секунд (у рыб) до нескольких месяцев (у пчел), У человека гаметы также недолговечны: у мужчин созревшие сперматозоиды живут до 48 ч, а у женщин в норме яйцеклетка сохраняет способность к оплодотворению от 12 до 24 ч.
Взаимодействие сперматозоида с яйцеклеткой регулируется специальными веществами, вырабатываемыми самими гаметами. При встрече с яйцеклеткой один из сперматозоидов растворяет ее оболочку специальными ферментами, а затем проникает внутрь. После этого вокруг яйцеклетки образуется оболочка оплодотворения, препятствующая проникновению других сперматозоидов (рис. 25).
Второй этап представляет собой слияние гаплоидных наборов хромосом гамет. При этом ядерные оболочки обеих гамет разрушаются, отцовские и материнские хромосомы, прикрепленные к нитям веретена деления, выстраиваются в плоскости экватора яйцеклетки. На этой стадии восстанавливается диплоидное число хромосом (2n).
На третьем этапе оплодотворение заканчивается активацией оплодотворенной клетки (зиготы) к дроблению и дальнейшему развитию (подробнее об этом будет сказано в § 25).
Оплодотворение у растений. Оплодотворение свойственно большинству растений. Ему обычно предшествует образование половых органов – гаметангиев, в которых развиваются гаметы. Половой процесс у низших растений разнообразен.
У высших споровых растений образуются гаметангии – антеридии (мужские) и архегонии (женские), в которых развиваются гаметы. В архегониях образуются яйцеклетки, в антеридиях – много сперматозоидов. У мхов и папоротникообразных вышедшие из антеридиев сперматозоиды подплывают в воде к вскрывшимся архегониям и сливаются с яйцеклетками внутри архегоний. У голосеменных растений в процессе эволюции исчезают антеридии. У покрытосеменных уже нет ни антеридиев, ни архегоний. Постепенное исчезновение у семенных растений гаметангиев связано с тем, что при наземном существовании (где часто изменяются условия среды) слишком рискованно оставаться в гаплоидном состоянии, поскольку вероятность оплодотворения в этом случае невысокая.
У семенных (голосеменных и покрытосеменных) растений оплодотворению предшествует опыление. Оплодотворение голосеменных и покрытосеменных растений, в отличие от споровых (мхов и папоротников), не зависит от воды. Именно поэтому в ходе эволюции сперматозоиды лишились жгутиков и превратились в спермии. Исчезновение зависимости полового процесса от капельной воды позволило семенным растениям широко расселиться по планете и занять территории с сухим климатом.
Двойное оплодотворение у покрытосеменных. У покрытосеменных (цветковых) растений мужские гаметы, или спермии, развиваются в пыльниках тычинок. Здесь происходит деление мейозом исходных, материнских клеток с образованием гаплоидных микроспор. Каждая микроспора делится митозом на две клетки – вегетативную и генеративную. Генеративная клетка вновь делится митозом и дает два спермия. В результате из микроспоры образуется пыльцевое зерно, которое имеет три гаплоидные клетки – вегетативную клетку и два спермия (рис. 26).
 Рис. 26. Схема двойного оплодотворения у цветковых растений
Рис. 26. Схема двойного оплодотворения у цветковых растений Яйцеклетки формируются в семязачатках завязи пестика. Материнская диплоидная клетка каждого семязачатка делится мейозом с образованием четырех гаплоидных клеток, три из которых погибают, а одна растет и превращается в мегаспору (будущий зародышевый мешок). Мегаспора претерпевает три последовательных деления митозом, в результате чего образуется восемь гаплоидных ядер (восьмиядерный зародышевый мешок). Два из них сливаются в центральное диплоидное ядро. Затем вокруг каждого ядра обособляется участок цитоплазмы, образуется клеточная стенка. В результате формируется 7-клеточный зародышевый мешок. Зрелый зародышевый мешок содержит одну диплоидную клетку (она имеет центральное диплоидное ядро) и шесть гаплоидных. Среди гаплоидных клеток можно выделить следующие: три клетки – антиподы (в оплодотворении они не участвуют, а обеспечивают питание зародышевого мешка), две – синергиды (они обеспечивают контакт одного из спермиев с яйцеклеткой) и одна – яйцеклетка (см. рис. 26).
Оплодотворение у покрытосеменных растений возможно лишь после опыления – перенесения пыльцевых зерен на рыльце пестика цветка. Здесь вегетативная клетка пыльцевого зерна образует вырост – пыльцевую трубку – и прорастает в столбик пестика, проникая в семязачаток через его отверстие – пыльцевход (микропиле).
В зародышевом мешке семязачатка один спермий сливается с яйцеклеткой, образуя зиготу, второй спермий сливается с центральной клеткой, образуя триплоидную клетку. Из диплоидной зиготы впоследствии формируется зародыш семени, из триплоидной клетки – эндосперм. Клетки эндосперма содержат большой запас питательных веществ. Таким образом семязачаток превращается в семя.
Этот своеобразный процесс, свойственный только покрытосеменным растениям, впервые описал русский ученый С. Г. Навашин в 1898 г. и назвал его двойным оплодотворением. Триплоидность эндосперма открыл его сын М. С. Навашин в 1915 г.
Значение триплоидного эндосперма заключается в очень быстром и эффективном накоплении питательного материала в семени, а значит, в очень быстром созревании самого семени. Это одна из причин господства покрытосеменных растений на Земле.
Вопросы и задания
- Какой процесс называют гаметогенезом?
- Что такое сперматогенез? Каково его значение?
- Что такое овогенез?
- В чем заключается процесс оплодотворения?
- Что понимает под наружным оплодотворением? У каких животных оно встречается?
- Какой еще тип оплодотворения существует у животных? У каких животных он встречается?
- Какие приспособления имеют животные к внутреннему оплодотворению?
- Какие этапы можно выделить в ходе оплодотворения животных?
- Как вы думаете, почему с одной и той же яйцеклеткой не могут слиться два и более сперматозоидов? В чем заключается биологический смысл данного явления?
- Что такое зигота? Какой набор хромосом у зиготы? Ответ обоснуйте.
dya4ckova.blogspot.com
Гаметы образование - Справочник химика 21
У животных мейоз происходит непосредственно перед образованием гамет. Иными словами, в клетках каждой особи данного вида на протяжении большей части жизненного цикла содержится 2N хромосом. Такие организмы называют диплоидными. Однако это отнюдь не всеобщее правило у эукариот, размножающихся половым путем. У многих протистов мейоз происходит сразу после образования зиготы, так что эти организмы на протяжении большей части жизненного цикла содержат N хромосом. Такие организмы называют гаплоидными. У многих водорослей и растений, а также у некоторых грибов и простейших происходит чередование гаплоидных и диплоидных поколений. При таком типе жизненного цикла из диплоидной зиготы возникает диплоидная особь, образующая путем мейоза гаплоидные клетки, предназначенные для бесполого размножения. Из каждой такой гаплоидной клетки возникает гаплоид- [c.50] РИС. 15-27. Мейоз. Деление клетки, приводящее к образованию гаплоидных гамет. [c.266]Самооплодотворение. Объединение мужской и женской.гамет, образованных одной и той же особью. [c.314]
Споры ипи гаметы, образованные гетеро-зиготой [c.252]
Отношение доминантных фенотипов к рецессивным как 3 1 характерно для поколения Рг при моногибридном скрещивании. Выводы Менделя о передаче каждой гаметой одного аллеля и о его фенотипическом проявлении соответствуют вероятностным законам. Вероятность того, что гамета, образованная гетерозиготной родительской особью Рь будет нести доминант- [c.183]
У человека и высших животных в результате мейоза образуются гаметы— яйцеклетка и сперматозоиды. При их слиянии возникает снова диплоидное ядро, из которого путем последовательных митозов развивается взрослый организм. Стадия мейоза характерна для жизненного цикла всех эукариот, однако отнюдь не всегда этот процесс протекает в период, аналогичный соответствующему моменту жизненного цикла человека. Так, клетки многих простейших и грибов обычно гаплоидны. После слияния двух гаплоидных ядер с образованием диплоидной клетки быстро наступает мейотическое деление, в результате которого вновь возникают гаплоидные особи. Чередование гаплоидных и диплоидных фаз жизненного цикла часто встречается у низших растений и примитивных животных. Например, гаметы папоротника падают на почву и [c.42]
Соотношение гамет, образованных растением с неизвестным генотипом, отражает обычно независимое расщепление аллелей и независимую комбинацию генов. Генотип каждой гаметы выясняется непосредственно в скрещивании с гаметой, несущей только рецессивные аллели. [c.11]
Механизм, при помощи которого хромосомы распределяются в половых клетках (гаметах), например при формировании яйцеклетки и сперматозоидов, называется мейозом (гл. I, разд. В, 3). При образовании гамет число хромосом [c.265]
Частота образования различных типов гамет будет различна в зависимости от расстояния между генами. Однако в [c.91]
Необходимость уменьшения числа хромосом вдвое при образовании гамет объясняется тем, что при оплодотворении происходит суммирование хромосом сливающихся гамет. Если бы это суммирование не компенсировалось уменьшением [c.33]
ОБРАЗОВАНИЕ ГАМЕТ У ВЫСШИХ ЖИВОТНЫХ и У ЧЕЛОВЕКА [c.38]
Половые клетки родительских форм будут соответственно иметь конституцию АВ и аЬ, и скрещивание приведет поэтому к образованию гибридов с конституцией АВ + аЬ = АаВЬ. Вследствие явления доминирования такие гибриды будут иметь красные цветки и гладкие семена, т. е. соверщенно те же признаки, как и одна из родительских форм. Среди образованных гибридами гамет одна половина будет содержать ген Л, а другая — ген а. То же самое произойдет и в отношении генов В и 6. Так как каждая гамета должна содержать по одному представителю от пары генов А-а и от пары генов В-Ь, то возможны четыре различных сочетания. Ген Л может объединиться либо с геном В, либо с геном Ь, и равным образом ген а имеет равную вероятность оказаться совмещенным с геном В или с геном Ь. В результате получатся четыре типа гамет, содержащих следующие сочетания генов АВ, АЬ, аВ [c.51]
И аЬ. Это одинаково относится как к пыльцевым зернам, так и к яйцеклеткам. Ввиду того что соединение гамет различных типов при образовании гибридами потомства происходит совершенно случайно, это случайное соединение можно выразить формулой АВ АЬ + аВ + аЬ) х АВ + АЬ + аВ - - аЬ). Результаты такого соединения станут более ясными, если 16 образующихся при этом комбинаций представить в виде следующей таблицы [c.52]
Однако довольно часто образуется небольшое число функционирующих гамет, причем у этих гамет число хромосом не редуцировано они возникают благодаря образованию реституционных ядер и способны функционировать, так как содержат все хромосомы. Таким образом, они имеют ту же наследственную конституцию, как и гаметы, нормально образуемые соответствующим диплоидом. Так, нормальный диплоид дурмана имеет 24 хромосомы, а соответствующий гаплоид—12 хромосом. Все эти хромосомы качественно различны и поэтому могут функционировать лишь нередуцированные гаметы, содержащие все хромосомы. Если от подобных гаплоидов удается получить потомство, то некоторые из развившихся растений будут диплоидными и, таким образом, будет восстановлено число хромосом, типичное для данного вида. [c.331]
В ОДНОМ опыте четыре указанные категории мух образовались с частотой 1) 145 2) 150 3) 33 4) 28, Общее число мух в этом случае составляло 356. Две кроссоверные категории мух (3 и 4) вместе были представлены 61 особью, составляя всего 17,1% общего числа мух. Очевидно, что это число показывает процент гамет кроссове рного типа в Таким образом, степень сцепления генов определяется путем скрещивания самок из Рх с самцами, рецессивными по обоим генам. При этом получаются четыре внешне различные категории потомства, которые точно соответствуют четырем типам гамет, образованным самкой р1. Процент перекреста определяется подсчетом числа полученных кроссоверных особей, которое выражается в процентах к общему числу особей. [c.89]
Скрещивание самок без Р-элемента с самцами, несущими Р-элементы, приводит у гибридов к транспозициям Р-элемента, которые наблюдаются только в клетках зародышевого пути. В потомстве таких гибридов обнаруживается достаточно много мутаций, вызванных внедрением элемента. Эги мутации часто приводят к стерильности потомства. Поэтому линии с Р-элементом и без него выглядят как репродуктивно изолированные, по крайней мере частично. Биологическая изоляция играет огромную роль в процессе эволюции. В этом случае она объясняется на молекулярном уровне изоляция линий вызвана активацией транспозиций Р-элемента, присутствующего в одной из них. Механизм активации транспозиций не расшифрован, однако выяснена причина, почему транспозиции Р-элемента ограничены зародышевыми клетками. Оказалось, что только в клетках—предшественниках гамет — осуществляется такой ход сплайсинга транскрипта Р-элемента, который приведет к образованию непрерывной открытой рамки трансляции, кодирующей транспозазу (рис. 120, а). Ограничение транспозиции зародышевыми клетками, по-видимому, имеет определенный смысл, поскольку обеспечивает выживание особей, несущих гаметы, в которых произошли геномные перестройки вследствие транспозиции Р-элемента. Подобный геномный шок , сопровождающийся высокой частотой мутагенеза, может обеспечить большую степень геномной изменчивости, которая послужит материалом для отбора в процессе эволюции. [c.232]
Путем последовательных митотических делений из одной оплодотворенной яйцеклетки формируется взрослый организм. Для формирования организма человека достаточно всего 40—50 последовательных митозов. Однако образование гамет (половых клеток), имеющих гаплоидный набор хромосом, осуществляется путем мейоза — специального процесса, в ходе которого число хромосом делится надвое. При мейозе одна хромосома из каждой гомологичной пары, содержащейся в диплоидной клетке, переходит в одну из образующихся гамет. В организме, подобном As aris, который содержит единственную пару хромосом, гамета получает хромосому либо от отцовского организма, либо от материнского, но не от обоих сразу. В организмах, имеющих несколько пар хромосом, хромосомы при мейозе распределяются случайным образом, так что в каждой гамете имеются как материнские, так и отцовские хромосомы. [c.40]
Некоторые одноклеточные водоросли достигают значительных размеров. Примером может служить Асе1аЬи1аг1а (рис. 1-9), произрастающая в теплых водах Средиземноморья и других тропических морей. Клетка этой водоросли содержит одно ядро, расположенное в ее основании (ризоиде). У взрослой водоросли, жизненный цикл которой длится от 6 месяцев (в лабораторных условиях) до 1 года (в природе), формируется характерного вида вырост (шляпка). По завершении развития этого образования ядро делится примерно на Ю вторичных ядер, которые мигрируют вверх по стебельку и в радиальные лучи шляпки, где образуются цисты. Затем шляпка отмирает и цисты высвобождаются в них происходит мейоз, и образовавшиеся жгутиковые гаметы попарно сливаются, формируя зиготу, из которой вновь вырастает диплоидная водоросль. [c.49]
Независимое распределение геиов (Independent assortment) Распределение генов, локализованных на разных хромосомах, по гаплоидным гаметам с образованием всех возможных комбинаций генов. Лежит в основе закона Менделя о независимом распределении признаков. [c.554]
Наличие квадривалентов, тривалентов и унивалентов в мейозе у тетраплоидов ведет к нарушениям в распределении хромосом и к образованию гамет с измененными числами [c.319]
У низших грибов фаза полового размножения начинается с образования половых клеток, или гамет. Если гаметы, происходящие от мужской и женской родительских клеток, морфологически неразличимы, их называют изогаметами. Гаметы образуются часто в особых морфологически дифференцированных клетках — гаметангиях. Если эти последние различны по своей форме, то мужские гаметангии называют антери-диями, а женские — оогониями. [c.57]
Грибы размножаются вегетативным, т. е. бесполым путем, но у них есть и половое размножение. Бесполое раампожекке возможно- спорами, почкованием клеток, фрагментацией гиф с образованием оидий или артроспор, а также в результате механического разрыва мицелия. Грибам свойственна высокая регенеративная способность любой обрывок мицелия в благоприятных условиях дает рост, превращаясь в организм следующего поколения. Половое размножение грибов происходит в результате слияния двух клеток — гамет (плазмогамия), сопро- [c.72]
Изменчивость прокариотов и эукариотов должна рассматриваться раздельно, так как прокариотам не свойствен тот ха-зактер изменчивости, который сопряжен с половым процессом. 3 то же время для грибов и водорослей (кроме синезеленых) как эукариотических организмов характерно множество возможностей, вытекающих из комбинации признаков родительских организмов при образовании диплоидных клеток в результате слияния гомозиготных и гетерозиготных гамет. [c.97]
Теперь образование гаплоидных ядер гамет может очень просто происходить в результате второго делеппя мейоза, при котором хромосомы выстраиваются на экваторе нового веретена и без дальнейшей репликации ДНК сестринские хроматиды отделяются друг от друга, как при обычном мнтозе, образуя клетки с гаплоидщ1м набором ДНК. Таким образом, мейоз состоит из двух клеточных делений, следующих за единственной фазой удвоения хромосом, так что из каждой клетки, вступающей в мейоз, получаются в итоге четыре гаплоидные клетки. [c.17]
К моменту вьшупления личинки в ее организме имеются две первичные половые клетки, нз которых в дальнейшем образуется около 2000 клеток, заполняющих гонаду взрослой особи. Вблизи дистального конца гонады продолжается образование первичных половых клбток путем митотических делений, в то время как в остальной части гонады они вступают в мейоз (рис. 15-68). Пролиферирующие клетки дистального конца, подобно сперматогониям у самцов млекопитающих, играют роль стволовых клеток, восполняющих убыль гамет, по мере того как те созревают и используются для размножения. [c.118]
У всех высших растений и животных в процессе полового размножения происходит смена ядерных фаз. При оплодотворении половые клетки (гаметы) и их ядра сливаются, образуя зиготу. Отцовское и материнское ядра вносят при оплодотворении одинаковое число хромосом (п) таким образом, ядро зиготы содержит двойной хромосомный набор (2п). Иными словами, гаметы-гаплоидные клетки (т.е. клетки с одним набором хромосом), а соматические клетки-диплоидные (с двумя наборами). Поэтому при образовании гамет следующего поколения число хромосом в клетке (2и) должно уменьшиться вдвое (2и/2 = и). Совокупность процессов, приводящих к уменьшению числа хромосом, называют мейозом или редукционным делением (рис. 2.3). Мейоз - важнейший процесс у организмов, размножающихся половым путем он приводит к двум результатам 1) к перекомбинированию отцовских и материнских наследственных факторов (генов) и 2) к уменьшению числа хромосом. Мейоз начинается с конъюгации хромосом-каждая хромосома соединяется с соответствующей (гомологичной) хромосомой, происходящей от дфугого родителя. Во время конъюгации путем разрыва и перекрестного воссоединения (кроссинговера) может происходить обмен фрагментами одинаковой длины между гомологичными хромосомами. Затем следует двукратное разделение спаренных расщепившихся хромосом, и в результате образуются четыре клетки, каждая из которых имеет гаплоидное ядро. Таким образом, в процессе мейоза не только происходит перетасовка хромосом материнского и отцовского происхождения, но может произойти и обмен сегментами между гомологичными хромосомами. Оба процесса приводят к новым сочетаниям генов (к их рекомбинации). [c.24]
У родительских особей наследственные детерминанты, 0пределя10ш ие окраску цветков, существуют в двух устойчивых альтернативных формах, или аллелях. Пурпурной окраске соответствует гомозиготная пара аллелей РР, а белой окраске — также гомозиготная пара аллелей рр. При образовании гамет (половых клеток) эти аллели распределяются независимо друг от друга при оплодотворении аллели рекомбинируют. [c.477]
ПРЯМОЕ ДОКАЗАТЕЛЬСТВО ОБРАЗОВАНИЯ ГЕТЕРОЗИГОТОИ ГАМЕТ Л И а В РАВНОМ КОЛИЧЕСТВЕ [c.44]
Чрезвычайно важное положение, следующее из таблицы дигибридного расщепления, состоит в том, что гены скрещиваемых сортов могут рекомбинироваться при образовании гибридами гамет. В результате становится возможным возникновение новых константных сортов с новыми сочетаниями признаков. Так, скрещивание (красные гладкие) X (белые морщинистые) приведет к возникновению в Ра новых сочетаний (красные морщинистые) и (белые гладкие). Если это выразить в виде формул, то скрещивание ААВВ У(ааЬЬ, помимо всего прочего, даст рекомбинации ААЬЬ и ааВВ. В эти сочетания входят те же самые гены, что и в прежние, однако они были перегруппированы, и эта перегруппировка, или рекомбинация, привела к образованию особей (а с их помощью, если мы захотим, и константных новых сортов) с совершенно новыми сочетаниями признаков. Такая рекомбинация представляет собой следствие того, что при. мейозе у гибридов доминантные и рецессивные гены разделяются и что гены, входящие в различные пары аллелей, наследуются независимо друг от друга. [c.53]
В только что приведенном примере дигибридного расщепления одна из родительских форм несла два доминантных гена (ААВВ), а другая была рецессивна по обои.м этим генам (ааЬЬ). Точно такое же расщепление произойдет в том случае, если каждая из родительских форм имеет только один доминантный ген ААЬЬ, ааВВ) при этом родительские формы будут иметь соответственно следующие признаки красные цветки, морщинистые семена и белые цветки, гладкие семена. Соединение при скрещивании гамет АЬ и аВ приведет к образованию в р1 особей, которые, как и в первом случае, будут иметь конституцию АаВЬ. Внешними признаками этих потомков будут красные цветки и гладкие семена, и, следовательно, в этом случае особи Р1 будут отличаться от обеих родительских форм. Однако расщепление во втором поколении будет такое же, как и при скрещивании ААВВ X ааЬЬ. Самая распространенная категория растений в Рг (соответствующая [c.53]
В гл. III подчеркивалось, что в мейозе гомологичные хромосомы от отца и от матери (у самооплодотворяющихся организмов — от мужской и женской половых клеток) сочетаются парами и что только одна из хромосом каждой пары попадает в гамету. Если особь гетерозиготна по одной паре аллелей А-а), то это означает, что одна из хромосом данной пары несет ген А, а другая — ген а. Отсюда следует, что в среднем одна половина гамет будет содержать хромосомы с геном А, а другая половина — хромосомы с геном а. Если мы имеем дигибриды АаВЬ, у которых две пары аллелей находятся в разных хромосомах, то аллели А-а и В-Ь распределятся в половых клетках независимо друг от друга. Это уже обсуждалось выше (см. стр. 33 и фиг. 8) и еще раз представлено на фиг. 29, где показано также распределение хромосом у моно-и тригибридов. Если допустить, что гены А w В лежат в черных хромосомах, а гены а и 6 — в соответствующих белых хромосомах, то распределение хромосом по типу / приведет, очевидно, к образованию гамет АВ и аЬ. Однако столь же часто распределение хромосом между гаметами будет происходить и по типу //, что приведет к образованию гамет АЬ и аВ. Таким образом, у дигибрида в среднем с одинаковой частотой образуются гаметы четырех типов. [c.83]
Если мы имеем дело с тригибридом АаВЬСс, у которого все три пары аллелей расположены в разных парах хромосом, то легко представить, что в этом случае возникнет 8 различных типов гамет. В первой метафазе мейоза в среднем с одинаковой частотой будут наблюдаться 4 типа распределения хромосом. Каждое распределение приведет к образованию гамет двух типов (фиг. 29). В случае гетерозиготности по четырем различным парам генов, расположенным в разных хромосомах, возможны 8 типов распределения хромосом в метафазе мейоза, которые приведут к образованию гамет 16 различных типов. Ясно, что все сказанное отлично согласуется с разобранной ранее формулой 2", показывающей число [c.83]
Это приводит к образованию с равной частотой гамет четырех типов. Таким же образом у тригибридов возможны и равновероятны четыре различных типа распределения хромосом. В результате образуются с равной частотой гаметы восьми разных типов. [c.84]
Как правило, подобные гетерозиготы по транслокации имеют пониженную плодовитость. Это представляет собой другое, хотя и менее четкое указание на наличие взаимного обмена между негомологичными хромосомами. Причина сниг жения плодовитости связана с образованием гамет, в которых отсутствуют определенные участки хромосом, тогда как другие присутствуют в двойном числе. [c.169]
Когда три хромосомы тривалента расходятся в мейозе, то две из них пойдут к одному полюсу, а одна — к другому. Если все семь тривалентов ориентированы одинаково (одна хромосома направлена к верхнему полюсу, а две — к нижнему), то это приведет к образованию гамет с 7 и 14 хромосомами соответственно. Однако этот исключительный случай наблюдается крайне редко. Ориентация тривалентов происходит случайно, и поэтому, например, у четырех тривалентов к верхнему полюсу направится лишь одна хромосома, а у трех других к этому полюсу направятся две хромосомы. При этом в одну клетку попадет 10 хромосом, а в другую — 11. Эти числа хромосом, приблизительно соответствующие половине сомати [c.323]
ААВВ вместо АВ. В мейозе наборы АА конъюгировали бы друг с другом с образованием 7 бивалентов, а два набора ВВ давали бы другие 7 бивалентов. Таким образом, мейоз приобрел бы закономерность, давая начало гаметам с 14 хромосомами и конституцией АВ. После оплодотворения, т. е. слияния таких гамет, возникли бы новые особи ААВВ. Иными словами, тип ААВВ явился бы новым видом с 28 хромосомами, который по своему составу представлял бы синтез исходных родительских видов АА и ВВ. Новое звено в этом ряду могло бы возникнуть при скрещивании нового вида ААВВ с третьим видом СС, имеющим 14 хромосом. Тем же уже описанным путем первичный гибрид АВС мог дать начало новому виду с 42 хромосомами ААВВСС. [c.334]
chem21.info
Гаметы
Гамета — зародышевая клетка (яйцеклетка или сперматозоид).[ ...]
Донация гамет (спермы, яйцеклеток), которая является одной из форм донации генетического материала (см. также гл. Если, например, нравственный характер безвозмездного донорства крови в настоящее время практически ни у кого не вызывает сомнений (хотя поначалу оно также порождало много моральных сомнений у сторонников традиционализма), то относительно донорства спермы или яйцеклеток имеются противоречивые оценки. Позитивные: как и любое другое донорство в медицине, это - проявление альтруизма; оно оправдывается счастьем семей, в которых с его помощью родились дети, и т.д. Негативные: это безответственное отношение к высшему дару (Бога, Природы), которым обладает всякий здоровый человек, - дару продления жизни; это вообще элементарная распущенность и свидетельство невысокого нравственного уровня человека; это проявление эгоизма или способ компенсировать свою физическую непривлекательность; в нем могут участвовать лица со скрытыми гомосексуальными наклонностями и психическими расстройствами и т.д.[ ...]
Таким образом, диморфизм гамет имеет большое биологическое значение и усиливался в процессе естественного отбора.[ ...]
Вопросы созревания и формирования гамет у корбикулы японской, обитающей в зал. Петра Великого, не изучены. В связи с этим в данной работе исследованы гаметогенез и репродуктивный цикл этого моллюска.[ ...]
При гетеро- и оогамии мужские и женские гаметы могут развиваться на одной особи или колонии (обоеполые, или однодомные, виды) или на разных (раздельно-полые, или двудомные, виды). Среди водорослей, для которых характерна изогамия, существуют гомоталличные виды (у них сливаются гаметы из одного слоевища или колонии) и гетероталличные (слияние возможно только между гаметами из разных особей), которые ввиду отсутствия морфологических различий обозначаются знаками + и —, соответственно различают - -гаметы и —гаметы.[ ...]
Как видим, с моральной точки зрения донация гамет оценивается далеко неоднозначно. Называя ИОД методом лечения, врачи и сами доноры морально оправдывают такую практику. Противники же ИОД все свои контраргументы сводят к категорическому неприятию “суррогатного отцовства и материнства”.[ ...]
Женские гаметы крупные, неподвижные; мужские — более мелкие, двужгутиковые. При переходе к половому размножению часть клеток в верхней половипе слоевища подвергается последовательному делению и дает начало многослойной половой ткани. В пределах этой ткани хорошо различаются темные и светлые участки, располагающиеся попарно и соответствующие скоплениям женских и мужских клеток. В каждой клетке образуется в конечном итоге по одной гамете. Зигота, как и апланоспоры, прорастает в пластинчатое слоевище. Гаметофит, вплоть до образования половых клеток, морфологически подобен спорофиту, поэтому цикл развития празиолы можно считать изоморфным. Своеобразие его заключается в необычном строении половой ткани.[ ...]
Поскольку при половом процессе в результате слияния гамет и их ядер происходит удвоение набора хромосом в ядре, то в последующем в какой-то момент цикла развития наступает редукционное деление ядра (мейоз), в результате которого дочерние ядра получают одинарный набор хромосом. Спорофиты многих водорослей диплоидные, и мейоз в цикле их развития совпадает с моментом образования спор, из которых развиваются гаплоидные гаметоспо-рофиты или гаметофиты. Такой мейоз называют спорической редукцией (рис. 2Ъ,1).[ ...]
Совокупность всех генов во всех их аллельных формах в гаметах организмов, составляющих популяцию, представляет собой пул генов, тогда как совокупность всех генотипов популяций называют их генофондом. В случае индивидуального диплоидного организма частота какого-либо аллеля может составлять 100%, 50% или 0%, но в популяции организмов частота этого аллеля является функцией количества (%) индивидуумов в популяции, которые имеют этот ген. Поэтому в популяции организмов частота аллеля может составлять от 0 до 100%. Стойким является тот генетический пул, в котором частоты генов, независимо от их функций, являются постоянными во всех следующих одно за другим поколениях организмов.[ ...]
Следовательно, если допущения случайного объединения гамет при оплодотворении и равной во всех случаях жизнеспособности гибридов справедливы, тогда среди гибридов Р2 три четверти особей должны давать круглые семена и одна четверть — шероховатые, т. е. растения, дающие семена разной формы, среди гибридов должны встречаться в отношении 3:1, которое является фенотипическим отношением.[ ...]
Как правило, мутации количества хромосом происходят в гаметах одного из родителей. Поэтому, все клетки организма, в зачатии которого принимала участие одна из мутантных гамет, будут содержать аномальный хромосомный набор. Однако иногда количественные хромосомные мутации могут случаться в процессе первых делений зиготы, образованной нормальными гаметами. Из такой зиготы разовьется организм, часть клеток которого будет иметь нормальный диплоидный набор, другая же часть — аномальный. Это явление называют хромосомным мозаицизмом, а индивидов, обладающих мозаицизмом — хромосомными мозаиками. Мозаицизм более част по половым хромосомам. Такие мозаики имеют генотип Х/ХХ, Х/ХУ, ХХ/ХУ, ХХУ/ХХ.[ ...]
С усилением половой дифференциации количество мужских гамет в гаметангии возрастало. Так получился антеридий высших растений с его многочисленными мелкими сперматозоидами. Количество женских гамет, напротив, уменьшилось. Таким путем возник в конце концов архегоний с одной большой и неподвижной яйцеклеткой. В эволюции архегония стерилизация сказалась весьма сильно, а сама женская гамета лишилась жгутиков и стала неподвижной. Шейковая и брюшная канальцевые клетки архегония также были когда-то гаметами, но в процессе эволюции стерилизовались. У разноспоровых растений в связи с редукцией их гаметофитов редуцируются и видоизменяются также аптеридии и архегонии, а у голосеменных родов вельвичия и гнетум и у всех цветковых растений они уже вовсе не образуются.[ ...]
Оплодотворение посредством объединения ядер разнополых гамет обеспечивает сочетание генетических вкладов обеих родительских особей в одной клетке. В результате у образующихся особей следующего поколения наблюдается генетическая изменчивость, определяемая этими вкладами. При этом передача потомку признаков родителей является не случайным явлением и определяется законами генетики. Именно на эту наследственную изменчивость действует естественный отбор, что и влечет за собой эволюционное развитие организмов.[ ...]
Самое главное событие, происходящее при объединении двух гамет в результате оплодотворения, - это слияние гаметных ядер. Особенностью такого слияния является то, что в результате слияния ядер двух клеток - гамет в ядре зиготы сохраняется число хромосом, которое характерно и постоянно для данного вида. Следовательно, для того, чтобы после слияния двух гамет зигота имела число хромосом, характерное для генома данного вида, гаметы должны содержать вдвое меньше хромосом, чем зигота и все соматические клетки организма.[ ...]
У водорослевого предка высших растений мужские и женские гаметы отличались друг от друга, вероятно, очень слабо и были все снабжены жгутиками. Но у высших растений половая дифференциация усилилась и привела к резкому диморфизму гамет. Женские гаметы (яйцеклетки) накапливают питательные вещества и поэтому более крупны и неподвижны. Мужские гаметы (сперматозоиды) почти совершенно лишены запасных питательных веществ. У низших групп высших растений они сохраняют свою подвижность. Мужские гаметы потеряли свои жгутики у большинства голосеменных и у всех цветковых растений, но здесь это компенсировалось специальным приспособлением для их доставки — пыльцевой трубкой. Такое неравномерное распределение питательного материала между женскими и мужскими гаметами обеспечивает большее число копуляций (слияний) при одной и той же массе, чем если бы эта масса питательных веществ была распределена равномерно между гаметами.[ ...]
В животном царстве мейоз ведет к образованию половых клеток - гамет и обычно только эти клетки содержат гаилоидный набор хромосом. У растений мейоз может происходить на разных этапах жизненного цикла, причем в качестве гаплоидных продуктов у них образуются как половые клетки - гаметы, так и бесполовые споры. Продолжительность жизни гамет ограничена и составляет от нескольких минут до нескольких суток, после чего неоплодотворенные гаметы пропадают.[ ...]
Кроме вегетативного, известно и половое размножение — анизогамия. Гаметы образуются в верхней части пузырей в результате преобразования и деления участков цитоплазмы, которые отделяются от остальной части их лишь тонким цитоплазматическим слоем. Гаметы выходят под давлением через несколько отверстий. Один и тот же пузырь плодоносит обычно несколько раз. Бесполого размножения нет.[ ...]
Биологическое значение оплодотворения состоит в том, что при слиянии гамет восстанавливается диплоидный набор хромосом, а новый организм несет наследственную информацию и признаки двух родителей. Это увеличивает разнообразие признаков организмов, повышает их жизнестойкость.[ ...]
Для выхода гамет служит отверстие с неровными краями во внешней стенке клетки (рис. 219,7). Зигота развивается в одноклеточный спорофит. При этом она силь-по увеличивается в размерах и покрывается толстой оболочкой (рис. 219, 5). Через более или менее длительное время содержимое этого одноклеточного растения делится с образованием зооспор. Таким образом, здесь смена форм развития гетероморфная.[ ...]
Ганлоидный набор, входящий в состав диплоидного и происходящий из отцовской гаметы, несет отцовскую наследственность с ее индивидуальными чертами, а гаплоидный набор из материнской гаметы — материнскую. Сложное взаимодействие однородных, но не всегда тождественных генов двух гаплоидных наборов, в сумме образующих один диплоидный, определяет, какие признаки проявятся у диплоидпого потомства, которое, по существу, является гибридом отца и матери.[ ...]
Имеются сведения о том, что у медаки Oryzias latipes и фундулюоа Fundulus heteroclitus мужские гаметы более чувствительны к облучению, чем женские (Solberg, 1938а, Ь).[ ...]
Биологическое значение оплодотворения состоит в том, что при слиянии разнополых гамет (происходящих, как правило, от двух разных особей) образуется новый организм, несущий в себе признаки и матери и отца. Образующиеся после оплодотворения новые организмы могут сочетать в себе признаки обоих родителей в самых различных комбинациях, что обеспечивает колоссальное увеличение наследственного разнообразия организмов. Место мейоза оплодотворения в жизненном цикле животных, размножающихся половым путём, показано на рис. /.[ ...]
Возникновение зародышей из синергид дает право утверждать, что эти клетки являются потенциальными гаметами и при соответствующих условиях способны дать начало новому растению. Значительно реже происходит развитие зародышей из антипод. Такой тип размножения описан лишь у Allium odorum и Ulmus glabra.[ ...]
Полостная жидкость содержит биологически активные вещества, способствующие сближению и соединению гамет в процессе осеменения. Эти вещества, или «гамоны» (гормоны гамет) (Hartmann, Schartau, 1939), являются продуктом секреции яйцеклетки. Различают гиногамоны и андрогамоны — вещества, содержащиеся соответственно в икринках и спер-миях. Гиногамоны локализованы в разных частях яйца и не всегда переходят в полостную жидкость (табл. 13).[ ...]
Проблеме плодовитости полиплоидов придается громадное значение. Тем не менее, бывают случаи, когда стерильность гамет не вызывает опасений и даже желательна. Так, возделываемая в качестве салата триплоидная настурция (Nasturtium officinale) отличается пониженной плодовитостью, но прекрасно размножается вегетативным путем.. Многие культурные растения, размножаемые луковицами (тюльпаны, нарциссы, канна и желтые лилии), также являются триплоидами. Например, значительное число ценных американских сортов яблонь является триплоидами. У двулетних овощных культур, образующих в 1-й год жизни корнеплоды и клубни, стерильность не является препятствием при промышенном возделывав нии. Так обстоит дело и с триплоидным гибридом сахарной свеклы, отличающимся высоким содержанием сахара в соке, большой массой корня, а следовательно, и большой урожайна-стью сахара с 1 га. Триплоидные гибриды, полученные таким образом, по урожайности превышают диплоидные сорта.[ ...]
Имея в виду соображения, приведенные выше, вернемся к гибридам Р1, которые являются гетерозиготами Иг и продуцируют гаметы К и г в равных количествах. Допуская, что при оплодотворении гаметы объединяются случайно (нет преимуществ для объединения гамет К и Я или гиг, или И и г) и что все формирующиеся гибридные растения жизнеспособны, можно предположить, что в результате расщепления генов формируемые гибриды будут различными как по фенотипу, так и по генотипу (см. рис. 124).[ ...]
Род характеризуется изоморфной сменой форм развития и отличить половые растения от бесполых по внешнему виду невозможно. Зрелые гаметы и зооспоры выходят в окружающую среду через одно округлое отверстие — пору, образующуюся на вершине сосочкообразного изгиба во внешней стенке клетки (рис. 218, 2). При прорастании зооспор и зигот энтероморфа, как и все водоросли семейства ульвовых, проходит стадию однорядной нити (рис. 218, 3—7). Клетка, от которой начинается развитие, делится на две — апикальную и базальную. В результате поперечных делений первой образуется вертикальная нить, вторая превращается в первичный ризоид. Позднее нить преобразуется в трубчатое слоевище. При изменении условий среды первые деления могут вести к образованию стелющихся нитей, расположенных в форме диска, из клеток которого уже позднее вырастает одна или несколько вертикальных однорядных нитей.[ ...]
Половое размножение — это образование нового организма при участии двух родительских особей. При половом размножении происходит слияние половых клеток — гамет мужского и женского организма. Новый организм несет наследственную информацию от обоих родителей. Половые клетки формируются в результате особого типа деления. В этом случае в отличие от клеток взрослого организма, которые несут диплоидный (двойной) набор хромосом, образующиеся гаметы имеют гаплоидный (одинарный) набор. В результате оплодотворения парный, диплоидный набор хромосом восстанавливается. Одна хромосома из пары является отцовской, а другая — материнской. Гаметы образуются в половых железах или в специализированных клетках в процессе мейоза.[ ...]
Мужские половые клетки — сперматозоиды, мелкие, часто подвижные, имеют жгутики, их образуется значительно больше, чем яйцеклеток. У семенных растений мужские гаметы неподвижны и называются спермиями.[ ...]
У одних видов органы бесполого и полового размножения развиваются на разных особях; тогда растения, образующие споры, называют спорофитами, а растения, производящие гаметы,— гаметофитами. У других водорослей споры и гаметы образуются на одних и тех же растениях; одновременно у таких видов могут существовать и особи, дающие только споры, т. е. спорофиты (порфира). Сейчас растения, способные производить и споры (зооспоры) и гаметы, как правило, называют гаметофитами. Однако во избежание путаницы с настоящими гаметофитами, производящими только гаметы, их лучше называть гаметоспо-рофитами.[ ...]
Деление сперматогенной клетки обычно происходит перед оплодотворением. Она переходит в пыльцевую трубку, сопровождаемая клеткой-ножкой, и, делясь, дает начало двум мужским гаметам. В отличие от сперматозоидов саговниковых и гинкго, снабженных эффективно действующим локомоторным аппаратом — жгутиками, они лишены жгутиков и поэтому способны лишь к слабым амебоидным движениям. Как мы уже знаем, такие без-жгутиковые мужские гаметы называют спер-миями. Потеря жгутиков связана с тем, что в процессе эволюции функция доставки гамет к яйцеклеткам перешла к пыльцевой трубке. Этот переход функции доставки гамет к пыльцевой трубке не только более эффективен, но он, кроме того, привел к значительному упрощению спермиев.[ ...]
Мейоз представляет логически необходимую часть жизненного цикла, размножающегося половым путем, Мейоз обеспечивает расщепление генов - отдельных участков ДНК по отдельным гаметам, в результате чего происходит разнообразное сочетание генов в гаметах. В отношении поддержания постоянства хромосом в клетке оплодотворение составляет антитезу (противоположное) мейозу, о процессе оплодотворения происходит слияние гаплоидных ядер двух разнополых гамет с образованием одной клетки - зиготы с диплоидным ядром.[ ...]
Как видно, гибриды ?! являются гетерозиготными растениями по двум парам генов, т. е. растениями ИгУу. Если иметь в виду лишь форму семян, то гибриды Г с генотипом Иг будут продуцировать гаметы И и г поровну. Если же иметь в виду одновременно форму и окраску семян, то каждая гамета, продуцируемая гибридом ?! с генотипом КгУу, будет нести не только один генный аллель для формы семян, но и один генный аллель для окраски семян.[ ...]
Определение частот спонтанных мутаций позволяет определить вероятность мутаций в каждом новом поколении людей: 1 х 10 ® мутаций на ген х5х 10“4 генов (гаплоидный геном) = 5 х 10“2 мутаций на гамету (5 : 100 или 1 : 20). Далее, 1 : 20 х 2 гаметы на зиготу = 1 : 10 случаев, что каждая гамета несет новую мутацию. Возможно, что это очень высокая частота, но ее достоверность определяется тем, что большинство мутаций рецессивно и, следовательно, не экспрессируется у гетерозигот.[ ...]
Как мы уже отмечали, в соответствии с заключением Т. Моргана гены сцеплены тогда, когда локализованы на одной хромосомной паре. Допуская, что хромосомы остаются интактными при вступлении их в гаметы, локализованные на них гены всегда наследуются вместе. В этом случае можно говорить об их полном сцеплении. Однако, рассматривая сцепление генов у кукурузы, мы видели, что полного сцепления генов не бывает, поскольку происходит формирование гамет не только двух родительских типов, но и гамет рекомбинантных типов (вследствие рекомбинации генов). Об отсутствии полного сцепления свидетельствуют также данные, полученные при изучении других организмов, механизм этого явления заключается в том, что в процессе гаметогенеза хромосомы клеток могут подвергаться разрывам в одном или нескольких местах, а сегменты, образующиеся в результате разрыва одной хромосомы, могут смыкаться с сегментами гомологичной хромосомы при условии, что в последней тоже были разрывы, причем в аналогичных местах. Как мы видели, гены С и в у кукурузы в 97% случаев (гамет) остаются сцепленными в родительских комбинациях и примерно в 3% случаев (гамет) они не связаны между собой и находятся в рекомбинантных сочетаниях. Обмен между хромосомными сегментами гомологичных хромосом, сопровождаемый рекомбинацией сцепленных генов, получил название кроссинговера (Т. Морган), а явление, обусловленное этим механизмом, называют генетической рекомбинацией. В результате рекомбинации из двух исходных комбинаций генов создается новая комбинация. Возвращаясь к случаю рекомбинации генов у кукурузы, можно сказать, что кроссинговер произошел в сегменте хромосомы между локуса-ми (местами), занимаемыми генами Сив, вследствие двух разрывов в этих участках хромосомы у отдельных клеток.[ ...]
Очевидно, такие гаметангии не могли удовлетворить потребностям наземного существования. С переходом водорослевого предка высших растений к наземному образу жизни возникла прежде всего потребность в защите гамет от высыхания. Как в мужских, так и в женских гаметангиях самые наружные клетки должны были в зтих условиях стерилизоваться, т. е. потерять способность производить гаметы, и после соответствующих видоизменений превратиться в защитный слой. Предполагают, что именно таким путем, т. е. путем стерилизации наружного слоя клеток, развилась стенка ан-теридиев и архегониев.[ ...]
Самая яркая отличительная черта конъюгат, позволяющая объединять их вместе в особый класс, несмотря на разнообразие внешнего вида,— это полное отсутствие подвижных стадий размножения, т. е. зооспор и подвижных гамет, и наличие своеобразного полового процесса, получившего специальное название — конъюгация (лат. сощ аНо — слияние, соединение). Во время конъюгации происходит слияние протопластов двух клеток, которые выполняют здесь функцию гамет. В результате такого слияния, происходящего через специально образующийся канал, получается одноклеточная, покрытая толстой оболочкой зигота. Внешне две сливающиеся (конъюгирующие) клетки совершенно неразличимы; физиологическая дифференциация на мужскую и женскую клетки хотя и определенна, но узнать ее можно лишь косвенно и спустя некоторое время — по положению зиготы. В том случае, когда скорости передвижения (вернее, перетекания) сжавшихся протопластов-гамет одинаковы, зигота образуется в конъюгационном канале, иногда разрастаясь вплоть до степок конъюгирующих клеток-гаметангиев. В этом случае разделение клеток на мужскую и женскую крайне условно, а сам процесс слияния равноценных по морфологии и скорости перетекания гамет принято называть изогамией (греч. «изос»— равный, «гамос»— брак). Эта форма полового процесса характерна для большинства водорослей-конъюгат. И здесь протопласты-гаметы обеих конъюгирующих клеток одинаковы по размерам, но одна из них (мужская) активно перетекает через конъюгационный канал из отдающей клетки в воспринимающую, где и сливается со сравнительно неподвижной гаметой (женской). В этом случае зигота формируется только в воспринимающей (женской) клетке.[ ...]
Жизненные циклы. Жизнь организма состоит из различных форм и стадий развития. Жизненный цикл высших организмов, включая человека, тождествен половому циклу. О последовательности стадий развития более правильно будет сказать не «от чрева до могилы», а «от гамет до оплодотворенной яйцеклетки к взрослой особи до гамет». Жизненный цикл низших организмов может быть значительно сложнее и часто состоит из ряда длительных стадий существования. Половой цикл у них может включать и ряд бесполых форм размножения. Кроме того, многие низшие организмы, например бактерии, обычно не проходят через половую стадию, а существуют в одной длительной форме или используют только бесполые споры, как, например, у несовершенных грибов.[ ...]
Итак, генные пары двух скрещиваемых организмов расщепляются (сегрегируют) независимо одна от другой и перераспределяются, подвергаясь свободной рекомбинации между собой, причем гибридные организмы в процессе их гаметогенеза дают начало комбинациям родительских гамет и рекомбинациям гамет в равных количествах. Данное правило, установленное Г. Менделем, в настоящее время называют вторым законом наследственности (вторым законом Г. Менделя), законом независимого перераспределения генных аллелей или законом рекомбинации генов.[ ...]
В пыльниках М 2 обычно только часть пыльцы является фертильной, причем количество ее, а также число растрескивающихся пыльников изменяются в широких пределах в зависимости от генотипа, температуры, влажности и других условий среды, при которых происходит формирование гамет и цветение.[ ...]
Бесполое размножение у каулерповых отсутствует, по-видимому, оно было утрачено у современных форм в процессе эволюции. Половое размножение — анизогамия — характеризуется рядом особенностей, отличающих каулерповых от других семейств порядка. Во-первых, у каулерпы нет специальных органов размножения — гаметангиев. Гаметы образуются непосредственно в ассимиляционных нитях в любой их части. На отдельных участках цитоплазма сгущается, приобретает темно-зеленую окраску, затем сетчатое строение и наконец делится с образованием одноядерных гамет. Какие-либо перегородки, отделяющие место образования гамет, отсутствуют. Такое размножение называют г о л о к а р п и е й. Для выхода гамет на поверхность слоевища образуются довольно длинные выросты — папиллы; гаметы освобождаются в результате разрыва оболочки на вершинах папилл.[ ...]
Весь ;>тот процесс, как известно, принято намывать двойным оплодотворением. Ведь согласно общепринятому в литературе определению, оплодотворение (еипгамин) — это процесс слияния, мужской п женской половых клеток (гамет) с образованием зиготы, из которой и дальнейшем разнимается новый организм. Такое определение оплодотворения можно найти в любом учебнике биологии и в любом :нщнклопедичоском словаре (в том число в советских шщпклопедиях, в Большой и Малой). И даже в известном «Словаре русского языка» С. И. Ожегова (1973) мы читаем: «Оплодотворить. 1. Создать зародыш в ком-чем-н. слиянием мужской и женской половых клеток. 2. Послужить источником развития, совершенствования». Слияние одного из сиормпев с яйцеклеткой является, несомненно, оплодотворением, но тройное слияние по является, строго говоря, оплодотворением, так как 1) центральная клетка — не гамета и 2) в результате ¡»того слияния но образуется зигота, из которой и дальнейшем развивался бы новый организм. Очевидно, тройное слияние является оплодотворением лишь в указанном Ожеговым, втором, переносном смысле. Другими словами, в выражении «двойное оплодотворение» термин «оплодотворение» применяется в двух разных, смыслах — прямом и переносном. Тем не менее выражение «двойное онлодотиороние» настолько широко вошло в литературу, что было бы нецелесообразно его заменить (а попытки такого рода делались, в том числе известным, немецким ботаником. Достаточно, если мы будем помнить, что речь идет здесь о двух разных биологических процессах, условно объединяемых общим .названием.[ ...]
Редуцированный партеногенез отличается полной стерильностью образующихся растений и не передается из поколения в поколение, т. е. является нерегулярным и ненаследственным. Редуцированный партеногенез тесно связан с псевдогамией, поскольку партеногенетическое развитие зародыша из яйцеклетки всегда происходит после опыления своей или чужой пыльцой без слияния гамет. Растения, имеющие свойственное гаметам уменьшенное вдвое число хромосом, получены многими исследователями. В частности, гаплоидные растения были выращены из пыльцевых зерен при культуре на искусственной питательной среде у 20 видов покрытосеменных растений. Редуцированный партеногенез, вызываемый действием низкой температуры на растение, впервые был описан у дурмана (Datura stramonium). За последнее время в роде Datura разными способами было получено более 200 гаплоидных растений, главным образом при межродовых и межвидовых скрещиваниях, а также при действии во время процесса оплодотворения низкой или высокой температуры, Х-лучей, опылении пыльцой, облученной рентгеновскими лучами.[ ...]
Размножение представляет собой контролируемое воспроизведение растений. Оно преследует две основные цели: увеличение числа растений и сохранение у них ценных признаков и свойств. Имеются два существенно различающихся между собой способа размножения — половой и бесполый. Половое размножение обеспечивает увеличение числа растений путем посева семян, образовавшихся в результате слияния гамет; при бесполом размножении увеличение числа растений происходит за счет простого деления клеток и их дифференциации. Существенной особенностью бесполого размножения является то, что растения способны регенерировать недостающие части. Так, у стеблевого черенка образуются корни, у корневого — развиваются стеблевые почки, а листовой черенок образует и корни и побеги.[ ...]
При изоляции колосьев М 2 пергаментными колпачками озер-ненность их значительно ниже, чем при обычном свободном цветении, и в среднем составляет 21,8% с варьированием от 0 до 32% от числа нормально развитых цветков (табл. 28). Такая широкая амплитуда колебания по озерненности изолированных колосьев объясняется, с одной стороны, генетическими факторами, а с другой — различием температуры и влажности среды во время; формирования гамет и цветения. Попытки повысить плодовитость М 2 путем систематических отборов растений с применением изоляции колосьев не дали желаемых результатов.[ ...]
Экспериментально получепные пшенично-пырейные амфидип-лоиды представляют интерес только для дальнейшего скрещивания их с тем или другим видом пшеницы в зависимости от задач, стоящих перед селекционером. Но необходимо отметить, что при скрещивании пшеницы с амфидиплоидами дальнейшее формообразование характеризуется теми же закономерностями, какие наблюдаются при беккроссе амфигаплоидов с пшеницей. Объясняется это тем, что у амфигаплоидов жизнеспособными гаметами являются те, которые имеют реституционное ядро с нередуцированным числом хромосом. Таким образом, эти гаметы по хромосомному составу такие же, какие формируются у амфидиплоидов соответствующих гибридных комбинаций. Поэтому у пшенично-пырейных гибридов полпплоидизация не имеет того значения, как у большинства межродовых совершенно стерильных гибридов.[ ...]
Гаплоидный набор у дурмана состоит из 12 хромосом. На основе этого Блексли выделили 12 первичных мутантов, отличающихся между собой как по одной лишней хромосоме, так и по некоторым морфологическим признакам, вполне согласующимся с индивидуальными особенностями каждой из 12 хромосом.[ ...]
Многие растения являются естественными полиплоидами. Для избежания путаницы в этом отношении основное число хромосом в наборе принято обозначать через х. Так, основное число хромосом в роде Fragaria, к которому относится земляника, равно 7 (х = 7). Число хромосом в вегетативных клетках культурной земляники составляет 56 (8 наборов по 7 хромосом в каждом), или 2п=56=8х. Число хромосом в гаметах называется гаплоидным, или тг-числом, составляет п 28=4г. Под п понимается половинное число, под х — основное число хромосом.[ ...]
Антеридиальная клетка у сосновых, араукариевых и подокарповых или же вся клетка микроспоры у остальных хвойных делится на маленькую генеративную клетку и более крупную клетку трубки. Дальнейшая судьба этих двух клеток очень различна. Клетка трубки дальше не делится, но зато она дает начало пыльцевой трубке. Генеративная клетка рано или поздно делится на две: стерильную клетку, или клетку-ножку (иногда ее называют дис-локатором), и сперматогенную клетку (называемую часто клеткой-телом). Сперматогенная клетка дает начало двум мужским гаметам. Перед оплодотворением она набухает и тем самым способствует разрыву как ее самой, так и сперматогенной клетки. Таким образом, клетка-ножка способствует высвобождению мужских гамет из их материнской сперматогенной клетки. Вот почему знаменитый ботаник К. Гёбель (1933) назвал ее «дислокатором» — она дислоцирует (перемещает) сперматогенную клетку и сами мужские гаметы.[ ...]
Голосеменные отличаются от папоротников также развитием мужского гаметофита, строением и способом прорастания микроспор. У папоротников, где развитие гаметофита происходит обычно лишь после высеивания спор, прорастание спор происходит через так называемый тетрадный рубец, расположенный на проксимальном полюсе споры. У голосеменных, где мужской гаметофит сильно упрощается и его развитие ускоряется, первые деления ядра микроспоры происходят уже внутри микроспорангия. В связи с ранним развитием мужского гаметофита и образованием гамет еще внутри оболочки споры возникает необходимость в приспособлении, посредством которого микроспора может изменять свой объем. Таким приспособлением оказывается борозда на дистальном полюсе микроспоры, впервые возникающая у некоторых семенных папоротников и характерная для огромного большинства голосеменных. Борозда служит не только для регулирования объема пыльцевого зерна. Она становится местом выхода из микроспоры гаустории (у низших групп) или пыльцевой трубки (у гнетовых и хвойных), также являющихся новообразованиями. Таким образом, у голосеменных, в отличие от папоротников, отверстие для выхода содержимого микроспоры образуется на дистальном полюсе. Гаустория (присоска) типа саговниковых растет горизонтально и служит лишь для прикрепления и питания мужского гаметофита; настоящая пыльцевая трубка хвойных и гнетовых растет вертикально и служит главным образом для проведения спермиев к яйцеклеткам, т. е. является проводником (вектором), а не только присоской. Хотя обычно оба эти образования называют пыльцевыми трубками, но морфологически и функционально они очень различны.[ ...]
ru-ecology.info
Растения жизненный цикл и образование гамет
У человека и высших животных в результате мейоза образуются гаметы— яйцеклетка и сперматозоиды. При их слиянии возникает снова диплоидное ядро, из которого путем последовательных митозов развивается взрослый организм. Стадия мейоза характерна для жизненного цикла всех эукариот, однако отнюдь не всегда этот процесс протекает в период, аналогичный соответствующему моменту жизненного цикла человека. Так, клетки многих простейших и грибов обычно гаплоидны. После слияния двух гаплоидных ядер с образованием диплоидной клетки быстро наступает мейотическое деление, в результате которого вновь возникают гаплоидные особи. Чередование гаплоидных и диплоидных фаз жизненного цикла часто встречается у низших растений и примитивных животных. Например, гаметы папоротника падают на почву и [c.42] У животных мейоз происходит непосредственно перед образованием гамет. Иными словами, в клетках каждой особи данного вида на протяжении большей части жизненного цикла содержится 2N хромосом. Такие организмы называют диплоидными. Однако это отнюдь не всеобщее правило у эукариот, размножающихся половым путем. У многих протистов мейоз происходит сразу после образования зиготы, так что эти организмы на протяжении большей части жизненного цикла содержат N хромосом. Такие организмы называют гаплоидными. У многих водорослей и растений, а также у некоторых грибов и простейших происходит чередование гаплоидных и диплоидных поколений. При таком типе жизненного цикла из диплоидной зиготы возникает диплоидная особь, образующая путем мейоза гаплоидные клетки, предназначенные для бесполого размножения. Из каждой такой гаплоидной клетки возникает гаплоид- [c.50] Есть данные, подтверждающие, что у некоторых водорослей и грибов, равно как и у высших растений, в образовании половых органов, их созревании, освобождении гамет и, наконец, в привлечении сперматозоидов яйцеклеткой участвуют какие-то особые вещества, вырабатываемые организмом в определенной фазе его жизненного цикла. Эти данные говорят в пользу кон- [c.100]Внимательно изучите рис. 2.34, так как на нем в обобщенном виде представлен жизненный цикл всех наземных растений, в том числе и наиболее высокоорганизованных цветковых растений. Никогда не забывайте, что гаметы у растений образуются не в результате мейоза, как у животных, а в результате митоза мейоти-ческое деление происходит при образовании спор. [c.57]
Перед растениями, вышедшими на сушу, встает задача обеспечения встречи мужской и женской гамет, произведенных желательно разными особями. Эта задача решалась по-разному. Отметим поэтому лишь конвергенцию, связанную с одним из возможных решений. Хорошо известно, что цветки большинства покрытосеменных опыляются насекомыми. Исходно насекомых привлекала сама пыльца, а затем растения научились выделять особую сладкую жидкость — нектар. Для улавливания переносимой насекомыми пыльцы образовался специальный орган—пестик (точнее его рыльце) или несколько стило-диев. Оказывается аналогичные образования возникли и у некоторых ржавчинных грибов (дальнейшее относится к виду Пукциния граминис). В жизненном цикле этого гриба чередуются две формы. Одиа из них, назовем ее первой, диплоидна (дикарионна) и живет на хлебных злаках. Другая, вторая форма, гаплоидна и живет иа листьях барбариса. Вторая форма развивается из гаплоидных спор, образуемых первой формой в результате редукционного деления. Первая форма в свою очередь развивается из диплоидных спор, возникающих в ходе жизни второй формы. Для формирования диплоидных спор у второй формы необходимо оплодотворение. Оно может происходить при встрече мицелиев разных полов (точнее разных знаков —полы здесь ие различима морфологически). Такая встреча возможна, если мицелин разных знаков вы- [c.238]
У животных мейоз проходит при образовании гамет и является составной частью овогенеза и сперматогенеза, а у цветковых растений — во время микро- и мегаспорогенеза, т. е. до образования гамет. У растений продуктом мейоза являются гаплоидные микро- и мегаспоры, а гаметы образуются позднее, после двух митотических делений гаплоидных микроспор или трех митотических делений гаплоидных мегаспор. Кроме того, у растений мейоз связан с процессом чередования поколений — спорофита и гаметофита — в одном жизненном цикле. [c.190]
chem21.info