
Зависимость роста растений от экзогенных факторов. Реакция растений на температуру
Актиноритмическая реакция растений на низкую температуру
Актиноритмические условия выращивания влияют и на такие важные практические признаки, как морозоустойчивость и зимостойкость многолетних растений.
Известно, что многие древесные виды у себя на родине выдерживают значительные морозы, доходящие часто до —25 °С, и в то же время, если их перенести в более высокие широты, они начинают вымерзать при менее низких температурах. Например, многие маньчжурские и вообще дальневосточные виды, хорошо переносящие зимы в пределах своих естественных ареалов, сильно страдают от морозов на широте Ленинграда, хотя здесь зимы менее суровы. То же самое можно сказать об акации белой, хорошо зимующей под Харьковом и вымерзающей под Ленинградом, и т. д.
Южные формы акации белой, грецкого ореха, абрикоса, миндаля и винограда, выращенные при естественных условиях на широте Ленинграда и оставленные на зиму без укрытия, полностью вымерзали. Сеянцы же образцов, росших в течение лета при укороченных до определенного периода днях, становились зимоустойчивыми и не вымерзали. Более того, в большинстве случаев сеянцы и саженцы южных древесных видов, выращенные при коротких днях, не только не вымерзали, но и вообще не страдали от перезимовки в открытом грунте. Весной их рост начинался из вершинных почек, так же, как и у себя на родине.
Растения акации белой после выращивания при 14-часовом дне совершенно не страдали от перезимовки, в то время как контрольные растения вымерзали полностью — от верхней почки до конца корневой системы.
Хинное дерево (Cinchona succirubra), дающее наибольшие приросты при 10-часовом дне, в этих же условиях оказывается наиболее холодоустойчивым, перенося —1,8 °С, тогда как при других актиноритмах, как с более длинным, так и с более коротким днями, когда его приросты бывают меньшими, оно погибает при температуре близкой к 0 °С.
У лимона лучшие приросты отмечены при 12-часовом дне. При выращивании в таких же условиях растение было наиболее холодостойким и переносило —7—9 °С почти без повреждений.
Однако и среди вечнозеленых субтропических видов есть такие, которые оказываются наиболее морозоустойчивыми при длине дня, меньшей по сравнению с той, что определяет условия для их максимальных приростов. Так, апельсин (сеянец Швецова), дающий наибольшие приросты при 15—16-часовом дне, в таких условиях далеко не морозостоек, но вполне устойчив, когда его выращивают при 8-часовом дне.
С другой стороны, среди листопадных видов есть такие, которые, подобно лимону и хинному дереву, обладают наибольшей морозоустойчивостью при тех же актиноритмических условиях, которые способствуют их максимальному росту. Примером таких листопадных видов может служить миндаль обыкновенный. Опыт с этой культурой проводили в Пушкине. Проросшие семена миндаля весной посадили по одному в горшки диаметром 15 см. Повторность в группе — 12 сосудов.
С момента всходов растения миндаля выращивали при 5-, 6-, 7-, 8-, 9-, 10-, 11-, 12-, 13-, 14-, 15-, 19-часовом дне и при непрерывном освещении. При всех указанных актиноритмических условиях рост сеянцев миндаля окончился почти одновременно на 80—85-е сутки. Наибольшие приросты стеблей были в группах при 13-, 14- и 15-часовом дне, но и они мало отличались от приростов других групп, так как нивелировались небольшим объемом горшков, в которых они находились. Выращивали миндаль в горшках для того, чтобы иметь возможность проморозить растения в холодильных шкафах, но этого сделать не удалось. Поэтому все растения были оставлены на зимовку просто в грядках участка.
В начале лета, после отрастания сеянцев, при учете их перезимовки была установлена совершенно правильная зависимость зимостойкости миндаля от актиноритмических условий предшествующей вегетации. Сеянцы миндаля, выращенные при актиноритмах с длиной дня, равной 5, 6, 7, 15, 16, 17, 19 и 24 ч, вымерзли полностью с корнями.
Не погибло зимой ни одного растения в группах, выращиваемых при длине дня 8, 9, 10, 11, 12, 13 и 14 ч. Только у некоторых растений в различной степени подмерзли верхние части побегов, причем меньше всего пострадали сеянцы, выращиваемые при 13-часовом дне, у некоторых особей их сохранились даже верхушечные почки. Можно сказать, что оптимальными актиноритмическими условиями для прохождения процессов, определяющих высокую морозоустойчивость миндаля, являются такие, где длина дня не менее 8 и не более 14 ч или, наоборот, где ночь не более 16 и не менее 10 ч.
Как избыток, так и недостаток ежесуточной темноты привели к одному и тому же отрицательному эффекту — вымерзанию сеянцев миндаля. Но можно сказать и так, что недобор ежесуточного освещения (7-часовой день и ниже), так же как и чрезмерность его (15-часовой день и выше), действуют одинаково отрицательно. Только оптимальные суточные сочетания света и темноты способствуют прохождению физиологических процессов, определяющих морозоустойчивость данного вида.
В нашем примере с морозоустойчивостью миндаля очень отчетливо выявились критические актиноритмические условия, лежащие: нижний — между 7- и 8-часовым днем и верхний — между 14- и 15-часовым днем.
Актиноритмическая реакция листьев растений путем изменения морозоустойчивости проливает свет на ряд ранее непонятных фактов. В частности, она объясняет широко распространенное явление хорошей перезимовки многих видов при весьма суровых зимах в одних районах и вымерзание их в других пунктах с более мягкими зимами. Данное явление наблюдается только в тех случаях, когда упомянутые районы находятся в различных географических широтах. Причина его — различие в длине дней — более коротких в южных широтах, чем в северных.
При переносе культуры какого-нибудь относительно южного вида в северные районы фактором, отбирающим наиболее морозоустойчивые формы, являются условия не зимы, а лета, т. е. актиноритмы. Нам удалось доказать это прямыми опытами. Вот один из них. Под Москвой при посеве большого числа образцов акации белой сохранилось несколько десятков растений, дошедших до цветения и плодоношения. Естественно, что плодоносили только наиболее морозоустойчивые формы. Когда семена, собранные с одного из плодоносивших растений, были высеяны параллельно с семенами исходной южной популяции в различных актиноритмических условиях, то при коротком дне (10-часовом) значительно больше угнетался рост сеянцев московского образца, чем южного.
Совершенно ясно, что плодоносящие под Москвой экземпляры акации белой явились продуктом отбора длинных московских дней, и поэтому прирост их сеянцев сильно снижался при коротком дне, чего не наблюдалось у подавляющего большинства растений акации в исходном образце южного происхождения.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Действие на растения высоких температур
Реакция растений на высокую температуру тесно связана с состоянием их обводненности и включает изменение многих обменных процессов. В онтогенезе растений отмечаются периоды,
более чувствительные к высоким температурам и водному дефициту. Как правило, они совпадают с периодом наиболее интенсивного роста и формирования гамет.
Перегрев может вызывать водный дефицит и характерные для него изменения у растений, но высокая температура и сама по себе приводит к гибели из-за повреждения мембран, инактивации и денатурации белков. При перегреве в растении обычно падает активность биосинтеза и усиливаются гидролитические процессы. Даже если из строя выходят немногие термолабильные ферменты, то это ведет к гибели клеток от расстройства обмена нуклеиновых кислот и белков. Растворимые азотистые соединения накапливаются в таких концентрациях, что диффундируют из клеток. Образуются ядовитые продукты распада, которые больше не обезвреживаются: например, могут гидролизоваться белки до аммиака, вызывающего сильное отравление клеток и гибель растений (П.А.Генкель, 1982). Но при этом у жаростойких видов увеличивается также содержание органических кислот, связывающих избыточный аммиак.
Характерно, что большинство растений повреждается температурами немного выше 30 °С, хотя ферменты и органеллы, выделенные из растений, при таких температурах обычно не повреждаются. Вероятно, при перегреве быстро распадаются белковолипидные комплексы и изменяется структура мембран. Чувствительность мембран клеток и органелл к изменениям температуры объясняется плавлением или затвердеванием жирных кислот в их фосфолипидах. Известно, что при высоких температурах растения синтезируют больше насыщенных жиров с более высокой температурой плавления (Г. Косулина и др., 1993).
Другая причина повреждения клеток при перегреве состоит в том, что при повышенных температурах некоторые биологически активные вещества — необходимые для роста витамины, аминокислоты, гормоны — быстро разрушаются или не образуются в нужном количестве. Поэтому, зная химическую природу индуцированного температурой замедления роста, можно улучшать состояние растений внесением нужных веществ (А.Гэлстон и др., 1983).
eco-rasteniya.ru
Действие температурного стресса на растения
Приспособленность онтогенеза растений к условиям среды является результатом их эволюционного развития (изменчивости, наследственности, отбора). На протяжении филогенеза каждого вида растений в процессе эволюции выработались определенные потребности индивидуума к условиям существования и приспособленность к занимаемой им экологической нише. Жароустойчивость, холодоустойчивость и другие экологические особенности конкретных видов растений сформировались в ходе эволюции в результате длительного действия соответствующих условий. Так, теплолюбивые растения и растения короткого дня характерны для южных широт, менее требовательные к теплу и растения длинного дня - для северных. В природе в одном географическом регионе каждый вид растений занимает экологическую нишу, соответствующую его биологическим особенностям. Наследственность растений формируется под влиянием определенных условий внешней среды. Важное значение имеют и внешние условия онтогенеза растений.
В большинстве случаев растения и посевы (посадки) сельскохозяйственных культур, испытывая действие тех или иных неблагоприятных факторов, проявляют устойчивость к ним как результат приспособления к условиям существования, сложившимся исторически, что отмечал еще К. А. Тимирязев. Все физиологические и биохимические процессы идут лишь в определенных температурных границах, которые обычно лежат в довольно узких пределах. Фактор тепла имеет большое значение и в географическом распределении растений. Составляя существенную часть климатических условий, он тем самым определяет северные и южные границы ареалов, зональную структуру растительного покрова.
Действие температурного стресса на растения
Жара и мороз вредят жизненным функциям и ограничивают распространение вида в зависимости от их интенсивности, продолжительности и периодичности, но прежде всего от состояния активности и степени закалки растений. Стресс — это всегда необычная нагрузка, которая не обязательно должна быть опасной для жизни, но которая непременно вызывает в организме «реакцию тревоги», если только он не находится в выраженном состоянии оцепенения. Покоящиеся стадии, такие, как сухие споры, а также пойкилогидрические растения в высохшем состоянии, нечувствительны, так что они могут пережить без повреждения любую отмеченную на Земле температуру.
Протоплазма вначале отвечает на стресс резким усилением метаболизма. Повышение интенсивности дыхания, которое наблюдается в качестве стрессовой реакции отражает попытку исправить уже появившиеся дефекты и создать ультраструктурные предпосылки для приспособления к новой ситуации. Стрессовая реакция — это борьба механизмов адаптации, с деструктивными процессами в протоплазме, ведущими к ее гибели.
Гибель клеток от перегрева и холода. Если температура переходит критическую точку, клеточные структуры и функции могут повреждаться так внезапно, что протоплазма тотчас же отмирает. В природе такое внезапное разрушение нередко происходит при эпизодических морозах, например при поздних заморозках весной. Но повреждения могут возникать и постепенно; отдельные жизненные функции выводятся из равновесия и угнетаются, пока, наконец, клетка не отомрет в результате прекращения жизненно важных процессов.
Картина повреждения
Различные жизненные процессы неодинаково чувствительны к температуре. Сначала прекращается движение протоплазмы, интенсивность которого непосредственно зависит от энергоснабжения за счет процессов дыхания и от наличия высокоэнергетических фосфатов. Затем снижаются фотосинтез и дыхание. Для фотосинтеза особенно опасна жара, дыхание же наиболее чувствительно к холоду. У поврежденных холодом или жарой растений после возвращения в умеренные условия уровень дыхания сильно колеблется и часто бывает ненормально повышен. Повреждение хлоропластов ведет к длительному или необратимому угнетению фотосинтеза. В конечной стадии утрачивается полупроницаемость биомембран, разрушаются клеточные компартменты, особенно тилакоиды пластид, и клеточный сок выходит в межклетники.
Причины гибели при перегреве
Высокая температура быстро приводит к гибели из-за повреждения мембран и прежде всего в результате инактивации и денатурации белков. Даже если из строя выходят только немногие, особо термолабильные ферменты, это ведет к расстройству обмена нуклеиновых кислот и белков и в конце концов — тоже к гибели клеток. Растворимые азотистые соединения накапливаются при этом в таких больших концентрациях, что они диффундируют из клеток и теряются; кроме того, образуются ядовитые продукты распада, которые не могут больше обезвреживаться в ходе обмена веществ.
Гибель от охлаждения и от мороза
При повреждении протоплазмы холодом следует различать, вызвано ли оно самой по себе низкой температурой или же замерзанием. Некоторые растения тропического происхождения повреждаются уже при снижении температуры до нескольких градусов выше нуля. Подобно гибели от перегрева, гибель от охлаждения тоже бывает прежде всего связана с дезорганизацией обмена нуклеиновых кислот и белков, но здесь играют роль также нарушения, проницаемости и прекращение тока ассимилятов.
Растения, которым охлаждение до температур выше нуля не причиняет вреда, повреждаются только при температурах ниже нуля, т. е. в результате образования льда в тканях. Богатые водой, незакаленные протопласты могут легко замерзать; при этом внутри клетки мгновенно образуются ледяные кристаллики, и клетка погибает. Чаще всего лед образуется не в протопластах, а в межклетниках и клеточных стенках. Такое образование льда называют внеклеточным. Выкристаллизовавшийся лед действует как сухой воздух, так как упругость пара надо льдом ниже, чем над переохлажденным раствором. В результате от протопластов отнимается вода, они сильно сжимаются (на 2/з своего объема) и концентрация растворенных веществ в них возрастает. Перемещение воды и замерзание продолжаются до тех пор, пока в протоплазме не установится равновесие сосущих сил между льдом и водой.
Положение равновесия зависит от температуры; при температуре —5°С равновесие наступает примерно при; 60 бар, а при —10°С — уже при 120 бар. Таким образом, низкие температуры действуют на протоплазму так же, как высыхание. Морозоустойчивость клетки более высока, если вода прочно связана со структурами протоплазмы и осмотически связана. При обезвоживании цитоплазмы (безразлично, в результате засухи или замерзания) инактивируются ферментные системы, ассоциированные с мембранами, - системы, участвующие главным образом в синтезе АТФ и в процессах фосфорилирования. Инактивацию, вызывают чрезмерные и потому токсичные концентрации ионов солей и органических кислот в незамерзшем остаточном растворе. Напротив, сахара, производные сахаров, определенные аминокислоты и белки защищают биомембраны и ферменты от вредных веществ. Наряду с этим есть указания на то, что при замерзании белки денатурируются, что также ведет к повреждению мембран.
Термоустойчивость
Термоустойчивость — это способность организма переносить большую жару или холод без необратимого повреждения. Термоустойчивость растения складывается из способности протоплазмы переносить экстремальные температуры (толерантность по Дж. Левитту) и из эффективности мер, которые замедляют или предотвращают развитие повреждений (избегание).
Меры, позволяющие избегать повреждения
Возможные способы защиты клеток от температурного повреждения немногочисленны и не очень эффективны. Изоляция от перегрева и охлаждения может дать лишь кратковременную защиту. Так. например, в густых кронах деревьев или у подушечных растений почки листья и цветки, находящиеся в глубине и ближе к земле, меньше подвергаются опасности замерзнуть в результате отдачи тепла путем излучения, чем наружные части растения. Виды хвойных с особенно толстой коркой лучше выдерживают пожары в подлеске. Общее же значение имеют главным образом две защитные меры: замедление образования льда в тканях и (при жаре) охлаждение путем отражения падающих лучей и с помощью транспирации.
Устойчивость протоплазмы
Длительное и регулярно повторяющееся воздействие крайних. температур растения могут выдерживать только в том случае, если сама протоплазма жаро- или морозоустойчива. Эта особенность обусловлена генетически и поэтому у разных видов и даже сортов выражена в разной степени. Однако это не такое свойство, которое присуще растению постоянно и всегда в одинаковой мере. Проростки, весенние побеги древесных растений в период их интенсивного растяжения, культуры микроорганизмов в фазе экспоненциального роста вряд ли способны закаливаться и поэтому чрезвычайно чувствительны к температуре.
Устойчивость к образованию льда и закаливание к действию мороза
В областях с сезонным климатом наземные растения приобретают осенью «льдоустойчивость», т. е. способность переносить образование льда в тканях. Весной, с распусканием почек они снова утрачивают эту способность, и теперь замерзание приводит к их вымерзанию. Таким образом, холодостойкость многолетних растений вне тропиков регулярно колеблется в течение года между минимальной величиной в период вегетации и максимумом в зимнее время. Льдоустойчивость формируется постепенно осенью. Первая предпосылка для этого—переход растения в состояние готовности к закаливанию, наступающее только тогда, когда заканчивается рост. Если готовность к закаливанию достигнута, то процесс закаливания может начинаться.
Этот процесс состоит из нескольких фаз, каждая из которых подготавливает переход к следующей. По теории, разработанной И.И. Тумановым, закаливание к морозу, у озимых злаков и плодовых; деревьев (эти растения изучались наиболее основательно) начинается многодневным (до нескольких недель) воздействием температур чуть выше нуля. На этой фазе, предшествующей закаливанию, в протоплазме накапливаются сахара и другие защитные вещества, клетки становятся беднее водой, а центральная вакуоль распадается на множество мелких вакуолей. Благодаря этому протоплазма оказывается подготовленной к следующей фазе, проходящей при регулярных слабых морозах от —3 до—5°С. При этом ультраструктуры и ферменты протоплазмы перестраиваются таким образом, что клетки переносят обезвоживание, связанное с образованием льда. Только после этого растения могут, не подвергаясь, опасности, вступать в заключительную фазу процесса; закаливания, которая при непрерывном морозе по меньшей мере от —10 до—15 °С делает протоплазму в высшей степени морозоустойчивой.
Зоны эффективных температур для разных видов различны. Готовые к закаливанию сеянцы березы, которые до начала процесса закаливания вымерзали бы при температуре от —15-до —20°С, переносят после окончания первой фазы закаливания; уже —35 °С, а при полной закалке они выдерживают даже охлаждение до —195°С. Таким образом, холод сам по себе стимулирует процесс закаливания. Если мороз ослабевает, то протоплазма опять переходит в первую фазу закаливания, однако устойчивость, снова может быть поднята холодными периодами до наивысшего уровня, пока растения остаются в состоянии покоя.
.В зимний период на сезонный ход морозостойкости накладываются кратковременные (индуцированные), адаптации, благодаря которым уровень устойчивости быстро приспосабливается к изменениям погоды. Холод больше всего способствует закаливанию в начале зимы. В это время устойчивость может за несколько дней подняться до наивысшего уровня. Оттепель, особенно в конце зимы, вызывает быстрое снижение устойчивости растений, но в середине зимы после выдерживания в течение нескольких дней при температуре от +10 до +20 °С растения в значительной мере теряют закалку. Способность изменять морозоустойчивость под влиянием холода и тепла, т. е. диапазон индуцируемых адаптации устойчивости, является конституционным признаком отдельных видов растений.
После окончания зимнего покоя способность к закаливанию и вместе с тем высокая степень закалки быстро утрачиваются. Весной существует тесная связь между активированием распускания почек и ходом изменения устойчивости.
biofile.ru
Влияние температуры на рост растений
Рост растении возможен в сравнительно широком диапазоне температур и определяется географическим происхождением данного вида. Требования растения к температуре меняются с возрастом, различны у отдельных органов растения (листья, корни, плодоэлементы и др.). Для роста большинства сельскохозяйственных растений России нижняя температурная граница соответствует температуре замерзания клеточного сока (около -1...-3 °С), а верхняя - коагуляции белков протоплазмы (около 60 "С). Вспомним, что температура влияет на биохимические процессы дыхания, фотосинтеза и других метаболических систем растений, а графики зависимости роста растений и активности ферментов от температуры близки по форме (колоколообразная кривая).
Температурные оптимумы для роста. Для появления всходов требуется более высокая температура, чем для прорастания семян (табл. 22).
22. Потребность семян полевых культур в биологически минимальных температурах (по В. Н. Степанову)
Температура, "С
прорастання семян 1 появления всходов
Горчица, конопля, рыжик 0-1 2-3
Рожь, пшеница, ячмень, овес, 1-2 4-5
горох, вика, чечевица, чина
Лен, гречиха, люпин, бобы, 3-4 5-6
нуг, свекла, сафлор
Подсолнечник, перилла 5-6 7-8
Кукуруза, просо, соя 8-10 10-11
Фасоль, клещевина, сорго 10-12 12-15
Х-волчатник, рис, кунжут 12-14 14-15
При анализе роста растений выделяют три кардинальные температурные точки: минимальную (рост только начинается), оптимальную (наиболее благоприятная для роста) и максимальную температуру (рост прекращается).
Различают растения тешолюбивые- с минимальными температурами для роста более 10 "С и оптимальными 30-35 "С (кукуруза, огурец, дыня, тыква), холодостойкие - с минимальными температурами для роста в пределам 0-5 "С н оптимальными 25-31 "С. Максимальные температуры для большинства растений 37-44 "С, для южных 44-50 "С. При увеличении температуры на 10 °С в зоне оптимальных значений скорость роста увеличивается в 2-3 раза. Повышение температуры выше оптимальной замедляет рост и сокращает его период. Оптимальная температура для роста корневых систем ниже, чем для надземных органов. Оптимум для роста выше, чем для фотосинтеза.
Можно предположить, что при высокой температуре имеет место недостаток АТФ и НАДФН, необходимых для восстановительных процессов, что вызывает торможение роста. Температура, оптимальная для роста, может быть неблагоприятной для развития растения. Оптимум для роста меняется на протяжении вегетационного периода и в течение суток, что объясняется закрепленной в геноме растений потребностью к смене температур, имевшей место на исторической родине растений. Многие растения интенсивнее растут в ночной период суток.
Термопериодизм. Росту многих растений благоприятствуем смена температуры в течение суток: днем повышенная, а ночью пониженная. Так, для растений томата оптимальная температур_) днем 26 "С, а ночью 17-19 _С. Это явление Ф. Вент (1957) назвал термопериоднзмом. Термопериодии! - реакция растение) на периодическую смену повышенных и пониженных температур, выражающаяся в изменении процессов роста и развитие! (М. *. Чайлахян, 1982). Различают суточный и сезонный термопериоднзм. Для тропических растений разница между дневными и ночными температурами составляет 3-6 °С, для растений умеренного пояса - 5-7 "С. Это важно учитывать при выращивании растений в поле, теплицах и фитотронах, районировании культур и сортов сельскохозяйственных растений.
Чередование высоких и низких температур служит регулятора?__ внутренних часов растений, как п фотопе1_иодизм. Относительно низкие ночные температуры повышают унижай картофеля (Ф. Вент. 1959), сахаристость корнеплолок сахарной свеклы, ускоряют рост корневой системы н боковых побегов * растений томата (Н. И. Якушкмна, 1980). Низкие температуры, возможно, повышают активность ферментов, осуществляющих гидролиз) крахмала в листьях, а образующиеся растворимые формы углеводов передвигаются в корни н боковые побеги.
studfiles.net
Устойчивость растений к низким температурам
Под устойчивостью к низким температурам подразумевают холодостойкость и морозоустойчивость, т. е. способность растений выживать в условиях низких положительных температур и при заморозках. В листьях растений, повреждающихся при охлаждении, нарушаются процессы фотосинтеза, транспорта ассимилятов, дыхания, синтеза белков. Диапазон повреждающих температур сильно варьирует в зависимости от вида растения. Особенно устойчивы к низким температурам древесные виды в состоянии покоя. Семена, другие обезвоженные ткани растений и споры грибов способны переживать температуры около абсолютного нуля. Тургесцентные вегетирующие клетки также могут выдерживать замораживание, если их охлаждать очень быстро, поскольку формирующиеся в таких условиях кристаллы льда очень малы и не вызывают механических повреждений.
Холодостойкость
Большинство видов тропических и субтропических растений не выдерживают не только заморозков, но даже низких положительных температур. Такие растения, как кукуруза, фасоль, рис, томаты, огурцы и хлопок, останавливаются в росте при температуре 10–15 °С и повреждаются, если температура падает ниже 10 °С. Холодостойкость теплолюбивых растений можно также повысить предпосевным закаливанием. Для этого наклюнувшиеся семена (например, огурцов, томатов) в течение нескольких суток выдерживают в чередующихся (через 12 ч) условиях низких положительных (около 5 °С) и более высоких (10–20 °С) температур. Таким же образом можно закалять и рассаду. Обычное время для закаливания, например, картофеля составляет 15 дней обработки низкими положительными температурами. После такой закалки растения картофеля хорошо переносят даже слабые заморозки. В процессе закаливания изменяется гормональный баланс растения, ростовые процессы затормаживаются, возрастает количество ненасыщенных жирных кислот, включаются механизмы защиты клеток от обезвоживания и замерзания.
Основной причиной повреждающего действия низких положительных температур на теплолюбивые растения является нарушение функционирования клеточных мембран из-за их «затвердевания», связанного с фазовыми переходами жирных кислот, поскольку при достаточно низких температурах липидные бислои ведут себя, как твердые тела. При температуре выше фазового перехода структура бислоя сохраняется, однако при этом жирные кислоты «плавятся», в результате чего вращение и скручивание молекул происходит легче, чем при низких температурах. Затвердевание или «плавление» мембранных липидов зависит от состава содержащихся в них жирных кислот. Для насыщенных жирных кислот с длинной цепью характерны более высокие температуры фазовых переходов. Однако фазовые переходы в мембране индуцируются не только изменениями температуры. Они могут быть вызваны сдвигами рН и мембранного потенциала, двухвалентными катионами и гормонами.
Различная реакция устойчивых и неустойчивых растений на низкие температуры определяется в первую очередь различиями в составе жирных кислот, входящих в состав мембранных фосфолипидов. Выявлено, что у холодостойких растений содержание ненасыщенных жирных кислот (таких, как линоленовая и линолевая) гораздо выше, чем у растений, чувствительных к холоду. Увеличение количества ненасыщенных жирных кислот в составе мембран приводит к снижению температуры фазового перехода мембранных липидов.
У растений, чувствительных к холоду, в составе мембранных липидов велико содержание насыщенных жирных кислот (пальмитиновой, стеариновой). Мембраны такого типа стремятся к затвердеванию до квазикристаллического состояния уже при низких положительных температурах. При этом они становятся менее текучими, что нарушает функционирование многих белков-каналоформеров, переносчиков, рецепторов, ферментов и т. п. Схема влияния низких температур на мембраны от силы воздействия приведена на рис. 7.3.

Рис. 7.3. Схема влияния низких температур на мембраны
Итак, холодостойкими называются растения, которые не повреждаются и не снижают своей продуктивности при температуре от 0 до +10 оС.
Следует отметить, что холодостойкость свойственна и растениям умеренной зоны. Для большинства сельскохозяйственных культур низкие положительные температуры почти безвредны. Наиболее холодоустойчивыми являются яровые пшеница, ячмень, овес. Картофель, томаты, гречиха, просо, кукуруза, соя переносят температуры +5 оС и ниже без значительных повреждений. Фасоль, сорго, клещевина, арахис, рис, арбуз, дыня, тыква, огурцы, кабачки, перец – теплолюбивые растения, погибающие при температуре ниже +10 оС.
В ходе холодового воздействия синтезируются стрессовые белки холодового шока. Так, в проростках озимой пшеницы трехчасовая гипотермия (+3 оС) индуцирует синтез большого набора стрессовых низкомолекулярных и гидрофильных белков.
Морозоустойчивость
Заморозки вызывают повреждение клеток кристаллами льда, образующимися при температуре ниже 0 °С. Повреждающий эффект усугубляется также обезвоживанием тканей, которое при этом происходит. На первых этапах замерзания сильных повреждений растительных клеток не наблюдается, так как вначале кристаллы льда формируются в межклетниках и сосудах ксилемы. Если же заморозки длятся долго, вода переходит из цитоплазмы в апопласт, клетки обезвоживаются, кристаллы льда растут, и повреждают их.
Для предотвращения замерзания в растительных клетках и тканях функционирует система антифризов, представленная различными белками, углеводами и гликопротеинами. Криопротекторы, синтезирующиеся в растительных клетках при понижении температуры, могут предотвратить или резко замедлить рост кристаллов льда. Гидрофильные белки, моно- и олигосахариды, обладающие криопротекторным эффектом, способны связывать значительные количества воды. Связанная таким образом вода уже не замерзает и не транспортируется. Белки и углеводы, обладающие криопротекторным эффектом, способны стабилизировать другие белки и клеточные мембраны при дегидратации клеток, инициированной низкими температурами. Криопротекторы начинают синтезироваться, прежде всего, в эпидермисе и клетках, окружающих межклеточные полости, где происходит наиболее интенсивное образование кристаллов льда при замерзании тканей.
Важная роль в адаптации растений к низким температурам принадлежит фитогормону АБК. Устойчивость многих растений к заморозкам может быть повышена не только закаливанием, но также обработкой их АБК. При воздействии низких положительных температур и (или) при уменьшении светового периода концентрация этого гормона в растительных тканях резко возрастает. Мутанты арабидопсиса, нечувствительные к АБК (abi 1)или с нарушенным синтезом АБК (аbа1), не могут приспособиться к заморозкам и не закаливаются.
Активность многих генов и белков, которые экспрессируются при низкой температуре или водном дефиците, может быть индуцирована обработкой АБК. При изучении этих генов обнаружилось, что белки, появляющиеся при охлаждении тканей, имеют высокую степень гомологии с семейством белков RAB/LEA/DHN (responsive to ABA, late embryo abudant and dehydrin), синтезируются при созревании семян, водном дефиците и обработке растений АБК. Они очень гидрофильны и обладают необычным свойством: сохраняют свою структуру при резких колебаниях температуры. Продуктами генов LЕА являются гидрофильные белки, способные очень прочно удерживать воду. Считается, что именно эта группа белков обеспечивает защиту клетки при обезвоживании и заморозках, стабилизируя другие белки и клеточные мембраны.
У морозостойких растений при действии низких температур усиливается гидролиз крахмала и накопление в цитоплазме растворимых cахаров. У озимых форм пшеницы и многих других видов растений функцию криопротектора выполняет сахароза, накапливающаяся в больших концентрациях. В качестве антифризов могут также выступать и другие растворимые сахара: рафиноза, фруктозиды, сорбит или маннит, которые накапливаются в клеточных стенках растений и таким образом предотвращают образование кристаллов льда.
Наиболее чувствительны к морозу корни, корневища, клубни и луковицы большинства растений, которые повреждаются при температуре ниже –10 – –15°С. Покоящиеся почки древесных растений способны выдерживать морозы до –25 – –30 °С. У многолетних растений, устойчивых к заморозкам, в период подготовки к зиме в клубнях, луковицах и корневищах накапливаются запасные вещества, которые начинают использоваться с началом вегетационного периода.
biofile.ru
Влияние температуры | Земная флора
Реакции на температуру, так же как и на свет, у растений; могут быть и качественными и количественными. Скорости почти всех химических процессов в растении с повышением температуры градуально (т. е. плавно) возрастают, достигают некоторого максимума, а затем снижаются. В отличие от этого многие онтогенетические процессы, например прорастание семян и прерывание покоя почек, часто регулируются по принципу «всё или ничего». В последних случаях для низкотемпературной индукции требуется непрерывное воздействие в течение какого-то минимального периода; это напоминает фотопериодическую индукцию, при которой тоже необходимы совершенно определенные периоды темноты.Градуальные реакции
Скорость большинства химических процессов неуклонно возрастает с повышением температуры. Температурный коэффициент Qio показывает, во сколько раз увеличивается скорость реакции при повышении температуры на 10° по сравнению с первоначальной: Qio для большинства химических (и в том числе ферментативных) реакций равно по меньшей мере 2, тогда как для физических процессов, таких как диффузия или фотохимические реакции, Qio составляет около 1,1 или 1,2. У растений, выращиваемых при температурах от 0 до 30 °С, с повышением температуры постепенно увеличивается скорость вытягивания и QiQ равно около 2,0 или больше. По неизвестным нам причинам температурные оптимумы для разных растений очень сильно различаются, и это указывает на то, что какой-то фундаментальный биохимический процесс у них обладает различной чувствительностью к температуре. В зоне выше оптимума абсолютная скорость роста у всех растений снижается, иногда очень резко. Для большей части растений этот оптимум лежит в области от 28 до 32 °С.
Мы не знаем, почему большинство растений повреждается температурами выше примерно 30 °С, хотя ферменты или орга-неллы, выделенные из растений, при таких температурах обычно не повреждаются. Одно из возможных объяснений состоит в том, что мембраны клеток или их органелл чувствительны к изменениям температуры из-за плавления или затвердевания жирных кислот в фосфолипидах. Известно, что растения при более низких температурах синтезируют больше ненасыщенных жиров с соответственно более низкой температурой плавления.
Обратное явление наблюдается при высоких температурах. Другая возможность состоит в том, что при повышенных температурах какие-то материалы, необходимые для роста, очень быстро разрушаются или не образуются в нужном количестве. У многих организмов имеются гены «чувствительности к температуре». У плесневого гриба Neurospora ген, ответственный за образование витамина В2 (рибофлавина), хорошо функционирует при низких температурах, но не действует нормально при более высоких. Поэтому при 35 °С гриб должен непременно получать рибофлавин извне, тогда как при 25 °С он может расти и в отсутствие этого вещества. Подобным образом дело могло бы обстоять с другими витаминами, аминокислотами или гормонами у высших растений. Если это так, то, зная химическую основу индуцированного температурой замедления роста при супраоптимальных температурах, можно было бы улучшить рост путем внесения нужных веществ.
Морфогенетические эффекты
Очень малая скорость химических реакций в растении при низких температурах обеспечивает координацию изменений роста с климатическими изменениями. Кроме того, температура влияет на многие процессы, чувствительные к фотопериоду, изменяя критическую длину темного периода, хотя механизм этого явления не выяснен. Так как наличие Фдк ночью могло бы нарушать индукцию цветения у растения короткого дня, можно было бы ожидать, что высокие ночные температуры, ускоряющие разрушение фитохрома и превращение Фдк в Фк, будут благоприятствовать индукции цветения у таких растений. На самом же деле обычно наблюдается противоположный эффект. Некоторые растения можно заставить цвести с помощью длинных темных ночей или низких ночных температур. Возможно,, что каждое из этих воздействий способно активировать какой-то процесс, ведущий к синтезу флоригена.
geo-plant.ru
Зависимость роста растений от экзогенных факторов
Рост, являясь интегральной функцией растения, зависит от влияния абиотических факторов внешней среды: света (его интенсивности, спектрального состава, продолжительности и периодичности), температуры (величины и периодичности), питательных веществ (количества и сбалансированности), механических воздействий ветер и др.).
Регуляция роста светом. Разносторонние процессы, которые определяют влияние света на форму растений, были названы фотоморфогенезом.
Растения, выросшие в темноте, называются этиолированными. Они отличаются от растений, выращиваемых на свету, рядом морфологических и биохимических особенностей.
Программа развития нормального зеленого растения контролируется специальными световыми реакциями, лежащими в основе фотоморфогенеза. Хлорофилл и разные вспомогательные пигменты фотосинтетического аппарата не играют в этих процессах главной роли, посколько начальные этапы развития растений на свету не могут осуществляться за счет фотосинтеза: структурная и биохимическая основы фотосинтетического аппарата в этот период еще не сформированы. Для устранения эффекта этиоляции и включения процессов фотоморфогенеза достаточно ежедневного 5–10 минутного освещения растений.
Термин «фотоморфогенез» объединяет процессы, которые не зависят от направления и периодического освещения, а определяются длиной волны и интенсивностью светового потока. Вероятно, в процессах фотоморфогенеза растений играют роль определенные соединения – пигменты.
Содержание этих пигментов в растениях очень малое, поэтому количество энергии, необходимое для насыщения соответствующих фотопроцессов, на несколько порядков ниже, чем при фотосинтезе. В то время как при фотосинтезе 8–10 квантов света необходимо для выделения только одной молекулы О2, такое же количество квантов на одну клетку может полностью определить репродуктивную судьбу растения или направление роста всего стебля.
Реакция растений на длительность светового периода (или длину дня) разная. Эта реакция растений на продолжительность дня получила название фотопериодизма. Выделяют растения короткого дня, когда растения зацветают при небольшой продолжительности освещения – менее некоторой критической величины. Когда же длина дня превышает определенную критическую величину – это растения длинного дня. Имеется и группа растений, цветение которых не зависит от длины дня – нейтральные растения.
Критический фотопериод у разных видов и даже сортов растений как короткого, так и длинного дня значительно варьирует. Эти различия в критическом фотопериодизме играют важную роль в размещении растений на Земле.
После открытия фотопериодизма выявили, что многие растения реагируют на длительность непрерывной темноты, а не на протяженность светового периода. Иными словами, так называемые растения короткого дня – это на самом деле растения длинной ночи. Для закладки цветковых бугорков им необходимо определенная минимальная длительность темноты, которая не прерывается светом. Подобным образом растение длинного дня может быть в действительности растением короткой ночи: оно будет цвести только в том случае, если ночной период не продолжительней некоторого максимума.
Всего только один подходящий период темноты может привести растение короткого дня к цветению, даже тогда, когда последующие темновые периоды недостаточно длительны. Этот феномен известен как фотопериодическая индукция. У многих растений длинного дня имеет место подобный, но обратно направленный феномен: прерывание очень длительного темнового периода вспышкой света приводит к индукции и заложению цветков.
Последнее говорит о том, что растения длинного дня и короткого дня, вероятно, владеют одинаковыми фотопериодическим механизмом, но последний каким-то образом действует в разных направлениях.
Это нашло дальнейшее подтверждение при изучении действия света разной длины волны – спектр действия. Для ингибирования цветения короткодневных растений и для активации цветения длиннодневных растений во всех случаях был определен максимум активности в красной области спектра (около 660 нм) при почти полной неэффективности других длин волн. Подобие спектров позволило считать, что зацветание растений как короткого, так и длинного дня контролируется одним и тем же пигментом.
Далее, свет сильно влияет на прорастание семян. Некоторые семена плохо прорастают в полной темноте, но быстро и хорошо при экспонировании на свету на протяжении нескольких минут. Оказалось, что спектр действия для этого эффекта подобен спектру действия для цветения.
Проросток (гороха), который выращивается в полной темноте, имеет очень длинный тонкий стебель, апикальный изгиб и почти не распустившиеся листья. Когда такой этиолированный проросток выставить только на короткое время на свет, листья после этого разворачиваются, изгиб начинает выпрямляться, удлинение стебля замедляется. Поэтому, появилась возможность заключить, что такие разносторонние реакции как прорастание семян, изменение этиолированного проростка (его деэтиолирование) и индукция цветения регулируются одним и тем же пигментом – рецептором.
Как определили, таким пигментом является фитохром. Фитохром – от греческого слова «растение» и «краситель». Фитохром представлен растворимыми в воде соединениями, относящимися к билихромпротеинам. Как и у фикобилинпротеинов (фикоцианобелин), хроматофор (пигментная часть молекулы) фитохрома представлен незамкнутыми порфиринами (тетрапирол). Другая часть молекулы образуется бесцветным белком. После поглощения фитохромом света с определенной длинной волны форма хроматофора изменяется, и, это в свою очередь, изменяет форму белкового компонента. Молекулярная масса фитохрома колеблется в зависимости от вида растения от 120 до 170 кДа. В составе фитохрома – одна полипептидная цепь. Существует, как предполагают, фитохром в клетке в виде димера.
Фитохром регулирует многие физиологические реакции, наблюдаемые при поглощении малых доз красного и дальнего красного света растениями, которые находились в полной темноте.
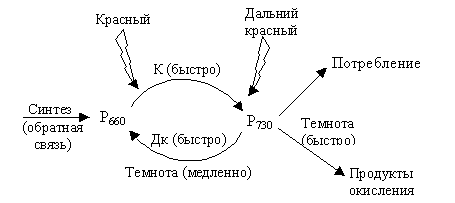
Фитохром находится в растениях в двух формах: Р660 – поглощает красные лучи и превращается в Р730; последний поглощает дальние красные лучи и переходит в Р660 (рис. 6.16). Р660 (Рк) – физиологически неактивный.
 |
| Рис. 6.16. Схема биологического ответа при действии красного и дальнего красного света |
Структура фитохрома представлена на рис. 6.17.
 |
| Рис. 6.17. Структура фитохрома |
Реакции, управляемые фитохромом, зависят от концентрации Р730 (Рдк).
Полагают, что Ф660 содержит на один протон больше, чем Ф730 (см. рис. 6.17). Эта разница влияет на конформацию хромотофора и белковой молекулы, с которой он связан.
Фитохром присутствует в растениях, как уже отмечалось, в небольших количествах: его концентрация составляет около 1 мкМ. При этом он размещен в растении неравномерно, больше всего его в растущих, делящихся тканях, например в клетках меристемы. В клетках стебля он практически отсутствует.
До освещения красным светом фитохром обычно распределен по всей цитоплазме и ее мембранах, а после короткого освещения выявляется только в определенных частях клетки. Вероятно, эти части – ни ядра, ни пластиды, ни митохондрии, а, скорее, ЭР, разбросанный по всей цитоплазме. Длительное воздействие света приводит к появлению фитохрома уже и в ядрах. Подтверждают эти выводы эксперименты с изолированными субклеточными фракциями; однако, не все согласны с интерпретацией полученных данных.
Все же данные физиологических опытов позволяют предполагать, что фитохром может находиться в клетке во многих местах в соответствии с его многочисленными функциями. Вероятно, он локализован внутри этиопластов и митохондрий, или в их наружных мембранах, так как изолированные органеллы реагируют на воздействие красным и дальним красным светом, после их выделения. Предполагают, что фитохром находится (или каким-то образом связан) в плазмалемме.
Считают даже, что хроматофору фитохрома свойственна определенная ориентация в плазмалемме. Кроме того, вероятно, что фитохром изменяет свою ориентацию в мембране на 90 о при фототрансформации
Обычно реакции достигают насыщения, когда 50 % фитохрома представлена формой Р730, и продолжается до той поры, пока имеется достаточное количество Р730.
Превращение Р660 ↔ Р730 происходит также и in vitro. Как считают, оно связано с конформационными изменениями участков белковой части молекулы фитохрома. Этот процесс происходит через промежуточные этапы, потому что пигмент исчезает быстрее, чем появляется другой.
Из-за того, что Р730 неустойчив, его количество в темноте уменьшается, что объясняется медленным тепловым превращением в Р660 и довольно быстрым, по всей вероятности, окислительным разрушением, катализируемым металлами. Очень маленькое количество Р730 остается при выдерживании растений в темноте.
 |
| Рис. 6.18. Схема превращения фитохрома |
Потери фитохрома компенсируются in vivo его синтезом, который начинается по принципу обратной связи, когда суммарное количество фитохромов (Р660 + Р730) становится ниже порогового (рис. 6.18). Синтез пигмента (незамкнутого порфирина) происходит, как и синтез хлорофилла (циклического порфирина), через δ-аминолевулиновую кислоту.
Под влиянием Дк света Р730 быстро и почти целиком (около 90 %) превращается в Р660, а при красном свете Р660 превращается в Р730 примерно на 80 %.
Солнечный свет содержит приблизительно одинаковые доли К и Дк. Это приводит к равновесию Р730/Р660, при котором содержание Р730 доходит до 50 % (во всяком случае, больше, чем в темноте). Поэтому в реакциях, регулируемых фитохромом, солнечный свет действует как красный.
Важная роль фитохрома обусловлена тем, что это единственный из растительных пигментов, который использует световые сигналы для управления развитием растения. Ряд ферментативных реакций, связанных с процессом фотосинтеза регулируются фитохромом.
В соответствии с общими представлениями, впервые высказанными Х. Мором (1966 г.), действие фитохрома связано в первую очередь с регуляторным влиянием Дк-формы на активность так называемых фотоиндуцибильных генов. В общем виде генетический путь трансдукции может быть представлен следующей схемой:
К → Дк → активация или репрессии гена → рост или снижение содержания РНК → модификация содержания белкового продукта → биологический эффект.
Кроме того, на основании полученных данных считают, что Са2+ и кальмодулин принимают участие в передаче сигнала от Дк-формы фитохрома к генетическому аппарату, а так же в этом процессе участвует и G-белок. Генетический механизм обеспечивает у растений медленные фитохромные ответы.
Однако, быстрые фитохроминдуцируемые эффекты (фотоиндуцируемые изменения потенциала, набухание хлоропластов и т. д.), вероятно связаны с модификацией мембранных структур. В механизмах мембранной трансдукции фитохромного сигнала важную роль играет Са2+.
Мембранный механизм передачи фитохромного действия имеет следующий вид:
К → Дк → изменение проницаемости мембраны для ионов Са2+ → временное изменение концентрации Са2+ в цитоплазме → включение кальций-зависимой биохимической системы → биологический ответ.
Кальциевый поток действует на активность кальмодулина, последний изменяет активность ферментов, например, протеинкиназ. Через действие катаболических ферментов может происходить активация фитогормонов, запускающих механизм ростовой фотореакции.
Так состояние фитохрома может регулировать качественный и количественный состав гормонов четырех типов – этилена, цитокининов, ауксина и гиббереллинов – в разных органах растения. Кроме того, длительное облучение иногда контролирует уровень АБК. В некоторых случаях внесение гормона может снижать эффект трансформации фитохрома (например действие гиббереллина при деэтиляции).
В природе фитохром, вероятно, контролирует форму растений и прорастание семян под пологом листвы, так как листья очень сильно поглощают свет в области 660 нм и относительно слабо – 730 нм. Фитохром может регулировать прорастание семян, деэтилирование, цветение, опадение листьев и даже старение.
Синий свет также регулирует фотоморфогенетические реакции, в частности движение устьиц, фототропизм, ранние этапы синтеза хлорофилла и каротиноидов, движение листьев за солнцем, фототаксис хлоропластов, угнетение роста гипокотилей и стеблей в длину. Фоторецепторы, участвующие в восприятии синего света, называют криптохромами.
У растений арабидопсиса выявлены соединения (криптохром 1, криптохром 2 и фототропин), которые воспринимают и передают сигналы синего света. Криптохропы – рецепторы синего света, локализованные в ядре и цитоплазме, включающие белковую и хромофорную составляющие. Функции хромофоров у криптохромов выполняют флавины (ФАД, деазафлавин) и (или) птерин.
Криптохромная система замедляет рост гипокотеля (или междоузлия) на свету, помогает фитохрому контролировать процессы диэтиоляции. Обе фоторецепторные системы (криптохром и фитохром), действующие в ядерном компартменте, дополняют друг друга, что сказывается в интеграции информации о физиологически важных областях спектра – синей и красной.
Фототропин представляет собой ассоциированный с мембраной белок, т. е. является мебранным рецептором синего света, функцию хромофора у которого выполняет флавин ФМН. Фототропин ответственен за фототропизм, хемотропизм и потенциалзависимые мембранные процессы.
Характер ответных реакций растений при освещении синим светом определяется его интенсивностью и длиной волны.
У папоротника Adiantum capillus-veneris был найден ген, гомологичный как фитохромам, так и фототропинам. «Двойной» фоторецептор был назван суперхромом. Как и фототропин, суперхром обладает протеинкиназной активностью.
Следовательно, если у одних растений фототропин взаимодействует с фитохромами опосредованно, то у Adiantum проблема взаимодействия красного и синего световых сигналов решена на уровне «двойного» рецептора.
Таким образом, конечная форма зеленого растения, обусловленная в целом его генами, определяется участием света, который действует через разные фоторецепторы.
Влияние температуры. Яровизация. Стратификация.Реакции на температуру, так же как на свет, у растений могут быть количественными и качественными. Скорость почти всех химических процессов с повышением температуры возрастает, достигает некоторого оптимума, а затем снижается. В отличие от этого многие процессы онтогенеза, например, прорастание семян, прерывание покоя почек, часто регулируется по закону «все или ничего». Это означает, что, например для низкотемпературной индукции требуется воздействие на протяжении какого-то минимального периода; приведенная закономерность напоминает фотопериодическую индукцию, при которой также необходимы определенные периоды темноты.
Влияние температуры, как мы уже отмечали, на физиолого-биохимические процессы характеризуют температурным коэффициентом:
 |
При анализе зависимости роста растений от температуры выделяют три кардинальные точки: минимум (рост только начинается), оптимум (наиболее благоприятный период для роста) и максимум (прекращение роста). Кардинальные точки для различных растений неодинаковы. У теплолюбивых растений (кукуруза, огурец, дыня, тыква), все точки смещены в сторону более высоких температур.
В среднем минимальные температуры для роста растений умеренной зоны –5 – +15, оптимальные – +25 – +35, максимальные – +37 – +44 0С. Растения подразделяются на теплолюбивые и холодостойкие.
Кривые зависимости скорости роста растения от температуры представлены на рис. 6.19.
 |
| Рис. 6.19. Влияние температуры на рост корней гороха (1) и колеоптилей кукурузы (2) |
Пока еще окончательно неясно почему большинство растений повреждается температурами выше ~ 30 оС, тогда как ферменты или органеллы, выделенные из растений, при таких температурах обычно не повреждаются. Одно из возможных объяснений заключается в том, что мембраны клеток или органелл чувствительны к изменениям температуры из-за плавкости или затвердения жирных кислот в фосфолипидах. Известно, что растения при низких температурах синтезируют больше ненасыщенных жиров с более низкой температурой плавления. Обратное явление наблюдается при высоких температурах. Другая возможность состоит в том, что при повышенных температурах какие-то материалы, необходимые для роста, очень быстро разрушаются или не образуются в необходимых количествах. Во многих организмах имеются даже гены, «чувствительные к температуре».
У плесневого гриба Neurospora ген, ответственный за образование витамина В2 (рибофлавин), хорошо функционирует при низких температурах, но плохо действует при более высоких. Поэтому при 35 оС гриб должен получать рибофлавин из вне. Подобным же образом может происходить у высших растений при недостатке витаминов, аминокислот, гормонов и т. д. Таким образом, можно было бы улучшить рост путем внесения необходимых веществ.
Очень маленькая скорость химических реакций в растении при низких температурах обеспечивает координацию изменения роста с климатическими изменениями. Кроме того, температура влияет на многие процессы, чувствительные к фотосинтезу. Некоторые растения можно заставить цвести с помощью длинных темных ночей или низких ночных температур.
Низкая температура может вызвать прорастание семян, сдвиги температуры могут прервать покой почек и подготовку к закладке цветочных бугорков.
Для появления всходов требуются более высокие температуры, чем для прорастания семян (табл. 6.2).
Таблица 6.2
Похожие статьи:
poznayka.org









