Моховидные как особая линия эволюции высших растений. Почему моховидные рассматривают как отдельную ветвь эволюции растений
Моховидные как особая линия эволюции высших растений
Моховидные (Bryophyta) часто, даже в кругах специалистов, называют популярным и коротким именем — мхи. Однако в более точном, научном смысле собственно мхами называют представителей лишь одной, наиболее обширной группы отдела моховидных, а именно — листостебелъные, или настоящие мхи (Вгуopsida). К ним относятся, например, всем известный сфагновый мох и кукушкин лен.
Среди высших растений моховидные образуют наиболее обособленную группу. Не случайно и среди ботаников выделился особый профиль исследователей-бриологов и соответственно оформилась особая, посвященная изучению моховидных ботаническая наука — бриология.
Древность моховидных вполне убедительно подтверждается ископаемыми находками. Во всяком случае, в карбоне они уже определенно существовали. Можно предположить, что именно в палеозое наметились такие основные линии развития моховидных, как печеночники, сфагновые и бриевые мхи. Геологическая летопись содержит слишком мало сведений для восстановления картины их становления, выяснения филогенетического родства отдельных групп и путей их эволюции. Несомненно, однако, что у возникших в начале палеозоя моховидных современные их семейства, а возможно, и роды в основном сложились уже в середине мезозоя. Третичное время характеризуется появлением и расселением высокоспециализированных родов, уже очень близких или даже тождественных современным. В четвертичное время продолжалось расселение моховидных и трансформация их ареалов.
Моховидные представляют собой слепую ветвь развития растений. С давних пор они прочно заняли свое особое место в экономике природы и сохранили его в сложных условиях формирования континентов, изменений климата и растительного покрова. Они выдержали эти испытания в течение геологического времени, не оскудев в морфологическом разнообразии и таксономическом богатстве и достигнув весьма широкого распространения. Массовое участие моховидных в растительном покрове Земли оказывает существенное воздействие на среду обитания других растений и животных.
В местах повышенного увлажнения в умеренных зонах северного и южного полушарий постепенно накапливались и продолжают накапливаться значительные толщи торфяных отложений с преобладающим участием моховидных.
Общеизвестно ландшафтное значение моховидных в тундровой зоне, особенно в моховых тундрах. Но изобилие этих растений свойственно и областям тропическим. Здесь, высоко в горах, обычно на высоте более 3000 м над уровнем моря, расположен особый пояс высокогорных лесов, который по праву называют поясом мшистых лесов. Известный русский ботаник и путешественник А. Н. Краснов очень образно описывает это «царство археофитов», т. е. древних растений, так: «Вы не видите ни коры, ни стеблей, все скрыто в нежном зеленом футляре, придающем в туманном полумраке леса стволам чудовищную толщину и неясные неопределенные контуры. Тонкие веточки, как инеем в морозное утро ветви наших лесов Севера, обрастают футляром зеленого мха, а этот же зеленый иней превращает в толстые канаты тонкие стебельки лиан черешки папоротниковых вай и стебельки соцветий».
Моховидные в подавляющем большинстве многолетние и только сравнительно редко однолетние растения. Но независимо от продолжительности жизни все они низкорослы. Размеры моховидных, если не учитывать нижней отмирающей части тела гаметофита, колеблются от миллиметра до нескольких сантиметров у большинства видов. Значительно реже встречаются более крупные моховидные. Так, живущие в воде мхи рода фонтиналис (Fontinalis) могут иметь стебли длиной до 60 см и более. Такими же и более длинными являются стебли некоторых мхов-эпифитов, поселяющихся на стволах и ветвях деревьев.
Моховидные отличаются сравнительно простой внутренней организацией. В их теле мож но обнаружить ассимиляционную ткань, а также слабо по сравнению с другими современными высшими растениями выраженные проводящие, механические, запасающие и покровные ткани, которые к тому же, как будет показано, отчетливо прослеживаются отнюдь не во всех систематических группах этого отдела.
При наличии сложно устроенных органов размножения у моховидных, по существу, отсутствуют специализированные вегетативные органы. Только условно можно говорить о листьях и стеблях моховидных, которые следовало бы называть соответственно филлидиями и каулидиями (чисто бриологические термины).
Структурные возможности гаплоидного гаметофита — доминирующего поколения в жизненном цикле моховидных — вообще более узки, чем диплоидного спорофита других высших растений. Этим, очевидно, и объясняется отсутствие у моховидных развитой водопроводящей системы и, как следствие этого, их ограниченный рост в высоту.
Своеобразие моховидных, как самостоятельного отдела высших растений, особенно заметно проявляется в цикле их развития.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
dic.academic.ru
Моховидные как особая линия эволюции высших растений
Моховидные (Bryophyta) часто, даже в кругах специалистов, называют популярным и коротким именем — мхи. Однако в более точном, научном смысле собственно мхами называют представителей лишь одной, наиболее обширной группы отдела моховидных, а именно — листостебелъные, или настоящие мхи (Вгуopsida). К ним относятся, например, всем известный сфагновый мох и кукушкин лен.
Среди высших растений моховидные образуют наиболее обособленную группу. Не случайно и среди ботаников выделился особый профиль исследователей-бриологов и соответственно оформилась особая, посвященная изучению моховидных ботаническая наука — бриология.
Древность моховидных вполне убедительно подтверждается ископаемыми находками. Во всяком случае, в карбоне они уже определенно существовали. Можно предположить, что именно в палеозое наметились такие основные линии развития моховидных, как печеночники, сфагновые и бриевые мхи. Геологическая летопись содержит слишком мало сведений для восстановления картины их становления, выяснения филогенетического родства отдельных групп и путей их эволюции. Несомненно, однако, что у возникших в начале палеозоя моховидных современные их семейства, а возможно, и роды в основном сложились уже в середине мезозоя. Третичное время характеризуется появлением и расселением высокоспециализированных родов, уже очень близких или даже тождественных современным. В четвертичное время продолжалось расселение моховидных и трансформация их ареалов.
Моховидные представляют собой слепую ветвь развития растений. С давних пор они прочно заняли свое особое место в экономике природы и сохранили его в сложных условиях формирования континентов, изменений климата и растительного покрова. Они выдержали эти испытания в течение геологического времени, не оскудев в морфологическом разнообразии и таксономическом богатстве и достигнув весьма широкого распространения. Массовое участие моховидных в растительном покрове Земли оказывает существенное воздействие на среду обитания других растений и животных.
В местах повышенного увлажнения в умеренных зонах северного и южного полушарий постепенно накапливались и продолжают накапливаться значительные толщи торфяных отложений с преобладающим участием моховидных.
Общеизвестно ландшафтное значение моховидных в тундровой зоне, особенно в моховых тундрах. Но изобилие этих растений свойственно и областям тропическим. Здесь, высоко в горах, обычно на высоте более 3000 м над уровнем моря, расположен особый пояс высокогорных лесов, который по праву называют поясом мшистых лесов. Известный русский ботаник и путешественник А. Н. Краснов очень образно описывает это «царство археофитов», т. е. древних растений, так: «Вы не видите ни коры, ни стеблей, все скрыто в нежном зеленом футляре, придающем в туманном полумраке леса стволам чудовищную толщину и неясные неопределенные контуры. Тонкие веточки, как инеем в морозное утро ветви наших лесов Севера, обрастают футляром зеленого мха, а этот же зеленый иней превращает в толстые канаты тонкие стебельки лиан черешки папоротниковых вай и стебельки соцветий».
Моховидные в подавляющем большинстве многолетние и только сравнительно редко однолетние растения. Но независимо от продолжительности жизни все они низкорослы. Размеры моховидных, если не учитывать нижней отмирающей части тела гаметофита, колеблются от миллиметра до нескольких сантиметров у большинства видов. Значительно реже встречаются более крупные моховидные. Так, живущие в воде мхи рода фонтиналис (Fontinalis) могут иметь стебли длиной до 60 см и более. Такими же и более длинными являются стебли некоторых мхов-эпифитов, поселяющихся на стволах и ветвях деревьев.
Моховидные отличаются сравнительно простой внутренней организацией. В их теле мож но обнаружить ассимиляционную ткань, а также слабо по сравнению с другими современными высшими растениями выраженные проводящие, механические, запасающие и покровные ткани, которые к тому же, как будет показано, отчетливо прослеживаются отнюдь не во всех систематических группах этого отдела.
При наличии сложно устроенных органов размножения у моховидных, по существу, отсутствуют специализированные вегетативные органы. Только условно можно говорить о листьях и стеблях моховидных, которые следовало бы называть соответственно филлидиями и каулидиями (чисто бриологические термины).
Структурные возможности гаплоидного гаметофита — доминирующего поколения в жизненном цикле моховидных — вообще более узки, чем диплоидного спорофита других высших растений. Этим, очевидно, и объясняется отсутствие у моховидных развитой водопроводящей системы и, как следствие этого, их ограниченный рост в высоту.
Своеобразие моховидных, как самостоятельного отдела высших растений, особенно заметно проявляется в цикле их развития.
Поделитесь на страничкеslovar.wikireading.ru
моховидные как особая линия эволюции высших растений
Моховидные (Bryophyta) часто, даже в кругах специалистов, называют популярным и коротким именем — мхи. Однако в более точном, научном смысле собственно мхами называют представителей лишь одной, наиболее обширной группы отдела моховидных, а именно — листостебелъные, или настоящие мхи (Вгуopsida). К ним относятся, например, всем известный сфагновый мох и кукушкин лен.
Среди высших растений моховидные образуют наиболее обособленную группу. Не случайно и среди ботаников выделился особый профиль исследователей-бриологов и соответственно оформилась особая, посвященная изучению моховидных ботаническая наука — бриология.
Древность моховидных вполне убедительно подтверждается ископаемыми находками. Во всяком случае, в карбоне они уже определенно существовали. Можно предположить, что именно в палеозое наметились такие основные линии развития моховидных, как печеночники, сфагновые и бриевые мхи. Геологическая летопись содержит слишком мало сведений для восстановления картины их становления, выяснения филогенетического родства отдельных групп и путей их эволюции. Несомненно, однако, что у возникших в начале палеозоя моховидных современные их семейства, а возможно, и роды в основном сложились уже в середине мезозоя. Третичное время характеризуется появлением и расселением высокоспециализированных родов, уже очень близких или даже тождественных современным. В четвертичное время продолжалось расселение моховидных и трансформация их ареалов.
Моховидные представляют собой слепую ветвь развития растений. С давних пор они прочно заняли свое особое место в экономике природы и сохранили его в сложных условиях формирования континентов, изменений климата и растительного покрова. Они выдержали эти испытания в течение геологического времени, не оскудев в морфологическом разнообразии и таксономическом богатстве и достигнув весьма широкого распространения. Массовое участие моховидных в растительном покрове Земли оказывает существенное воздействие на среду обитания других растений и животных.
В местах повышенного увлажнения в умеренных зонах северного и южного полушарий постепенно накапливались и продолжают накапливаться значительные толщи торфяных отложений с преобладающим участием моховидных.
Общеизвестно ландшафтное значение моховидных в тундровой зоне, особенно в моховых тундрах. Но изобилие этих растений свойственно и областям тропическим. Здесь, высоко в горах, обычно на высоте более 3000 м над уровнем моря, расположен особый пояс высокогорных лесов, который по праву называют поясом мшистых лесов. Известный русский ботаник и путешественник А. Н. Краснов очень образно описывает это «царство археофитов», т. е. древних растений, так: «Вы не видите ни коры, ни стеблей, все скрыто в нежном зеленом футляре, придающем в туманном полумраке леса стволам чудовищную толщину и неясные неопределенные контуры. Тонкие веточки, как инеем в морозное утро ветви наших лесов Севера, обрастают футляром зеленого мха, а этот же зеленый иней превращает в толстые канаты тонкие стебельки лиан черешки папоротниковых вай и стебельки соцветий».
Моховидные в подавляющем большинстве многолетние и только сравнительно редко однолетние растения. Но независимо от продолжительности жизни все они низкорослы. Размеры моховидных, если не учитывать нижней отмирающей части тела гаметофита, колеблются от миллиметра до нескольких сантиметров у большинства видов. Значительно реже встречаются более крупные моховидные. Так, живущие в воде мхи рода фонтиналис (Fontinalis) могут иметь стебли длиной до 60 см и более. Такими же и более длинными являются стебли некоторых мхов-эпифитов, поселяющихся на стволах и ветвях деревьев.
Моховидные отличаются сравнительно простой внутренней организацией. В их теле мож но обнаружить ассимиляционную ткань, а также слабо по сравнению с другими современными высшими растениями выраженные проводящие, механические, запасающие и покровные ткани, которые к тому же, как будет показано, отчетливо прослеживаются отнюдь не во всех систематических группах этого отдела.
При наличии сложно устроенных органов размножения у моховидных, по существу, отсутствуют специализированные вегетативные органы. Только условно можно говорить о листьях и стеблях моховидных, которые следовало бы называть соответственно филлидиями и каулидиями (чисто бриологические термины).
Структурные возможности гаплоидного гаметофита — доминирующего поколения в жизненном цикле моховидных — вообще более узки, чем диплоидного спорофита других высших растений. Этим, очевидно, и объясняется отсутствие у моховидных развитой водопроводящей системы и, как следствие этого, их ограниченный рост в высоту.
Своеобразие моховидных, как самостоятельного отдела высших растений, особенно заметно проявляется в цикле их развития.
Источник: Биологическая энциклопедия на Gufo.megufo.me
биология
Моховидные / Bryophyta
| Общее число видов моховидных около 35 тыс. Строение моховидных растений: В жизненном цикле моховидных, как и других высших растений, имеется чередование двух фаз: спорофита и гаметофита. Однако доминирует (преобладает) гаметофит, в то время как у всех других высших растений доминирует спорофит. Именно поэтому моховидные рассматривают как самостоятельную боковую ветвь в эволюции высших растений. |
Гаметофит обычно многолетний. Он представляет собой листовидный таллом или растение в виде побега, расчлененного на стебель и листья. Корней нет, их функцию выполняют ризоиды - выросты поверхностных клеток тела. Органы полового размножения многоклеточные. Спорофит, называемый у моховидных спорогоном,играет подчиненную роль. Он представляет собой цилиндрическую ножку, заканчивающуюся шаровидной, эллиптической или цилиндрической коробочкой, внутри которой образуется спорангий со спорами. Спорогон паразитирует на гаметофите, так как получает от него воду и необходимую пищу. Максимальная длина тела (гаметофита и спорогона) 60 см.
Моховидные по своей организации и экологии еще близки к водорослям. Как и у водорослей, у них нет сосудов и корней. Некоторые примитивные представители имеют вегетативное тело в виде стелющегося таллома с верхушечным (дихотомическим) ветвлением, похожего на таллом водорослей. Оплодотворение связано с водой. Среди моховидных, как и среди водорослей, нет одревесневающих форм.
Распространение. Моховидные распространены на всех континентах мира, но неравномерно. В тропических странах - преимущественно в горах. Незначительное число видов растет в засушливых условиях, например в степях. Некоторые виды ведут эпифитный образ жизни на корке деревьев или водный. Основное же разнообразие видов сосредоточено во влажных местах Северного полушария, в областях с умеренным и холодным климатом. В сложении растительного покрова, особенно тундр, болот и лесов, им принадлежит важная роль.
Классификация. Моховидные подразделяют на три класса: Антоцеротовые, Печеночники, Листостебельные мхи. Наибольшее значение имеют два последних класса.
Класс Печеночники - Hepaticopsida.
Общее число видов около 10 тыс. Распространелы повсеместно. Примитивность строения тела печеночников свидетельствует об их древности.
Маршанция обыкновенная (Marchantia polymorpha) - типичный представитель класса. Гаметофит в виде пластинчатого таллома, длиной 10 - 12 см, ветвление верхушечное. С обеих сторон он покрыт эпидермой. Верхняя эпидерма имеет вентиляционные отверстия - устьица. Они окружены специальными клетками, расположенными в четыре ряда. Под устьицами имеются воздушные камеры. Нижняя эпидерма дает выросты - одноклеточные ризоиды и красноватые или зеленоватые чешуйки, которые иногда принимают за редуцированные листья. Под верхней эпидермой расположена ассимиляционная ткань, состоящая из вертикальных столбиков паренхимных клеток с хлоропластами. Ниже находится слой тонкостенных бесхлорофилльных паренхимных клеток. Следовательно, таллом маршанции имеет дорсивентральное строение.
На верхней стороне таллома образуются особые веточки - подставки, а на них - органы полового размножения. Маршанция - двудомное растение. На одних экземплярах подставки имеют форму сидящей на ножке девятилучевои звездочки, между лучами которой на нижней стороне расположены архегонии. На других - подставки имеют форму сидящего на ножке восьмиугольного щитка, на верхней стороне которого расположены антеридии, погруженные в антеридиальные полости. В брюшке архегония образуется яйцеклетка. После ее слияния со сперматозоидом из зиготы образуется спорогон. Он представляет собой коробочку на короткой ножке, которая прикреплена к гаметофиту гаусторией. Внутри коробочки из спорогенных клеток в результате мейоза образуются гаплоидные споры, а также элатеры - мертвые удлиненные клетки со спирально утолщенной стенкой, служащие для разрыхления массы спор, а также для выбрасывания их из коробочки. В благоприятных условиях из споры развивается предросток, или протонема. Это небольшая нить. Из ее верхушечной клетки вырастает таллом маршанции.
Вегетативное размножение осуществляется выводковыми тельцами линзовидной формы, имеющими зеленый цвет. Они образуются на верхней стороне таллома в особых корзиночках в результате деления клеток, выстилающих их дно.
.Виды маршанции имеют широкое распространение. Наиболее часто их можно встретить во влажных местах: на берегах озер и рек, по оврагам и в травянистом покрове под пологом леса.
Класс Листостебельные мхи - Bryopsida.
Общее число видов около 25 тыс. Многие виды распространены в приполярных странах Северного полушария. На огромных территориях в тундре, на болотах, в лесах они доминируют в растительном покрове, значительно влияя на обеспеченность сушивлагой.
Гаметофит представляет собой прямостоячую стеблевидную ось - каулидий, покрытую листовидными выростами - филлидиями. Условно их можно называть стебель и листья. На нижней части стебля образуются многоклеточные ризоиды (не у всех). Ветвление боковое. Нарастание осей происходит в результате деления пирамидальной верхушечной клетки. Оно может быть моноподиальным или симподиальным. В соответствии с этим органы полового размножения и спорогон размещаются на вершине гаметофита или на боковых разветвлениях.
Класс подразделяют на три подкласса: Андреевые мхи, Сфагновые мхи, Бриевые (Зеленые) мхи. Наибольшее значение имеют два последних подкласса.
Подкласс Сфагновые мхи - Sphaqnidae.
Сфагновые мхи имеют довольно однообразное строение и поэтому трудны для определения. Гаметофит их представляет собой сильноветвящееся, особенно в верхней части, растение. Разветвления густо покрыты листьями. Сфагновые мхи живут в очень влажной среде. В связи с этим у них нет ризоидов и влага поступает непосредственно в стебель, который при основании с течением времени отмирает. Строение стебля несложное. В центре его находится сердцевина из тонкостенных паренхимных клеток, выполняющих проводящую и запасающую функции. Она окружена корой, состоящей из двух слоев: склеродермы, выполняющей механическую функцию, и гиалодермы, выполняющей водозапасающую функцию. Клетки гиалодермы крупные, мертвые, стенки их имеют круглые отверстия, через которые полости смежных клеток сообщаются друг с другом, а также с внешней средой. Иногда эти клетки несут спиральные утолщения. Лист состоит из одного ряда клеток, резко различающихся как по структуре, так и по выполняемой функции. Одни из них живые, хлорофиллоносные, другие - мертвые, относительно более крупные, со спирально утолщенными стенками, пронизанные отверстиями, по строению сходные с водозапасающими клетками гиалодермы, их называют гиалиновыми. Гиалиновые клетки способны накапливать и долго удерживать огромное количество воды, в 30 - 40 раз превышающее массу самого растения.
Гаметофиты - однодомные и двудомные. Антеридии образуются в пазухах листьев на разветвлениях стебля. Около них листья окрашены в красноватый цвет. Архегонии на укороченных разветвлениях. В результате слияния сперматозоида с яйцеклеткой возникает зигота, которая представляет собой начало диплоидной фазы - спорогона. Спорогон состоит из ножки и коробочки. Ножка сильно укорочена, луковицеобразная, но ко времени созревания спор верхушка стебля гаметофита сильно вырастает и выносит коробочку кверху (ложная ножка). В центре коробочки помещается округлая колонка, над которой размещен в виде свода спорангий со спорогенной тканью. Стенка коробочки прочная, многослойная. Наружный хлорофиллоносный слой содержит большое число недоразвитых устьиц. Коробочка имеет крышечку, которая во время созревания спор отскакивает, и споры рассеиваются. Элатер нет. Из спор образуется сначала зеленая пластинчатая протонема, а затем из почек, расположенных на ней, - взрослый гаметофит, который и доминирует в жизненном цикле.
Структура сфагновых отличается примитивностью: пластинчатая протонема, отсутствие проводящего пучка и ризоидов, слабая дифференциация коробочки.
Значение сфагновых в природе очень велико. Накапливая огромное количество воды и разрастаясь плотными дернинами, они вызывают заболачивание огромных пространств, достигающих зоны тундры. Для осушения их осуществляют агромелиоративные работы. С другой стороны, старые болота имеют важное хозяйственное значение для разработки залежей торфа. Нарастание пласта торфа в наиболее благоприятных условиях происходит медленно - слой толщиной 1 см образуется примерно за 10 лет.
Подкласс Бриевые (зеленые) мхи - Bryidae.
Число видов 24,6 тыс. Распространены более широко, чем сфагновые мхи. Живут в разнообразных экологических условиях от тундры и лесотундры до степей и пустынь. Наиболее типичные местообитания бриевых мхов, где они доминируют или формируют сплошной покров, - тундра, болота и некоторые типы лесов. Каждому местообитанию свойственны свои виды. Бриевые мхи по сравнению со сфагновыми отличаются большим разнообразием строения. Органы полового размножения закладываются у одних видов на главной оси, у других - на боковых. У некоторых видов ветвление не выражено.
Политрих обыкновенный, кукушкин лен (Polytrichum commune) - это один из распространенных представителей бриевых мхов. Он растет в лесу, на полянах, окраинах болот.
Стебель гаметофита прямостоячий, неветвистый, высотой 15 см и более, густо покрыт листьями. Подземная часть его простирается в почве почти горизонтально, на ней образуются ризоиды. В центре стебля находится концентрический проводящий пучок, состоящий из вытянутых клеток, сходных с трахеидами и ситовидными трубками. Он окружен паренхимой, также выполняющей проводящую функцию. С наружной стороны паренхима граничит со склеродермой (корой). Внешний слой ее, состоящий из бесцветных клеток, называют гиалодермой.
Листья расположены по спирали. Они состоят из линейной пластинки с заостренной зубчатой верхушкой и пленчатого влагалища. На верхней стороне листа расположены ассимиляционные пластинки. Жилка с механическими и проводящими элементами расширена.
Гаметофит двудомный. Архегонии бутылковидной формы расположены на верхушке женского гаметофита, антеридии мешковидной формы - на верхушке мужского. Между архегониями и антеридиями имеются стерильные нити - парафизы. После оплодотворения из зиготы образуется спорогон, состоящий из длинной ножки и коробочки. Коробочка прямостоячая или более или менее косо расположенная, призматическая, четырех-пятиграниая, покрыта ржаво-войлочным колпачком, образующимся из стенок архегония. Коробочка состоит из урночки и крышечки. Нижняя часть урночки сужена в шейку. На границе урночки и шейки в эпидерме имеются устьица. В центре урночки расположена колонка, которая у крышечки расширяется и формирует эпифрагму - тонкостенную перегородку, закрывающую урночку. Вокруг колонки расположен спорангий в виде цилиндрического мешка, прикрепленного к стенке и колонке особыми нитевидными образованиями. Урночка имеет специальное приспособление для рассеивания спор - перистом, представляющий собой ряд зубчиков с тупыми верхушками, расположенных по краю урночки. Между зубцами, способными к гигроскопическим движениям, и эпифрагмой имеются отверстия, через которые в сухую погоду высыпаются споры. Из споры вырастает протонема в виде зеленой ветвящейся нити. На ней формируются почки, из которых со временем развиваются взрослые гаметофиты.
Отделы водорослей
Отдел организмов, рассматриваемая здесь как водоросли, очень разнообразен и не представляет единого таксона. Эти организмы неоднородны по своему строению и происхождению.
Водоросли — автотрофные растения, в их клетках присутствуют различные модификации хлорофилла и другие пигменты, обеспечивающие фотосинтез. Водоросли обитают в пресной и морской воде, а также на суше, на поверхности и в толще почвы, на коре деревьев, камнях и других субстратах.
Водоросли относятся к 10 отделам из двух царств: 1) Сине-зелёные, 2) Красные, 3) Пирофитовые, 4) Золотистые, 5) Диатомовые, 6) Желто-зелёные, 7) Бурые, 8) Эвгленовые, 9) Зеленые и 10) Харовые. Первый отдел относится к царству Прокариот, остальные — к царству Растений.
Отдел Сине-зёлёные водоросли, или Цианобактерии (Cyanophyta)
Сине-зелёных водорослей насчитывается около 2 тысяч видов, объединяемых примерно в 150 родов. Это древнейшие организмы, следы существования которых найдены в докембрийских отложениях, возраст их около 3 млрд. лет.
Среди сине-зелёных водорослей есть одноклеточные формы, но большинство видов являются колониальными и нитчатыми организмами. Они отличаются от других водорослей тем, что в их клетках нет оформленного ядра. У них отсутствуют митохондрии, вакуоли с клеточным соком, нет оформленных пластид, а пигменты, с помощью которых осуществляется фотосинтез, находятся в фотосинтетических пластинах — ламеллах. Пигменты сине-зелёных водорослей очень разнообразны: хлорофилла, каротины, ксантофиллы, а также специфические пигменты из группы фикобилинов — синий фикоцианин и красный фикоэритрин, встречающиеся помимо цианобактерий только у красных водорослей. Окраска этих организмов чаще всего сине-зеленая. Однако в зависимости от количественного соотношения различных пигментов окраска этих водорослей может быть не только сине-зеленой, но также фиолетовой, красноватой, желтой, бледно-голубой или почти черной.
Сине-зелёные водоросли распространены по всему земному шару и встречаются в самых разнообразных условиях. Они способны существовать даже в крайних условиях обитания. Эти организмы выносят продолжительное затемнение и анаэробиоз, могут жить в пещерах, в разных почвах, в богатых сероводородом слоях природного ила, в.термальных водах и др.
Вокруг клеток колониальных и нитчатых водорослей образуются слизистые влагалища, которые служат защитной обверткой, предохраняющей клетки от высыхания и являющейся светофильтром.
Многие нитчатые сине-зелёные водоросли имеют своеобразные клетки — гетероцисты. У этих клеток хорошо выражена двуслойная оболочка, и выглядят они пустыми. Но это живые клетки, заполненные прозрачным содержимым. Сине-зелёные водоросли, имеющие гетероцисты, способны фиксировать атмосферный азот. Некоторые виды сине-зелёных водорослей являются компонентами лишайников. Они могут находиться в качестве симбионтов в тканях и органах высших растений. Их способность к фиксации атмосферного азота используется высшими растениями.
Массовое развитие сине-зелёных водорослей в водоемах может иметь отрицательные последствия. Повышенная температура и загрязнение вод органическими веществами вызывают так называемое «цветение воды». Это делает воду непригодной для употребления человеком. Некоторые пресноводные цианобактерии токсичны для человека и животных.
Размножение сине-зелёных водорослей очень примитивно. Одноклеточные и многие колониальные формы размножаются только делением клеток пополам. Большинство нитчатых форм размножаются гормогониями (это короткие участки, отделившиеся от материнской нити, вырастающие во взрослые особи). Размножение может осуществляться и с помощью спор — разросшихся толстостенных клеток, способных переживать неблагоприятные условия и затем прорастать в новые нити.
Отдел Красные водоросли (или Багрянки) (Rhodophyta)
Красные водоросли (Багрянки) — многочисленная (около 3800 видов из более чем 600 родов) группа в основном морских обитателей. Их размеры варьируются от микроскопических до 1-2 м. Внешне красные водоросли очень разнообразны: есть нитевидные, пластинчатые, кораллоподобные формы, в разной степени рассеченные и разветвленные.
Красные водоросли имеют своеобразный набор пигментов: кроме хлорофилла a и b имеется хлорофилл d, известный только для этой группы растений, есть каротины, ксантофиллы, а также пигменты из группы фикобилинов: синий пигмент — фикоцианин, красный — фикоэритрин. Различное сочетание этих пигментов определяет окраску водорослей — от ярко-красной до голубовато-зеленой и желтой.
Размножаются красные водоросли вегетативным, бесполым и половым путем. Вегетативное размножение характерно лишь для наиболее низко организованных багрянок (одноклеточные и колониальные формы). У высокоорганизованных многоклеточных форм оторванные участки таллома погибают. Для бесполого размножения служат различного рода споры.
Половой процесс оогамный. На растении-гаметофите образуются мужские и женские половые клетки (гаметы), лишенные жгутиков. При оплодотворении женские гаметы не выходят в окружающую среду, а остаются на растении; мужские гаметы выбрасываются наружу и пассивно переносятся токами воды.
Диплоидные растения — спорофиты — имеют такой же внешний вид, как и гаметофиты (гаплоидные растения). Это изоморфная смена поколений. На спорофитах формируются органы бесполого размножения.
Многие красные водоросли широко используются человеком, они съедобны и полезны. В пищевой и медицинской промышленности широко используется получаемый из разных видов багрянок (около 30) полисахарид агар.
Отдел Пирофитовые (или Динофитовые) водоросли (Pyrrophyta (Dinophyta))
Отдел насчитывает около 1200 видов из 120 родов, объединяющих эукариотические одноклеточные (в том числе — двужгутиковые), коккоидные и нитчатые формы. Группа совмещает в себе признаки растений и животных: у некоторых видов имеются щупальца, псевдоподии и стрекательные клетки; некоторые имеют характерный для животных тип питания, обеспечивающийся глоткой. У многих имеется стигма, или глазок. Клетки часто покрыты твердой оболочкой. Хроматофоры буроватых и красноватых оттенков, содержат хлорофиллы а и с, а также каротины, ксантофиллы (иногда — фикоцианин и фикоэритрин). В качестве запасных веществ откладывается крахмал, иногда — масло. Жгутиконосные клетки имеют четко выраженные спинную и брюшную стороны. На поверхности клетки и в глотке имеются бороздки.
Размножаются делением в подвижном или неподвижном состоянии (вегетативно), зооспорами и автоспорами. Половое размножение известно у немногих форм; оно проходит в виде слияния изогамет.
Пирофитовые водоросли — обычные обитатели загрязненных водоемов: прудов, отстойников, некоторых водохранилищ и озер. Многие образуют фитопланктон в морях. При неблагоприятных условиях образуют цисты с толстыми целлюлозными оболочками.
Наиболее широко распространен и богат видами род Криптомонада (Cryptomonas).
Отдел Золотистые водоросли (Chrysophyta)
Микроскопические или мелкие (до 2 см длины) организмы золотисто-желтого цвета, обитающие в соленых и пресных водоемах по всему земному шару. Имеются одноклеточные, колониальные и многоклеточные формы. В России известно около 300 видов из 70 родов. Хроматофоры обычно золотисто-желтые или бурые. Они содержат хлорофиллы а и с, а также каротиноиды и фукоксантин. В качестве запасных веществ откладывается хризоламинарин и масло. Некоторые виды гетеротрофны. Большинство форм имеют 1-2 жгутика и в силу этого подвижны. Размножаются преимущественно бесполым путем — делением или зооспорами; половой процесс известен лишь у нескольких видов. Встречаются обычно в чистых пресных водах (кислые воды сфагновых болот), реже — в морях и в почвах. Типичные элементы фитопланктона.
Отдел Диатомовые водоросли (Bacillariophyta (Diatomea))
Диатомовые водоросли (диатомеи) насчитывают около 10 тысяч видов, относящихся примерно к 300 родам. Это микроскопические организмы, обитающие главным образом в водоемах. Диатомовые водоросли — особая группа одноклеточных организмов, отличная от других водорослей. Клетки диатомеи покрыты панцирем из кремнезема. В клетке находятся вакуоли с клеточным соком. В центре расположено ядро. Хроматофоры крупные. Окраска их имеет различные оттенки желто-бурого цвета, так как среди пигментов преобладают каротины и ксантофиллы, имеющие желтые и бурые оттенки, и маскирующие хлорофиллы а и с.
Для панцирей диатомеи характерны геометрическая правильность строения и большое разнообразие очертаний. Панцирь состоит из двух половинок. Большая — эпитека — покрывает меньшую — гипотеку, как крышка покрывает коробку.
Большинство диатомей, имеющих двустороннюю симметрию, способны передвигаться по поверхности субстрата. Движение осуществляется с помощью так называемого шва. Шов представляет собой щель, прорезающую стенку створки. Движение цитоплазмы в щели и трение ее о субстрат обеспечивают перемещение клетки. Клетки диатомей, имеющих радиальную симметрию, не способны к передвижению.
Размножаются диатомеи обычно делением клетки на две половинки. Протопласт увеличивается в объеме, вследствие чего эпитека и гипотека расходятся. Протопласт делится на две равные части, митотически делится ядро. В каждой половинке разделившейся клетки панцирьиграет роль эпитеки и достраивает недостающую половинку панциря, всегда гипотеку. В результате многочисленных делений происходит постепенное уменьшение размеров клеток у части популяции. Некоторые клетки примерно в три раза меньше по сравнению с первоначальными. Достигнув минимальных размеров, клетки развивают ауксоспоры («растущие споры»). Образование ауксоспор связано с половым процессом.
Клетки диатомовых водорослей в вегетативном состоянии диплоидны. Перед половым размножением происходит редукционное деление ядра (мейоз). Две клетки диатомей сближаются, створки раздвигаются, гаплоидные (после мейоза) ядра попарно сливаются, и образуются одна или две ауксоспоры. Ауксоспора некоторое время растет, а затем вырабатывает панцирь и превращается в вегетативную особь.
Среди диатомей есть светолюбивые и тенелюбивые виды, они живут в водоемах на разных глубинах. Диатомовые могут обитать и в почвах, особенно влажных и заболоченных. Наряду с другими водорослями диатомеи могут вызывать «цветение» снега.
Диатомеи играют большую роль в экономике природы. Они служат постоянной кормовой базой и первоначальным звеном в пищевых цепях для многих водных организмов. Ими питаются многие рыбы, особенно молодь.
Панцири диатомей, осаждаясь на дно в течение миллионов лет, формируют осадочную геологическую породу — диатомит. Он широко используется как строительный материал с высокими тепло- и звукоизоляционными свойствами, в качестве фильтров в пищевой, химической, медицинской промышленности.
Отдел желто-зеленые водоросли (Xanthophyta)
Эта группа водорослей насчитывает около 550 видов. В основном это обитатели пресных вод, реже встречаются в морях и на влажной почве. Среди них есть одноклеточные и многоклеточные формы, жгутиковые, коккоидные, нитчатые и пластинчатые, а также сифональные организмы. Для этих водорослей характерна желто-зеленая окраска, давшая название всей группе. Хлоропласты дисковидной формы. Характерные пигменты — хлорофиллы а и с, а и Ь каротиноиды, ксантофиллы. Запасные вещества — глюкан, жиры. Половое размножение — оогамное и изогамное. Вегетативно размножаются делением; бесполое размножение осуществляется специализированными подвижными или неподвижными клетками — зоо- и апланоспорами.
Отдел Бурые водоросли (Phaeophyta)
Бурые водоросли — высокоорганизованные многоклеточные организмы, обитающие в морях. Их около 1500 видов из примерно 250 родов. Самые крупные из бурых водорослей достигают нескольких десятков метров (до 60 м) длины. Однако в этой группе встречаются также виды микроскопических размеров. Форма слоевищ может быть очень многообразна.
Общий признак всех водорослей, принадлежащих к этой группе, — желтовато-бурая окраска. Она обусловлена пигментами каротином и ксантофиллом (фукоксантин и др.), которые маскируют зеленый цвет хлорофиллов а и с. Клеточная оболочка целлюлозная с наружным пектиновым слоем, способным к сильному ослизнению.
У бурых водорослей встречаются все формы размножения: вегетативное, бесполое и половое. Вегетативное размножение происходит отделившимися частями слоевища. Бесполое размножение осуществляется при помощи зооспор (подвижных благодаря жгутикам спор). Половой процесс у бурых водорослей представлен изогамией (реже — анизогамия и оогамия).
У многих бурых водорослей гаметофит и спорофит различаются по форме, размерам и строению. У бурых водорослей наблюдается чередование поколений, или смена ядерных фаз в цикле развития. Бурые водоросли встречаются во всех морях земного шара. В зарослях бурых водорослей близ берегов находят укрытие, места размножения и питания многочисленные прибрежные животные. Бурые водоросли широко используются человеком. Из них получают альгинаты (соли альгиновой кислоты), применяемые как стабилизаторы растворов и суспензий в пищевой промышленности. Они используются при изготовлении пластмасс, смазочных материалов и т. д. Некоторые бурые водоросли (ламинарии, аларии и др.) используются в пище.
Отдел Эвгленовые водоросли (Euglenophyta)
В этой группе около 900 видов из примерно 40 родов. Это одноклеточные жгутиковые организмы, в основном обитатели пресных вод. В хлоропластах присутствуют хлорофиллы а и b и большая группа вспомогательных пигментов из группы каротиноидов. У этих водорослей на свету происходит фотосинтез, а в темноте они переходят на гетеротрофное питание.
Размножение этих водорослей происходит только за счет митотического деления клетки. Митоз у них отличается от этого процесса в других группах организмов.
Отдел Зеленые водоросли (Chlorophyta)
Зеленые водоросли — самый крупный отдел Водорослей, насчитывающий, по разным оценкам, от 13 до 20 тысяч видов из примерно 400 родов. Для этих водорослей характерна чисто зеленая, как у высших растений, окраска, так как среди пигментов преобладает хлорофилл. В хлоропластах (хроматофорах) присутствуют две модификации хлорофилла a и b, как и у высших растений, а также другие пигменты — каротины и ксантофиллы.
Жесткие клеточные стенки зеленых водорослей образованы целлюлозой и пектиновыми веществами. Запасные вещества — крахмал, реже масло. Многие особенности строения и жизни зеленых водорослей свидетельствуют об их родстве с высшими растениями. Зеленые водоросли отличаются наибольшим по сравнению с другими отделами многообразием. Они могут быть одноклеточными, колониальными, многоклеточными. В этой группе представлено все разнообразие морфологической дифференциации тела, известной для водорослей, — монадная, коккоидная, пальмеллоидная, нитчатая, пластинчатая, неклеточная (сифональная). Велик диапазон их размеров — от микроскопических одиночных клеток до крупных многоклеточных форм длиной в десятки сантиметров. Размножение вегетативное, бесполое и половое. Встречаются все основные типы смены форм развития.
Зеленые водоросли обитают чаще в пресных водоемах, однако немало солоноводных и морских форм, а также вневодных наземных и почвенных видов.
studfiles.net
Происхождение и эволюция мхов (мохообразных) | Биология. Реферат, доклад, сообщение, краткое содержание, лекция, шпаргалка, конспект, ГДЗ, тест
Мохообразные — особая группа высших растений. Они отличаются преобладанием в цикле развития гаметофита — полового поколения. Спорофит — бесполое поколение — имеет незначительные размеры, кратковременное существование и всегда связан с гаметофитом, прикреплен к нему и получает от гаметофита питательные вещества, необходимые для развития спор. Он получил особое название — спорогоний.
Мохообразные — древняя группа растений. Ископаемые их остатки достоверно известны из отложений каменноугольного периода палеозойской эры. Отсюда известны как талломные печеночники, похожие на маршанциевые, так и лиственные мхи, внешне напоминающие Polytrichum. Эти формы имеют уже сложное строение, и прошло, вероятно, очень много времени, прежде чем они могли сформироваться. В последнее время споры мохообразных (наряду со спорами папоротникообразных) найдены в еще более древних отложениях — силурийских и девонских.
Поэтому время возникновения мохообразных мы должны отнести на еще более ранние периоды жизни Земли, чем каменноугольный период палеозойской эры. Палеонтология, к сожалению, нам не открывает таких древних форм растений. В вопросе о происхождении и эволюции мохообразных нам приходится руководствоваться данными, добытыми при изучении современных растений (морфологии, цитологии, географии и т. д.). Эти данные не являются бесспорными. Поэтому имеются различные взгляды на эволюцию мохообразных. Наиболее распространенными и обоснованными являются следующие взгляды, разделяемые большинством биологов.
Мохообразные, как и остальные высшие растения, произошли от каких-то древних водорослей — бурых или зеленых, вышедших на сушу. Эти водоросли имели, вероятно, стелющиеся по субстрату талломы очень простого строения. Они имели вид дихотомически ветвящихся дорзовентральных пластинок, обладали сменой поколений с равноправным развитием спорофита и гаметофита. Половые органы имели очень простое строение и лишь постепенно приняли форму антеридиев и архегониев.
У мохообразных, развившихся из таких простейших форм, совершенствовался гаметофит. Он был сначала талломным, затем выработалась листостебельная структура. Спорофит, наоборот, претерпевал редукцию, упрощение. Он потерял способность к самостоятельному существованию после того, как оплодотворенная яйцеклетка стала делиться и развиваться внутри архегония. Он, внедрившись в ткань таллома, оказался постоянно связанным с гаметофитом и постепенно перешел к паразитическому существованию за его счет. Функции спорофита сводились к скорейшему и массовому развитию спор. Во внешнем строении спорофит все время упрощался, но зато у него развивались сложные приспособления к рассеиванию спор (пружинки у печеночников, перистом у лиственных мхов). Все остальные высшие растения базировались на спорофите, гаметофит же у них претерпевал упрощение (редукцию).
Среди мохообразных, и в частности среди печеночников, наиболее примитивными являются сферокарповые, с их простыми талломами. Они произошли из каких-то групп еще более простых талломных печеночников, но имевших уже антеридии и архегонии и связанных в свою очередь с водорослями. Маршанциевые и юнгерманниевые могут быть выведены из сферокарповых. Последние, как мы видели выше, обладают признаками тех и других (простое анатомическое строение, как у юнгерманниевых, и сходное с маршанциевыми устройство антеридиев и спорогония).
Маршанциевые усложнили внутреннее строение (наличие воздушных камер, простые и язычковые ризоиды и т. д.) но остались талломными растениями. Юнгерманниевые развивались в направлении выработки сложного внешнего строения. Они постепенно из талломных форм выработали листостебельную структуру при простом анатомическом строении
Антоцеротовые стоят особняком. По строению гаметофита их можно вывести непосредственно из водорослей (хроматофоры с пиреноидами и др.), но по наличию уже типичных антеридиев и архегониев они могут быть связаны и с простейшими талломными печеночниками Они обнаруживают связи и с лиственными мхами (по устройству спорогония).
Лиственные мхи являются более организованными растениями и могут быть связаны в происхождении с олиственными юнгерманниями. Так, сфагновые мхи обнаруживают тесные взаимоотношения с печеночниками (пластинчатая протонема, однослойные листья у Sphagnum, строение антеридиев, развитие спорогония, отсутствие перистома и др.). По общему строению, радиальной симметрии сфагновые (строение антеридиев и архегониев, общий облик) связаны с зелеными мхами. Это говорит о родстве всех классов и порядков Bryophyta, о том, что они произошли из одного источника. Сфагновые (и андреевые) более примитивны, зеленые мхи с их сложным внутренним строением, наличием перистома и др. более сложны и совершенны. Материал с сайта http://worldofschool.ru
В середине XX века некоторые ботаники за рубежом и у нас (Б. М. Козополянский, А. Л. Тахтаджян) развивали иной, прямо противоположный взгляд на эволюцию мохообразных. Основываясь на сходстве спорангиев Horneophyton со спорангиями сфагновых и андреевых мхов, они считали Bryophyta редуцированной группой, происшедшей из псилофитовых. По их мнению, развитие мохообразных шло в направлении от листостебельных форм к талломным в связи с приспособлением их к жизни в местообитаниях с повышенным увлажнением. С этой точки зрения сложный спорофит типа кукушкина льна является более примитивным, наиболее же совершенными мохообразными являются талломные формы типа маршанциевых.
Этот взгляд принять нельзя. Помимо противоречий данным сравнительной морфологии, эта точка зрения не подтверждается и палеонтологическими материалами. Листостебельные печеночники, например, появляются только в третичное время, а талломные формы типа Marchantia известны уже из каменноугольных отложений палеозойской эры.
На этой странице материал по темам:Происхождение и эволюция мохообразных, разные взгляды.
Эволюция мхов
Эволюция мохообразных
worldofschool.ru
Моховидные Как Особая Линия Эволюции Высших Растений - Биологическая энциклопедия
Моховидные (Bryophyta) часто, даже в кругах специалистов, называют популярным и коротким именем - мхи. Однако в более точном, научном смысле собственно мхами называют представителей лишь одной, наиболее обширной группы отдела моховидных, а именно - листостебелъные, или настоящие мхи (Вгуopsida). К ним относятся, например, всем известный сфагновый мох и кукушкин лен.
Среди высших растений моховидные образуют наиболее обособленную группу. Не случайно и среди ботаников выделился особый профиль исследователей-бриологов и соответственно оформилась особая, посвященная изучению моховидных ботаническая наука - бриология.
Древность моховидных вполне убедительно подтверждается ископаемыми находками. Во всяком случае, в карбоне они уже определенно существовали. Можно предположить, что именно в палеозое наметились такие основные линии развития моховидных, как печеночники, сфагновые и бриевые мхи. Геологическая летопись содержит слишком мало сведений для восстановления картины их становления, выяснения филогенетического родства отдельных групп и путей их эволюции. Несомненно, однако, что у возникших в начале палеозоя моховидных современные их семейства, а возможно, и роды в основном сложились уже в середине мезозоя. Третичное время характеризуется появлением и расселением высокоспециализированных родов, уже очень близких или даже тождественных современным. В четвертичное время продолжалось расселение моховидных и трансформация их ареалов.
Моховидные представляют собой слепую ветвь развития растений. С давних пор они прочно заняли свое особое место в экономике природы и сохранили его в сложных условиях формирования континентов, изменений климата и растительного покрова. Они выдержали эти испытания в течение геологического времени, не оскудев в морфологическом разнообразии и таксономическом богатстве и достигнув весьма широкого распространения. Массовое участие моховидных в растительном покрове Земли оказывает существенное воздействие на среду обитания других растений и животных.
В местах повышенного увлажнения в умеренных зонах северного и южного полушарий постепенно накапливались и продолжают накапливаться значительные толщи торфяных отложений с преобладающим участием моховидных.
Общеизвестно ландшафтное значение моховидных в тундровой зоне, особенно в моховых тундрах. Но изобилие этих растений свойственно и областям тропическим. Здесь, высоко в горах, обычно на высоте более 3000 м над уровнем моря, расположен особый пояс высокогорных лесов, который по праву называют поясом мшистых лесов. Известный русский ботаник и путешественник А. Н. Краснов очень образно описывает это «царство археофитов», т. е. древних растений, так: «Вы не видите ни коры, ни стеблей, все скрыто в нежном зеленом футляре, придающем в туманном полумраке леса стволам чудовищную толщину и неясные неопределенные контуры. Тонкие веточки, как инеем в морозное утро ветви наших лесов Севера, обрастают футляром зеленого мха, а этот же зеленый иней превращает в толстые канаты тонкие стебельки лиан черешки папоротниковых вай и стебельки соцветий».
Моховидные в подавляющем большинстве многолетние и только сравнительно редко однолетние растения. Но независимо от продолжительности жизни все они низкорослы. Размеры моховидных, если не учитывать нижней отмирающей части тела гаметофита, колеблются от миллиметра до нескольких сантиметров у большинства видов. Значительно реже встречаются более крупные моховидные. Так, живущие в воде мхи рода фонтиналис (Fontinalis) могут иметь стебли длиной до 60 см и более. Такими же и более длинными являются стебли некоторых мхов-эпифитов, поселяющихся на стволах и ветвях деревьев.
Моховидные отличаются сравнительно простой внутренней организацией. В их теле мож но обнаружить ассимиляционную ткань, а также слабо по сравнению с другими современными высшими растениями выраженные проводящие, механические, запасающие и покровные ткани, которые к тому же, как будет показано, отчетливо прослеживаются отнюдь не во всех систематических группах этого отдела.
При наличии сложно устроенных органов размножения у моховидных, по существу, отсутствуют специализированные вегетативные органы. Только условно можно говорить о листьях и стеблях моховидных, которые следовало бы называть соответственно филлидиями и каулидиями (чисто бриологические термины).
Структурные возможности гаплоидного гаметофита - доминирующего поколения в жизненном цикле моховидных - вообще более узки, чем диплоидного спорофита других высших растений. Этим, очевидно, и объясняется отсутствие у моховидных развитой водопроводящей системы и, как следствие этого, их ограниченный рост в высоту.
Своеобразие моховидных, как самостоятельного отдела высших растений, особенно заметно проявляется в цикле их развития.
На нашем сайте Вы найдете значение "Моховидные Как Особая Линия Эволюции Высших Растений" в словаре Биологическая энциклопедия, подробное описание, примеры использования, словосочетания с выражением Моховидные Как Особая Линия Эволюции Высших Растений, различные варианты толкований, скрытый смысл.
Первая буква "М". Общая длина 98 символа
my-dict.ru
Мохообразные болотных экосистем
Моховидные (Bryophyta) часто, даже в кругах специалистов, называют популярным и коротким именем – мхи. Однако в более точном, научном смысле собственно мхами называют представителей лишь одной, наиболее обширной группы отдела моховидных, а именно – листостебельные, или настоящие мхи (Bryopsida).
Среди высших растений моховидные образуют наиболее обособленную группу.
Моховидные иногда рассматривают в целом как растения, связанные в своей жизни с достаточным, часто избыточным увлажнением. Значение этой зависимости не следует преувеличивать. Прежде всего, все развитие гаметофита совершается у них в воздушной среде. Моховидные обнаруживают значительную устойчивость к длительному пересыханию и способны произрастать в местах неравномерного и даже весьма кратковременного сезонного увлажнения.
Мохообразные как особая линия эволюции высших растений
Моховидные представляют собой слепую ветвь развития растений. С давних пор они прочно заняли свое особое место в экономике природы и сохранили его в сложных условиях формирования континентов, изменения климата и растительного покрова. Массовое участие моховидных в растительном покрове земли оказывает существенное воздействие на среду обитания других растений и животных.
Общеизвестно ландшафтное значение моховидных в тундровой зоне, особенно в моховых тундрах.
Моховидные в подавляющем большинстве многолетние и только сравнительно редко однолетние растения. Но независимо от продолжительности жизни все они низкорослы.
Моховидные отличаются сравнительно простой организацией.
При наличии сложно устроенных органов размножения у моховидных, по существу, отсутствуют специализированные вегетативные органы.
Цикл развития моховидных
Особенность чередования поколений у моховидных заключается в том, что гаметофит у них значительно расширяет по сравнению с остальными высшими растениями, сферу своей физиологической деятельности. Наряду с обеспечением полового размножения гаметофит моховидных принимает на себя выполнение основных вегетативных функций – фотосинтеза, водоснабжение, минерального питания целого растения. Спорофит же фактически ограничивается спорообразованием.
Из одноклеточной споры у моховидных вырастает гаметофит. Прежде всего развивается многоклеточное ветвистое нитчатое или пластинчатое образование – протонема, на которой у мхов закладывается несколько или даже много почек.
Из почек в одних группах моховидных вырастают пластинчатые, в других – облиственные «побеги», которые в бриологии называют гаметофорами, поскольку они образуют половые органы. Таким образом, протонема, почки на ней и вырастающие из них гаметофоры представляют собой гаметофит (гаметофазу) моховидных.
На гаметофорах образуются половые органы – женские - архегонии и мужские - антеридии.
Оплодотворение яйцеклетки подвижными сперматозоидами происходит при наличии капельно-жидкой воды. Слияние этих гамет и дальнейшее развитие зиготы происходит внутри архегония.
Из зиготы за время от нескольких месяцев до двух лет развивается спорофит, у моховидных обычно называемый спорогоном.
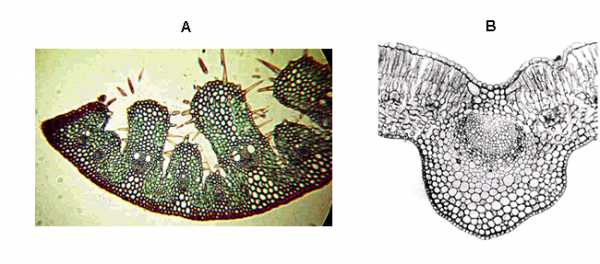
Класс - листостебельные мхи
Из всех моховидных листостебельные мхи включают наибольшее число видов. Среди них многие отличаются широким распространением, высокой жизненной устойчивостью и большой фитоценотической ролью в растительном покрове Земли. Экологическое своеобразие и особое значение мхов в природных процессах во многом зависит от присущих им групповых форм роста, благодаря чему мхи могут на больших площадях создавать рыхлые или плотные покровы различной мощности. Тем самым мхи активно участвуют в формировании на поверхности континентов многочисленных мощных влагоприемников в виде болот и замоховелых лесов, оказывающих существенное влияние на общую обеспеченность суши влагой.
Листостебельные мхи - самый крупный класс моховидных. Он состоит из 700 родов и включает 14500 или больше видов. Представители этого класса на суше встречаются почти повсеместно. Этот класс обычно делят на три подкласса: сфагновые мхи, андреевые мхи и бриевые мхи. Рассмотрим один из основных подклассов этого вида – сфагновые мхи.
Сфагновые мхи
Название происходит от латинизированного sphagnos — род мха.
Примерно 350 видов мхов, относящихся к единственному роду сфагнум Sphagnum (рисунок 1), составляют четко очерченную группу, очень давно отделившуюся от основной линии эволюции этого отдела. Стебли листостебельного гаметофита у них несут мутовки ветвей, часто по пять в узле, более тесно скученные у верхушки растения, так что возникает своего рода головка.
Рисунок 1 - Сфагновый мох
Многолетние, беловато-зеленые, желтоватые, буроватые или красноватые болотные мхи. Стебли мха высотой 5—30 см, ветвистые, ризоидов нет, листья однослойные без жилки, образованы двумя типами клеток: водоносными — крупными, мертвыми, бесцветными и пустыми с волокнами и порами в наружных стенках; и хлорофиллоносными, вытянутыми в длину, узкими, окрашенными, расположенными между водоносными.
Стеблевые и веточные листья различны по форме (языковидные, широкояйцевидные или овальные, равносторонние, заостренные, яйцевидно ланцетные), длиной 0,5—2 мм. Коробочки со спорами шаровидные, с крышечкой. Споры желтые или желтовато-бурые. Спороносят в июле — августе. Растения однодомные и двудомные. Имеют два поколения. Домиирует гаметофит (половое поколение) — облиственное растение с половыми органами — антеридиями, в которых развиваются мужские гаметы — сперматозоиды, и архегониями, в которых развиваются женские гаметы — яйцеклетки.
Бесполое поколение представлено безлистным стебельком; на его верхушке имеются коробочки со спорами. Проросшая спора образует пластинчатый заросток — протонему; из нее развивается растение сфагнума. Сфагнум растет медленно (за год вырастает до 3 см), а снизу стебель отмирает.
Сфагновые мхи – наименее требовательные к минеральному питанию болотные растения. Они довольствуются минеральными веществами, поступающими на поверхность почвы из атмосферы. Эти мхи отличаются очень высокой влагоемкостью, позволяющей некоторым видам сфагнума поглощать воды в 20-30 раз больше своего веса в сухом состоянии.
Высокая влагоемкость сфагнового мха объясняется тем, что ткани его листьев состоят в основном из мертвых гиалиновых клеток, между которыми зажаты очень мелкие живые хлорофиллоносные клетки. Из мертвых гиалиновых клеток состоит и наружный покров (гиалодермис) стеблей и ветвей (рисунок 2).
Рисунок 2 - Строение сфагнового мха
1 - общий облик Sphagnum magellanicum; 2- наружные клетки гиалодермиса стебля; 3 – наружные клетки гиалодермиса отстоящей ветки; 4 – часть поперечного среза через стебель; 5- веточный лист; 6- поперечный срез веточного листа; 7- часть ткани веточного листа (темные –хлорофилловые клетки, бесцветные – водоносные клетки с порами (а) и волокнами (б)
Так как сфагновый мох не имеет корней, минеральное питание в виде бедных солями водных растворов воспринимается всей поверхностью растения через гиалиновые клетки. Поселяясь на заболачивающихся местах, сфагнум уже сам, в условиях достаточно влажного климата, усиливает болотообразовательный процесс.
Торфяные мхи произрастают на торфяных болотах (низинах, переходных и особенно верховых). Они играют большую роль в зарастании водоемов и в заболачивании лесов и лугов. Побеги этих мхов, нарастая верхушкой и отмирая снизу, постепенно превращаются в торф. Растет по всей территории России.
Свойства.
Мхи – растения, не имеющие циркуляторной системы. Они получают влагу из осадков или атмосферы, используя осмотическое давление. Это означает также, что они одновременно поглощают все содержащиеся в окружающей среде вещества, в том числе вредные, не обладая механизмами освобождения от них. Поэтому мхи являются прекрасными индикаторами состояния окружающей среды. Сфагновые мхи способны повышать кислотность окружающей их среды, выделяя в воду ионы водорода. Наиболее важной особенностью сфагнума, приобретенной в ходе миллионов лет эволюции, является его способность впитывать и сохранять от 12 до 20 весовых частей воды на часть сухого веса (в зависимости от биологического вида сфагнума), а также его бактерицидные свойства.
Размножение.
Сфагнум может размножаться как спорами, так и вегетативно. Количество спор в спорофите может быть от 20 000 до 200 000 в зависимости от вида мха, а на квадратном метре болота – примерно 15 млн. Спорофит выбрасывает споры в июле. Коробочка как бы взрывается при сухой теплой погоде, и споры разносятся ветром на различные расстояния, так как имеют разный размер, 20-50 мкм. Еще один механизм переноса спор – потоком воды или брызгами от капель дождя. В последнем случае расстояние переноса не превышает десятка сантиметров. Размножение спорами является основным при расселении сфагнума на большие расстояния - новые или пострадавшие от пожара или хозяйственной деятельности участки. Для образования растения из споры необходимо, чтобы она попала на подходящую почву – влажный торф. Другой механизм распространения сфагнума – вегетативный, участками стебля или ветвей. Этот механизм эффективен на малых расстояниях.
Места обитания.
Основной средой обитания сфагнума в России являются болота, занимающие примерно пятую часть ее территории.
Поверхность моховой дернины очень живописна: на ней видны лишь головки сфагнума всевозможных оттенков, напоминающие узоры персидского ковра (рисунок 3).
Рисунок 3 - Головки сфагновых мхов
У сфагнума одновременно протекают процессы роста и разложения. Верхушка растет, вытягиваясь вверх на 1-3 см в год, а нижняя подводная часть отмирает и со временем превращается в торф, поэтому стебель постепенно опускается вниз. Однако, из-за постоянного накопления торфа (до 1 см в год в верхних слоях) поверхность болота медленно поднимается – формируются так называемые верховые болота, в которых обычно нет трясин, а уровень воды находится на 10-20 см ниже поверхности сфагновой дернины .
Отдельные растения мха вместе образуют могучую дернину (рисунок 4).
Рисунок 4 – Дернина сфагновых мхов
Отмирающие нижние части сфагновых мхов формируют многометровые отложения торфа. В верхних слоях идет постепенное разложение органических веществ, нижние под давлением верхних слоев уплотняются на глубине несколько метров, одному году соответствует уже слой толщиной несколько миллиметров, а возраст глубинных слоев составляет несколько тысячелетий.
Анатомия.
Гаметофор сфагнов состоит из двух типов клеток – живых, хлорофиллоносных, и мертвых, лишенных содержимого крупных водоносных, или гиалиновых, клеток.
Стенки последних укреплены спиральными волокнами, предохраняющими их от разрыва. У многих видов в стенках гиалиновых клеток имеются поры, облегчающие всасывание воды (рисунок 5).
Листья, произрастающие на веточке, налегают друг на друга, называют – черепитчато-налегающими.
Рисунок 5 - Клетки сфагна: 1 – водоносная клетка; 2 – хлорофиллоносная клетка; 3 – пора; 4 – спиральные волокна
Более сложное строение обнаруживает стебель. В нем клетки, хотя и дифференцированы, но не образуют специализированных тканей, как у сосудистых растений.
Спорогон сфагнов, как и у других мхов, состоит из ножки и коробочки. Ножка спорогона очень короткая а коробочка не имеет перистома.
Листостебельное растение (гаметофор) состоит из стебля с веточками или без них. Как на стебле, так и на веточках у бриофитов имеются листья, которые называются соответственно стеблевыми и веточными.
Экологические особенности и распространение.
Мхи способны заселять наиболее бедные и влажные местообитания. В умеренной зоне они образуют обширные торфяные болота. Торф – это неразложившиеся спрессованные остатки сфагновых мхов, которые сохраняются практически в неизменном виде столетиями и тысячелетиями. Многие болота таежной зоны имеют возраст от 8 000 до 10 000 лет. Вместе со сфагнами гниению не подвергаются и остатки других болотных растений. Происходит это потому, что торфяные мхи выделяют в окружающую среду целый букет органических кислот (яблочную, лимонную, уксусную, муравьиную и др.).
Но сфагны способны расти не только в местообитаниях с бедным минеральным питанием. Часто их можно встретить и в поймах лесных речек, и на озерах, и у родников (выходах ключевых вод). В таких местообитаниях они не образуют сплошных ковров, а также торфа, так как биохимический состав растений тех видов мхов, которые здесь произрастают, не столь химически агрессивен.
Разновидности сфагновых мхов
Sphagnum angustifolium (Russ. ex Russ. ) C. Jens – типично болотный вид, облигатно связанный с торфяными болотами на всем протяжении его ареала. Является доминантом мохового покрова в рослых рямах и мезоолиготрофных осоково – сфагновых топей верховых и переходных болот. С высоким постоянством растет на грядах ГМК и в рямах олиготрофных сфагновых болот.
Sphagnum balticum (Russ.) Russ.ex C.Jens – типично болотный вид, облигатно связанный с торфяными болотами на всем протяжении его ареала. Растет в сфагновых топях и мочажинах верховых водораздельных болот атмосферного питания, где является одним из основных доминантов и торфообразователей. Встречается часто, по всей территории.
Spagnum compactum DC. in Lam. et DC –характерный болотный вид, облигатно связанный на юго – востоке Западной Сибири с торфяными болотами. Растет в обширных сфагновых топях верховых водораздельных болот атмосферного питания. Предпочитает местообитания с угнетенным сфагновым покровом и доминированием печеночных мхов в местах выхода болотных газов.
Sphagnum fallax (Klinggr.) Klinggr –характерный болотный вид, облигатно связанный на юго – востоке Западной Сибири с торфяными болотами. Растет в мезоолиготрофных осоково – сфагновых топях верховых и переходных болот, где является основным доминантом и торфообразователем. Реже поселяется в мезотрофных осоково – сфагновых топях. Встречается часто, по всей территории.
Sphagnum lindbergii Schimp. ex Lindb (рисунок 6) –гипоарктический характерный болотный вид, облигатно связанный на юго – востоке Западной Сибири с торфяными болотами. Растет в обширных топях и мочажинах верховых водораздельных сфагновых болот.
Рисунок 6 - Сфагновый мох Sphagnum lindbergii
Sphagnum fuscum (Schimp.) Klinggr (рисунок 7) –характерный болотный вид, облигатно связанный в лесной зоне Западной Сибири с торфяными болотами. Растет на грядах и пушицево – сфагновых топях олиготрофных болотных комплексов и в рямах, являясь основным доминантом и торфообразователем верховых сфагновых болот.
Рисунок 7 - Сфагновый мох Sphagnum fuscum
Sphagnum jensenii Lindb –типично болотный вид, облигатно связанный с торфяными болотами на всем протяжении его ареала. Растет в застойных топях и мочажинах верховых сфагновых болот, где нередко является доминантом мохового яруса. Поселяется также в проточных мезо – олиготрофных и мезотрофных осоково – сфагновых топях верховых и переходных болот. Встречается сравнительно часто, по всей территории.
Sphagnum magellanicum Brid (рисунок 8) –типично болотный вид, облигатно связанный с торфяными болотами на всем протяжении его ареала. Растет в рослых и типичных рямах, в олиготрофных болотных комплексах и проточных осоково – сфагновых топях верховых сфагновых болот, реже на кочках и грядах в мезотрофных осоково – сфагновых топях и мелколесьях, а также на мезотрофных лесных сфагновых болотах. Встречается часто, по всей территории.
Рисунок 8 - Сфагновый мох Sphagnum magellanicum
biofile.ru