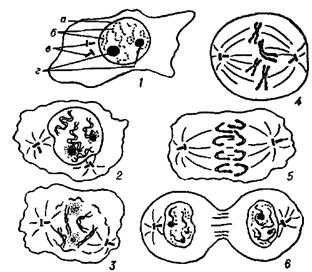
Выносливость растений к повреждению различных органов. Способность к росту у растений сохраняется до
Ответы@Mail.Ru: помогите срочно!! ! ПЛИЗ!!!
максим москалик Профи (586), на голосовании 4 года назадЗадание 1. Выберите один правильный ответ. 1. К грибам относится: А – аспергиллБ – навикулаВ – трахеломонасГ – эуструм. 2. Хламидомонада отличается от хлореллы наличием: А – клеточной стенкиБ – аппарата ГольджиВ – ядраГ – стигмы. 3. К трубчатым грибам относятся: А – сыроежкиБ – лисичкиВ – подосиновикиГ – шампиньоны. 4. У дрожжей вегетативное тело представлено: А – одноклеточным мицелиемБ – многоклеточным мицелиемВ – неподвижной шаровидной клеткойГ – подвижной клеткой со жгутиками. 5. Способность к росту у растений сохраняется: А – до определенного возрастаБ – в течение всей жизниВ – до достижения репродуктивного возрастаГ – до достижения определенных размеров. 6. Трахеды входят в состав: А – ксилемыБ – проводящих тканейВ – флоэмыГ – верно А и Б. 7. Клетки энтодермы корня: А – всасывают водуБ – не пропускают водуВ –проводят водуГ – верно А и Б. 8. Вечнозеленые растения Беларуси: А – вереск обыкновенныйБ – грушанка круглолистнаяВ –копытень европейскийГ – верно А и Б. 9. У земляники плод: А – многокостянкаБ – многоорешекВ – костянкаГ – ягода. 10. Травы бывают: А – только однолетниеБ – одно- дву- и многолетниеВ – однолетние и двулетние11. Двулетнее дикорастущее растение: А – лопух большойБ – мать-и-мачехаВ – одуванчик12. Какие признаки характерны для сем. Сложноцветные: А – соцветие кисть, плод семянкаБ – одиночные цветки, плод ягодаВ – соцветие корзинка, плод зерновкаГ – соцветие головка, плод семянка. 13. Какие грибы вызывают гниль картофеля: А – мучнисторосяныеБ – фитофтораВ – головневыеГ – ржавчинные. 14. К масличным растениям не относятся: А – клещевинаБ – горчицаВ – рапсГ – лютик. 15. Корневища образуют: А – ландыш майскийБ – пырей ползучийВ – кукурузаГ – А и Б. 16. Сальвиния относится к: А – водорослямБ – мхамВ – папоротникамГ – покрытосеменным. 17. Семенная кожура развивается из: А – покровов семязачаткаБ – стенок завязиВ – цветоложа и цветоножкиГ –верно Б и В. 18. Рамфоринкус – это: А – гриб-паразитБ – летающий динозаврВ – улотриксовая водоросльГ – эпифитный лишайник. 19. Назовите животное, не относящееся к ракообразным: А – дафнияБ – циклопВ – мокрицаГ – дрейсена20. Переносчик возбудителя клещевого энцефалита: А – таежный КЛЕЩБ – чесоточный клещВ – паутинный клещ. За каждый верный ответ 1 балл. Задание 2. Выберите правильные суждения. 1. Анабена – нитчатая цианобактерия. 2. Паренхима – это ткань, которая покрывает молодые части растения. 3. В генеративных почках находятся только зачатки цветков. 4. Семя состоит только из зародыша и запаса питательных веществ. 5. Столбчатая ткань теневого листа развита лучше, чем столбчатая ткань светового листа. 6. Корневой чехлик покрывает всю зону деления. 7. Кисть – соцветие, в котором цветки крепятся при помощи цветоножек на главной оси в очередном порядке. 8. Все мхи – листостебельные растения. 9. Саргассум относится к бурым водорослям. 10. Гаметофит папоротника называется заросток. За каждое верно выбранное суждение 2 балла. Задание 3. Ответьте на вопрос. Существует пословица: «Чем больше, тем лучше» . Можно ли получить богатый урожай, если посадить семена свеклы, моркови погуще? Ответ обоснуйте. (10 баллов)
Голосование за лучший ответ
Похожие вопросы
Также спрашивают
otvet.mail.ru
84. Локализация роста у растений
Характерной чертой ростовых процессов растительных организмов является их локализация в определенных тканях - меристемах. Меристемы различны по расположению в отдельных органах. При повреждении у растений активируются раневые меристемы, за счет которых регенерируют ткани, органы и целые организмы. Апикальные, или верхушечные, меристемы расположены в окончаниях (верхушках) стебля и корня. Вставочные, или интеркалярные, меристемы характерны для стебля (рост междоузлий у злаков) и для некоторых листьев. Латеральные меристемы обеспечивают рост стебля в толщину (камбий, феллоген). Клетки меристемы делятся; дочерние клетки достигают размеров материнской и снова делятся. Однако размер и объем меристем остаются постоянными. Это связано с тем, что большинство меристематических клеток через несколько делений (4-5) переходят к росту растяжением. Однако есть инициальные клетки, которые делятся в течение всей жизни органа. Показано также, что апикальные меристемы корня и стебля состоят из двух типов клеток, резко отличающихся по функциям и по способности к делению. Меристематические клетки, расположенные на самом верху стебля или корня, не прекращают делиться в течение всего периода роста. Эту зону меристематических клеток называют покоящимся центром (для корня) или меристемой ожидания (для стебля). Более длительная способность к делению является следствием меньшей частоты делений и большей длительности интерфазы. Одновременно данные клетки характеризуются и большей длительностью митотического цикла. Вместе с тем эти клетки более устойчивы к неблагоприятным воздействиям. Так, в них реже возникают хромосомные аберрации, что очень важно для сохранения жизнеспособности организма. Клетки меристемы ожидания стебля менее дифференцированы, что облегчает их последующую дифференциацию и имеет значение при переходе к образованию генеративных органов. Уже на первой фазе роста — фазе деления — клетки, находящиеся в нижней части меристемы, начинают дифференцироваться. В них постепенно накапливаются физиологические, а затем и морфологические различия. Вопрос о причинах дифференциации клеток - один из наиболее сложных вопросов физиологии. Все клетки данного организма обладают одинаковым геномом, следовательно, все клетки должны обладать и одинаковыми потенциальными возможностями (тотипотентностью). Тотипотентность проявляется, в способности дифференцированных растительных клеток, превращаться в эмбриональные (дедифференцированные) и при определенных условиях, давать начало целому организму. В частности, тотипотентность клеток проявляется в широко распространенной способности растительного организма к восстановлению утраченных частей - регенерации. Хорошо известным примером регенерации является образование придаточных корней при черенковании растений. Черенками называется любая часть растения, стебля, корня, листа, которая, будучи отделена; способна дать новую особь. После отделения черенка клетки, прилегающие к поверхности среза, начинают усиленно делиться, и образуется наплыв, или каллюс. Вначале каллюс - это недифференцированная ткань, а затем в ней обособляются проводящие элементы и закладываются корневые зачатки, из которых развиваются придаточные корни. Однако наиболее ярко тотипотентность клеток проявляется при культуре изолированных тканей.
studfiles.net
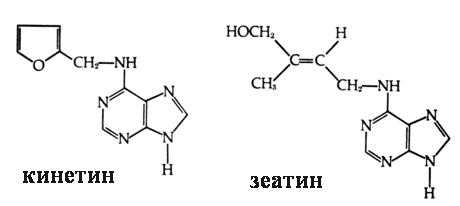
| АрхеологияАрхитектураАстрономияАудитБиологияБотаникаБухгалтерский учётВойное делоГенетикаГеографияГеологияДизайнИскусствоИсторияКиноКулинарияКультураЛитератураМатематикаМедицинаМеталлургияМифологияМузыкаПсихологияРелигияСпортСтроительствоТехникаТранспортТуризмУсадьбаФизикаФотографияХимияЭкологияЭлектричествоЭлектроникаЭнергетика | В основе роста и развития целого организма и отдельной клетки лежит обмен веществ. В процессе жизни каждого организма происходят постоянные качественные и количественные изменения, прерываемые периодами покоя. Необратимое количественное увеличение структур, объема и массы живого тела и его частей получило название роста. Развитие – это качественные изменения организма. Рост и развитие тесно связаны между собой, оба процесса регулируются на клеточном уровне. Рост органов и всего организма слагается из роста его клеток. Основные этапы роста, а также и развития на клеточном уровне – деление клеток и их растяжение, то есть увеличение клеточного потомства и увеличение их размеров. В многоклеточных организмах одним из показателей роста будет увеличение числа клеток в результате клеточного деления. Растительная клетка способна к росту растяжением, чему содействуют особенности строения ее оболочки. Особенности роста различны у разных систематических групп организмов. У высших растений рост тесно связан с деятельностью меристем. Рост, так же как и развитие, контролируется фитогормонами - химическими соединениями, вырабатываемыми в малых количествах, но способных давать значительный физиологический эффект. Фитогормоны, выработанные в одной части растения, транспортируются в другую часть, вызывая там соответствующие изменения в зависимости от генной модели воспринимающей клетки. Известны три класса фитогормонов, действующих, по преимуществу, как стимуляторы: ауксины (индолилуксусная, нафтилуксусная кислоты) (рис. 5.6), цитокинины (кинетин, зеатин) (рис. 5.7 ) и гиббереллины (С10 – гибериллин). Два класса гормонов (абсцизовая кислота и этилен) оказывают тормозящее действие (рис. 5.8). Заметное воздействие на рост и развитие растений оказывают ведущие факторы среды: свет, тепло и влага. Комплекс факторов и фитогормонов действует либо независимо, либо взаимодействуя друг с другом. Рис. 5.6. Структурные формулы ауксинов .
Рис. 5.7. Структурные формулы цитокининов
Рис. 5.8. Структурная формула абсцизовой кислоты
Интенсивность роста существенным образом связана с питанием растений, особенно с азотным и фосфорным. Типы роста различных органов определяются характером расположения меристем. Стебли и корни растут верхушками, они имеют апикальный рост. Зона нарастания листьев часто находится у их основания и они имеют базальный рост. Характер роста органа зависит от видовой специфичности. У злаков, например, рост стебля осуществляется у основания междоузлий, преобладает интеркалярный рост. Важная особенность роста растений – его ритмичность (чередование процессов интенсивного и замедленного роста). Она зависит не только от изменений внешних факторов среды, но и контролируется внутренними факторами (эндогенно), закрепленными в процессе эволюции. В целом рост растения складывается из четырех фаз: начальной, интенсивного роста, замедления роста и стационарного состояния. Это связано с особенностями различных стадий онтогенеза (индивидуального развития) растений. Так, переход растения к репродуктивному состоянию обычно сопровождается ослаблением активности меристем. Процессы роста могут прерываться продолжительными периодами торможения, наступление которых в северных широтах связано с концом лета и приближением зимы. Иногда у растений наблюдается как бы остановка роста – состояние покоя. Покой у растений – это такое физиологическое состояние, при котором резко снижаются скорость роста и интенсивность обмена веществ. Оно возникло в ходе эволюции как приспособление для переживания неблагоприятных условий среды в разные периоды жизненного цикла или сезона года. Покоящееся растение устойчиво к морозам, жаре, засухе. В состоянии покоя могут находиться растения (зимой, во время засухи), их семена, почки, клубни, корневища, луковицы, споры. Семена многих растений способны к длительному покою, обусловливающему их длительную сохранность в почве. Известен случай вызревания растения из семени одного из бобовых, пролежавшего в условиях вечной мерзлоты 10000 лет. В состоянии покоя находятся, например, клубни картофеля, благодаря чему они долго не прорастают. В понятие «развитие» вкладываются два смысла: индивидуальное развитие отдельного организма (онтогенез) и развитие организмов в ходе эволюции (филогенез). Физиология растений занимается изучением, главным образом, развития в онтогенезе. Меристематические клетки тотипотентны (омнипотентны) – любая живая клетка может дать начало недифференцированным клеткам, способным развиваться самыми различными путями (рис. 5.9 ). Переход меристематической клетки к росту сопровождается появлением в ней вакуолей и их слиянием в центральную вакуоль, растяжением клеточных оболочек.
Рис. 5.9. Тотипотентность меристематической клетки. Производные клетки: 1 – паренхима, 2 – эпидерма, 3 – флоэма, 4 – членик сосуда ксилемы, 5 – трахеида ксилемы, 6 – склеренхимное волокно, 7 – идиобласт, 8 – колленхима, 9 – хлоренхима.
Наиболее важный момент в развитии клеток высшего растения – их дифференцировка, или специализация, то есть возникновение структурной и функциональной разнокачественности. В результате дифференцировки образуются специализированные клетки, присущие отдельным тканям. Дифференцировка осуществляется как во время растяжения, так и после окончания видимого роста клеток и определяется дифференциальной активностью генов. Дифференцировки и рост контролируется фитогормонами. Развитие отдельных органов у растения получило название органогенеза. В целом цикле генетически обусловленное образование морфологических структур в онтогенезе называется морфогенезом. Внешние факторы, или факторы среды, также оказывают заметный эффект на рост и развитие. Свет оказывает глубокое влияние на внешнее строение растений. Свет влияет на дыхание и прорастание семян, образование корневищ и клубней, формирование цветков, на листопад, переход почек в состояние покоя. Растения, выращенные при отсутствии света (этиолированные), обгоняют в росте растения, выросшие на свету. Интенсивное освещение нередко усиливает процессы дифференцировки. Для каждого растения существует температурный оптимум роста и развития. Температурные минимумы роста и развития в среднем лежат в интервале 5-15 ° С, оптимумы – при 35° С, максимумы – в пределах 55° С. Низкая и высокая темпера-тура может нарушать покой семян, почек, сделать возможным их прорастание и распускание. Образование цветков – это переход из вегетативного состояния в генеративное. Индуцирование (ускорение) этого процесса холодом, называют яровизацией. Без процесса яровизации многие растения (свекла, репа, сельдерей, злаки) не способны к цветению. Огромное значение для роста, прежде всего, в фазе растяжения, имеет обеспеченность водой. Недостаток воды приводит к мелкоклеточности, отставанию в росте. Перемещение растений в пространстве имеет ограниченный характер. Для растений свойственно, прежде всего, вегетативное движение, связанное с особенностями роста, развития и обмена веществ. Одним из примеров движения служит фототропизм – направленная реакция искривления, вызываемая односторонним освещением: при росте побеги и черешки листьев искривляются в сторону света. Многие процессы обмена веществ, роста, развития и движения подвержены ритмическим колебаниям. Иногда эти колебания следуют смене дня и ночи (циркадные ритмы), иногда связаны с длиной дня (фотопериодизм). Пример ритмических движений – ночное закрывание или открывание цветков, опускание и продольное складывание листьев, раскрытых и приподнятых в дневное время. Такие движения связаны с неравномерным тургором. Эти процессы контролируются внутренней хронометрической системой – физиологическими часами, по-видимому, существующими у всех эукариотов. У растений важнейшая функция физиологических часов – регистрация длины дня и, вместе с тем, времени года, что определяет переход к цветению или подготовку к зимнему покою (фотопериодизм). Виды, растущие на севере (севернее 60° с. ш.), должны быть преимущественно длиннодневными, поскольку их короткий вегетационный период совпадает с продолжительной длиной дня. В средних широтах (35-40° с. ш.) встречаются растения как длиннодневные, так и короткодневные. Здесь весеннее- или осеннецветущие виды относятся к короткодневным, а цветущие в разгар лета – к длиннодневным. Фотопериодизм имеет большое значение для характера распространения растений. В процессе естественного отбора у видов генетически закрепилась информация о длине дня своих местообитаний и об оптимальных сроках начала цветения. Даже у растений, размножающихся вегетативно, длина дня определяет соотношение между сезонными изменениями и накоплением запасных веществ. Виды, индифферентные к длине дня, являются потенциальными космополитами и нередко они цветут с ранней весны до поздней осени. Некоторые виды не могут выходить за пределы географической широты, определяющей их способность к цветению при соответствующей длине дня. Фотопериодизм важен и в практическом отношении, поскольку он определяет возможности продвижения южных растений на север, а северных – на юг. Одним из важных процессов, осуществляющихся в ходе индивидуального развития, является морфогенез. Морфогенез (от греческого «морфе» – вид, форма), то есть становление формы, образование морфологических структур и целостного организма в процессе индивидуального развития. Морфогенез растений обусловливается непрерывной активностью меристем, благодаря чему рост растения продолжается в течение всего онтогенеза, хотя и с разной интенсивностью. Процесс и результат морфогенеза определяются генотипом организма, взаимодействием с индивидуальными условиями развития и закономерностями развития, общими для всех живых существ (полярность, симметрия, морфогенетическая корреляция). Вследствие полярности, например, верхушечная меристема корня производит только корень, а апекс побега – стебель, листья и репродуктивные структуры (стробилы, цветки). С законами симметрии связана форма различных органов, листорасположение, актиноморфность или зигоморфность цветков. Действие корреляции, то есть взаимосвязи разных признаков в целостном организме, сказывается на характерном для каждого вида внешнем облике. Естественное нарушение корреляций в ходе морфогенеза приводит к различным тератологиям (уродствам) в строении организмов, а искусственное (путем прищипки, обрезки) – к получению растения с полезными для человека признаками. В онтогенезе растение претерпевает возрастные изменения от эмбрионального состояния до генеративного (способного давать потомство путем образования специализированных клеток бесполого или полового размножения – спор, гамет), а затем – до глубокой старости. Выделяют 2 группы цветковых растений по типу репродуктивных процессов: монокарпики и поликарпики. К первой группе (монокарпики) относят однолетники, часть многолетников (бамбуки), которые цветут и плодоносят только один раз в жизни. Ко второй группе (поликарпики) принадлежат многолетние травы, древесные и полудревесные растения, способные плодоносить многократно. Онтогенез цветкового растения от возникновения зародыша в семени до естественной смерти особи подразделяют на возрастные периоды – этапы онтогенеза. 1. Латентный (скрытый) – покоящиеся семена. 2. Прегенеративный, или виргинильный, – от прорастания семени до первого цветения. 3. Генеративный – от первого до последнего цветения. 4. Сенильный, или старческий, – с момента потери способности к цветению до отмирания. В пределах этих периодов различают этапы. В группе виргинильных растений выделяют проростки (P), недавно появившиеся из семян и сохраняющие зародышевые листья – семядоли и остатки эндосперма. Ювенильные растения (Yuv), несущие еще семядольные листья, и следующие за ними ювенильные листья – более мелкие и иногда по форме еще не вполне похожие на листья взрослых особей. Имматурными (Im) считают особи, уже потерявшие ювенильные черты, но еще не вполне оформившиеся, полувзрослые. В группе генеративных растений (G) по обилию цветущих побегов, их размерам, соотношению живых и мертвых частей корней и корневищ различают молодые (G1), средневзрослые зрелые (G2) и старые генеративные особи (G3). Для высших растений очень важны процессы органогенеза. Под органогенезом понимают формирование и развитие основных органов (корня, побегов, цветков). Каждому виду растений свойствен свой темп заложения и развития органов. У голосеменных формирование репродуктивных органов, ход оплодотворения и развития зародыша достигают одного года (у ели), а иногда и больше (у сосны). У некоторых высших споровых, например у равноспоровых плаунов, этот процесс длится около 12-15 лет. У покрытосеменных процессы споро- и гаметогенеза, оплодотворения и развития зародыша происходят интенсивно, особенно у эфемеров (однолетних растений засушливых районов) – за 3-4 недели. Для цветковых растений установлен ряд этапов органогенеза. Главнейшие из них: дифференциация стебля, закладка листьев и побегов второго порядка; дифференциация соцветия; дифференциация цветка и образование археспория в семязачатках; мега- и микроспорогенез; мега- и микрогаметогенез; зиготогенез; формирование плода и семени. В онтогенезе организмов закономерно повторяются некоторые этапы развития, свойственные их отдаленным предкам (явление рекапитуляции). Впервые естественнонаучное объяснение рекапитуляциям дал Ч. Дарвин (1859). В 1866 г. Э. Геккель фактам повторения этапов филогенеза в онтогенезе придал форму биогенетического закона. В основе биогенетического закона лежит индивидуальное развитие особи (онтогенез), которое, в той или иной степени, представляет короткое и быстрое повторение важнейших этапов эволюции вида (филогенеза). Имеется множество примеров проявления биогенетического закона в мире растений. Так, протонема мхов, образующаяся на первых этапах прорастания споры, напоминает водоросль и свидетельствует о том, что предками мхов были, вероятнее всего, зеленые водоросли. У многих папоротников первые листья имеют дихотомическое (вильчатое) жилкование, которое было свойственно листьям ископаемых форм древних папоротников из среднего и верхнего девона. Зигоморфные цветки покрытосеменных при своем заложении проходят актиноморфную стадию. Биогенетический закон используется для выяснения особенностей филогенеза.
|
studopedya.ru
Читать книгу Рост и развитие растений
UntitledВ.В. Чуб
РОСТ И РАЗВИТИЕ РАСТЕНИЙ.
Оглавление
7.1. Общее представление о росте и развитии.
7.1.1. Параметры роста.
7.1.2. Кривая роста.
7.1.3. Дифференцировка.
7.1.4. Тотипотентность. Терминальная дифференцировка.
7.1.5. Периодизация индивидуального развития.
7.1.6. Проблема старения у растений.
7.2. Гормональная система растений.
7.2.1. Введение.
7.2.1.1. Регуляторные молекулы растений.
7.2.1.2. Рецепция и усиление сигнала.
7.2.1.3. Фосфатидил-инозитольная система вторичных мессенджеров.
7.2.1.4. Взаимодействие сигналов.
7.2.2. Ауксины - гормоны роста побега.
7.2.2.1. История открытия ауксинов.
7.2.2.2. Биосинтез и деградация ауксинов.
7.2.2.3. Транспорт ауксинов.
7.2.2.4. Физиологические эффекты ауксинов.
7.2.2.5. Ауксины и неоднородность внешней среды.
7.2.2.6. Ауксин и плоды.
7.2.2.7. Ауксин как гербицид.
7.2.2.8. Гравитропизм.
7.2.3. Цитокинины - гормоны корневого апекса.
7.2.3.1. История открытия.
7.2.3.2. Биосинтез и инактивация цитокининов.
7.2.3.3. Эффекты цитокининов от апекса корня до апекса побега.
7.2.4. Взаимодействие ауксинов и цитокининов.
7.2.4.1. Физиологическое действие ауксинов и цитокининов в культуре in vitro.
7.2.4.2. Баланс между ауксинами и цитокининами в интактном растении.
7.2.4.3. Цитокинины и паразиты растений.
7.2.5. Гиббереллины - гормоны листа.
7.2.5.1. История открытия.
7.2.5.2. Биосинтез гиббереллинов.
7.2.5.3. Основные физиологические эффекты гиббереллинов.
7.2.5.4. Гиббереллины и прорастание зерна.
7.2.5.5. Гиббереллин и проявление пола у растений.
7.2.5.6. Гиббереллин и цветение растений.
7.2.6. Абсцизовая кислота - сигнал водного стресса.
7.2.6.1. Окрытие абсцизовой кислоты.
7.2.6.2. Биосинтез абсцизовой кислоты.
7.2.6.3. Передача АБК-сигнала.
7.2.6.4. АБК-гормон осмотического стресса.
7.2.6.5. Регуляция покоя семян АБК.
7.2.6.6. Синдром дифицита АБК.
7.2.6.7. АБК и форма листьев.
7.2.7. Этилен - сигнал механического стресса.
7.2.7.1. Открытие физиологической роли этилена.
7.2.7.2. Биосинтез этилена.
7.2.7.3. Рецепция и передача сигнала.
7.2.7.4. Этилен как гормон механического стресса.
7.2.7.5. Этилен и прикосновение.
7.2.7.6. Этилен и заживление ран.
7.2.7.7. Регуляция листопада в умеренных широтах.
7.2.7.8. Формирование и созревание плодов.
7.2.7.9. Биотический стресс.
7.2.7.10. Этилен и цветение ананасов.
7.2.8. Другие гормональные вещества растений.
7.2.8.1. Брассиностероиды.
7.2.8.2. Жасминовая кислота.
7.2.8.3. Салициловая кислота.
7.2.8.4. Олигосахарины.
7.2.8.5. Короткие пептиды.
7.3. Рецепция световых сигналов.
7.3.1. Введение.
7.3.1.1. Основные принципы фоторецепции.
7.3.1.2. Физиологически важные области спектра. Фитохром и криптохром.
7.3.2. Фоторецепция в красной области спектра: фитохромная система.
7.3.2.1. История открытия фитохрома.
7.3.2.2. Фотоконверсия фитохрома. Фитохром А и фитохром В.
7.3.2.3. Этиоляция и деэтиоляция.
7.3.2.4. Избегание тени.
7.3.2.5. Регуляция прорастания семян.
7.3.2.6. Взаимодействие внутренних часов и фитохромная система.
7.3.3. Фоторецепция в синей области спектра: криптохром и фототропин.
7.3.3.1. История изучения фоторецепции в синей области спектра.
7.3.3.2. Криптохром - рецептор синего света, локализованный в ядре и цитоплазме.
7.3.3.3. Фототропин - мембранный рецептор синего света .
7.3.3.4. Суперхром - "кентавр" с головой фитохрома и туловищем фототропина.
7.4. Регуляция роста и развития растений.
7.4.1. Эндогенные факторы развития растений.
7.4.1.1. Образование листьев.
7.4.1.2. Переход к цветению.
7.4.1.3. Образование цветка.
7.4.1.4. Заключение.
7.4.2. Влияние внешних факторов на рост и развитие.
7.4.2.1. Пищевые ресурсы экотопа. Регуляция цветения элеменами минерального питания.
7.4.2.2. Фотопериодизм и климатические факторы.
7.4.3. Фотопериодизм.
7.4.3.1. История открытия фотопериодизма.
7.4.3.2. Опыты М.Х.Чайлахяна.
7.4.3.3. Гормональная теория цветения М.Х.Чайлахяна.
7.4.4. Термопериодизм. Стресс-периодизм.
7.4.4.1. Сезонная специализация растений.
7.4.4.2. Термопериодизм: стратификация и яровизация.
7.4.4.3. Опыты М.Х.Чайлахяна.
7.4.4.4. Стресс-периодизм.
7.5. Заключение
7.6. Физиология растений и прогресс сельскохозяйственных технологий.
7.6.1. Минеральные удобрения и зеленая революция.
7.6.2. Вторая зеленая революция.
7.6.3. Биотехнология.
7.6.4. Генная инженерия растений.
В настоящее время наблюдается дефицит учебной литературы по физиологии растений для ВУЗов. Последний раз большим тиражом учебник издавался в 1986 году, и многие сведения с тех пор успели устареть или существенно пополниться. В результате преподаватели МГУ не могут рекомендовать учебник, который соответствовал бы современным требованиям, а подготовка к экзамену исключительно по конспектам лекций имеет большие недостатки. Чтобы повлиять на создавшееся положение кафедра физиологии растений МГУ подготовила учебник, в который вошли традиционные разделы физиологии растений в современном изложении.
Начальные главы посвящены 1) клетке, 2) энергетическим процессам метаболизма, 3) фотосинтезу, 4) дыханию, 5) водному режиму, 6) минеральному питанию растений. Далее следует представленная глава "Рост и развитие растений", которая обозначена как "Глава 7". Заключают учебник главы по 8) устойчивости растений и 9) вторичному метаболизму.
Издание учебников - медленный процесс, который очень трудно корректировать и обновлять по мере подготовки к печати. Кроме того, очень существенны технические ограничения (например, объем издаваемой книги), которые не позволяют изложить материал достаточно полно, что приводит к конспективному "сухому" изложению. А конспект воспринимается намного хуже, чем полный текст.
Приемлемый выход из создавшейся ситуации - размещение учебных материалов на сайте кафедры. Глава 7 "Рост и развитие растений" является пробным шаром, и мы надеемся, что со временем здесь же будут размещены другие главы нашего электронного учебника. По мере обновления курса лекций главы будут расширяться и обновляться.
Материалами могут воспользоваться студенты при подготовке к семинарам и экзаменам, а также преподаватели, перед которыми стоит задача прочитать курс физиологии растений. Все материалы защищены авторским правом и не могут быть использованы в коммерческих целях.
Глава 7. РОСТ И РАЗВИТИЕ РАСТЕНИЙ.
7.1. Общее представление о росте и развитии.
7.1.1. Параметры роста.
В русском названии "растение" заключено фундаментальное свойство растений - способность к росту в течение всей жизни. Если растение не растет, это означает, что оно находится в стрессовых условиях или погибло. Таким образом, феномен роста сопровождает растение всю жизнь.
Казалось бы, о росте можно судить по увеличению общей (сырой) биомассы растения. Однако, сырая биомасса может не только увеличиваться, но и уменьшаться. Это случается в период засухи (запасающие воду ткани постепенно теряют воду). При выходе из стрессового состояния клетки
www.bookol.ru
Выносливость растений к повреждению различных органов
По К. И, Попову (1965, 1968), отчуждение листовой площади может приводить к уменьшению ассимиляционной поверхности, нарушению транспортных связей в передвижении ассимилятов по листу и между листьями и питающимися за их счет органами.
Основным звеном в цепи биохимических нарушений у растений при повреждении листьев является недостаток углеводов вследствие снижения их синтеза и возросшей траты на усилившееся дыхание. Это тормозит синтез белков и процессы роста растений.
Нарушения в работе листового аппарата отрицательно сказываются на корневой системе (корни испытывают недостаток в ассимилятах при обмене веществ). Растения при сильном повреждении листового аппарата испытывают не только углеводное, но и азотное голодание. У растений с хорошо развитым ассимиляционным аппаратом восстановительные процессы осуществляются на более широкой основе (В. Н. Любименко, 1921). Это происходит за счет повышения продуктивности фотосинтеза неповрежденных листьев и других зеленых органов растений, влагалищ, прилистников, прицветников и покровных тканей. У оставшихся после повреждения и вновь появляющихся листьев омолаживается структура хлоропластов. Они имеют более мелкозернистое строение, что увеличивает их удельную поверхность и Фотосинтетическую способность (А. А. Табнецкий, 1947; А. В. Ляшенко, 1960).
Повышение продуктивности фотосинтеза у поврежденных растений может также происходить благодаря повышению его интенсивности, увеличению размеров и массы листьев, а также продолжительности их активной жизни.
На завершающих этапах роста растений, когда идет формирование и созревание урожая, их выносливость к повреждению в большей мере зависит от сохранения листьев, обеспечивающих наиболее полный отток ассимилятов в обслуживаемые ими репродуктивные органы.
Как известно, корни выполняют функции поглощения воды и питательных веществ, закрепления растений в почве, а также аккумулируют резервные пластические вещества. Повреждение корней приводит к ухудшению общего состояния растений. Разные виды и сорта растений могут существенно различаться по выносливости к повреждению корневой системы. Наибольшее значение для выносливости растений имеют: 1) скорость роста и характер формирования корневой системы; 2) способность и скорость репродукции новых корней; 3) темпы заживления ран и устойчивость корней к гниению. Наиболее критическим периодом повреждения корней является период перехода всходов растений от гетеротрофного к автотрофному питанию, т. е, когда растения очень чувствительны к ослаблению функционирования еще слабо развитой настоящей корневой системы.
У кукурузы наибольшую выносливость к повреждению проволочниками (Elateridae) проявляют зубовидные сорта по сравнению с кремнистыми (К. Я. Калашников, И. Д. Шапиро, 1962). Эти различия обусловлены сортовым своеобразием формирования зародышевой и настоящей корневой системы у кукурузы в этот период.
Выносливость белокочанной капусты к повреждению личинками капустных мух зависит от темпов одревеснения центрального цилиндра (Е. Г. Рапопорт, 1979; Б. П. Асякин, Р. В. Пухаев, 1981) и от потенциальных возможностей растений формировать дополнительную корневую систему взамен утраченной. Анализ корневой системы растений, поврежденных личинками капустных мух, показал, что образование дополнительных корней в верхнем ярусе является активной реакцией растений на повреждения.
Стебли растений и особенно злаков становятся привлекательными для фитофагов благодаря наличию естественных полостей. Здесь личинки насекомых обеспечены относительно легкодоступной пищей и хорошо защищены от внешних неблагоприятных условий и энтомофагов. В то же время при проникновении личинок скрытностеблевых вредителей к местам питания создаются нередко специфические препятствия, обусловленные морфологическими или ростовыми барьерами растений. Так, личинки шведских мух для поселения во всходах зерновых культур должны преодолеть многие слои покровных тканей зачатков листьев, окружающих конус нарастания, и энергично происходящие в этой зоне ростовые процессы.
В соответствии с представлениями о регулирующей роли фитогормонов в онтогенезе растений, выносливость всходов злаков к повреждению шведской мухой можно себе представить следующим образом. После повреждения конуса нарастания усиливается приток гормона кинетина к месту повреждения. Это приводит к подавлению апикального доминирования главного стебля, в результате чего пробуждается боковая почка.
На следующем этапе к новому стеблю начинается приток гиббереллина (гормона роста стебля), который совместно с другим гормоном — ауксином — обеспечивает ускорение роста стебля. Гормоны усиливают поступление к новому побегу питательных веществ,
В отличие от шведских мух личинки гессенской мухи поселяются за влагалищами листьев. Наиболее часто они располагаются поблизости от места прикрепления влагалища к стеблю, т. е. на том участке, где дольше всего у междоузлия сохраняется способность к интеркалярному росту, и там, куда обеспечивается наиболее активный приток ассимилятов. На всходах личинки заселяют нижние междоузлия. Личинки летних поколений вредителя питаются в более высокорасположенных междоузлиях. В результате питания личинок растения приобретают интенсивно-зеленый цвет. Лист у поврежденного междоузлия становится более широким и коротким. Это, очевидно, происходит вследствие нарушения личинкой гессенской мухи соотношения гормонов в растении в сторону ослабления активности ауксина и усиления влияния кинетина.
При повреждении всходов пшеннцы личинками озимой (Delia coarctata) и пшеничной мух (Phorbia securis) степень выносливости растений в основном определяется способностью сортов к раннему компенсаторному кущению — формированию дополнительных стеблей.
Выносливость пшеницы и ячменя к хлебным стеблевым пилильщикам (Cephus pygmaeus L. и Tracheitis tafaidus F.) связана со степенью склеренхимизации проводящей системы к началу питания личинок (IX—XI этапы органогенеза растений), проходимостью личинок через междоузлия, а также с толщиной и прочностью соломины, которая позволяет удерживаться на корню подпиленным личинками стеблям.
У кукурузы выносливость к кукурузному мотыльку (Ostrinia nubilalis) в значительной мере основывается на устойчивости стебля и ножки початка к слому.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Как зависят рост и развитие растений от условий окружающей среды? — Науколандия
Рост и развитие растений зависят от условий, в которых они растут. Поскольку условия среды бывают разными, то и растения могут вырастать на разную высоту, иметь различные темпы развития. В определенных условиях растения могут проявлять свойства, которые не проявляют в других условиях.
Так семена растений могут прорастать только при наличии в почве влаги в достаточном количестве. А для размножения споровых растений необходима вода на поверхности почвы. Фотосинтез в растениях идет только на свету и у большинства растений только в теплое время года.
У растений в процессе их исторического развития выработалась периодичность в процессах роста и развития. Суточные ритмы, связанные с чередованием дня и ночи, выражаются в повышенной активности и замедлением процессов жизнедеятельности в зависимости от времени суток. Например, клетки образовательной ткани растений активно делятся днем. От времени суток также зависят фотосинтез, дыхание, испарение и другое.
Сезонная периодичность связана со сменой времен года. Неблагоприятные периоды растения обычно переживают в состоянии покоя. С сезонными ритмами связаны образование годичных побегов, активный рост в летом, сокодвижение весной, листопад осенью и др.
Если растения перенести в другие условия, то их суточная и сезонная периодичность сохраняются. Это значит, что она выработалась в процессе исторического развития растений и хранится в их наследственной информации.
Периодичность в жизни растений позволяет им быть хорошо приспособленными к климатическим особенностям среды их обитания.
Однако условия среды бывают разными не только в зависимости от времени суток и сезона. Из года в год похода в одно и то же время года может быть немного разной, местности, в которых растут растения, также могут отличаться между собой. Все это экологические факторы, оказывающие влияние на рост и развитие растений.
Большое влияние на растения оказывает свет, его яркость, продолжительность, периодичность. Так, например, в лесу, где свет плохо проходит сквозь кроны деревьев, у деревьев боковые ветви на стволе есть только вверху, а ниже ствол «голый». Однако деревья того же вида, но растущие в хорошо освещенных местах, ствол имеет боковые ветви почти на всем своем протяжении.
Если ветры в какой-то местности дуют примерно в одну и туже сторону, то у деревьев крона может как бы изгибаться в одну сторону, иметь больше ветвей с одной стороны.Рост и развитие растений сильно зависят от достаточного содержания в почве минеральных веществ. Например, дефицит азота замедляет рост растения.
На растения влияют животные, которые ими питаются. Обычно животные снижают скорость роста и развития растений. Однако многие животные необходимы растениям. Так насекомые переносят пыльцу с цветка на цветок и опыляют растения. Животные, которые питаются плодами и семенами, способствуют распространению растений.
Человек также оказывает влияние на рост и развитие растений. Внесение удобрений ускоряет рост и развитие растений, влияет на урожайность. Однако растениям, которые не являются сельско-хозяйственными, человек больше вредит. Вырубаются леса, загрязняется воздух и т. д.
Таким образом, на растения влияют совокупности абиотических (неживая природа), биотических (живая природа) и антропогенных (человек) факторов.
scienceland.info
способность к размножению
Способность к размножению все куриные, кроме глухаря, приобретают на первом году жизни. Продолжительность жизни невелика, представители большинства видов редко живут до 4—5 лет, глухари — дольше, до 10—12 лет. Все представители отряда — объекты любительской и промысловой охоты.[ ...]
Потенциальная способность к размножению у многих организмов огромна. Вспомним закон максимального давления жизни. У простейших в благоприятных условиях промежуток между последовательными делениями может сокращаться до нескольких минут. Гриб склеропора, паразитирующий на кукурузе, порождает до 6 млрд спор на одно растение в день. Однолетний мак производит до миллиона семян. Среди насекомых рекордсмен — матка термитов: она кладет по одному яйцу в секунду на протяжении всей жизни (у некоторых видов — до 12 лет). У рыб треска откладывает до 4 млн икринок в год, сельдь на протяжении жизни — от 8 до 75 млрд. У млекопитающих в одном помете от 1 (киты, слоны, приматы) до 20 особей у серой крысы.[ ...]
Часто растения обладают природной способностью к размножению отводками. У черной малины верхушки пониклых ветвей склонны к окоренению при соприкосновении их с почвой. У земляники усы представляют собой естественные отводки. Поскольку регенерирующий стебель прикреплен и продолжает получать питательные вещества от материнского растения, время и техника отведения не имеют такого большого значения, как при черенковании, когда вегетативная часть, которая должна будет регенерировать, полностью отделяется от материнского растения. Ускорить окоренение можно такими приемами, как поранение, кольцевание, этиолирование, изменение ориентации побега, оказывающими влияние на передвижение и накопление углеводов и ауксинов, стимулирующих возникновение корней.[ ...]
Когда растение становится потенциально способным к размножению, говорят, что оно стало взрослым. Бесспорным подтверждением наступления данной стадии является образование цветков. Однако у многих растений проявлению цветения предшествуют физиологические и морфологические изменения. Например, у плюща обыкновенного при достижении взрослой стадии заметно изменяется форма листа: лопастные листья, характерные для ювенильной стадии, становятся цельными. Последний лист, образующийся раньше цветочной почки, почти всегда редуцируется в прицветник.[ ...]
Большинство специализированных клеток не способно к размножению. Однако растение растет всю свою жизнь, и в течение всей жизни в нем образуются новые клетки. Они развиваются из клеток образовательных — меристематических — тканей. Размножение делением является специализацией меристематических клеток, их функцией в организме. Происшедшие из них клетки развиваются, превращаясь в те или иные специализированные клетки — клетки ситовидных трубок, клетки пробки, волокна и т. д. Образовательная ткань находится в разных частях растения (например, в точках роста — на верхушках побегов, корней). В толще стебля обычно есть несколько слоев образовательной ткани. Под слоем пробки находятся клетки феллогена, которые, делясь, пополняют число клеток пробки, корки. Внутрь от луба располагается слой клеток камбия. Те из порождаемых ими клеток, которые образуются снаружи от камбия, развиваются в клетки луба (рис. 25). Оказавшиеся внутри, т. е. ближе к оси ствола, дают начало клеткам древесины.[ ...]
Некоторые патогенные микроорганизмы сохраняют способность к размножению даже в воде водоемов, образуют споры. Выделяют следующие группы факторов, обусловливающих сроки выживания патогенных микроорганизмов в воде: 1) биологические особенности возбудителей заболевания; 2) количество попадающих в водоем микроорганизмов; 3) одновременное попадание в водоем биологического субстрата их естественного обитания; 4) особенности водоема; 5) температурный фактор; 6) комплекс гидрометеорологических факторов; 7) сопутствующая микрофлора и гидробионты (Л. В. Григорьева).[ ...]
В отличие от зигогенеза, многие животные организмы способны к размножению в естественных условиях путем партеногенеза (от греч. parthenos — девственница и genesis — рождение). Различают облигатный и факультативный партеногенез. Облигатный партеногенез — это размножение организмов из неоплодотворен-ной яйцеклетки. Такой партеногенез служит способом размножения животных более 90 видов, включая некоторых позвоночных. Примером облигатного партеногенеза является размножение кавказской скальной породы ящерицы, представленной только женскими особями. Напротив, факультативный партеногенез заключается в том, что яйцеклетки способны развиваться как без оплодотворения, так и после оплодотворения. Факультативный партеногенез в свою очередь бывает женским и мужским. Женский партеногенез част у пчел, муравьев, коловраток, у которых из неоплодотворенных яйцеклеток развиваются самцы. Мужской партеногенез встречается у некоторых изогамных водорослей.[ ...]
Полная послебрачная линька взрослых птиц, с потерей способности к полету, происходит в конце лета. Зимуют на юге Каспийского моря, порой улетая дальше на юг — до Африки, Ближнего Востока и Южной Азии. Способность к размножению приобретают в возрасте 3 лет. Максимальный известный возраст — 20 лет.[ ...]
Существенными адаптациями паразитов являются повышенная способность к размножению, выработка сложных жизненных циклов, использование переносчиков и промежуточных хозяев. Например, полупаразит Б1п§а (колдунья трава) широко распространен в Африке, Южной Азии, Австралии на зерновых культурах, сорго, сахарном тростнике, образует на одном растении до полумиллиона очень мелких семян. Семена паразитов весьма долговечны, что дает им возможность длительного «выжидания» контакта с растением-хозяином (семена 1п§а могут сохранять всхожесть в почве до 20 лет).[ ...]
Весь путь от оплодотворенной яйцеклетки до взрослого животного, способного к размножению и продуцированию, состоит из двух периодов: эмбрионального и постэмбрионального. Многие ученые называют их стадиями развития. Эмбриональный период животного организма включает в себя зародышевую, пред-илодпую и плодную фазы. Зародышевая фаза продолжается от оплодотворения яйцеклетки до полной имплантации эмбриона.[ ...]
Рождаемость (плодовитость) — число новых особей, появившихся за единицу времени в результате размножения. Живые организмы обладают огромной способностью к размножению. Она характеризуется т.н. биотическим потенциалом, представляющим собой скорость, с которой при беспрерывном размножении (возможном только теоретически при идеальных экологических условиях существования) особи определенного вида могут покрыть земной шар равномерным слоем. Это важнейший, хотя и условный показатель имеет самые различные значения. Так, для слонов он составляет 0,3 м/с, а для некоторых микроорганизмов — сотни метров в секунду. Удивительный факт: один одуванчик менее чем за 10 лет способен заселить своими потомками земную поверхность, если все семена прорастут (Р. Дажо, 1975). На практике такая громадная плодовитость никогда не реализуется.[ ...]
Поразительную картину изменения окраски можно наблюдать, у самцов бойцовой рыбки (Betta splend.es) в период размножения. Если в аквариум поместить 3—5 взрослых, способных к размножению самцов, то они в течение довольно длительного "времени мирно уживаются и не дерутся между собой. Окраска их тела достаточно разнообразная и красочная, но все же бледноватая. Плавники (спинной и хвостовой) повисшие, как бы смятые. Но картина резко меняется, если к ним поместить взрослую самку.[ ...]
Быстро возобновимые исчерпаемые ресурсы воссоздаются популяциями, имеющими большой биотический потенциал (видовая способность к размножению при отсутствии ограничений со стороны среды) и большую скорость роста (травяная растительность, животные).[ ...]
Эффект Ропартца: запах, исходящий от достаточно большой группы мышей,- вызывает у них гипертрофию надпочечников, что приводит к интенсификации биосинтеза кор-тикоидов и уменьшает их способность к размножению.[ ...]
В основе круговорота химических элементов лежит наличие в биосфере двух основных типов питания — аутотроф-ного и гетеротрофного, способность организмов аккумулировать солнечную энергию в другие формы энергии, а также способность живых организмов совершать химические реакции с такой скоростью, которая превышает скорость реакций в Неживой природе во много раз. Непрерывность и накопление результатов деятельности живых организмов в этом направлении обеспечивается их способностью к размножению. По мнению В.И. Вернадского, « ...жизнь захватывает значительную часть атомов, составляющих материю земной поверхности, Под влиянием ее эти атомы находятся в непрерывном Интенсивном движении. Из них все время создаются миллионы разнообразнейших соединений. И этот процесс длится без перерыва десятки миллионов лет, от древнейших, археозойских эр, до нашего времени. На земной поверхности нет химической силы, более постоянно действующей, а поэтому и более могущественной по своим конечным результатам, чем живые организмы, взятые в целом».[ ...]
Желаемые признаки очень часто обнаруживаются у растений, которые в остальных отношениях мало перспективны для садоводства. Устойчивость к парше свойственна таким видам яблони, которые образуют почти несъедобные плоды величиной менее 2,5 см. Кроме того, имеется много и других признаков, которые следует принимать во внимание. Для получения признания сорт земляники должен удовлетворять: 1) садовода — в отношении урожайности, срока созревания и устойчивости к болезням; 2) покупателя — внешним видом и качеством; 3) питомниковода — способностью к размножению; 4) грузоотправителя, продавцов и технологов.[ ...]
Определения», — это теоретически возможный верхний предел, которого популяция или способная к размножению часть популяции могла бы достигнуть при идеальных условиях. Легко себе представить трудности, связанные с определением этого термина, но он представляет интерес по двум причинам.[ ...]
Дифференциальные уравнения роста — приблизительные уравнения, так как они не учитывают различия между особями, сезонные колебания количества пищи, способности к размножению и т. п.[ ...]
Описанные модели роста популяции и дифференциальные уравнения предполагают, что все организмы сходны между собой, имеют равную вероятность погибнуть и равную способность к размножению, так что скорость роста популяции в экспоненциальной фазе зависит только от ее численности и не ограничена условиями среды, которые остаются постоянными. Идеальность» всех экологических факторов в исходных условиях предопределила то, что рассматриваемые модели называют идеальными.[ ...]
При рассмотрении плодовитости В следует расценивать как «общее число потомков, которое было бы произведено в отсутствие внутривидовой конкуренции», т. е. если каждая способная к размножению особь дала бы столько же потомков, сколько она давала бы в условиях без конкуренции. Практически В можно оценивать в популяции, испытывающей очень слабую конкуренцию — не обязательно в лишенной конкуренции). При рассмотрении роста В должно характеризовать общую биомассу или общее число модулей, которые появились бы, если бы все особи выросли в среде без конкуренции (а практически в среде с очень слабой конкуренцией). В этом случае А представляет собой общую биомассу или общее число действительно образованных модулей.[ ...]
На уровне особи абиотические факторы оказывают влияние прежде всего на поведение животных, изменяют трофические отношения со средой и характер метаболизма, воздействуют на способность к размножению и плодовитость, в значительной степени определяют развитие, скорость роста и продолжительность жизни животных и растений. Существует два основных способа адаптации особи к изменениям абиотических факторов: поведенческие реакции и физиологическая перестройка организма.[ ...]
Взрослые, как правило, возвращаются на свои прежние места гнездования. Молодые держатся с родителями всю зиму, возвращаются с ними на родину, но потом отделяются и вскоре летят к местам линьки. Способными к размножению они становятся на 3—4-й год жизни, некоторые — к 5—7-му годам. В возрасте 1—4 лет (обычно — в 2 года) многие молодые уже формируют пару и взрослеют дальше вместе, «помолвленными». Максимальный известный возраст — 17 лет.[ ...]
Животных обычно не держат до глубокой старости, так как со старостью они дряхлеют, теряют зубы, плохо переваривают и используют корм; довольно резко снижается их продуктивность, а способность к размножению утрачивается. Пользовательных животных в хозяйстве обычно держат более короткий срок, чем племенных, так как значительное снижение продуктивности (молочной, рабочей) наступает несколько раньше, чем потеря воспроизводительной способности. Таких животных выбраковывают из стада, как только их продуктивность, вследствие старости, снизится настолько, что дальнейшее содержание и использование их в хозяйстве станет экономически невыгодным. Племенных животных, особенно высокоценных, оставляют в хозяйстве, несмотря на значительное снижение их продуктивности, до тех пор, пока они способны воспроизводить хорошее потомство.[ ...]
Вышеизложенное говорит о том, что популяции присуще важнейшее свойство — саморегуляция. Она осуществляется действующими в природе двумя взаимно уравновешивающимися буферными силами: способностью к размножению и зависящей от плотности популяции реакции, напротив, ограничивающей воспроизводство.[ ...]
Зимой белые совы зимуют в тундре только при наличии корма, чаще откочевывают на юг за куропатками, нередко летят дальше, долетают до степей и пустынь, где странствуют до весны. Становятся способными к размножению в возрасте 2—3 лет, иногда гнездятся в неполном годовалом возрасте.[ ...]
Жизнь — одна из форм существования материи, закономерно возникающая при определенных условиях в процессе ее развития. Организмы отличаются от неживых объектов обменом веществ, раздражимостью, способностью к размножению, росту, развитию, активной регуляции своего состава и функций, к различным формам движения, приспособляемостью к среде.[ ...]
Вегетативное размно?кение халицистисов происходит путем образования дополнительных пузырей на одном ризоме или нескольких новых пузырей па одном старом. Пузыри легко отделяются от слоевища, сохраняя при этом тургор и способность к размножению.[ ...]
В популяции со сложной возрастной структурой представлены все возрастные группы, одновременно живут несколько поколений. Так, в стадах слонов, например, есть и новорожденные, и подростки, и молодые крепнущие животные, и способные к размножению самцы и самки, и старые особи. Такие популяции не подвержены резким колебаниям численности. Экстремальные внешние условия могут изменить их возрастной состав за счет гибели наиболее слабых, но самые устойчивые возрастные группы выживают и затем восстанавливают структуру популяции.[ ...]
Современная микробиологическая наука располагает достаточно обширным набором методов и приемов разделения живых и мертвых микроорганизмов. В основу этих методов положены следующие критерии жизнеспособности бактерий: 1) способность к размножению и образованию микроколоний на плотных средах; 2) ферментативная активность клеток; 3) изменение свойств клетки (проницаемости клеточной стенки, показателя преломления, восприятия того или иного красителя и т. д.). Однако задачу нельзя считать окончательно решенной даже в отношении суспензии чистой культуры микроорганизмов. Совершенно не разработана эта задача в отношении микроорганизмов природной воды, где исследователь имеет дело с самым разнообразным микробным биоценозом, до сих пор еще в полной мере не изученным. Оценка уже имеющихся методов разделения живых и мертвых бактерий позволяет выбрать для дальнейшей разработки приемы, наиболее подходящие для исследования микрофлоры воды.[ ...]
Большинство хлорорганических соединений до сих пор используются в качестве пестицидов. Хотя применяются эти вещества на суше, но ветры и реки быстро переносят их в океан. Помимо прямого действия на организм (снижение жизнестойкости, способности к размножению и т. д.), ксенобиотики вызывают изменения в наследственном аппарате, предвидеть которые весьма сложно. Однако особенно опасно то, что хлорорганические соединения почти не разлагаются и накапливаются в организмах, занимающих верхние уровни пищевой цепи (хищные рыбы, морские млекопитающие, рыбоядные птицы). Концентрация пестицидов в организмах морских животных в десятки и сотни тысяч раз превышает содержание этих веществ в морской воде. Часто это становится причиной их гибели.[ ...]
Весьма существенное влияние оказывают солнечные лучи на ход обмена веществ у рыб. Опыты, проведенные на гамбузии (Gambusia affinis Baird, et Gir.),. показали, что у гамбузий, лишенных света, довольно быстро развивается авитаминоз вызывающий, в первую очередь, потерю способности к размножению.[ ...]
Различные по своим свойствам когорты играют неодинаковую роль в жизни и динамике популяции. Долго живущие и поздно созревающие животные из осенних выводков обеспечивают переживание популяцией трудного по экологическим условиям зимнего периода, сохраняя способность к размножению. Именно эти зверьки составляют основу первого ранневесеннего цикла размножения и, дав потомство, довольно быстро отмирают. Их потомки —животные ранневесенних когорт,— отличаясь высокой ‘скоростью созревания, выполняют функцию максимально быстрого пополнения изреженной в течение нерепродуктивного периода популяции. Именно эти когорты представляют собой активную продуцирующую часть популяции и ответственны за темпы и величину нарастания численности и биомассы от весны к осени. К концу репродуктивного сезона большинство особей весенне-летних когорт отмирает; в зиму в основном уходит их потомство — зверьки, родившиеся во второй половине лета и осенью. Зимующая когорта начинает новый цикл.[ ...]
Максимальный температурный диапазон активной жизни чуть меньше диапазона жидкого состояния воды; для большинства многоклеточных организмов — от 0 до 50°С. Некоторые микроорганизмы, обитающие в горячих поверхностных и глубинных источниках могут сохранять способность к размножению при температуре до 85°С.[ ...]
Ориентация растения в пространстве оказывает заметное физиологическое влияние на рост и плодоношение. Плодовые деревья, посаженные наклонно под углом примерно 45°, становятся карликовыми и зацветают раньше. Такой же эффект дает горизонтальное размещение ветвей. К естественному снижению роста и увеличению цветения приводит пониклость скелетных веток, образующаяся под тяжестью обильного урожая. Таким образом, плодоношение действует как тормозной механизм, сохраняет у растений способность к размножению. Точного объяснения этому явлению пока не найдено, но полагают, что оно связано с нарушением нормального передвижения ауксинов, которое, в свою очередь, влияет на передвижение веществ по флоэме. Допускают, что на характер распределения ауксинов в стебле влияет сила тяжести.[ ...]
Действие ядовитых (токсичных) соединений на гидробионты проявляется в зависимости от их концентрации. При больших концентрациях ядовитых соединений наступает гибель гидробионтов, при малых — изменяются обмен веществ, темп развития, мутагенез (наследственные признаки), потеря способности к размножению и др. Так как отдельные популяции (например, зоопланктон) очень чувствительны к токсичным веществам, то уже небольшие концентрации последних вызывают гибель отдельных популяций, что влияет на биоценоз в целом.[ ...]
Нельзя полагать, что популяция — просто сумма отдельных особей, а ее свойства — лишь сложение свойств этих особей. У популяции в результате сложного взаимодействия входящих в нее организмов появляются только ей присущие свойства, которые совершенно не присущи отдельной особи (например, способность к размножению, а следовательно, к изменению численности и полового состава).[ ...]
Возрастная структура популяции является ее важной характеристикой, которая, как это было показано на примерах, рассмотренных в предыдущем разделе, оказывает влияние как на рождаемость, так и на смертность. Соотношение различных возрастных групп в популяции определяет, следовательно, ее способность к размножению в данный момент и показывает, чего можно ожидать в этом отношении в будущем. Обычно ¡в быстро растущих популяциях значительную часть составляют молодые особи, в стабильных популяциях распределение возрастных групп более равномерно, а в популяциях с уменьшающейся численностью больше старых особей. Однако возрастная структура популяции может меняться и без изменения ее численности. Для каждой популяции характерна некоторая «нормальная», или стабильная, возрастная структура, к которой направлено изменение ее реальной возрастной структуры. Коль скоро достигается стабильная возрастная структура, необычное увеличение рождаемости или смертности вызывает ее временное изменение, после чего происходит спонтанное возвращение к стабильному состоянию.[ ...]
Под развитием животного понимают процесс усложнения структуры организма, специализацию и дифференциацию его органов и тканей. Иными словами, развитие — это качественные изменения содержимого клеток, органообразовательные процессы, которые проходит каждый организм от оплодотворенного яйца до взрослого, способного к размножению и сходного в основных чертах с родительским организма.[ ...]
В природе и лабораторных условиях микроорганизмы могут подвергаться кратковременному воздействию высоких температур. Во время такого теплового воздействия клетки обычно не размножаются. После прекращения действия этого неблагоприятного для развития микроорганизма фактора одни штаммы могут сохранить репродуктивную способность (способность к размножению), другие оказываются менее устойчивыми и погибают.[ ...]
Рождаемость обычно выражают в виде скорости, определяемой путем деления числа вновь образовавшихся особей за определенный промежуток времени (йИп/М — абсолютная рождаемость) или числом новых особей в единицу времени на единицу популяции (ёШ/ЫсИ — специфическая, удельная рождаемость), где —размер популяции или только части, способной к размножению. Например, для высших организмов рождаемость выражают в расчете на одну самку, а для популяции человека — в расчете на 1000 человек.[ ...]
В законодательстве и литературе для обозначения объекта (или частей объекта) правового регулирования употребляются разные термины, иногда являющиеся смежными или подчиненными понятиями, иногда синонимами: генно-инженерно-модифицированный организм, трансгенный организм, продукты генной инженерии, продукция, содержащая результаты генно-инженерной деятельности, т. е. неживые объекты, не способные к размножению, но полученные генно-инженерным путем (лекарственные средства, корма и т. п.), микробиологические (биологические) агенты или токсины, не свойственные ественным экологическим системам организмы, искусственно созданные организмы и т. п. Ю. Р. Храмова, анализируя приведенные и другие понятия, указывает, что их имеет смысл различать применительно к основным типам рисков. Так, с трансгенными растениями связаны риски в результате: передачи чужеродных генов в природные растения за счет трансгенной пыльцы, т. е. при бесконтрольном ее переносе; поражения трансгенными организмами не тех вредителей, для уничтожения которых они были созданы; проявления токсических или аллергеноопасных свойств по отношению к человеку, животным или растениям. С животными связаны риски появления у них и в получаемых от них продуктах нежелательных физиологически активных веществ (гормонов), индуцирования новых вирусов и т. п.[ ...]
И наконец, несколько слов о динамическом хаосе в популяционной динамике. Используя дифференциальные (а также дискретные по времени разностные) уравнения для описания популяций и экосистем, мы тем самым принимаем и гипотезу о бесконечной делимости биомассы популяций, о существовании бесконечно малых количеств биомассы, которые, тем не менее, обладают всеми свойствами отдельной особи и, в частности, одним из основных свойств — способностью к размножению. С другой стороны, для мира живых систем характерна именно дискретность: существование биомассы в виде клеток, особей, популяций различных видов (для справедливости заметим, что и физический мир также дискретен — элементарные частицы, атомы, молекулы, кристаллы. . .). Как правило, мы закрываем глаза на это противоречие, поскольку ньютоновско-лейбницевский формализм непрерывного описания, созданный для классической механики и внутри нее непротиворечивый, дает вполне удовлетворительные результаты и при описании, например, популяционной динамики, когда число особей в популяции велико, а само это число меняется достаточно медленно. Не слишком четкие эти критерии, тем не менее, позволяют нам использовать дифференциальные (и разностные по времени) уравнения в моделях популяционной динамики в тех случаях, когда эта динамика более или менее регулярна. Но слишком задумываясь над обидами проблемами, мы использовали модели подобного типа и для описания динамического хаоса в популяциях и сообществах. Интуитивно кажется, что и в этом случае модели-будут давать удовлетворительное описание реальности. Но здесь возникают сомнения, на которые я не знаю, как ответить, но о которых необходимо сказать.[ ...]
Особый интерес для рыбоводства, как деликатесная рыба, вкусовые качества которой в несколько раз выше, чем форели, представляет севанская, или гокчинская, храмуля. Производителей этого вида храмули заготавливают в оз. Севан, нерестовых речках Мартуникского района и в Ар-данишской бухте. Искусственное воспроизводство севанской храмули освоено на рыбоводных предприятиях Грузии и Армении. Храмуля отличается невосприимчивостью к заболеваниям, предпочитает горные реки и озера, очень плохо скатывается из водохранилищ, поэтому представляет интерес для холодноводных бассейнов, прудов и других категорий водоемов. В водохранилищах храмуля выживает при температуре 26,4 °С, а зимой — 0,6—0,3 °С. Храмуля способна к размножению в возрасте 3—5 лет при достижении размеров самок 25—55 см (38,5 см), самцов—18—45 см, при массе 350—2100 (960 г) и 150—1500 г (650 г) соответственно. Производителей заготавливают в соотношении самок и самцов 1:1с помощью невода.[ ...]
Отношения по типу 2 могут быть причиной относительной стабильности популяции некоторых видов, в частности, они распространяются иа грибы под пологом леса. Их можно распространить и на некоторых животных из числа тех, кто питается падалью, а не нападает непосредственно на свою жертву. Для животных это отношение может быть не столь простым, если питающаяся популяция некоторым образом влияет на популяцию-пищу, но при этом не уменьшает ее способность к размножению и самосохранению. Конкуренция в этом случае возникает между особями питающейся популяции, которые используют только «процент», или излишки, не влияющие на поддержание устойчивости кормящего вида.[ ...]
Проведено три серии экспериментов: первая серия исследований - в апреле месяце до помещения приборов был снят фон микробного загрязнения воды выбранных колодцев; две серии исследований выполнены с интервалом экспозиции 1 и 2 месяца. Пробы воды исследовали по всем показателям СанПиН 2.1.4.559-96, а также использовали методику выделения «пин-пойнтс» (выращивание микроорганизмов методом заливки на питательном агаре при температуре 20° С). Определяли также Ps. aeruginosa как микроорганизм, способный к размножению в воде при повышении температуры воды, а также ряд вирусологических и паразитологических показателей.[ ...]
Экология изучает правую часть «спектра», изображенного на рис. 1.1, т. е. уровни биологической организации от организмов до экосистем. В экологии организм рассматривается как целостная система, взаимодействующая с внешней средой, как абиотической, так и биотической. В этом случае в наше поле зрения попадает такая совокупность, как биологический вид, состоящий из сходных особей, которые, тем не менее, как индивидуумы отличаются друг от друга. Они точно так же непохожи, как непохож один человек на другого, тоже относящиеся к одному виду. Но всех их объединяет единый для всех генофонд, обеспечивающий их способность к размножению в пределах вида. Не может быть потомства от особей различных видов, даже близкородственных, объединенных в один род, не говоря уже о семействе н более крупных таксонах, объединяющих еще более «далеких родственников».[ ...]
Независимый от плотности рост - это простейшая модель изменения численности популяции во времени, основанная на устранения из рассмотрения множества факторов, которые усложняют этот процесс в природе. Так, на динамику естественных популяций оказывают влияние две совокупности взаимодействующих процессов: рождаемость и иммиграция увеличивают число особей в популяции, тогда как смертность и эмиграция уменьшают его. Для того, чтобы упростить ситуацию, предположим, что: (1) процессы иммиграции и эмиграции уравновешивают друг друга так, что лишь рождение и гибель особей влияют на плотность популяции; (2) все особи идентичны друг другу, в особенности в отношении их способности к размножению и вероятности гибели; (3) популяция состоит лишь из партеногенетических самок, т.е. мы можем игнорировать все сложности, связанные с обоеполым размножением, и (4) ресурсы среды бесконечны и поэтому только врожденные способности особей к размножению и их смертность влияют на величину популяции. Эти предположения позволяют построить упрощенную модель роста популяции. Представляется очень полезным проанализировать подобную модель в двух вариантах в соответствии с двумя основными типами жизненных циклов организмов в природе.[ ...]
Пища чрезвычайно разнообразна, но основу составляют полевки, мыши и другие мелкие зверьки. При их низкой численности главным замещающим кормом чаще всего служат лягушки, а также ящерицы, змеи, птицы (чаще — птенцы), насекомые, черви, моллюски. Меню зависит от условий и от индивидуальных склонностей самих птиц. Основных способов высматривания добычи два. Часто канюки парят на высоте 30—50 м и выше, при ветре могут зависать на месте, почти не шевеля крыльями. Нередко занимаются подкарауливанием, сидя на столбах или деревьях. В лесу никогда не охотятся, разве что в совсем редком древостое. Улетают на юг в сентябре, некоторые задерживаются до октября. Дальность миграции невелика — до ближайших бесснежных или малоснежных краев. Способность к размножению приобретают в возрасте неполных 2 лет, часть птиц размножается в конце первого года жизни.[ ...]
Гнездо устраивают где-либо на балке, карнизе, на полу чердака, в других местах. Их размеры — 35—43 х 25—32 мм. Насиживают 17—19 дней, начиная с появления первого яйца, участвуют обе птицы, но больше самка. Птенцы в одном гнезде заметно различаются по размерам. Когда им исполняется 2 недели или несколько больше, самка уже может отложить новую кладку и садится ее насиживать, а птенцов докармливает самец. Маленьких птенцов родители кормят большими порциями 3—4 раза в сутки, подрастающих — всего дважды, обычно утром и вечером. Птенцы начинают летать в возрасте 4—5 недель и тогда же переходят к самостоятельному питанию. Способными к размножению становятся в возрасте 5—7 месяцев, самки несколько раньше самцов. Осенью и зимой большинство птиц не размножаются и держатся стаями, собираясь на ночевку на чердаках, крышах и карнизах.[ ...]
В роде стафилея И видов, довольно широко распространенных преимущественно в умеренных областях северного полушария. Наиболее известна стафилея перистая, или клекачка обыкновенная (S. pinnafca), произрастающая в Центральной и Южной Европе и в Закавказье. Небольшое дерево или кустарник, стафилея перистая весьма декоративна и успешно разводится в садах Европы с XVI в. Но гораздо шире распространена в культуре очень близкая и легко скрещиваемая с ней стафилея колхидская (S. colchica). Гибрид между этими видами известен в садоводстве под названием стафилеи изящной (S. ologans). В диком виде стафилея колхидская встречается в лесах Западного Закавказья, а также в Турции в окрестностях Трабзона. Недавно стафилея колхидская открыта и в Азербайджане на реке Гуручай. Кавказская приправа джоиджоли квашеные, маринованные и соленые соцветия, собранные в стадии бутонов,—по вкусу сравнима с каперсами. Семена стафилеи используют как лакомство (по вкусу напоминают фисташку, но обладают слабительным действием), из них выжимают масло. Кроме того, семена применяют в качестве украшений (бусы) и для изготовления четок. Такое интенсивное использование стафилеи, особенно стафилеи колхидской, привело бы к полному ее истреблению, если бы не ее способность к размножению корневыми отпрысками. Все же стафилея колхидская объявлена охраняемым растением. Из американских Видов упомянем стафилею трехлистную (S. trifolia, рис. 133), обитающую на Атлантическом побережье (эндемик Аппалачской провинции) и стафилею мексиканскую (S. mexicana).[ ...]
ru-ecology.info