
Органоиды движения: функции и строение, особенности движения простейших. Жгутики реснички у растений
ЖГУТИКИ И РЕСНИЧКИ — Юнциклопедия
Жгутики и реснички — специализированные органоиды движения клеток. Различают два типа жгутиков: одни — в клетках животных и низших растений, кроме цианобактерий, а другие — у бактерий. Реснички встречаются только в клетках животных.
Жгутики бактерий — это тонкие (15— 20 нм), полые в середине нити, построенные из одного белка — флагеллина. Нити волнообразно изогнуты, имеют постоянную форму и сами не способны двигаться. В мембране бактериальной клетки находится базальное тельце жгутика, имеющее сложный белковый состав и строение. Жгутик соединяется с базальным тельцем с помощью крюка. Он совершает вращательные движения (истинное вращение). Вращающим элементом служит базальное тельце, движущееся внутри бактериальной мембраны, подобно ротору в электромоторе. Интересно, что и источник энергии для вращения жгутика не энергия химических веществ (типа АТФ), а разность электрохимических потенциалов на мембране бактериальной клетки.
Жгутики и реснички животных и растений гораздо крупнее. Они имеют диаметр около 250 нм и достигают в длину нескольких миллиметров. В отличие от бактериальных эти жгутики покрыты мембраной и обладают собственной подвижностью.
Строение и принцип работы жгутиков и ресничек совершенно одинаковы. Различия между ними лишь в количестве: обычно на одну клетку приходится один или несколько жгутиков, а ресничек до нескольких тысяч.
Под мембраной у жгутиков (ресничек) располагается стержневая структура — аксоне-ма. Она состоит из 9 спаренных микротрубочек, расположенных по окружности, и 2 одиночных микротрубочек в центре. Центральные и периферические микротрубочки соединены между собой системой связок. В состав периферических микротрубочек входит белок ту-булин и специфический для жгутиков белок — динеин. Динеин использует энергию АТФ (см. Аденозинтрифосфорная кислота (АТФ), расщепляя ее до АДФ (аденозиндифосфорной кислоты), и, взаимодействуя с тубулином, превращает энергию в механическую работу по перемещению спаренных микротрубочек относительно соседних — скольжению. Две центральные микротрубочки и система связок превращают скольжение отдельных микротрубочек внутри аксонемы в изгибание всей ак-сонемы, а это и приводит в движение жгутик (ресничку). Движение состоит либо из псевдовращения (такое движение мы производим, например, делая вращательные движения рукой) — так чаще всего бьются жгутики; либо из возвратно-поступательных колебаний в одной плоскости, типичных для ресничек.
В основании жгутиков (ресничек) лежат базальные тельца, но в отличие от бактериальных они не связаны с движением, а служат для роста аксонемы и закрепления жгутика (реснички) в клетке. Часто от базальных телец в глубь цитоплазмы отходят дополнительные заякоривающие структуры — исчерченные корешки.
Жгутики и реснички обеспечивают движение свободноживущих клеток. Реснички создают поток жидкости вдоль поверхности неподвижных клеток (например, в дыхательных путях согласованное биение миллионов ресничек обеспечивает удаление из легких пылевых частиц) .
У некоторых животных (например, млекопитающих) имеются наряду с подвижными ресничками еще и неподвижные — чувствительные реснички. Последние не имеют двух центральных микротрубочек и иногда вообще лишены аксонемы, но всегда имеют в основании базальное тельце. Таковы палочки сетчатки глаза, реснички обонятельных луковиц, реснички во внутреннем ухе (орган равновесия).
yunc.org
20. Строение и значение микроворсинок, ресничек и жгутиков. Представление о базальной исчерчености (базальных впячиваниях).
Микроворсинки - структуры клетки, располагающиеся на ее свободной поверхности и выступающие во внеклеточное пространство,
Эти выпячивания значительно расширяют поверхность клетки. Микроворсинки активно всасывают и (или) активно переваривают вещества на поверхности клеток. Микроворсинкам присущи некоторым популяциям эпителиоцитов. Нередко для клеток, имеющих много микроворсинок, характерна высокая ферментативная активность и счет белков, прикрепляющихся к гликокаликсу (столбчатые энтероциты кишечника, тироциты щитовидной железы и т. д.).
Микроворсинки сформированы клеточной мембраной к гиалоплазмой.
Структурной основой каркаса микроворсинок служат тонкие микрофиламенты. (+)-конец микрофиламентов направлен на периферию и стыкуется с электронноплотным аморфным веществом дистальной части микроворсинки. Тонкие микрофиламенты в микроворсинке лежат параллельно друг к другу на расстоянии около 10 нм, регулярно соединяясь между собой с помощью белков - фимбрина и фаоцима. С мембранами они взаимодействуют с помощью минимиозина и фасцина. На поверхности микроворсинок нередко выявляется высокая концентрация белков с транспортно-насосными функциями (всасывающие эпителии).
Сходное строение имеют стереоциллии, однако в отличие от микроворсинок, стереоцилии имеют гораздо большие размеры и не обладают всасывающими функциями.
Реснички и жгутики. Реснички и жгутики на ультраструктурном уровне имеют сходные принципы строения, ко могут иметь разные функции. Реснички перемещают поверхностный субстрат полого органа, тогда как жгутик сперматозоида позволяет передвигаться самой клетке.
Реснички состоят из базалъного тельца и выступающей из клетки аксонемы. Базальное тельце по строению аналогично дочерней центриоле и состоит из девяти триплетов микротрубочек.
В основе аксонемы тоже лежат микротрубочки. Они формируются от базального тельца и достигают своим дистальным (-) концом гомогенного матрикса. Имеется девять дуплетов микротрубочек и две центральные микротрубочки. Каждый периферический дуплет представлен: а- микротрубочкой (полной) и б - микротрубочкой (неполной). Соседние дуплеты соединены между собой линейном (обладающим АТФ-азной активностью и позволяющим микро-трубочкам скользить по отношению друг к другу) и индогеном.
К центральным микротрубочкам от периферических радиально направляются спицы. Центральные же микротрубочки объединены втулкой.
Базальные инвагинации (впячивания).
Базальные впячивания – это внедрения цитолеммы в цитоплазму, прилежащие к базальной мембране или иной плотной структуре, например, впячивание цитолеммы на остеокласте со стороны разрушаемой (резорбцируемой) кости, существенно расширяющие поверхность клетки с внутренней (базальной) поверхности. Нередко в зонах впячиваний цитолеммы видны многочисленные мнтохондрии. Совокупность базальных инвагинаций и митохондрий формирует базальную исчерченность.
Наличие базальной исчерченности - признак активных процессов трансмембранного перекоса веществ, когда наряду с увеличением поверхности значение имеет и высокий уровень энергетического потребления, обеспечиваемый АТФ.
Базальные впячивания формируют сложный лабиринт каналов и ходов, взаимопереплетений. На их поверхности обнаруживаете высокая концентрация ионных каналов, рецепторов, нередко - ферментативная активность. Форму базальных впячиваний поддерживают структуры цитоскелета - в первую очередь промежуточные филаменты и тонкие микрофиламенты. Состав промежуточных филаментов разнообразен и зависит от тканевой принадлежности клетки. В эпителиях - это цитокератины. Они формируют микрофибриллы. В зоне впячиваний часто можно встретить полудесмосомальные контакты.
86
studfiles.net
Реснички жгутики - Справочник химика 21
Центриоли, реснички, жгутики и микротрубочки [c.34]Микротрубочки. Тонкие трубочки, собранные из двух типов субъединиц глобулярных белков присутствуют в ресничках, жгутиках и других сократительных и двигательных структурах. [c.1014]
Мерцательное движение распространено у растительных и животных организмов. Оно осуществляется жгутиками и ресничками. Жгутики встречаются у многих одноклеточных животных и растений и в клетках многоклеточных. [c.80]
Важной составной частью цитоплазмы являются микротрубочки— полые стерженьки, наружный диаметр которых составляет 24 2 нм, а внутренний 13—15 нм. Наиболее удивительна их форма в жгутиках и ресничках эукариотических клеток (рис. 1-5). Устойчивые микротрубочки ресничек являются, по-видимому, неотъемлемой частью аппарата, обеспечивающего движение жгутиков (Приведенный справа рисунок взят из работы .) Лабильные (т. е. образующиеся, а затем распадающиеся) микротрубочки обнаруживаются чаще всего в цитоплазме клеток, способных к перемещению (например, в псевдоподиях амеб). Митотическое веретено (гл. 15, разд. Г.9) представляет собой набор микротрубочек, обеспечивающих перемещения хромосом в делящейся клетке. Микротрубочки обнаруживаются также в плоскостях деления растительных клеток. [c.276]Некоторые клетки и организмы способны сокращаться и перемещаться благодаря специальным белкам. К таким белкам относятся, в частности, актин и миозин, функционирующие в сократительной системе мышечной ткани. Другим белком, обеспечивающим перемещение клеток, является тубулин, входящий в состав микротрубочек — основных компонентов ресничек и жгутиков клетки. [c.46]
Микротрубочки и микрофиламенты, по-видимому, исполняют роль цитоскелета и формируются из белка тубулина. Они входят в состав центриолей, играющих важную роль в делении ядра, а также в состав жгутиков и ресничек. [c.42]
Реснички и жгутики позволяют клеткам передвигаться [c.42]
| Рис. 2-18. Реснички и жгутики имеют одинаковое строение, но реснички значительно короче. |  |
Еще одна система длинных нитевидных белков имеется в микротрубочках, о которых речь шла вьппе (разд. 2.14 и 2.15). Микротрубочки-это длинные полые трубки, каждая из которых построена из 13 белковых нитей, уложенных параллельно друг другу вокруг центральной полости. Каждая нить состоит из чередующихся молекул двух глобулярных белков-а-тубулина и р-тубулина. Микротрубочки входят в состав ресничек и жгутиков эукариот их взаимное скольжение или скручивание относительно друг друга сообщает ресничкам и жгутикам характерное винтообразное, вращательное или волнообразное движение, обеспечивающее перемещение клеток. Микротрубочки участвуют во многих других видах клеточной активности, например в делении клеток некоторым клеткам они придают ту или иную форму. Движение микротрубочек в жгутиках тоже зависит от гидролиза АТР. [c.183]
Реснички и жгутики позволяют [c.360]
Двигательные органеллы. У всех эукариотических клеток, имеющих жгутики или реснички (у простейших, водорослей, сперматозоидов и клеток мерцательного эпителия), строение их одинаково. На поперечном срезе видно, что на периферии жгутика располагаются девять двойных нитей, а в центре-две одиночные нити (структура 9 + 2 ). Снаружи вся эта система одета плазматической мембраной. Основание жгутика закреплено в наружном слое цитоплазмы при помощи базального тельца, или блефаропласта. Блефаропласт представляет собой производное самовоспроизводящейся органеллы (центриоли). [c.27]
Жгутики (или реснички) типа 9 + 2. [c.522]
Перемещаются с помощью жгутиков или ресничек. Некоторые питаются бактериями. Размеры 5—80 мкм [c.230]
Некоторые палочковидные формы имеют жгутики или реснички, с помощью которых они двигаются. [c.23]
Проблемы, связанные с молекулярными основами превращений химической энергии АТФ в механическую энергию процессов сокращения и движения, чрезвычайно сложны [3, 15]. Это объясняется тем, что вне живого организма отсутствуют примеры непосредственного превращения химической энергии в механическую. Механическая работа может быть представлена сокращением мышц, а также движениями ресничек и жгутиков у простейших. Большинство клеток содержат сократительные нити (фибриллы), которые осуществляют организацию содержимого клетки, движение и перенос клеточных веществ, процессы клеточного деления и т. д. В качестве примера преобразования энергии АТФ в механическую работу можно привести процессы мышечного сокращения, связанные с использованием энергии АТФ [3, 15, 18], при этом важную функцию выполняют белковые компоненты мышечных клеток — комплекс миозина и актина, названный актомиозином. Актомиозин и его компонент миозин обладают АТФ-азной активностью, т. е. способны гидролизовать концевую фосфатную группу АТФ. Однако АТФ-азную активность актомиозина стимулируют ионы Mg +, а миозина — ионы Са +. Сигналом для сокращения мышц является электрический импульс, приходящий из двигательного нерва через нервномышечное соединение. До получения импульса по обе стороны мембраны (сарколемма) мышечной клетки поддерживается, разность потенциалов (с наружной стороны имеется избыточный положительный заряд). При распространении импульса по мембране разность потенциалов сразу исчезает. Считают, что это является результатом резкого повышения проницаемости мембраны для ионов К+, Na+ и Са2+ при этом направление потоков ионов вызывает разряд трансмембранного потенциала. После этого мембрана вновь возвращается в поляризованное состояние, а ионы Са + входят внутрь саркоплазматической сети мышечной клетки. Подобный перенос ионов Са + осуществляется за счет свободной энергии гидролиза АТФ (АТФ-азный кальциевый насос мембраны). Поставщиками АТФ в мышечных клетках служат как гликолиз, так и дыхание. Однако при нарушении этих процессов мышца (скелетная мышца позвоночных животных) при стимуляции продолжает сокращаться благодаря тому, что в ней содержится богатое энергией вещество — креатинфосфат (см. стр. 416), концентрация которого более чем в 4 раза превышает концентрацию АТФ. В мышце идет реакция [c.430]
Базальные тельца, реснички и жгутики [c.203]
Различные рецепторные клетки имеют выросты — антенны, плазматические мембраны которых содержат белки, специфические для данного вида рецепции. Антенны могут состоять из так называемых микровилл и являющихся результатами их диффе-ренцировки стереоциллий или киноциллий — ресничек, жгутиков и их производных. В эти образования входят фибриллярные белки. Жгутики обычно построены по принципу 9-2 + 2, т. е. имеют 9 пар фибрилл на периферии и одну в центре (см. 12.6). В других случаях структурный аппарат антенн представлен формулой 9-2 + 0. [c.355]
Различные группы водных организмов (гидробионтов) присп.о-собили ь к определенным зонам обитания, образуя биоценозы в предел единой экологической системы (водоема). Толща воды населена микроорганизмами планктона. Растительные формы называются фитопланктоном, а животные организмы — зоопланктоном. Мно микроорганизмы зоопланктона (простейшие) обладают определенной способностью к перемещению с помощью ресничек, жгутиков. Коловратки совершают прыгательные движения или плавают, исйользуя реснички. [c.231]
Осн. ф-ция К.-активация мн. ферментов аденилатциклазы, фосфодиэстеразы циклич. нуклеотидов, киназы фосфо-рилаз и легких цепей миозина (киназы-ферменты, катализирующие перенос фосфорильной группы с АТФ на субстрат), Са -зависимой протеинкиназы цитоплазмы и мембран, фосфолипазы Aj и др. Благодаря этому он влияет на гликогенолиз и липолиз, секрецию нейромедиаторов, адренергич. передачу регуляторного сигнала, изменяет функциональные св-ва рецепторов, ускоряет активный транспорт Са в сердце и мозге, препятствует гуанозинтрифосфат-зависимой полимеризации тубулина (белок, из к-рого состоят жгутики и реснички клеток животных и растений), влияет на скорость деления клеток. [c.293]
По структуре центриоли сходны со жгутиками или более короткими образованиями — ресничками (эти термины, в сущности, синонимы), обычно находятся на поверхности клеток эукариот и являются органами движения. Неподвижные клетки тела человека также нередко имеют реснички. Например, эпителий бронхов несет 10 ресничек на 1 см Г26]. Модифицированные жгутики образуют светочувствительные рецепторы нашего глаза и рецепторы вкуса на языке. Жгутики и реснички несколько больше по диаметру (около 0,2 мкм), чем центриоли, и обладают характерной внутренней структурой они состоят из И полых микротрубочек диаметром 24 нм, организованных по схеме 9 + 2 (рис. 1-5 и 1-6). Каждая микротрубочка внешне похожа на жгутик бактерии, но существенно отличается от него по химическому составу. Базальное тельце, называемое также кинетосомой (рис. 1-5), по структуре, размерам и способу воспроизведения сходно с центриолью. Микротрубочки, подобные тем, которые входят в состав жгутиков, обнаружены также в цитоплазме клеток [27]. Они выглядят как маленькие канальцы, но действительно ли играют такую роль — неясно. Скорее всего микротрубочки выполняют опорную функцию цитоокелета . В аксоне нерва микротрубочки расположены по всей длине аксона и, вероятно, составляют часть механической системы переноса клеточных компонентов. [c.37]
Микротрубочки ресничек (и жгутиков) эукариот имеют другую структуру. В большей части реснички они находятся в форме слившихся inap А-трубочка содержит ручку , и В-трубочка присоединяется к ней за счет общих субъединиц, расположенных в центре структуры. Как и из лабильных микроррубочек, было выделено два типа молекул тубулина, однако вопрос о том, как они вписываются в структуру опа-ренных микротрубочек, остается открытым. [c.279]
Мышечное сокращение — лучше всего изученное механохи-мическое явление. К таким же явлениям относится множество биологических процессов движения растений движения клеток с помощью жгутиков и ресничек-, вся совокупность движений в дроцессах митоза и мейоза движения внутри не делящейся клетки сократительные процессы в хвостах фаговых частиц механохимические процессы в мембранах движение рибосом относительно мРНК в полисомах акустическая и механорецепция. [c.411]
Белки жгутиков и ресничек пока недостаточно изучены. Установлена их АТФ-азная активность. Формы и частоты волнообразных движений, распространяющихся вдоль жгутика от основания к концу, зависят от концентрации АТФ. Белки являются сократительными, но отличными от миозина или актина. [c.412]
Модельные теории работы жгутиков и ресггичек рассматривают изменения конформационного состояния белковых сократительных единиц, когда до них доходит сократительная волна. В каждой единице происходит расщепление АТФ. В жгутиках и ресничках реализуется скольжение субфибрилл, ручки функционируют подобно мостикам в актомиозине мышцы. Детальная молекулярная теория, количественно объясняющая волновое движение жгутиков и ресничек, еще не построена. [c.412]
При митозе и мейозе происходит перемещение протоплазмы Т1 хромосом. Митотический аппарат клетки состоит из видимых под микроскопом тяжей, соединяющих друг с другом центриоли л хромосомы с центриолями. Центриоли имеют структуру 9 + 2 , подобную структуре жгутиков и ресничек. Показано, что жгутики сперматозоидов вырастают из центриолей и кинетохоров хромосом. [c.414]
Простейший вид фоторецепции — это фототаксис. Одноклеточное эвглена обладает оранжево-красным фоторецепторным пятном — стигмой. Можно думать, что этот фоторецептор возник эволюционно из жгутика или реснички, ассоциировавшей пигмент. Под действием света стигма г.о.чдяет своего рода норнный импульс в жгутике, и эвглена движется по направлению к свету. Как указывает Уолкен, детально исследовавший фототаксис, система стигма—жгутик может рассматриваться как серво-механизм с обратной связью, обеспечивающий максимальную освещенность организма. [c.467]
Родственный класс составляют так называемые двигательные белки. Из них наиболее известны белки сократительного аппарата мышц—актин и миозин. Их разновидностью являются динеин ресничек и жгутиков простейших, спектрин мембран эритроцитов, нейростенин пресинаптпческих мембран и т. п. Сюда можно отнести и белки бактерий, ответственные за движение в градиенте концентраций различных веществ (хемотаксис), в частности мальтозу-связывающий белок Е.со(1, [c.22]
Жгутиковые протозоа (рис. 3.3,а) — мельчайшие организмы, размер которых колеблется от 10 до 50 мкм. Их высокая подвижность обусловлена резкими маховыми движениями жгутиков (подобными движениям хлыста). Многие из этих организмов питаются твердой пищей, хотя некоторые разновидности используют для своего питания растворенные органические вещества. Амебы (рис. 3.3,6) передвигаются и всасывают пищу при помощи подвижной протоплазмы. Хотя амебы не так широко распространены, как другие формы протозоа, они часто присутствуют в слизистом налете на материале загрузки биофильтров и на стенках аэротенков. Свободно плавающие протозоа (рис. 3.3,6) имеют реснички, используемые для перемещения и захватывания пищи (органических веществ). Их легко наблюдать в воде под микроскопом благодаря быстрому перемещению и довольно большим размерам — от 50 до 300 мкм. Стебельковые формы (рис. 3.3,г) прикрепляются к взвешенным в воде твердым частицам и иапользуют реснички для передвижения н добычи пищи. [c.56]
Реснички и жгутики-подвижные структуры, или отростки, выступающие с поверхности многих одноклеточных эукариот и некоторых клеток животных (но не растений), построены по одному общему архитектурному плану (рис. 2-18). Важно, однако, подчеркнуть, что жгутики эукариот очень сильно отличаются от жгутиков прокариот. Жгутики прокариот намного тоньше (10-20 нм) и состоят из отдельных белковьгх нитей. Они представляют собой упругие, изогнутые стерженьки, вращательное движение которых целиком зависит от расположенных в клеточной мембране моторов . Жгутики эукариот гораздо толще (200 нм), имеют более сложную структуру и способны самостоятельно вращаться по всей своей длине. Реснички и жгутики эукариот окружены выступами клеточной мембраны и содержат по 9 пар микротрубочек, расположенных вокруг 2 цен-тральньЕс трубочек при этом образуется так назьшаемая структура 9 + 2 (рис. 2-18). Реснички й жгутики имеют одинаковый диаметр, но длина ресничек (не превышающая 10 мкм) значительно меньше длины жгутиков (не более 200 мкм). В большинстве случаев реснички служат для того, чтобы передвигать вещества вдоль поверхности клетки с помощью волнообразных, напоминающих греблю движений, тогда как жгутики действуют как пропеллеры, проталкиваю- [c.42]Пары микротрубочек окружены выступающими за пределы клетки цитоплазмой и клеточной мембраной. Скольжение и закручивание микротрубочек относительно друг друга, стимулиро-ва нные АТР, вызывают волнообразные движения жгутиков. А. Параллельно расположенные микротрубочки ресничек на продольном срезе. [c.43]
Б. Поперечный срез ресничек. Видна структура 9-1-2, образованная девятью парами или девятью двойными микротрубочками, расположенными вокруг двух центральньгх трубочек. В. Принцип действия жгутика сперматозоида, проталкивающего вперед всю клетку. [c.43]
Скольжение нитей или микротрубочек относительно друг друга за счет энергии АТР-это процесс, лежащий в основе сокращения скелетной мьшщы, вращательных движений ресничек и жгутиков, а также образования характерных вьшя-чиваний, углублений и складок при дви- [c.43]
Растительные клетки содержат также пластиды других типов. В бесцветных лейкопластах запасаются крахмал и масла. Значительное место во многих растительных клетках занимают окруженные одиночной мембраной крупные пузьфь-ш-вакуоли (см. рис. 2-21). Они заполнены клеточным соком и различными продуктами, являющимися отходами метаболизма. Эти продукты часто агрегируют с образованием кристаллических отложений. В молодых клетках вакуоли имеют небольшую величину, но по мере старения клеток их размеры увеличиваются, и часто они заполняют весь объем клетки. Вакуоли встречаются также и в некоторых животных клетках, но здесь они, как правило, значительно мельче. У растительных клеток нет ни ресничек, ни жгутиков. [c.47]
Одна пз самых важных функций белков, с которой мы встречаемся у всех видов живых существ, — прямое превращение химической энергии в механическую работу. В этом отношении самый совершенный механизм — поперечно-полосатая скелетная мышца высших животных. Примеры механохимической активности в гораздо более простых организмах — движение жгутиков (флагелл) у бактерий и мерцательных ресничек у парамеций, сокращение хвоста бактериофага при инъекции ДНК в заражаемую клетку. Наконец, процесс сжатия и расширения митохондрий, [c.186]
Простейшие Protozoa Golfuss—одноклеточные животные. Единственная клетка выполняет все жизненные функции организма, которые у многоклеточных животных осуществляются разнообразными тканями и органами. Эти функции у простейших выполняются рядом мелких органелл — жгутиков, ресничек, мембран, скелетных образований, развитых у различных групп совершенно по-разному. [c.386]
Функции белков в клетках и в организме в целом очень разнообразны. Белки образуют такой инертный материал, как волос, рог или кость, и из белков же состоит сократимое вещество мышечного волокна. Эти последние белки обладают одной характерной особенностью — способностью превращать химическую энергию в механическую поэтому именно они обусловливают подвижность высших организмов. Точно так же у низших организмов механическое движение (движение жгутиков, координированные удары ресничек, амёбоидное движение) тесно связано с наличием сократимого белка. Касаясь других функций белка, необходимо указать, что ферменты — эти крайне важные для живых организмюв катализаторы, — дыхательные пигменты, гормоны некоторых желез (поджелудочной, щитовидной и гипофиза), антитела и токсины некоторых бактерий представляют собой белки. Из сказанного ясно, что значение белков для живого организма трудно переоценить. [c.6]
У трехслойных животных значительно совершеннее становится работа мьшщ. Необходимость в этом связана с увеличением размеров животного, поскольку реснички и жгутики теперь уже не могут обеспечить локомоторные функции. [c.80]
Реснички и жгутики идентичны по своему строению, но жгутики длиннее ресничек. Обе эти органеллы представляют собой выросты клеток. Движутся они либо однонаправленно (биение ресничек), либо волнообразно (движения жгутиков). Служат реснички и жгутики как для передвижения отдельных клеток, так и для того, чтобы перегонять жидкость вдоль поверхности клеток (так перегоняют ресничю слизь в дыхатель-ньгх путях). В основании каждой реснички и жгутика всегда обнаруживается базальное тельце. По своему строению базальные тельца идентичны центриолям и можно думать, что они образуются путем удвоения центриолей. Вероятно, они также действуют как центры организации микротрубочек, потому что ресничкам и жгутикам тоже свойственно характерное расположение микротрубочек ( 9 + 2 гл. 18). [c.203]
chem21.info
Реснички и жгутики
⇐ ПредыдущаяСтр 28 из 78Следующая ⇒Ресничка представляет собой вырост клетки, окруженный плазмалеммой. У основания реснички на уровне кортикального слоя цитоплазмы находится базальное тельце (кинетосома), которое образовано девятью периферическими триплетами коротких микротрубочек, окружающих один белковый центральный цилиндр. Каждый периферический триплет соединен с ним посредством белковых «спиц».
От каждой наружной микротрубочки отходит отросток, напоминающий флажок. Над базальным тельцем в названный цитоцлазматический вырост направляется осевая нить (осевой филамент, аксонема), тоже образованная микротрубочками (строение последних описано ранее). От каждого базального тельца к соседнему (к основанию другой реснички) отходит короткий боковой отросток, а в глубь цитоплазмы — поперечно исчерченный базальный корешок (периодичность исчерченности равна 65 нм).
Рис. 42. Ресничка I - общая структура, II - срез через внеклеточную часть, III - срез через внутриклеточную часть, IV - срез через базальное тельце, 1 - корешок, 2 - базальное тельце, 3 - плазмалемма поверхности клетки, 4 - микроворсинки, окружающие ресничку, 5 - центральна? капсула, 6 - дуплеты периферических микротрубочек, 7 - центральная пара микротрубочек, 8 - плазмалемма на поверхности реснички, 9 - динеиновые "ручки", 10-радиальная спица, 11 - нексии, 12 - субьединицаА, 13- субъединица В. 14- субъединица С, 15 - центральный цилиндр (по Р. Крстичуи по Ю. С. Ченцову, с изменениями).
Сразу над базальным тельцем микротрубочки аксонемы тоже образуют девять периферических триплетов, но над уровнем плазмалеммы в периферических комплексах одна из микротрубочек редуцируется; в центральной группе появляется пара микротрубочек, окруженная белковой оболочкой, имеющей вид цилиндра. Поэтому на большей части протяжения реснички тянутся дуплеты микротрубочек. Девять дуплетов располагаются на периферии, один — в центре.
В результате на поперечном сечении реснички ее возвышающаяся часть напоминает колесо с девятью спицами, в центре которого лежит нечетко контурирующаяся центральная капсула, окружающая две центральные одиночные микротрубочки, с которыми она связана белковыми отростками. Периферические дуплеты состоят из двух микротрубочек (А и В) и окружают центральную капсулу (см. рис. 41).
От микротрубочки А к центральной капсуле отходит радиальная «спица» с периодичностью расположения вдоль аксонемы, равной 29 нм. По направлению к соседней трубочке В отходит пара «ручек», образованных, как и «спицы», белком динеином с периодичностью 24 нм. Соседние дуплеты микротрубочки соединены между собой с периодичностью 86 нм поперечными мостиками, образованными белком нексином.
Реснички являются производными не только поверхностного комплекса клетки, но и клеточного центра. В начале их развития происходит многократная редупликация цен-триолей. Новые центриоли парами мигрируют к поверхности клетки. Здесь происходит их модификация.
Одна из центриолей (проксимальная) ложится в поверхностном комплексе вблизи от плазмалеммы у основания будущей реснички. Другая центриоль (дистальна] ложится между ней и плазмалеммой. Триплеты микротрубочек дистальной центриоли становятся дуплетами, в проксимальной центриоли триплеты сохраняются. В клеточном центре идет сборка микротрубочек, они направляются к дистальной центриоли и наращивают ее длину. Дистальная центриоль растет и влечет за собой плазмалемму. Рост аксонемы в длину происходит за счет полимеризации тубулина у дистальных концов двух внутренних трубочек каждого триплета базального тельца.
Жгутик (flagellum) эукариотической клетки (например, сдермия) напоминает ресничку, но он длиннее.
Реснички и жгутики выполняют функцию движения. Все реснички клетки совершают координированные колебания. Это достигается посредством скольжения дуплетов микротрубочек относительно друг друга. Оно обусловлено изменениями конфигурации молекул белка динеина- Динеин обладает аденозинтрифосфатазной активностью. При гидролизе АТФ выделяется свободная энергия, за счет которой динеиновые ручки выпрямляются, контактируют с соседним дуплетом микротрубочек и сдвигают его по направлению к верхушке реснички. При регенерации АТФ ручки отделяются от соседнего дуплета и опускаются вниз к основанию реснички.
Все реснички клетки совершают координированные колебательные движения. Они похожи на движения рук пловца брассом. Сначала ресничка резко наклоняется над поверхностью клетки. При этом слизь, которой обычно здесь покрыта поверхность, прогоняется в направлении наклона. Далее наклоненная ресничка совершает поворот на 180°. Поскольку она скользит параллельно поверхности клетки, слизь над ней назад не возвращается. Затем ресничка снова выпрямляется и перемещает слизь. Потом начинается новый цикл.
Число ресничек достигает нескольких сотен. Так, до 250 ресничек длиной 5—15 мкм и диаметром 0,15—0,25 мкм покрывают апикальную поверхность реснитчатых эпителиоцитов верхних дыхательных путей, маточных труб, семявыводящих канальцев.
mykonspekts.ru
Органоиды движения простейших: строение и функции
Клетки могут перемещаться при помощи специализированных органоидов, к которым относятся реснички и жгутики. Реснички клеток всегда многочисленны (у простейших их количество исчисляется сотнями и тысячами), а длина составляет 10-15мкм. Жгутиков же чаще всего 1-8, длина их — 20-50мкм.
Строение и функции органоидов движения
Строение ресничек и жгутиков, как у растительных, так и животных клеток сходно. Под электронным микроскопом обнаружено, что реснички и жгутики это немембранные органоиды, состоящие из микротрубочек. Две из них располагаются в центре, а вокруг них по периферии лежат еще 9 пар микротрубочек. Вся эта структура покрыта цитоплазматической мембраной, являющейся продолжением клеточной мембраны.
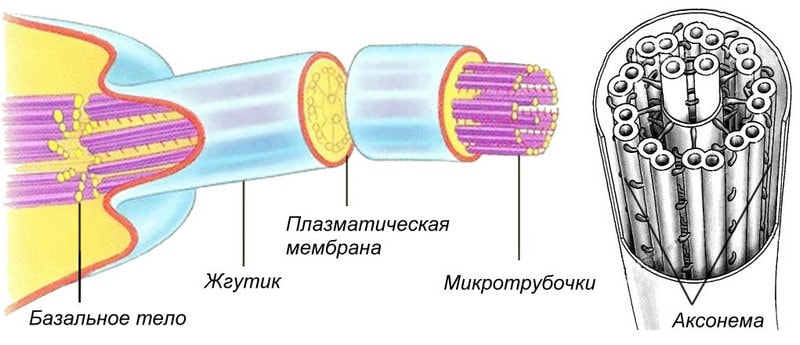
Жгутики и реснички обеспечивают не только передвижение клеток в пространстве, но и перемещение различных веществ на поверхности клеток, а также попадание пищевых частиц в клетку. У основания ресничек и жгутиков находятся базальные тельца, которые тоже состоят из микротрубочек.
Предполагают, что базальные тельца являются центром формирования микротрубочек жгутиков и ресничек. Базальные тельца, в свою очередь, нередко происходят из клеточного центра.
Большое количество одноклеточных организмов и некоторые клетки многоклеточных не имеют специальных органоидов движения и передвигаются при помощи псевдоподий (ложноножек), которое получило название амебоидного. В основе его лежит движение молекул особых белков, называемых сократимыми.
Особенности движения простейших
Одноклеточные организмы также способны передвигаться (инфузория туфелька, эвглена зеленая, амеба обыкновенная). Для перемещения в толще воды каждая особь наделена специфическими органоидами. У простейших такими органоидами являются реснички, жгутики, ложноножки.
Эвглена зелёная
Эвглена зелёная — представитель простейших из класса жгутиковых. Тело эвглены веретенообразной формы, удлиненное с заостренным концом. Органоиды движения эвглены зеленой представлены жгутиком, который находится на тупом конце. Жгутики — это тонкие выросты тела, число которых варьирует от одного до десятков.
Механизм движения при помощи жгутика отличается у разных видов. В основном это вращение в виде конуса, вершина которого обращена к телу. Перемещение наиболее эффективно при достижении углом вершины конуса 45°. Скорость колеблется в пределах от 10 до 40 оборотов за секунду. Часто наблюдается помимо вращательного движения жгутика, также его волнообразные покачивания.
Такой характер движения свойствен для одножгутиковых видов. У многожгутиковых нередко жгутики располагаются в одной плоскости и не формируют конуса вращения.
Микроскопическое строение жгутиков довольно сложное. Они окружены тонкой оболочкой, которая является продолжением наружного слоя эктоплазмы — пелликулы. Внутреннее пространство жгутика заполнено цитоплазмой и продольно расположенными нитями — фибриллами.
Периферически расположенные фибриллы отвечают за осуществление движения, а центральные выполняют опорную функцию.
Инфузория туфелька
Передвигается инфузория туфелька за счет ресничек, осуществляя ими волнообразные движения. Направляется вперед тупым концом.
Реснички двигаются в одной плоскости и делают прямой удар после полного выпрямления, а возвратный — в выгнутом положении. Удары идут последовательно один за другим с небольшой задержкой. Во время плаванья, инфузория осуществляет вращательные движения вокруг продольной оси.
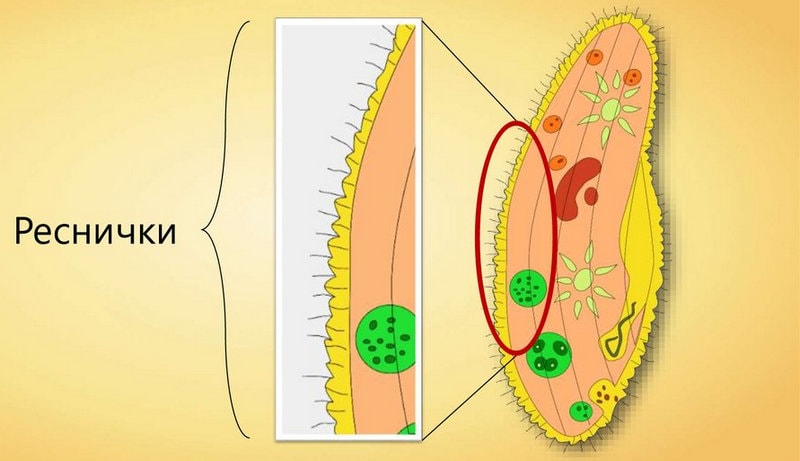 Реснички инфузории туфельки
Реснички инфузории туфелькиПеремещается туфелька со скоростью до 2,5мм/c. Направленность меняется за счёт перегибов тела. Если на пути будет преграда, то после столкновения инфузория начинает двигаться в противоположную сторону.
Все реснички инфузорииимеют сходное строение с жгутиками эвглены зеленой. Ресничка у основания образует базальное зерно, которое играет важную роль в механизме движения организма.
У некоторых инфузорий реснички соединяются между собой и таким образом позволяют развить большую скорость.
Инфузории относятся к высокоорганизованным простейшим и свою двигательную активность они осуществляют с помощью сокращений. Форма тела простейшего может меняться, а после возвращаться в прежнее состояние. Быстрые сократительные движения возможны благодаря наличию особых волокон — мионем.
Амеба обыкновенная
Амеба — простейшее довольно крупных размеров (до 0,5мм). Форма тела полиподиальная, обусловлена наличием множественных псевдоподий — это выросты с внутренней циркуляцией цитоплазмы.
У амебы обыкновенной псевдоподии еще называют ложноножками. Направляя ложноножки в разные стороны, амёба развивает скорость в 0,2 мм/минуту.
К органоидам движения простейших не относятся цитоплазма, ядро, вакуоли, рибосомы, лизосомы, ЭПР, Аппарат Гольджи.
animals-world.ru
Строение и механизм сокращения ресничек, жгутиков и мышечных волокон
Жгутики и реснички представляют собой подвижные цитоплазматические отростки, служащие либо для передвижения всего организма (у протист, ресничных червей) или репродуктивных клеток, либо для транспорта частиц и жидкостей (например, реснички мерцательных клеток слизистой оболочки носовых полостей и трахей, яйцеводов и т. д.). Жгутики эукариотических клеток по всей длине содержат 20 микротрубочек ( 9 периферических дублетов и 2 центральные одиночные), у основания жгутика в цитоплазме располагается базальное тельце. Длина жгутика до 100 мкм и больше. Более короткие (10-20 мкм) жгутики, которых бывает много на одной клетке, называются ресничками. Скольжение микротрубочек, входящих в состав жгутиков или ресничек, вызывает их биение, что обеспечивает перемещение клетки либо продвижение частиц.
Специализированные сократительные клетки мышечных волокон животных имеют в своей цитоплазме сократительные фибриллы — миофибриллы. Особенно много миофибрилл в скелетных мышечных волокнах, в клетках сердечной мышцы и гладкой мускулатуры. Пучки миофибрилл окрашены неравномерно: через равные промежутки чередуются темные и светлые участки. Темные участки названы анизотропными дисками (A-диски), светлые — изотропными дисками (I-диски). Светлый I-диск пересекается темным Z-диском (Z-полоска). Таким образом, миофибрилла представляет собой нить толщиной около 0,5 мкм с чередующимися участками:
A + ½I + Z + ½I + A + ½I + Z…
Единицей строения и функционирования миофибрилл является саркомер — участок между двумя Z-дисками. Величина саркомера в расслабленном состоянии всегда одинакова (1,8—2,8 мкм в зависимости от вида животного). Электронной микроскопией выявлено, что миофибрилла состоит из ряда более тонких нитей – протофибрилл. Их диаметр в разных частях саркомера разный. В I-дисках встречаются тонкие нити, а в A-дисках кроме тонких присутствуют толстые нити (диаметром порядка 16 нм). Все протофибриллы расположены параллельно и одна в другую не переходят. Тонкие нити – актиновые, а толстые – миозиновые. В состав последних входит около 300 молекул белка миозина. Нити актина и миозина могут скользить одна относительно другой, в результате чего происходит сокращение или расслабление саркомеров. Суммарное сокращение саркомеров приводит к сокращению мышечного волокна.
Скольжение нитей актина и миозина осуществляется с затратой большего количества АТФ и в присутствии ионов Са2+. Периодическое скольжение нитей в одну и другую стороны обеспечивает сокращение и расслабление миофибрилл и мышц в целом и, таким образом, обеспечивает работу мышц.
jbio.ru
| Мембраны – динамические, подвижные структуры, которые постоянно изменяют свою форму и площадь. На подвижности мембран основана концепция эндоплазматической системы. Согласно этой концепции, внутренние мембраны цитоплазмы, кроме мембран митохондрий и пластид, представляют собой единое целое и берут начало от эндоплазматического ретикулума. Новые цистерны диктиосом образуются из эндоплазматического ретикулума через стадию промежуточных пузырьков, а секреторные пузырьки, отделяющиеся от диктиосом, в конечном итоге способствуют формированию плазматической мембраны. Таким образом, эндоплазматический ретикулум и диктиосомы образуют функциональное целое, в котором диктиосомы играют роль промежуточных структур в процессе преобразования мембран, подобных эндоплазматическому ретикулуму, в мембраны, подобные плазматической. В тканях, клетки которых слабо растут и делятся, постоянно происходит обновление мембранных компонентов. Микротрубочки обнаружены практически во всех эукариотических клетках. Представляют собой цилиндрические структуры диаметром около 24 нм. Длина их варьирует. Каждая трубочка состоит из субъединиц белка, называемого тубулином. Субъединицы образуют 13 продольных нитей, окружающих центральную полость. Микротрубочки – это динамические структуры, они регулярно разрушаются и образуются на определенных стадиях клеточного цикла. Их сборка происходит в особых местах, которые называются центрами организации микротрубочек. В растительных клетках они имеют слабовыраженную аморфную структуру. Функции микротрубочек: участвуют в образовании клеточной оболочки; направляют пузырьки диктиосом к формирующейся оболочке, подобно нитям веретена, которые образуются в делящейся клетке; играют определенную роль в формировании клеточной пластинки (первоначальной границы между дочерними клетками). Кроме того, микротрубочки – важный компонент жгутиков и ресничек, в движении которых, играют немаловажную роль. Микрофиламенты, подобно микротрубочкам, найдены практически во всех эукариотических клетках. Представляют собой длинные нити толщиной 5 – 7 нм, состоящие из сократительного белка актина. Пучки микрофиламентов встречаются во многих клетках высших растений. По-видимому, играют важную роль в токах цитоплазмы. Микрофиламенты вместе с микротрубочками образуют гибкую сеть, называемую цитоскелетом. Основное вещество довольно долго считали гомогенным (однородный) богатым белком раствором с малым количеством структур или вообще бесструктурным. Однако в настоящее время, используя высоковольтный электронный микроскоп, было установлено, что основное вещество представляет трехмерную решетку, построенную из тонких (диаметром 3 – 6 нм) тяжей, заполняющих всю клетку. Другие компоненты цитоплазмы, включая микротрубочки и микрофиламенты, подвешены к этой микротрабекулярной решетке. Микротрабекулярная структура представляет собой решетку из белковых тяжей, пространство между которыми заполнено водой. Вместе с водой решетка имеет консистенцию геля, гель имеет вид студенистых тел. К микротрабекулярной решетке прикреплены органеллы. Решетка осуществляет связь между отдельными частями клетки и направляет внутриклеточный транспорт. Липидные капли – структуры сферической формы, придающие гранулярность цитоплазме растительной клетки под световым микроскопом. На электронных микрофотографиях они выглядят аморфными. Очень похожие, но более мелкие капли встречаются в пластидах. Липидные капли, принимая за органеллы, называли их сферосомами и считали, что они окружены одно- или двуслойной мембраной. Однако последние данные показывают, что у липидных капель мембран нет, но они могут быть покрыты белком. Эргастические вещества – это «пассивные продукты» протопласта: запасные вещества или отходы. Они могут появляться и исчезать в разные периоды клеточного цикла. Кроме зерен крахмала, кристаллов, антоциановых пигментов и липидных капель. К ним относятся смолы, камеди, танины и белковые вещества. Эргастические вещества входят в состав клеточной оболочки, основного вещества цитоплазмы и органелл, в том числе вакуолей. Жгутики и реснички – это тонкие, похожие на волоски структуры, которые отходят от поверхности многих эукариотических клеток. Имеют постоянный диаметр, но длина колеблется от 2 до 150 мкм. Условно более длинные и немногочисленные из них называют жгутиками, а более короткие и многочисленные - ресничками. Четких различий между этими двумя типами структур не существует, поэтому для обозначения обоих используют термин жгутик. У некоторых водорослей и грибов жгутики являются локомоторными органами, с помощью которых они передвигаются в воде. У растений (например, мхов, печеночников, папоротников, некоторых голосеменных) только половые клетки (гаметы) имеют жгутики. Каждый жгутик имеет определенную организацию. Наружное кольцо из 9 пар микротрубочек окружает две дополнительные микротрубочки, расположенные в центре жгутика. Содержащие ферменты «ручки» отходят от одной микротрубочки каждой из наружных пар. Это основная схема организации 9 + 2 обнаружена во всех жгутиках эукариотических организмов. Считают, что движение жгутиков основано на скольжении микротрубочек, при этом наружные пары микротрубочек движутся одна вдоль другой без сокращения. Скольжение пар микротрубочек относительно друг друга вызывает локальное изгибание жгутика. Жгутики «вырастают» из цитоплазматических цилиндрических структур, называемых базальными тельцами, образующимися и базальную часть жгутика. Базальные тельца имеют внутреннее строение, напоминающее строение жгутика, за исключением того, что наружные трубочки собраны в тройки, а не в пары, а центральные трубочки отсутствуют. Клеточная стенка. Клеточная стенка отграничивает размер протопласта и предохраняет его разрыв за счет поглощения воды вакуолью. Клеточная стенка имеет специфические функции, которые важны не только для клетки и ткани, в которой клетка находится, но и для всего растения. Клеточные стенки играют существенную роль в поглощении, транспорте и выделении веществ, а, кроме того, в них может быть сосредоточена лизосомальная, или переваривающая активность. Компоненты клеточной стенки. Наиболее типичным компонентом клеточной стенки является целлюлоза, которая в значительной степени определяет её архитектуру. молекулы целлюлозы состоят из повторяющихся молекул глюкозы, соединенных конец к концу. Длинные тонкие молекулы целлюлозы объединены в микрофибриллы толщиной 10 – 25 нм. Микрофибриллы перевиваются и образуют тонкие нити, которые в свою очередь могут обматываться одна вокруг другой, как пряди в канате. Каждый такой «канат», или макрофибрилла, имеет толщину около 0,5 мкм, достигая в длину 4 мкм. Макрофибриллы прочны, как равная по величине стальная проволока. Целлюлозный каркас клеточной стенки заполнен переплетающимися с ним целлюлозными молекулами матрикса. В его состав входят полисахариды, называемые гемицеллюлозами, и пектиновые вещества, или пектины, химически очень близкие к гемицеллюлозам. Другой компонент клеточной стенки – лигнин – является самым распространенным после целлюлозы полимером растительных клеток. Лигнин увеличивает жесткость стенки и обычно содержится в клетках, выполняющих опорную или механическую, функцию. Кутин, суберин, воска – обычно откладываются в оболочках защитных тканей растений. Кутин, например, содержится в клеточных оболочках эпидермы, а суберин - вторичной защитной ткани, пробки. Оба вещества встречаются в комбинации с восками и предотвращают чрезмерную потерю воды растением. Слои клеточной стенки. Толщина стенки растительных клеток варьирует в широких пределах в зависимости от роли клеток в структуре растений и возраста самой клетки. Под электронным микроскопом просматривается в растительной клеточной стенке два слоя: срединная пластинка (называемая также межклеточным веществом), и первичной клеточной стенки. Многие клетки откладывают ещё один слой – вторичную клеточную стенку. Срединная пластинка располагается между первичными стенками соседних клеток. Вторичная стенка, если она есть, откладывается протопластом клетки на внутреннюю поверхность первичной клеточной стенки. Срединная пластинка. Срединная пластинка состоит в основном из пектиновых веществ. Там, где должна возникнуть клеточная стенка, между двумя вновь образующимися клетками, вначале отмечается густое сплетение из канальцев эндоплазматической сети и цистерны аппарата Гольджи (диктиосом). Затем в этом месте появляются пузырьки, заполнены пектиновым веществом (из полисахаридов). Пузырьки эти отделяются от цистерн аппарата Гольджи. Ранняя клеточная стенка содержит различные полисахариды, основные из которых пектины и гемицеллюлоза. Позже в её состав входят более плотные вещества – целлюлоза и лигнин. Первичная клеточная оболочка. Это слой целлюлозной оболочки, который откладывается до начала или во время роста клетки. Помимо целлюлозы, гемицеллюлоз и пектина первичные оболочки содержат гликопротеин. Первичные оболочки могут лигнифицироваться. Пектиновый компонент придаёт пластичность, которая позволяет первичной оболочке, растягивается по мере удлинения корня, стебля или листа. Активно делящиеся клетки (большинство зрелых клеток, вовлеченных в процессы фотосинтеза, дыхания и секреции) имеют первичные оболочки. Такие клетки с первичной оболочкой и живым протопластом способны утрачивать характерную форму, делиться и дифференцироваться в новый тип клеток. Именно они участвуют в заживлении ран и регенерации тканей у растений. Первичные клеточные оболочки не одинаковы по толщине на всем своем протяжении, а имеют тонкие участки, которые называются первичными поровыми полями. Тяжи цитоплазмы, или плазмодесмы, соединяющие протопласты соседних клеток, обычно проходят через первичные поровые поля. Вторичная клеточная оболочка. Несмотря на то, что многие растительные клетки имеют только первичную оболочку, у некоторых к центру клетки протопласт откладывает вторичную оболочку. Обычно это происходит после прекращения роста клетки и площадь первичной оболочки более не увеличивается. По этой причине вторичная оболочка отличается от первичной. Вторичные оболочки особенно нужны специализированным клеткам, укрепляющим растение и проводящим воду. После отложения вторичной оболочки протопласт этих клеток, как правило, отмирает. Во вторичных оболочках больше целлюлозы, чем в первичных, а пектиновые вещества и гликопротеины в них отсутствуют. Вторичная оболочка растягивается с трудом, ее матрикс состоит из гемицеллюлозы. Во вторичной оболочке можно выделить три слоя – наружный, средний и внутренний (S1, S2, S3). Слоистая структура вторичных оболочек значительно увеличивает их прочность. Микрофибриллы целлюлозы во вторичной оболочке откладывается плотнее, чем в первичной. Лигнин – обычный компонент вторичных оболочек древесины. Поры в оболочках контактирующих клеток расположены напротив друг друга. Две лежащие друг против друга поры и поровая мембрана образуют пару пор. В клетках, имеющих вторичные оболочки, существуют два основных типа пор: простые и окаймленные. В окаймленных порах вторичная оболочка нависает над полостью поры. В простых порах этого нет. Рост клеточной оболочки. По мере роста клетки увеличивается толщина и площадь клеточной оболочки. Растяжение оболочки – процесс сложный. Он контролируется протопластом и регулируется гормоном ауксином. В клетках, растущих во всех направлениях равномерно, отложение миофибрилл носит случайный характер. Эти миофибриллы образуют неправильную сеть. Такие клетки обнаружены в сердцевине стебля, запасающих тканях и при культивировании клеток in vitro. В удлиняющихся клетках миофибриллы боковых оболочек откладывается под прямым углом к оси удлинения. Вещества матрикса – пектины, гемицеллюлозы и гликопротеины переносятся к оболочке в пузырьках диктиосом. При этом пектины более характерны для растущих клеток, а гемицеллюлозы преобладают в не растущих клетках. Целлюлозные микрофибриллы синтезируются на поверхности клетки с помощью ферментного комплекса, связанного с плазматической мембраной. Ориентация микрофибрилл контролируется микротрубочками, расположенными у внутренней поверхности плазматической мембраны. Плазмодесмы. Это тонкие нити цитоплазмы, которые связывают между собой протопласты соседних клеток. Плазмодесмы либо проходят сквозь клеточную оболочку в любом месте, либо сосредоточены на первичных поровых полях или в мембранах между парами пор. Под электронным микроскопом плазмодесмы выглядят как узкие каналы, выстланные плазматической мембранной. По оси канала из одной клетки в другую тянется цилиндрическая трубочка меньшего размера – десмотрубочка, которая сообщается с эндоплазматическим ретикулумом обеих смежных клеток. Многие плазмодесмы формируются во время клеточного деления, когда трубчатый эндоплазматический ретикулум захватывается развивающейся клеточной пластинкой. Плазмодесмы могут образовываться и в оболочках неделящихся клеток. Эти структуры обеспечивают эффективный перенос некоторых веществ от клетки к клетке. Деление клеток. У многоклеточных организмов деление клеток наряду с увеличением их размеров является способом роста всего организма. Новые клетки, образовавшиеся во время деления, сходны по структуре и функциям, как с родительской клеткой, так и между собой. Процесс деления у эукариот можно подразделить на две частично перекрывающиеся стадии: митоз и цитокинез. Митоз – это образование из одного ядра двух дочерних ядер, морфологически и генетически эквивалентных друг другу. Цитокинез – это деление цитоплазматической части клетки с образованием дочерних клеток. Клеточный цикл. Живая клетка проходи ряд последовательных событий, составляющих клеточный цикл. Продолжительность самого цикла варьирует в зависимости от типа клетки и внешних факторов, например от температуры или обеспеченности питательными веществами. Обычно цикл делится на интерфазу и четыре фазы митоза. Интерфаза. Период между последовательными митотическими делениями. Интерфазу делят на три периода, обозначаемые как G1, S, G2. В период G1, который начинается после митоза. В этот период увеличивается количество цитоплазмы, включая различные органеллы. Кроме того, согласно современной гипотезе, в период G1 синтезируются вещества, которые либо стимулируют, либо ингибируют период S и остальную часть цикла, определяя, таким образом, процесс деления. В период S следует за периодом G1, в это время происходит удвоение генетического материала (ДНК). В период G2, который следует за S, формируются структуры, непосредственно участвующие в митозе, например, компоненты веретена. Некоторые клетки проходит неограниченный ряд клеточных циклов. Это одноклеточные организмы и некоторые клетки зон активного роста (меристем). Некоторые специализированные клетки после созревания теряет способность к размножению. Третья группа клеток, например образующих раневую ткань (каллус), сохраняет способность делиться только в специальных условиях. Митоз, или деление ядра. Это непрерывный процесс, подразделяемый на четыре фазы: профазу, метафазу, анафазу, телофазу. В результате митоза генетический материал, удвоившийся в интерфазе, делится поровну между двумя дочерними ядрами. Одним из самых ранних признаков перехода клетки к делению служит появление узкого, кольцеобразного пояска из микротрубочек непосредственно под плазматической мембраной. Это относительно плотный поясок окружает ядро в экваториальной плоскости будущего митотического веретена. Так как он проявляется перед профазой, его называют препрофазным пояском. Он исчезает после митотического веретена, задолго до появления в поздней телофазе клеточной пластинки, которая растет от центра к периферии и сливается с оболочкой материнской клетки в области, ранее занятой препрофазным пояском. Профаза. В начале профазы хромосомы напоминают длинные нити, разбросанные внутри ядра. Затем, по мере того как нити укорачиваются и утолщаются, можно увидеть, что каждая хромосома состоит не из одной, а из двух переплетенных нитей, называемых хроматидами. В поздней профазе две укороченные спаренные хроматиды каждой хромосомы лежат рядом параллельно, соединённые узким участком, называемым центромерой. Она имеет определённое положение на каждой хромосоме и делит хромосому на два плеча различной длины. Микротрубочки располагаются параллельно поверхности ядра вдоль оси веретена. Это само раннее проявление сборки митотического веретена. К концу профазы ядрышко постепенно теряет чёткие очертания и наконец исчезает. Вскоре после этого распадается и ядерная оболочка. продолжение |
www.coolreferat.com





