11. Строение хромосомы. Типы хромосом. Эу– и гетерохроматин. У растений хромосомы
Сколько хромосом у картошки? Особенности генома растения

Генетические исследования различных культур растений важны для сельскохозяйственной промышленности. Каждое живое существо на земле имеет свой генетический код. От того, сколько хромосом у картошки, зависят последующие исследования в селекции. Создание новых усовершенствованных сортов необходимо для произрастания в современных условиях.
Количество хромосом у картошки
Клетки, содержащие наследственную память, называются хромосомами. Они включают в себя нуклеиновую кислоту, функция которой заключается в сохранении, воплощении и последующем продолжении информации. В большинстве случаев хромосомы в клетках содержатся попарно. У картофеля 48 хромосом или 24 пары.
Для сравнения число пар хромосом в других растениях:


Хромосомы картошки
Горох посевной – 14.- Земляника лесная – 14.
- Гречиха культурная – 16.
- Черешня – 16.
- Редис – 18.
- Арбуз – 20.
- Фасоль – 22.
История хромосомного анализа насчитывает более 140 лет. Термин хромосома впервые употребил немецкий ученый В. Вальдейер в 1888г.
Особенности генома растений

Нарезанная картошка
Основной биологической характеристикой растения является количество показателя ДНК на гаплоидный геном. Эукариот – это клетки живых организмов, содержащих ядро. Все живое имеет такие клетки, кроме бактерий и архей. Изучение эукариот растений выявило парадоксы. Их суть состоит в значительном колебании размера ДНК, несоответствие между их размерами у различных видов. Был обнаружен избыток ДНК по сравнению с необходимым количеством для кодирования белков. Эти несоответствия объясняются плоидией. Данный термин обозначает хромосомную аномалию, которая может возникать в силу внешних и внутренних факторов. Активное изменение структуры хромосом присуще растительному миру.
Методы исследования в генетике растений
Существует множество методик исследований в науке. Из широко распространенных выделяются:

Четыре картошки
моносомный анализ;- экспериментальный мутагенез;
- гибридизация;
- мскусственный ресинтез;
- автополиплоидия.
Перечисленные методы играют важнейшую роль при изучении наследственности и изменчивости растений. Интересен метод экспериментального мутагенеза. В процессе работы на геном происходит воздействие химическими веществами или радиацией. Полученные результаты составляют генетические карты хромосом. Измененный геном впоследствии проявляет себя по-другому в различных условиях внешней среды. Так называемая карта памяти оставляет новые физиологические и биохимические особенности различных видов растений.
В настоящий момент передовым методом стала гибридизация. Его суть заключается в скрещивании генетического материала в одну клетку зиготу. Тем не менее, вклад мутации в генетику растений немаловажен. С помощью этого метода в мире создано порядка 2 700 мутантных сортов, среди которых только зерновые культуры занимают 50 %. С помощью радиации создано 89 % сортов от общего количества средств, которые применяются для мутации. Зеленая революция позволила удвоить урожаи пшеницы и риса в развивающихся странах. Использование генетического материала позволяет создавать эффективные и ресурсосберегающие сорта растений. Советуем также прочитать нашу статью Гликемический индекс картофеля: жареного, варёного, пюре, чипсов. Калорийность.
© Lifeo.ru
Предыдущая:Как научить собаку команде «Лежать». Пошаговое руководство, ошибки при обученииДальше:Сколько хромосом у собаки Голос за пост - плюсик в карму! :) Загрузка...
Загрузка...lifeo.ru
Половые хромосомы у растений - Справочник химика 21
Половые хромосомы обнаружены у 70 видов покрытосеменных растений, принадлежащих к 25 семействам. [c.94]Существует несколько причин, по которым полиплоидия среди животных встречается реже, чем среди растений. Во-первых, полиплоидия нарушает баланс между аутосомами и половыми хромосомами, необходимый для определения пола. Во-вторых, большинство животных размножается посредством перекрестного оплодотворения единичный мутант-полиплоид не может размножаться сам по себе. В-третьих, [c.67]
Такого же типа сцепленное с полом наследование было обнаружено у многих других животных, двудомных растений, а также у человека. Среди 23 различных пар хромосом человека лучше всего изучены в отношении содержащихся в них генов половые хромосомы. У самцов дрозофилы между X- и -хромосомами перекреста не бывает, а следовательно, и обмена между находящимися в них генами не происходит. У человека же, по-видимому, возможен обмен некоторыми генами, [c.147]Двудомных видов мало. К ним относятся конопля, хмель, тополь, ива. У некоторых двудомных видов в клетках обнаружены половые хромосомы типа XX и XV (некоторые мхи, элодея, дрема). Генетической основой определения пола у этих двудомных растений являются X- и У-хромосомы, однако гены, ответственные за детерминацию пола, локализуются не только в половых хромосомах, но и в аутосомах (т. е. во всех других хромосомах). Тем более это относится к растениям, у которых специализированных половых хромосом нет. Гены, ответственные за определение пола, играют важную роль, позволяя клеткам по-разному реагировать на действие факторов внутренней и внешней среды. В отличие от животных процесс формирования пола в ходе онтогенеза растений может сильно меняться в зависимости от условий вплоть до превращения одного пола в другой. [c.375]
У двудомных растений также обнаружены половые хромосомы. [c.129]
У двудомных растений, так же как и у животных, в ряде случаев обнаружены специфические половые хромосомы. Впервые половые хромосомы у растений были обнаружены Алленом в 1917 г. при исследовании печеночного мха Sphaero- arpus. Выше было указано (стр. 35), что растения мхов всегда гаплоидны, тогда как спорофит — т. е. спорангий и его ножка — диплоидны. У вида, изученного Алленом, было обнаружено, что мужское растение имеет 7 обычных хромосом и одну маленькую точечную Y-хромосому, тогда как хромосомный набор женских растений состоит из тех же семи обычных хромосом и одной очень длинной Х-хромосомы (фиг. 52). При оплодотворении эти два хромосомных набора соединяются, и таким образом спорофит имеет набор 14А + X -Ь Y. Во время мейоза образуется 7 пар аутосом и одна пара X = У. Следовательно, половина спор получит набор 7А + X, а другая по- [c.135]
Наследственность корпускулярна (дискретна), и каждая из родительских особей передает своему потомку одинаковое число генов, за исключением тех животных и растений, у которых имеют-ся половые хромосомы. У этих организмов вклады родителей [c.58]
Перемещения хромосом во время митоза и мейоза осуществляются также с помощью сократительных элементов, так называемых микротрубочек. Микротрубочки — это вытянутые полые структуры длиной в несколько микрометров при диаметре всего 15—25 нм и толщине стенки около 6 нм. В микротрубочках содержится белок тубулин, изменяющий свою конфигу рацию в ответ на некоторые химические воздействия, например под влиянием ионов Са +. Микротрубочки прикрепляются к особому участку хромосом, к так называемому кинетохору, и помогают растаскивать хромосомы к противоположным полюсам клетки во время клеточного деления. Снабженные жгутиками клетки водорослей и подвижные половые клетки гаметы) различных растений движутся также благодаря сокращению микротрубочек. В поперечном сечении жгутики обычно имеют характерное строение 9 пар микротрубочек образуют кольцо, окружающее 2 пары, находящиеся в центре. Плазмалемма (а, быть может, также и тонопласт) находится в непрерывном движении. На ней возникают как бы волдыри , которыми она окружает и захватывает находящиеся снаружи частички или же крупные молекулы, после чего эти частички в процессе пино-цитоза транспортируются в цитоплазму в маленьких мембранных пузырьках. Аналогичным образом протекает и обратный процесс — выведение различных материалов из клетки наружу. [c.75]
Отдаленная гибридизация между культурными растениями и родственными дикими видами позволяет не только расширить генетическую изменчивость культурного вида, но, что наиболее важно, и привнести отдельные ценные гены от дикого вида, отсутствующие у культурного вида. Подобные скрещивания обычно являются весьма сложным делом, поскольку между видами существуют жесткие репродуктивные барьеры. Чаще всего пыльца чужого вида не может расти на пестике и оплодотворить яйцеклетку. Если оплодотворение все же произошло, то семена получаются нежизнеспособными из-за недоразвитого эндосперма (питающего элемента семени), или из семени развиваются стерильные, не способные формировать жизнеспособные половые клетки гибриды. Между видами существуют также жесткие барьеры, затрудняющие естественную рекомбинацию. Это означает, что хромосомы межвидового гибрида, полученные от разных видов, могут отличаться по количеству и гомологии (сходству). Отсутствие гомологии между хромосомами приводит к тому, что они не способны сближаться и обмениваться отдельными участками (а следовательно, и отдельными генами) в процессе образования половых клеток. В результате становится невозможным перенос нужных ге- [c.21]
Гибридизация соматических клеток. Следующий метод клеточной селекции — создание неполовых гибридов путем слияния изолированных протопластов, полученных из соматических клеток. Этот метод позволяет скрещивать филогенетически отдаленные виды растений, которые невозможно скрестить обычным половым путем, вызывать слияние трех и более родительских клеток, получать асимметричные гибриды, несущие весь генный набор одного из родителей наряду с несколькими хромосомами или генами, или только органеллами и цитоплазмой другого. Гибридизация соматических клеток дает возможность не только соеди-нить в одном ядре гены далеких видов растений, но и сочетать в гибридной клетке цитоплазматические гены партнеров. [c.154]
Кроме того, среди двудомных растений найдены формы, у которых не удается различить половые хромосомы. К таким относится, например, конопля annabis). У грибов, а также у некоторых двудомных цветковых растений имеются формы, промежуточные между чисто мужскими и чисто женскими растениями. Определение пола в таких случаях сложное, и в настоящее время механизм его еще не вполне выяснен. [c.138]
Половые клетки растения или животного содержат п хромосом. Это число широко варьирует у разных видов — от одной хромосомы до нескольких сотен, но оно постоянно для каждого вида, например у человека п 24. Когда при оплодотворении сперматозоид сливается с яйцеклеткой, образуется клетка, содержащая 2п хромосом так, для человека 2п 48 . Хромосомы половой клетки, как правило, все разные и в благоприятных случаях их можно различить по величине и форме под микроскопом. Каждая хромосома, внесенная мужской половой клеткой (за одним исключением), в основном идентична гомологична) соответствующей хромосоме, внесенной женской половой клеткой. Одна пара хромосом — половые хромосомы у многих животных, включая человека и плодовую мушку дрозофилу, — является исключением в том отношении, что в оплодотворенном яйце (или зиготе), которое разовьется в мужской организм, две хромосомы, составляющие пару, различны можно отличить Х-хромосому, внесенную яйцеклеткой, и У-хромосому, внесенную спермием. С другой стороны, зигота, которая должна дать женский организм, имеет две Х-хромосомы. Имеются спермин двух типов одни несут Х-хромосому и дают начало зиготам, развивающимся в самок, другие несут У-хромосому и образуют зиготы, дающие самцов. У птиц и бабочек гетерогаметная самка имеет хромосомы XY. Семенные растения и многие низшие животные не всегда имеют механизм определения пола типа XV. [c.105]
Отсутствие половых хромосом у диких и культурных растений устраняет самый важный барьер на пути полиплоидии, распространенной среди них в сотни раз чаще, чем среди культурных животных, особенно млекопитающих. Наличие строгого контроля половых хромосом, свойственного животным, резко уменьшает возможности полиплоидии. В подавляющем большинстве случаев ее возникновение в гомогаметпческом поле ведет к очень редко преодолимым аномалиям баланса половых хромосом у гетерохромосомного пола, поэтому полиплоидия в эволюции высших животных представляет большую редкость. У растений, не имеющих, за известными исключениями, половых хромосом, полиплоидия возникает чаще и является важной формой эволюции растений. Надо все же заметить, что даже у растений с хромосомной детерминацией пола последняя является гораздо менее строгой, и у большинства родов растений, несущих ноло-вые хромосомы, имеются полиплоидные виды. Скорее всего это возникает потому, что наличие половой детерминации пола у растений не уподобляет их животным по нормировке непрерывности зародышевой плазмы. [c.24]
Генетика человека опирается на общие принципы, полученные первоначально в исследованиях на растениях и животных. Как и у них, у человека имеются менделирующие, т.е. наследуемые по законам, установленным Г Менделем, признаки. Для человека, как и для других эукариот, характерны все типы наследования аутосомно-доминант-ный, аутосомно-рецессивный, наследование признаков, сцепленных с половыми хромосомами, и за счет взаимодействия неаллельных генов. Разработал Г.Мендель и основной метод генетики - гибридологический. Он основан на скрещивании особей одного вида, обладающих альтернативными признаками, и количественном анализе полученных фенотипических классов. Естественно, этот метод не может использоваться в генетике человека. [c.104]
Анеуплоидия, или гетероплоидия, возникает вследствие изменения числа хромосом, не кратного гаплоидному набору. Об этом уже упоминалось в гл. 5, где обсуждалось нерасхождение и потери Х-хромосом у D. melanogaster. В результате нерасхождения хромосом при гаметогенезе могут возникать половые клетки с лишними хромосомами, и тогда при последующем слиянии с нормальными гаплоидными гаметами они образуют зиготы 2п- -1, или трисомики, по определенной хромосоме. Если в гамете оказалось меньше на одну хромосому, то последующее оплодотворение приведет к образованию зиготы 2п— 1, или моносомика, по какой-либо из хромосом. Полисомия и моносомия могут иметь самостоятельное фенотипическое проявление вследствие изменения соотношений доз некоторых генов или нарушения генного баланса. Так, А. Блексли и Дж. Бел-линг в 20-х годах показали, что создание трисомиков по каждой из 12 хромосом у дурмана (Datura stramonium) приводит к появлению характерного, отличного от других типа растения. В частности, это выражалось в специфическом изменении формы семенной коробочки. [c.362]
При половом размножении растений и животных (в том числе ,Й человека) преемственность между поколениями обеспечивается Только через половые клетки — яйцеклетку и сперматозоид. Если яйцеклетка и сперматозоид обладали полным набором генети-веских характеристик (2п2с), свойственных клеткам тела, то при ИХ слиянии образовался бы организм с удвоенным набором (4п4с). Йапример, в соматических клетках организма человека содержится 46 хромосом. Если бы яйцеклетка и сперматозоид человека содержали по 46 хромосом, то при их слиянии образовалась бы зиго- Га с 92 хромосомами. В следующем поколении проявились бы по-,Томки со 184 хромосомами и т.д. [c.35]
У всех высших растений и животных в процессе полового размножения происходит смена ядерных фаз. При оплодотворении половые клетки (гаметы) и их ядра сливаются, образуя зиготу. Отцовское и материнское ядра вносят при оплодотворении одинаковое число хромосом (п) таким образом, ядро зиготы содержит двойной хромосомный набор (2п). Иными словами, гаметы-гаплоидные клетки (т.е. клетки с одним набором хромосом), а соматические клетки-диплоидные (с двумя наборами). Поэтому при образовании гамет следующего поколения число хромосом в клетке (2и) должно уменьшиться вдвое (2и/2 = и). Совокупность процессов, приводящих к уменьшению числа хромосом, называют мейозом или редукционным делением (рис. 2.3). Мейоз - важнейший процесс у организмов, размножающихся половым путем он приводит к двум результатам 1) к перекомбинированию отцовских и материнских наследственных факторов (генов) и 2) к уменьшению числа хромосом. Мейоз начинается с конъюгации хромосом-каждая хромосома соединяется с соответствующей (гомологичной) хромосомой, происходящей от дфугого родителя. Во время конъюгации путем разрыва и перекрестного воссоединения (кроссинговера) может происходить обмен фрагментами одинаковой длины между гомологичными хромосомами. Затем следует двукратное разделение спаренных расщепившихся хромосом, и в результате образуются четыре клетки, каждая из которых имеет гаплоидное ядро. Таким образом, в процессе мейоза не только происходит перетасовка хромосом материнского и отцовского происхождения, но может произойти и обмен сегментами между гомологичными хромосомами. Оба процесса приводят к новым сочетаниям генов (к их рекомбинации). [c.24]
Несколько легче понять данные Элькерса, полученные при скрещивании между разными видами южноафриканских растений рода Strepto arpus. Здесь также реципрокные скрещивания дают различный результат, в частности в отношении развития пыльников и пестиков. Анализируя возвратные скрещивания между Fl и обоими родителями, Элькерс смог показать, что пол у этих видов определяется равновесием между факторами, содержащимися в хромосомах и в цитоплазме. Все эти виды — гермафродиты, но у некоторых из них развитие пыльников обусловлено главным образом цитоплазмой, а развитие пестиков — хромосомными факторами. У других видов положение может быть противоположным. Поэтому, когда виды, принадлежащие к этим различным группам, скрещивают, а затем производят возвратные скрещивания, возникают различные нарушения либо в мужском, либо в женском половом аппарате. [c.369]
В клеточном ядре на определенной стадии развития клетки становятся видимыми под миюроскояом (под обычным оптическим микроскопом) хромосомы. Это червеобразные структуры число и форма хромосом однозначно характеризуют вид растения или животного. Во всех клетках человеческого тела, кроме половых, содержится по 23 пары хромосом, всего 46 хромосом. В половых клетках их вдвое меньше 23. в сперматозоиде и 23 1В яйцеклетке. При оплодотворении, т. е. при слиянии сперматозоида с яйцеклеткой, образуется полпый набор хромосом — 46. Все последующие клетки тела возникают 1В ршультате многократных делений, начавшихся с первой зародышевой клетки. [c.256]
Внутри ядра имеются длинные образованпя — хроматиновые нити, с трудом видимые в покоящейся клетке, т. е. существующей в период между двумя делениями. Перед делением клетки хроматиновые нити образуют палочкообразные тельца — хромосомы. Число и форма их постоянны для данного вида организмов. В соматических (не половых) клетках высших растений и животных хромосомы существуют попарно, т. е. но две копии каждого типа. Такая пара хромосом называется гомологичной, подобный пабор хромосом — диплоидным. Клетки человека содержат 23 нары хромосом (46). Из общего правила, что в клетках с диплоидным набором каждая хромосома представлена двумя копиями, имеется исключение. Оно заключается в том, что в клетках женских особей одна пара хромосом содержит две одинаковые Х-хромосомы, а в клетках мужских особой — одну X- и одну У-хромосому, отсутствующую в женских клетках. В половых клетках (гаметах) число хромосом вдвое меньше — по одной хромосоме от каждой пары. Этот набор хромосом называют гаплоидным. [c.11]
Блексли установил, что каждый первичный мутант дурмана способен дать два вторичных мутанта, т. е. всего их может быть 24 (рис. 82). Третичные мутанты по той же формуле (2я+1) способны возникать лишь в тех случаях, когда лишняя хромосома образована сегментами, принадлежащими не гомологичным хромосомам, а хромосомам из разных пар. На рисунке 83 представлен набор хромосом дурмана с обозначением их половинок числамЕ от 1 до 24. При этом третичный мутант может иметь лишнюю хромосому, возникающую путем транслокации между хромосомами 1, 2) и (17, 18), что приводит к новым комбинациям 1, 18) и 2, 17). Соответственно этому растения будут иметь набор хромосом, определяемый формулами [2я+(/, 18)] и [2п+ 2, 17)1 что в действительности наблюдалось и было описано. [c.140]
Принадлежность насекомых к империи ядерных соответствует признанию, что клетки их тела имеют оформленные ядра с хромосомами и другие органеллы, облеченные двухслойными биологическими мембранами. Наряду с другими свойствами, отличающими всех представителей империи (растения, грибы, животные), они противопоставляются доядерным (Prokaryota) специфичными формами клеточного деления (митоз, мейоз) и жизненными циклами, предполагающими развитие полового процесса и полового размножения. В соответствии с этим они демонстрируют признанные в генетике и биохимии стандарты, поскольку сами послужили главными объектами исследований такого рода. [c.55]
Для каждого вида растений и животных характерны свое число и свои морфологические особенности хромосомного набора, т.е. определенный кариотип. В норме все хромосомы клеток организма эукариот парны, т. е. каждая хромосома имеет своего аналогичного по размеру, форме и особенностям расположения генов гомолога, и составляют двойной (2п), или диплоидный (от греч. dipioos — двойной), набор. Только зрелые половые клетки содержат одинарный (1п), или гаплоидный, набор хромосом (от греч. haplous — одинарный). [c.39]
К ограниченной полом стерильности и нежизнеспособности гибридов у двудомных растений или раздельнополых животных приложимо обобщение, известное под названием правила Холдейна. Гибриды Fi от межвидовых скрещиваний у раздельнополых жи-. вотных должны состоять, во всяком случае потенциально, из гетерогаметного пола (несущего хромосомы XY) и гомогаметного (XX) пола. Правило Холдейна гласит, что в тех случаях, когда в проявлении стерильности или нежизнеспособности гибридов существуют половые различия, они проявляются чаще у гетерогаметного, чем у гомогаметного пола. У большинства животных, в том числе у млекопитающих и у двукрылых, гетерогаметны самцы мы только что упоминали о гибридной стерильности у лошадей и у дрозофилы. Из правила Холдейна имеются, однако, многочисленные исключения (см. White, 1973). [c.189]
chem21.info
Пол и половые хромосомы у растений
Генетика  Пол и половые хромосомы у растений
Пол и половые хромосомы у растений
Количество просмотров публикации Пол и половые хромосомы у растений - 689
| Наименование параметра | Значение |
| Тема статьи: | Пол и половые хромосомы у растений |
| Рубрика (тематическая категория) | Генетика |
Определение развития пола
Каждому виду свойственно определенное соотношение полов. Чаще оно равно 1:1. Хромосомы, составляющие одну пару в хромосомном наборе мужской и женской особей, по форме и величине очень похожи, кроме одной пары, в которой хромосомы значительно отличаются друг от друга. Установлено, что с этими необычными хромосомами связано определение пола и их называют половыми хромосомами. В хромосомном наборе животных и раздельнополых растений различают обычные хромосомы или аутосомы и половые X и Y – хромосомы. По хромосомной теории наследственности пол организма определяется в момент оплодотворения.
У многих видов особи женского пола имеют парные хромосомы, и в результате редукционного деления у них образуется только один сорт гамет X , а в гаметогенезе мужского пола образуется два сорта гамет: X и О либо X и Y при равном числе аутосом. Пол, дающий гаметы одного сорта - X X– принято называть гомогаметным, дающий два сорта гамет X и О и X и Y – гетерогаметным. Следовательно, в случае если самка определяется наличием в зиготе двух Х – хромосом, а самец – наличием лишь одной Х – хромосомы и мужские гаметы с Х и О образуются в равном числе, то в потомстве окажется соотношение полов.
Р ♀ХХ х ♂ХО ♀ХХ х ♂ХY
F1 1♀XX х 1♂XO или 1♀ХХ : 1♂XY
Это явилось первым научным объяснением расщепления по полу 1:1.
Существует четыре базовых типа хромосомного определения пола (таблица 1).
Таблица 1 – Типы соотношения половых хромосом у животных и растений
| Организмы | Гетерогаметный пол | Гаметы | Зиготы | ||
| яйцеклетки | спермии | самки | самцы | ||
| Человек, дрозофила, древесные виды (раздельнополые) | мужской | Х и Х | Х и Y | Х Х | Х Y |
| Клоп | мужской | Х и Х | Х и О | Х Х | Х О |
| Кузнечик | мужской | Х и Х | Х и О | Х Х | Х О |
| Птицы, бабочки | женский | Х и Y | Х и Х | Х Y | Х Х |
| Моль | женский | Х и О | Х и Х | Х О | Х Х |
Подавляющее большинство высших растений гермафродитные. Гаметы таких организмов генетически совершенно идентичны. Но около 5% цветковых растений двудомные: тычинковые и пестичные цветки у них находятся на различных особях (женских и мужских). У таких организмов выражен половой диморфизм. Среди двудомных видов имеются ценные сельскохозяйственные растения: виноград, конопля, дынное дерево и др. Размещено на реф.рфЗависимость между полом и структурой хромосом у таких растений была обнаружена позднее, чем у животных. Вначале установили сцепленное с полом наследование у дрёмы белой, а затем у этого растения, а также у щавеля, хмеля и др., были открыты половые хромосомы. У растений существует два базовых типа генетического контроля пола (таблица 2).
Таблица 2 – Хромосомный механизм определения пола у двудомных растений
| Вид | Число хромосом (2п) | Пол | |
| женский | мужской | ||
| Конопля посевная | ХХ | ХY | |
| Щавель малый | ХХ | ХY | |
| Спаржа | ХХ | ХY или YY | |
| Дрема белая | ХХ | ХY |
Первый из них определяется присутствием или отсутствием Х – хромосомы. Второй тип связан с факторами, находящимися в половых хромосомах и аутосомах, и развитие признаков того или иного пола определяется соотношением половых хромосом и аутосом.
Пол и половые хромосомы у растений - понятие и виды. Классификация и особенности категории "Пол и половые хромосомы у растений" 2014, 2015.
referatwork.ru
11. Строение хромосомы. Типы хромосом. Эу– и гетерохроматин.
Хромосома- постоянный компонент ядра, отличающийся особой структурой, индивидуальностью, функцией и способностью к самовоспроизведению, что обеспечивает их преемственность, а тем самым и передачу наследственной информации от одного поколения растительных и животных организмов к другому.
Размеры хромосом у разных организмов варьируют в широких пределах. Длина хромосом может колебаться от 0,2 до 50 мкм. Число хромосом у различных объектов также значительно колеблется, но характерно для каждого вида животных или растений. Совокупность числа, величины и морфологии хромосом называется кариотипомданного вида.
Хромосомы животных и растений представляют собой палочковидные структуры разной длины с довольно постоянной толщиной, у большей части хромосом удается легко найти зону первичной перетяжки, которая делит хромосому на два плеча. В области первичной перетяжки находится центромера, где расположенкинетохор. Некоторые хромосомы имеютвторичную перетяжку.
В конце интерфазы каждая хромосома состоит из двух сестринских хроматид. Каждая из них, в свою очередь, состоит из двух половинок – полухроматид или хромонем. Хромонемы содержат уплотненные участки – хромомеры, которые в световом микроскопе имеют вид темноокрашенных гранул. Их число, положение и величина в обеих хроматидах одинаковы и для каждой хромосомы относиельно постоянны. Расстояния между хромомерами называются межхромомерными участками.
Когда говорят о морфологии хромосом, то принимают во внимание следующие признаки: длину плеч, положение центромеры, наличие вторичной перетяжки или спутника. Спутники разных хромосом отличаются по форме, величине и длине нити, соединятющей их с основным телом.
Спутник– это хромосомный сегмент, чаще всего гетерохроматический, расположенный дистально от вторичной перетяжки. По классическим определениям спутник – сферическое тельце с диаметром, равным диаметру хромосомы или меньше его, которое связано с хромосомой тонкой нитью. Выделяют следующие 5 типов спутников:
микроспутники – сфероидальной формы, маленькие спутники с диаметром вдвое или еще меньше диаметра хромосомы;
макроспутники – довольно крупные формы спутников с диаметром, превышающим половину диаметра хромосомы;
линейные С. - спутники, имеющие форму длинного хромосомного сегмента. Вторичная перетяжка значительно удалена от терминального конца;
терминальные С. – спутники, локализованные на конце хромосомы;
интеркалярные С. – спутники, локализованные между двумя вторичными перетяжками.
Вторичная перетяжка, соединяющая спутник с телом хромосомы, способна к участию в процессе формирования и сборки ядрышек. Такая вторичная перетяжка поэтому называется еще ядрышковым организатором.
Спутник вместе с вторичной перетяжкой составляют спутничный район.Вторичные перетяжки могут быть у одних хромосом на длинном плече, у других - на коротком. Концевые участки хромосомы называюттеломерами.Особенность их состоит в том, что они не способны к соединению с другими участками хромосом.
Нормальная длина каждой хромосомы и суммарная длина всех хромосом кариотипа постоянна. Морфология хромосомы определяется в первую очередь положением центромеры. В соответствии с местом расположения центромеры выделяют основные формы хромосом: метацентрические, субметацентрические, акроцентрические и изохромосомы.
Метацентрическиехромосомы отличаются тем что плечи у них одинаковой или почти одинаковой длины.Субметацентрическиехромосомы имеют плечи разной длины. Уакроцентрическиххромосом центромера расположена к близко к одной из теломер.
Изохромосомы – моноцентрические хромосомы с двумя генетически идентичными плечами, появляющиеся как результат неправильного деления центромеры после разрыва и воссоединения сестринских хроматид в области центромеры.Изохромосомаимеет одинаковые плечи в результате деления центромеры по горизонтали.Дицентрические и ацентрическиеизохромосомы образуются после разрыва сестринских хроматид вне центромерной области и воссоединения их в центрические и ацентрический фрагменты.
Хроматин - основной компонент клеточного ядра. В среднем в хроматине 40% приходится на ДНК и около 60% на белки. В структурном отношении хроматин представляет собой нитчатые комплексные молекулы дезоксирибонуклеопротеида, которые состоят из ДНК, ассоциированной с гистонами и иногда еще с негистоновыми белками. Способность к дифференциальному окрашиванию легла в основу выявления двух фракций хроматина – гетеро – и эухроматина. Хейтц, открывший это явление, нашел, что определенные участки хромосом остаются в конденсированном состоянии в течении всего клеточного цикла и назвал их гетерохроматин, а участки, деконденсирующиеся в конце митоза и слабо окрашенные – эухроматином. Гетерохроматиновые участки функционально менее активны, чем эухроматиновые, в которых и локализована большая часть известных генов. Однако, гетерохроматин имеет определенное генетическое влияние; к примеру, определяющие пол хромосомы не могут рассматриваться как генетически неактивные, хотя они часто полностью состоят из гетерохрома тина. Кроме того, установлено, что стабильность генетического выражения эухроматина обуславливается близостью к гетерохроматину.
studfiles.net
Почему и зачем у картофеля 48 хромосом, когда у человека, например, 46?
Вообще, ученые с этим пока не совсем разобрались. Уже где-то к 70-м годам накопились данные о том, что количество ДНК в ядре не очень-то отражает эволюционное положение вида. Это так называемый С-парадокс (С - количество ДНК в гаплоидном ядре, то есть одинарный набор хромосом). Он заключается в следующем, если упростить:
Количество хромосом весьма условно связано с систематическим положением организма.
«Много хромосом ≠ эволюционно продвинутый вид», вопреки распространенному представлению.
Ну и еще кое-что: ДНК, из которой хромосомы сделаны, это не сплошь структурные гены, кодирующие белки. До 80-90% ДНК может состоять из некодирующей части, раньше она называлась мусорной. Она представляет собой коротенькие «бессмысленные» последовательности, которые расположены блоками и повторены сотни тысяч и миллионы раз (в последнее время понемножку становится понятнее, зачем они нужны, но сейчас не об этом). Количество этой странной информации сильно влияет на общее число пар нуклеотидов.
Помимо этого, хромосомы неодинаковы по массе. У разных видов в хромосомы «расфасован» разный объем ДНК, соответственно, при одинаковом числе пар нуклеотидов число хромосом может отличаться.
В определенной степени количество ДНК все же соответствует сложности организмов. Например, у вирусов геном варьирует в пределах 1,3–20*10^3, у бактерий 9*10^5–10^6 пар нуклеотидов. В эволюции позвоночных тоже прослеживается тенденция наращивания количества ДНК: у оболочников и ланцетников размер генома составляет соответственно 6 и 17% от размера генома плацентарных млекопитающих. При этом у некоторых рыб и хвостатых земноводных в 25 раз больше ДНК, чем у любого из видов млекопитающих. В общем, всё довольно запутанно.
Отдельно стоит сказать о растениях.
Животное справляется со многими проблемами, меняя условия среды. Жарко – лёг в тень, голодно – перешел на новый источник пищи и всё такое. У растений нет возможности вот так запросто встать и уйти, поэтому большинство задач решается на химическом уровне. Жарко – синтезируешь воск на поверхности листьев, чтобы вода не испарялась. Голодно – договариваешься с азотфиксирующими бактериями, чтобы поделились азотом. В таком духе (это метафора, на самом деле на подобные адаптации уходят тысячи тысяч поколений и заранее неизвестно, что получится). Естественно, чем больше разнообразных циклов и синтезов "умеет" осуществлять растение, тем больше нужно ферментов для работы этих метаболических путей. У растений относительно много структурных генов, в них записаны все эти необходимые белки. Метаболизм животных устроен проще. Чтобы не завалить сессию Как во время сессии запомнить то, что мне абсолютно не нужно?Как научиться красиво и грамотно говорить?Когда выгоднее идти сдавать: в первой волне или ближе к концу?
А еще для растений характерна такая интересная вещь, как полиплоидия – кратное увеличение числа хромосом. То есть жил-был геном, а потом однажды взял – и умножился на 2, 3, 10 и так далее. И всё нормально, бывает что даже очень хорошо – удваиваются (утраиваются, удесятеряются) элементы цветка, размер плодов, общая биомасса растения. Процесс может запуститься случайно, но коварные селекционеры давно приспособились его провоцировать для получения культурных сортов.
Для большинства животных такая ситуация очень неполезна, у нас любые резкие отступления от стандартного набора хромосом ведут к уродствам. Суть в том, что число хромосом у растений может в разы отличаться даже внутри семейства. Конечно, всё имеет свою цену – чем больше хромосом, тем больше вероятность что какие-то из них при делении неправильно разойдутся. И меньше вероятность, что найдется другой такой же полиплоид в качестве партнера для размножения. Из-за этого полиплоиды временами оказываются бесплодны. Эволюционный процесс отбраковывает такие, а мы их размножаем вегетативно у себя на клумбах. Но иногда бывает, что всё складывается очень удачно, и образуется целый новый вид. Яркий пример - семейство Ужовниковые (это папоротники). В среднем у разных видов ужовников по 120 пар хромосом, но абсолютный рекордсмен - маленький Ophioglossum reticulatum с диплоидным набором в 631 пару хромосом на клетку (по другим данным, 720). Зачем ему столько и как он весь этот ворох ДНК организует, не ясно. Но раз он всё еще существует как вид, значит, это сработало.
Резюмируя: не завидуйте картошке. Не стоит упрекать её в том, что ей досталось на две хромосомы больше. Это не хорошо и не плохо, не много и не мало, не прогрессивно и не убого. В этом вопросе больше не значит лучше. Просто так сложились обстоятельства.
thequestion.ru
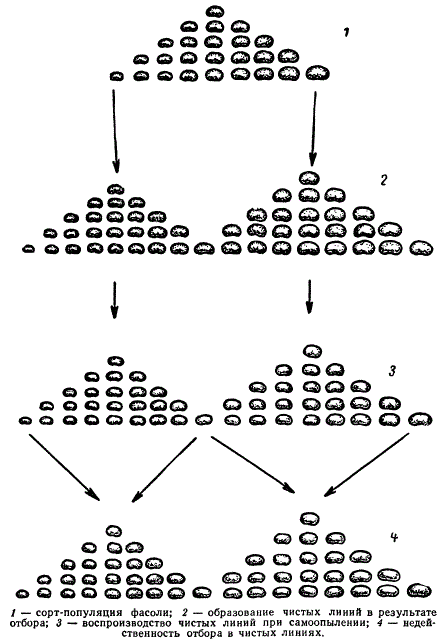
Хромосомы растений
Хромосомы — участки дезоксирибонуклеиновой кислоты, обособляющиеся прн делении клеток. Число их характерно для определения вида животного или вида растений.[ ...]
Хромосомы культурных растений и их дикорастущих сородичей,—М.: Изд-во Университета дружбы народов, 1984.[ ...]
Хромосомы имеют различную форму. Это либо прямые или изогнутые палочки, либо разнообразные крючки, овальные тельца, шарики (рис. 36). Сильно варьируют они и по размерам. Каждая клетка растений данного вида содержит в своем ядре одинаковый набор (или наборы) хромосом из строго определенного числа разных, но строго определенных хромосом. У всех высших и некоторых низших растений в течение их жизненного цикла чередуются два поколения: с клетками, содержащими в ядрах однократный (гаплоидный, п) набор хромосом, и с клетками, ядра которых имеют двойной (диплоидный, 2 п) набор хромосом. Гаплоидный набор состоит из п разных хромосом, по одной каждого типа. Численная величина строго постоянна для всех гаплоидных клеток всех растений данного вида. Так, для лесной земляники это число составляет 7, для гороха — тоже 7, но для фасоли — 11, для яблони — 17, для земляной груши — 51, для сахарного тростника — 60. Диплоидный набор — это два гаплоидных, сложенных вместе. В нем по две хромосомы каждого типа. Если у гороха в гаплоидном Наборе 7 разных хромосом, то в диплоидном 7 разных пар хромосом, причем две хромосомы в пределах каждой пары одинаковы. Легко сосчитать, что для перечисленных растений диплоидный набор (2п) составляет для земляники 14, для гороха — 14, для фасоли — 22, для яблони — 34, для земляной груши — 102, а для сахарного тростника — 120 хромосом.[ ...]
Все растения полученные от гибридизации Б-16 и Пирамидального с однолетними и озимыми сортами, были однотипны, выравнены в пределах каждой комбинации и имели в соматических клетках по 42 хромосомы.[ ...]
Растения с большим объемом хромосом гибнут при острой дозе ниже 1000 рад, тогда как растения с мелкими хромосомами или малым их количеством устойчивы к дозе 50 000 рад или выше. Такая зависимость свидетельствует о том, что при увеличении хромосомной «мишени» повышается вероятность прямого «попадания» атомных «выстрелов».[ ...]
Каждый вид растения содержит определенное число хромосом, известное как 2 п, или соматическое число (2 п= = 12 у шпината, 14 у гороха, 16 у лука, 18 у капусты, 20 у кукурузы, 22 у арбуза, 24 у томатов и т. д.). Репродуктивные клетки, или гаметы, содержат гаплоидное число (п) хромосом. В вегетативных клетках хромосомы присутствуют парами, составляя соматическое число. При мейозе одна хромосома из каждой пары переходит в гаметы, снижая диплоидное число наполовину. Впоследствии оплодотворение восстанавливает диплоидное число в зиготе (оплодотворенная яйцеклетка). Таким образом, в соматических клетках диплоидных растений каждый геи присутствует попарно. Отдельный ген, например С и его аллель с, может присутствовать в любом из трех сочетаний — СС, Сс или сс. Растение, содержащее два идентичных гена СС или сс, является гомозиготным по аллелям. Растение будет гетерозиготным, если аллели различны, например Сс.[ ...]
Гаплоидные растения высокостерильны, поскольку их хромосомная структура препятствует нормальному течению мейоза. В профазе мейоза в ядрах гаплоидов можно наблюдать образование петель в результате соединения сегментов одной и той же хромосомы, а также неправильное или частичное соединение различных хромосом (рис. 68).[ ...]
Поскольку растения содержат тысячи генов, но лишь ограниченное число хромосом, очевидно, каждая хромосома должна содержать много генов. Гены в каждой хромосоме сгруппированы не случайно, они имеют тенденцию наследоваться вместе. Это состояние известно как сцепление генов. Между физическим притяжением генов и интенсивностью или силой их сцепления имеется зависимость. Набор генов в хромосомах связан с подлинным обменом хромосомного вещества в мейозе.[ ...]
У бактерий, таких, как E. coli, хромосома содержит 10° пар нуклеотидов, что соответствует примерно 1000 генам, тогда как ядра растений и животных содержат гораздо большие количества ДНК. Следовательно, ДИК, содержащейся в ядре Vicia faba, достаточно для кодирования около 25-106 генов.[ ...]
Однако у многих раздельнополых растений, беспозвоночных, животных, рыб, птиц и всех млекопитающих различия между хромосомами мужских и женских особей связаны не с количеством хромосом, а с их качественным составом. Например, соматические клетки мужских и женских особей Б. те1аш аз1ег несут по четыре пары хромосом. Как у самцов, так и самок три пары хромосом являются одинаковыми и их называют аутосомами, но члены четвертой пары у самцов и самок неодинаковы по строению, ибо у самок оба члена этой пары являются прямыми палочковидными образованиями (Х-хромосомы), тогда как у самцов один член пары является прямым палочковидным образованием (Х-хромосома), а второй — изогнутым (У-хромосома). Все одинаковые хромосомы у самцов и самок называют аутосомами (А), тогда как хромосомы X и У получили название половых хромосом. Все яйцеклетки плодовой мушки несут четыре хромосомы (ЗА+Х), из которых три являются аутосомами, четвертая — Х-хромосомой. Напротив, сперматозоиды.также обладают четырьмя хромосомами, но они наполовину несут Х-хромосому (ЗА+Х) и наполовину — У-хромосому (ЗА+¥). Оплодотворение любой яйцеклетки сперматозоидом, обладающим Х-хромосомой, дает начало зиготе женского типа (6А+ХХ), тогда как оплодотворение любой яйцеклетки сперматозоидом, обладающим У-хромосомой, дает начало зиготе мужского типа (6А+ХУ).[ ...]
Совокупность генов, локализованных в хромосомах гаплоидного набора, называют геномом, нередко этим термином обозначают комплекс ядерно-генетических свойств клетки (организма). Число геномов, состоящих из различающихся по форме и величине гомологичных хромосом, можно определять по морфологическим признакам последних. Число хромосом — один из наиболее постоянных признаков при определении таксономического положения видов растений и животных. Закон специфичности числа хромосом был сформулирован впервые Т. Бовери в 1909 г. Начиная с этого времени морфологию хромосом стали использовать наряду с другими признаками в систематике. В некоторых случаях этим методом удавалось разрешить сложные таксономические проблемы.[ ...]
Необходимо отметить, что у большинства растений Е2(В1) мейоз имеет много нарушений в результате различных причин: общей перегрузки ядра и в таких условиях слабым проявлением гомеологичности геномов пырея Е1Е2. В тех случаях, когда многие хромосомы Е1Е2 остаются унивалентными в дополнение к унивалентам В1, они создают много нарушений, что ведет к образованию нежизнеспособных макро- и микроспор. В результате многие растения имеют очень слабую фертильность. Это простейший путь формирования растений типа многолетней пшеницы, которые имеют полный хромосомный комплекс пшеницы и дополнительно геном пырея, т. е. ААВВББЕЕ. Часто процесс формирования 56-хромосомных пшеничио-пырейных гибридов протекает более сложно. Особенно часты отклонения за счет недостаточно стабильной конъюгации хромосом геномов Е1Е2; иногда возникает недостаток или избыток этих хромосом. Но в конце концов с каждой новой генерацией в результате лучшей жизнеспособности гамет типа АББЕ! увеличивается и количество растений со стабильным октоплоидным числом хромосом.[ ...]
В большинстве случаев у видов двудомных растений женские растения в половой, паре имеют две идентичные хромосомы X гг X, мужские — гетерохромосомы X и Y, у двух типичных двудомных видов — конопли Cannabis sativa L. и шпината Spinacia oleracea L.— главные гены, определяющие сексуализацию, находятся в этих хромосомах [Frankel, Galmi, 1974]. Таким образом, генетический контроль определения пола у данных растений выявляется весьма четко и в настоящее время пи у кого по вызывает сомнений.[ ...]
Аномалии могут возникать при действии на растения и других химических и физических (радиационное, ультрафиолетовое, катодное и рентгеновское излучения) мутагенов. Большое число исследований посвящено изучению разрушительного и преобразовательного действия лучей Рентгена. Как известно, нормальный митоз отличается двуполярностью митотического аппарата, при применении же больших доз Рентгена отмечаются его монополярность и мультиполярность. В результате возникает асимметрия ядер, которая может достигать самых крайних пределов, когда диаметр одного ядра в несколько раз превышает диаметр другого. Добавочные ядра (микронуклеусы) образуются в результате пикноза, т. е. склеивания хромосом, который может быть частичным или полным. В случае полного пикноза хромосомы сливаются в бесформенные клубки и разрушаются. При неполном пикнозе в анафазе образуются мосты из хромосом, а также происходит неправильное их распределение (рис. 89).[ ...]
Рассмотрим это на примере с геномными формулами. Растения РгСВО дают гаметы, имеющие геномную структуру АВБЕВ2 (от 0 до 7). При оплодотворении их гаметой мягкой пшеницы с геномами АВВ возникают растения Р3(В2), которые имеют следующую геномную структуру: АА +ВВ + ВВ + Е(от 0 до 7) и В2 (от О до 7). Хромосомы геномов Е и В2, как не имеющие партнеров, будут элиминироваться уже при образовании гамет Р3. И в следующем, четвертом поколении многие растения будут иметь геномную структуру АА + ВВ + ВВ, т. е. это будут растения типа мягкой пшеницы, у которых еще содержатся единичные хромосомы геномов В2 и Е и поэтому сохранятся некоторые черты пырея. Но в дальнейшем и эти пырейные хромосомы будут элиминироваться. Пырейный наследственных! материал у пшенично-пырейных гибридов типа мягкой пшеницы может сохраняться в форме отдельных пырейных сегментов в пшеничных хромосомах, илит как теперь отмечают некоторые исследователи, отдельные пырейные хромосомы могут быть в пшенично-пырейных гибридах как дополнительные пли заменяющие пшенпчные хромосомы. Но во всех этих случаях растення относятся к типу однолетних мягких пшениц, а пырейные признаки у них проявляются в слабой форме.[ ...]
Вместе с тем данные генетических исследований на растениях и животных, а также на хромосомах слюнных желез Drosophila, где каждый диск, вероятно, соответствует одному гену, показывают, что число функциональных генов, кодирующих белки, не превышает нескольких тысяч. Представляется также удивительным, что очень близкие виды растений сильно различаются по содержанию ДНК. Так, клетки Vicia faba содержат в 7 раз больше ДНК, чем близкородственный вид V. sativa, хотя не ясно, зачем первому виду необходимо большее число генов.[ ...]
Политения обнаружена в семяпочках покрытосеменных растений. Политенные хромосомы найдены и изучаются в ядрах эндосперма, синергид и антипод представителей различных семейств растений. Они описаны в ядрах эндосперма некоторых видов семейства Роасеае (Zea mays, Hordeum vulgare), в эндосперме и синергидах рода Allium семейства Liliaceae, в эндосперме и клейких волосках завязи Bryonia dioica семейства Cucur-bitaceae, а также в подвесках представителей семейства Faba-сеае (род Phaseplus).[ ...]
Клеточные органеллы. Эти структуры представлены ядром, хромосомами, ядрышком, центриолями, митохондриями, рибосомами, лизосомами. Они характерны за некоторыми исключениями как для клеток животных, так и для клеток растений.[ ...]
В опытах Корреиса [Correns, 1906] со скрещиванием мужских и женских растений брионии двудомной (Brionia dioica) между собой и с растениями однодомной брионии (Brionia alba) была впервые доказана генетическая основа определения пола. Дальнейшие исследования в этом направлении привели к открытию половых хромосом, или гетерохромосом, обозначаемых как X- и Y-хромосомы [Морган, 1937]. Существование такого хромосомного механизма определения пола позволяет регулировать число мужских и женских особей у многих видов организмов.[ ...]
Основу исследований Г. Менделя, которые проводились, когда еще не были известны хромосомы, составляют скрещивания и изучение гибридов садового гороха. Г. Мендель начал исследования, располагая 22 чистыми линиями садового гороха, которые имели хорошо выраженные альтернативные (контрастирующие) различия между собой по семи парам признаков, а именно: форма семян (круглые —шероховатые), окраска семядолей (желтые — зеленые), окраска кожуры семян (серая — белая), форма бобов (выполненные — морщинистые), окраска незрелых бобов (зеленые — желтые), расположение цветков (пазушные — верхушечные), высота растения (высокие — карликовые).[ ...]
Изложенные материалы свидетельствуют о стерильности М 2 по мужской линии, подобно растениям, обладающим ЦМС. Но у нас нет оснований считать стерильность М 2 цитоплазматической, так как при создании этой пшеницы мягкая пшеница была опылена А. glaucшn, и, таким образом, хромосомы пырея были включены в цитоплазму пшеницы, а затем при беккроссе Е1 с пшеницей хромосомы пшеницы включались в цитоплазму гибрида, который своим источником имел цитоплазму пшеницы. Но несмотря на это, можно уже теперь с полной уверенностью предположить, что пырей, участвовавший в создании М 2, представляет интерес как новый источник получения форм с мужской стерильностью.[ ...]
В мейозе (последовательные деления клеток при формировании гамет) число хромосом уменьшается вдвое. Хромосомы в обычной вегетативной клетке высших растений, как правило, располагаются парами, образуя соматическое число. Обе хромосомы в паре (гомологи) морфологически подобны и содержат однотипные гены, хотя в результате некоторых отклонений (мутаций) гены внутри пары могут оказаться неидентичными. Генетическая изменчивость живых организмов обусловливается различными сочетаниями генов. Половой процесс — э о механизм, обеспечивающий такую перегруппировку и рекомбинацию генетических факторов, которая позволяет организмам приспосабливаться к изменяющимся условиям среды. Перегруппировка генетических факторов между и внутри хромосом совершается путем мейоза; рекомбинация завершается оплодотворением (см. гл. 10).[ ...]
Как было описано выше, первое поколение пшенично-пырейных гибридов имеет почти полное сходство с пыреем: растения: многолетние, мощные, их колос узкий, очень рыхлый с ломким стержнем. Для преодоления стерильности и окультуривания их колоса необходимо Р1 опылить искусственно пыльцой пшеницы, или фертильных пшенично-пырейных гибридов старших поколений, или предоставить Е1 свободному опылению указанных растений. Число завязывающихся зерновок у р! составляет доли процента с широким варьированием в зависимости от комбинации скрещивания, условий среды во время микро- и макроспорогенеза и цветения, а также от наличия в изобилии во время цветения соответствующей пыльцы у растений, окружающих первое поколение пшенично-пырейных гибридов.[ ...]
В клетках некоторых тканей при их развитии происходит незавершенный митоз: материал хромосом в ядрах удваивается, хромосомы делятся пополам, но, вместо того чтобы образовать два ядра, остаются в исходном ядре. С этого момента оно заключает в себе не диплоидный, а тетраплоидный (четверной) набор хромосом. Процесс, ведущий к подобному удвоепию хромосом внутри одного ядра, называется эндомитозом — внутренним митозом. Если он происходит с одним ядром дважды, то оно становится октоплоидпьтм (восьмикратным) набором и т. д. Клетки, ядра которых несут в себе больше двух наборов хромосом, называются полиплоидными, т. е. многоплоидными. Полиплоидия клеток в ряде случаев повышает их жизнеспособность, поскольку каждый ген дублируется несколькими другими такими же генами. Однокачественные гены действуют в унисон, и повреждение какого-нибудь из них пе ведет к выпадению определяемого им признака, так как компенсируется работой остальных однородных генов. Во многих случаях полиплоидные клетки крупнее и богаче содержимым, чем диплоидные. Выведены сорта полиплоидных культурных растений, которые обладают повышенными хозяйственными качествами.[ ...]
Открытие нерасхождения хромосом явилось окончательным доказательством в начале нашего века того, что гены локализованы на хромосомах. Однако теперь, когда с тех пор прошло много времени и когда к этому открытию возможен другой подход, можно сформулировать другое заключение. Фундаментальное значение открытия нерасхождения хромосом состоит в том, что оно положило начало изучению хромосомных мутаций и их механизмов. Можно добавить также, что оно означало объединение генетики и цитологии, т. е. формирование цитогенетики, оказавшейся в наше время исключительно плодотворной при изучении наследственности и наследственной патологии животных, растений и особенно человека (см. гл.[ ...]
Конъюгация хромосом обычно включает три хромосомы каждого типа (тривалентная конъюгация). При первом делении хромосом в мейозе две из них отходят к одному концу клетки, остальные к другому. Поскольку наборы хромосом тривалентов независимы между собой, число хромосом в формирующихся гаметах может составлять от п до 2п. Рассмотрим, например, шпинат, у которого п=6. В трип-лоиде (Зге = 18) в анафазе I мейоза возможны следующие типы набора хромосом: 6 — 12, 7 — 11, 8—10 и 9—9. При биномальном распределении образуется семь типов гамет, которые можно выразить числом хромосом (6, 7, 8, 9, 10, 11, 12), встречающиеся с частотой 1 : 6 : 15 : 20 : 15 : 6 : 1. Только 2/64 гамет содержат гаплоидное или диплоидное число хромосом (п или 2п). Поскольку число хромосом в растениях возрастает, частота гамет п и 2/г в триплоидах становится очень малой. Гаметофиты, имеющие несбалансированное число хромосом, или нежизнеспособны, или имеют низкую фертильность. Следовательно, триплоиды обычно полностью стерильны. Их можно размножать вегетативно (например, груша и яблоня) или ежегодно создавать вновь путем скрещивания тетраплоидов с диплоидами (например, бессемянный арбуз).[ ...]
Мейоз при образовании пыльцы и зародышевого мешка у гаплоидов протекает с большими нарушениями, что способствует стерильности гамет. У таких растений хромосомы в метафазе первого деления мейоза при микроспорогенезе остаются унива-лентами, располагаются неправильно и неравномерно расходятся в анафазе первого деления, вследствие чего образуются ядра с разными числами хромосом. Отмечены случаи возникновения реституционных ядер в результате выпадения редукционного деления и образования диад с диплоидным числом хромосом.[ ...]
Для выяснения вопроса о том, какие гаметы и с каким числом хромосом являются жизнеспособными, были проведены анализирующие скрещивания. Цветки растений первого поколения опыляли пыльцой мягкой пшеницы, у которой имеется определенное число хромосом (2и = 42). Полученные таким образом семена и затем растения Г2(В!) имели в соматических клетках 2гс = 63±1—2 хромосомы. Это подтвердило наше предположение » том, что жизнеспособными яйцеклетками являются те, которые-имеют реституционное ядро, т. е. ядро с нередуцированным числом хромосом, равным 42.[ ...]
Первое поколение гибридов, полученных от скрещивания A. glaucum с A. repens, по морфологическим признакам является промежуточным. Это очень мощные многолетние растения, высотой от 120 до 155 см с длинными ползучими корневищами, дающими большое число генеративных побегов (рис. 16). В соматических клетках гибрида, как и у обоих его родителей, содержится 42 хромосомы. Гибриды первого поколения частично плодовиты. В колосьях гибридов первого поколения встречается небольшое число цветков с нормально развитыми яйцеклетками. При просмотре колосьев гибридов иногда удается найти хорошо сформированные зерновки. Например, в 107 колосьях одного из гибридных растений Fi было найдено 20 зерен, завязавшихся, по-видимому, от свободного опыления пыльцой пыреев, растущих в питомнике в непосредственной близости от гибридов (рис. 17).[ ...]
С широким внедрением в сельское хоз-во гербицидов Появилась необходимость всестороннего их изучения и, в частности, изучения их влияния на наследственный аппарат растений, носителем кото poro являются хромосомы.[ ...]
Второе деление мейоза также протекает с большими отклонениями от нормы, особенно это сказывается на образовании микроядер, причем некоторые из них образованы многими хромосомами. Пыльца пустая, абортивная, но среди пустой пыльцы наблюдаются отдельные пыльцевые зерна, выполненные цито-тшазмой. При просмотре пыльцы под микроскопом в ацетокармине такие пыльцевые зерна хорошо красятся и в них можно видеть вегетативное ядро и два спермия. Но эти нормальные пыльцевые зерна остаются недеятельными, так как пыльники с малым количеством нормальной пыльцы кожистые и не растрескиваются. При вскрытии их пинцетом или препаровальной иглой и нанесении содержимого на рыльце цветков пшеницы завязываются семена, но в небольшом числе. Однако у отдельных гибридных растений иногда наблюдается растрескивание пыльников, в которых находится до 60% нормально выполненных цитоплазмой, хорошо окрашивающихся в ацетокармине пыльцевых зерен.[ ...]
Анализ длины главного колоса показал, что степень выраженности этого компонента продуктивности находится в зависимости от хромосомного состава 7 гомеологичной группы: в хромосоме 7В локализован ген (или гены), редуцирующий (е) длину колоса у растений. Выявлено положительное влияние на длину колоса 1А и 2А хромосом, однако наблюдаемый эффект статистически достоверен лишь по некоторым парам сравнения.[ ...]
На второй год жизни у гибридов, подвергавшихся действ; колхицина, кроме амфидиплоидных побегов, были и неизмеш ные, диплоидные. В соматических клетках амфидиплои содержится 84 хромосомы.[ ...]
Исключительное значение имеет биология цветения многолетних пшениц. Многие пшеннчно-пырейные гибриды, в том числе первые многолетние пшеницы, цветут открыто. И если с окружающих пшеничных растений пыльца попадает на рыльце цветков многолетней шпенпцы, происходит их оплодотворение. Таким образом, возникают гибриды между многолетней п мягкой пшеницей, у которых нарушена сбалансированность геномов и пырейные хромосомы элиминируются. Вместе с тем теряются и все пырейные признаки и, в первую очередь, многодетность. Поэтому перед нами стояла задача создать самоопыляющиеся многолетние пшеницы, чтобы при посеве в производственных условиях, где, как правило, большие площади заняты мягкой пшеницей, сорта многолетней пшеницы сохраняли бы все присущие ей черты. И такие самоопыляющиеся сорта нами получены.[ ...]
Геном эукариотов. Геном эукариотов по своим размерам больше генома E. coli лишь в 2-10 раз, однако ДНК в нем значительно больше. Установлено, что помимо «работающей» ДНК, т. е. помимо эксо-нов и интронов, в хромосомах клеток эукариот, в частности в гетерохроматиновых районах хромосом содержится много ДНК, которая является излишней, т. к. не транскрибируется. Такую ДНК называют сателлитной или «эгоистической (от англ. selfish — эгоистичный). Количество некодирующей ДНК у разных организмов является различным. Например, у дрожжей оно составляет 30% от общего количества ДНК, у насекомых — 67%, у цветковых растений (ара-бидопсис) — 69%, у нематод — 75%, у человека — 73-91%.[ ...]
Ни нуклеиновые кислоты, ни белки в отдельности не являются субстратами жизни. В настоящее время считают, что субстратом жизни являются нуклеопротеиды. Они входят в состав ядра и цитоплазмы клеток животных и растений. Из них построены хроматин (хромосомы) и рибосомы. Они обнаружены на протяжении всего органического мира — от вирусов до человека. Можно сказать, что нет живых систем, не содержащих нуклеопротеидов. Однако важно подчеркнуть, что нуклеопротеиды являются субстратом жизЯи лишь тогда, когда они находятся в клетке, функционируют и взаимодействуют там. Вне клеток (после выделения из клеток) они являются обычными химическими соединениями. Следовательно, жизнь есть, главным образом, функция взаимодействия нуклеиновых кислот и белков, а живым является то, что содержит само-воспроизводящую молекулярную систему в виде механизма воспроизводства нуклеиновых кислот и белков.[ ...]
Эндоспермальные гаустории способствуют повышению интенсивности обмена веществ во время роста и развития зародыша, так как облегчают приток питательных веществ в зародышевый мешок из окружающих тканей материнского растения. Ядра эндосперма и его гаусториев нередко становятся полиплоидными в результате следующих друг за другом эндомитозов. В них обнаружены также политенные хромосомы.[ ...]
Блексли установил, что каждый первичный мутант дурмана способен дать два вторичных мутанта, т. е. всего их может быть 24 (рис. 82). Третичные мутанты по той же формуле (2л+1) способны возникать лишь в тех случаях, когда лишняя хромосома образована сегментами, принадлежащими не гомологичным хромосомам, а хромосомам из разных пар. На рисунке 83 представлен набор хромосом дурмана с обозначением их половинок числами от 1 до 24. При этом третичный мутант может иметь лишнюю хромосому, возникающую путем транслокации между хромосомами (1, 2) и (17, 18), что приводит к новым комбинациям (1, 18) и (2, 17). Соответственно этому растения будут иметь набор хромосом, определяемый формулами [2я+ (1, /5)] и [2п+(2, 17)], что в действительности наблюдалось и было описано.[ ...]
Опубликовано много работ о передаче устойчивости против грибных заболеваний от пырея пшенице путем транслокаций у хромосом, принадлежащих разным родам. Однако во всех случаях исследователи применяли воздействие ионизирующими излучениями на гибридные растения для получения транслокаций между хромосомами, принадлежащими к разным родам (например, Triticale и Secale, Triticum и Agropyron и т. п.).[ ...]
И если обычные диплоидные растения шелковицы имеют в соматических клетках 28 хромосом (2га—28), то сейчас узко известии полиплоиды с 308 хромосомами (2л= ЗОВ).[ ...]
Полиплоидный ряд может быть образован как сбалансированными, так и несбалансированными типами хромосомных чисел (рис. 71). Большая плоидность среди растений встречается крайне редко. Приведенные обозначения характеризуют число хромосом в соматических клетках растений; так, у диплоидного колокольчика круглолистного (Campanula rotundifolia) в соматических клетках содержится 34 хромосомы, а у тетраплоидно-го — их 68.[ ...]
Именно так действуют антидоты, активирующие защитные системы кукурузы. В однодольных сорняках такая защитная система не действует, и они погибают. Прежде всего соединения этой группы подавляют митоз. Наиболее характерный симптом их действия — подавление роста проростков и деформация верхушек побегов. В метафазе хромосомы связываются с этими нитями и определенным образом ориентируются). При участии глутатиона БН-группы окисляются, образуя мостики — Б—Б.[ ...]
Раскрытие роли ДИК в передаче наследственных свойств представляется одним ил основных достижений современной биологии. Перечислим основные факты, подтверждающие, что именно это соединение является носителем паследстьеииой информации. 1. В половых клетках содержание ДНК в два раза меньше, чем в соматических. 2. Большая часть ДНК содержится в ядре и локализована в хромосомах. 3. ДИК представляет собой очень устойчивое соеднпекие. Вместе с тем воздействия, которые выпивают изменения в структуре ДНК (некоторые химические вещества, облучение и др.), изменяют наследуемые признаки организма. А. ДНК—единственное из известных веществ, способное к самовоспроизведению. 5. ДИК ответственна за изменение (трансформацию) оргапизмов. Это было доказано Эвери (1944) на эксперименте с двумя формами бактерий (пневмококков). Одна из пих обладала способностью образовывать капсулу п вызывать заболеваппе. Вторая форма не образовывала капсулы и не вызывала заболевапия. Оказалось, что после проникновения ДНК, выделенной пз вирулентных (вызывающих заболевание) клеток, некоторое количество клеток невкрулентпой формы образовало капсулу, причем эта способность передавалась по наследству. Естественно, на целых организмах высших растений такие опыты провести сложнее. Имеются лишь единичные результаты. Это был мутапт, потерявший способность образовывать антоциан. Цветки оказались розовыми, и эта способность передавалась по наследству. Опыты с высшими организмами единичны, однако сам факт возможности трансформации оргапизмов с помощью чужеродной ДНК пе вызывает сомнешш.[ ...]
Решающее прямое доказательство генетической рюли ДНК было обеспечено разработкой методов генной инженерии, создавшей возможность конструирования рекомбинантных молекул ДНК с заданными свойствами. К настоящему времени возможности генной инженерии показаны на примере клонирования многих генов самых различных организмов. Что касается косвенных доказательств, то они известны очень давно и их несколько. Для ДНК характерна специфичность локализации в клетках, поскольку она обнаруживается только в ядрах клеток (хромосомах), митохондриях (у животных) и хлоропластах (у растений). У многих микроорганизмов ДНК локализована только в ядерной области (нуклеоиде) или в цитоплазме в виде плазмид. Для организмов каждого вида характерно определенное количество ДНК на клетку (табл. 10).[ ...]
Значительные аномалии в течении мейоза обнаруживаются и в материнских клетках макроспор. Отличаясь неполным набором хромосом, они, естественно, нежизнеспособны и вскоре погибают. Возникновения вполне нормальных жизнеспособных спор у этих растений ожидать трудно, поскольку появиться они могут лишь в исключительных случаях, когда при первом делении мейоза все хромосомы группируются у одного полюса, в результате чего формируется ядро с гаплоидным числом хромосом. Изредка у гаплоидных растений можно наблюдать более или менее нормальное течение мейоза с образованием бивалентов.[ ...]
Итак, получить последующие поколения от гибридов многолетних пшениц с сортами озимой мягкой пшеницы легко, но эти гибриды не несут или почти не несут признаков пырея. Они по морфобиологическим признакам почти не отличаются от сортов озимой пшеницы и по образу жизни являются типично однолетними, относящимися к Т. аев1лушп. Только отдельные, главным образом, моногенные признаки иногда могут сохраняться от многолетней пшеницы. Поэтому улучшить колос многолетней пшеницы путем скрещивания ее с озимой пшеницей, как мы предполагали ранее, не представляется возможным. Это объясняется элиминацией пырейных хромосом, начиная с анафазы I у растений первого поколения. У гибридов второго и третьего поколения только в отдельных случаях можно наблюдать добавленные хромосомы к полному хромосомному комплексу пшеницы (2га = 42± ±1 — 2). У старших поколений этих гибридов сохраняются из пырейных признаков, присущих многолетней пшенице, лишь физиологические, которые детерминируются генами или блоками генов, находящихся в отдельных сегментах пырейных хромосом, включенных в пшеничные. Многодетность, являющаяся полиген-ным признаком, не может быть обеспечена единичными сегментами пырейных хромосом, включенными в пшеничные у рассматриваемых 42-хромосомных гибридов.[ ...]
И все же известно, например, что у кукурузы имеются гены, очевидно обладающие некоторыми чертами регуляторных генов. Так, локус генома кукурузы, названный Dissociation Dis), по-видимому, находится под контролем другого локуса Activator Ас) и в отсутствие последнего не может функционировать. Например, в некоторых случаях Dis обусловливает такое поведение гена С, детерминирующего окраску алейронового слоя в зернах, как если бы это был его рецессивный аллель, с, и алейроновый слой становится бесцветным. Таким образом, Dis, очевидно, подавляет активность гена С. На основании этих и других наблюдений Мак-Клииток предположила, что хромосомы высших растений содержат как гены, так и «контроллеры», регулирующие активность генов. ...[ ...]
ru-ecology.info
| Тупайя обыкновенная | Tupaia | 60 | Ю. Азия | |
| Тупайя филиппинская | Urogale | 44 | о. Минданао. Тупайеобразные | |
| Лемур серый | Hapalemur griseus | 54—58 | Мадагаскар. Лемуровые | |
| Лемуры обыкновенные | Lemur | 44—60 | Мадагаскар. 44, 46, 48, 52, 56, 58, 60 | |
| Лемур большой крысиный | Cheirogaleus major | 66 | Мадагаскар. Карликовые лемуры | |
| Лемуры мышиные | Mycrocebus | 66 | Мадагаскар | |
| Индри хохлатые | Propithecus | 48 | Мадагаскар | |
| Лори тонкие | Loris | 62 | Ю. Индия, Цейлон. Лориевые | |
| Лори толстые | Nycticebus | 50 | Ю. Азия. Лориевые | |
| Потто | Perodicticus | 62 | Африка | |
| Галаго сенегальский | Galago senegalensis | 38 | Африка. Галаговые | |
| Галаго толстохвостый | Galago crassicaudatus | 62 | Африка. Галаговые | |
| Долгопят западный | Tarsius bancanus | 80 | Суматра, Калимантан. Долгопяты | |
| Мирикина | Aotes trivirgatus | 54 | Ю. Америка | |
| Прыгун красный | Callicebus cupreus | 46 | Ю. Америка. Саковые | |
| Уакари красный | Cacajo rubicundus | 46 | Амазонка, Ориноко. Саковые | |
| Саки бледный | Pithecia pithecia | 46 | Север Ю. Америки | |
| Ревун рыжий | Alouatta seniculus | 44 | Ю. Америка Ревуны | |
| Ревун чёрный | Alouatta caraya | 52 | Ю. Америка. Ревуны | |
| Капуцин обыкновенныйКапуцин-фавн | Cebus capucinusCebus apella | 54 | Ю. Америка. Капуцины | |
| Саймири беличий | Saimiri sciureus | 44 | Север Ю. Америки | |
| Коата чёрнаяКоата Жоффруа | Ateles paniscusAteles geoffroyi | 34 | Север Ю. Америки. Коаты | |
| Обезьяны шерстистые | Lagothrix | 62 | Ю. Америка | |
| Мармозетка | Callimico goeldii | 48 | бассейн Амазонки | |
| Игрунка обыкновеннаяИгрунка желтоногая | Callithrix jacchusCallithrix flaviceps | 46 | Бразилия. Обыкновенные игрунки | |
| Игрунка золотистая | Leontideus rosalia | 46 | Бразилия | |
| Тамарин эдиповТамарин черноспинныйТамарин рыжий | Saguinus oedipusSaguinus nigricollisSaguinus illigeri | 46 | Ю. Америка. Тамарины | |
| Макаки | Macaca | 42 | Азия, С. Африка | |
| Павиан чёрный | Cynopithecus niger | 42 | о-в Сулавеси. Макаки | |
| Мангабеи | Cercocebus | 42 | Африка. Мартышковые | |
| Павианы | Papio | 42 | Африка | |
| Гелады | Terapithecus | 42 | Эфиопия | |
| Мартышки | Cercopithecus | 54—72 | Африка. 54, 58, 60, 62, 66, 68, 70, 72 | |
| Гульман | Pygathrix entellus | 50 | Ю. Азия. Тонкотелые обезьяны | |
| Носачи | Nasalis | 48 | Калимантан, 1 вид | |
| Орангутаны | Pongo | 48 | Суматра, Калимантан | |
| Шимпанзе | Pan | 48 | Африка | |
| Гориллы | Gorilla | 48 | Африка | |
| Сиаманги | Symphalangus | 50 | Ю. Азия | |
| Гиббоны | Hylobates | 44 | Ю. Азия. Кроме сиамангов | |
| Человек | Homo sapiens | 46 | Земля и частично космос | |
| Люцерна посевная | Medicago sativa | 32 | [8] | Cultivated alfalfa is tetraploid, with 2n=4x=32. Wild relatives have 2n=16.[8] |
| Американский барсук | Taxidea taxus | 32 | ||
| Американская куница | Martes americana | 38 | ||
| Американская норка | Neovison vison | 30 | ||
| Эквадорский рыбоядный хомячок | Anotomys leander | 92 | [9] | Один из самых больших хромосомных наборов среди млекопитающих |
| Резуховидка Таля | Arabidópsis thaliána | 10 | ||
| Ячмень обыкновенный | Hordeum vulgare | 14 | [8] | |
| Бобовые | Phaseolus sp. | 22 | [8] | All species in the genus have the same chromosome number, including P. vulgaris, P. coccineus, P. acutifolis, and P. lunatus[8]. |
| Канадский бобр | Castor canadensis | 40 | ||
| Каменная куница | Martes foina | 38 | ||
| Бенгальская лисица | Vulpes bengalensis | 60 | ||
| Гроздовник | Botrychium | 90 | ||
| Гроздовник прямой | Botrypus strictus | 88 | B. strictus and B. virginianus have been shown to be paraphyletic in the genus Botrypus | |
| Гроздовник виргинский | Botrypus virginianus | 184 | ||
| Капуста огородная | Brassica oleracea | 18 | [8] | Broccoli, cabbage, kale, kohlrabi, brussels sprouts, and cauliflower are all the same species and have the same chromosome number[8]. |
| Карп | Cyprinus carpio carpio | 104 | ||
| Кошка | Felis catus | 38 | ||
| Курица | Gallus gallus domesticus | 78 | ||
| Шиншиллы | Chinchilla lanigera | 64 | [10] | |
| Носухи | Nasua | 38 | ||
| Хлопчатник | Gossypium hirsutum | 52 | [8] | 2n=4x; Cultivated upland cotton is derived from an allotetraploid |
| Корова | Bos primigenius | 60 | ||
| Олений хомячок | Peromyscus maniculatus | 48 | ||
| Дельфин | Delphinidae Delphis | 44 | ||
| Осёл | Equus asinus | 62 | ||
| Голубь | Columbidae | 78 | [11] | Based on African collared dove |
| Муха-дрозофила | Drosophila melanogaster | 8 | [12] | 6 аутосом, 2 половые |
| Утконос | Ornithorhynchus anatinus | 52 | ||
| Земляные черви | Lumbricina | 36 | ||
| Вапити | Cervus canadensis | 68 | Подвид благородного оленя | |
| Барсук | Meles meles | 44 | ||
| Медоносная пчела | Apis mellifera | 32 | 32 у самок, самцы гаплоидны | |
| Европейская норка | Mustela lutreola | 38 | ||
| Лесной хорёк | Mustela putorius | 40 | ||
| Хорёк | Mustela putorius furo | 40 | ||
| Хвощ полевой | Equisétum arvénse | 216 | ||
| Куница-рыболов | Martes pennanti | 38 | ||
| Фосса | Cryptoprocta ferox | 42 | Хищное млекопитающее, обитающее на Мадагаскаре | |
| Жираф | Giraffa camelopardalis | 62 | ||
| Коза домашняя | Capra aegagrus hircus | 60 | ||
| Непарный шелкопряд | Lymantria dispar | 62 | ||
| Ястребинка | Hieracium | 8 | Род астровых | |
| Заяц[13][14] | Lepus | 48 | ||
| Африканские ежи | Atelerix | 90 | ||
| Евразийские ежи | Erinaceus | 88 | ||
| Червеколосник цейлонский | Helminthostachys zeylanica | 94 | Папоротник семейства ужовниковых | |
| Лошадь | Equus ferus caballus | 64 | ||
| Гиены | Hyaenidae | 40 | ||
| Крабоядный хомячок Питтье | Ichthyomys pittieri | 92 | [9] | Один из самых больших хромосомных наборов среди млекопитающих |
| Прыгающие муравьи | Myrmecia pilosula | 2 | [15] | 2 у самок. У самцов 1 хромосома (абсолютный минимум у организмов). У других муравьёв число хромосом больше.[15] |
| Кенгуру | Macropus | 12 | ||
| Лев | Panthera leo | 38 | ||
| Кузиманза | Crossarchus obscurus | 36 | Род мангустовых | |
| Кукуруза | Zea mays | 20 | [8] | |
| Мангифера индийская | Mangifera indica | 40 | [8] | Род манго |
| Сурикат | Suricata suricatta | 36 | Род мангустовых | |
| Комар | Aedes aegypti | 6 | [16] | 2n=6 у всех представителей семейства Culicidae, кроме вида Chagasia bathana, у которого 2n=8.[16] |
| Домовая мышь | Mus musculus | 40 | ||
| Мул | Mulus | 63 | Гибрид осла и кобылы. Стерилен | |
| Овёс | Avena sativa | 42 | [8] | Это гексаплоид с 2n=6x=42. Также культивируют диплоиды и тетраплоиды.[8] |
| Ужовниковые | Ophioglossum reticulatum | 1200 или 1260 | Семейство папоротниковых | |
| Восточная бескоготная выдра | Aonyx cinerea | 38 | ||
| Горох посевной | Pisum sativum | 14 | [8] | |
| Свиньи | Suidae | 38 | ||
| Голубь | Columbidae | 80 | ||
| Лесная куница | Martes martes | 38 | ||
| Ананас | Ananas comosus | 50 | [8] | |
| Картофель | Solanum tuberosum | 48 | [8] | Это тетраплоид; дикие формы чаще имеют 2n=24.[8] |
| Североамериканский дикобраз | Erethizon dorsatum | 34 | [10] | |
| Кролики | Leporidae | 44 | ||
| Енот-полоскун | Procyon lotor | 38 | [17] | |
| Енотовидная собака | Nyctereutes viverrinus | 42 | по разным источникам известно, что подвиды имеют 38, 54, и даже 56 хромосом | |
| Редис | Raphanus sativus | 18 | [8] | |
| Крысы | Rattus | 42 | ||
| Благородный олень | Cervus elaphus | 68 | ||
| Малая панда | Ailurus fulgens | 36 | ||
| Китайский мунтжак | Muntiacus reevesi | 46 | Вид оленей | |
| Рис посевной | Oryza sativa | 24 | [8] | |
| Рожь | Secale cereale | 14 | [8] | |
| Соболь | Martes zibellina | 38 | ||
| Чёрная антилопа | Hippotragus niger | 46 | ||
| Гроздовник | Sceptridum | 90 | Вид папоротника | |
| Калан | Enhydra lutris | 38 | ||
| Овца | Ovis aries | 54 | ||
| Креветка | Penaeus semisulcatus | 86-92 | [18] | |
| Клеточный слизевик | Dictyostelium discoideum | 12 | [19] | |
| Улитки | Gastropoda | 24 | ||
| Пятнистый скунс | Spilogale putorius | 64 | ||
| Морские звёзды | Asteroidea | 36 | ||
| Полосатый скунс | Mephitis mephitis | 50 | ||
| Болотный валлаби | Wallabia bicolor | 10/11 | Вид малых кенгуру, 10 у самцов, 11 у самок | |
| Тануки | Nyctereutes procyonoides albus | 38 | Вид енотовидной собаки | |
| Тигр | Panthera tigris | 38 | ||
| Табак | Nicotiana tabacum | 48 | [8] | Культурный вид тетраплоидный[8]. |
| Индейки | Meleagris | 82 | ||
| Виргинский опоссум | Didelphis virginiana | 22 | [20] | |
| Пшеница мягкая | Triticum aestivum | 42 | [8] | This is a hexaploid with 2n=6x=42. Durum wheat is Triticum turgidum var. durum, and is a tetraploid with 2n=4x=28[8]. |
| Белохвостый олень | Odocoileus virginianus | 70 | ||
| Мамонт | Mammuthus primigenius | 58 | Образцы из замороженных тканей | |
| Росомаха | Gulo gulo | 42 | ||
| Жёлтый мангуст | Cynictis penicillata | 36 | ||
| Дрожжи | 32 |
dic.academic.ru