
Доказательства эволюции: Переходные формы. Переходные формы растений
Переходная форма | Info-Farm.RU
Переходные (промежуточные) формы — организмы, которые сочетают в своем строении признаки двух больших систематических групп.
Переходные формы характеризуются наличием более древних и примитивных (в смысле первичных) черт, чем более поздние формы, но, в то же время, наличием более прогрессивных (в смысле более поздних) черт, чем их предки. Как правило, термин «переходные формы» употребляют по отношению к ископаемых форм, хотя промежуточные виды совсем не обязательно должны умирать.
Переходные формы используют как одно из доказательств существования биологической эволюции.
История понятия
В 1859 г.., Когда была издана работа Ч. Дарвина «Происхождение видов», количество ископаемых остатков была крайне малой, науке не были известны переходные формы. Дарвин описал отсутствие промежуточных форм «как наиболее очевидное и тяжелое возражение, которое может быть против теории», но объяснил это крайней неполнотой геологической летописи. Он отмечал ограниченное количество доступных коллекций в то время, в то же время описал имеющуюся информацию об имеющихся ископаемые образцы с точки зрения эволюции и действия естественного отбора. Только два года спустя, в 1961 году. Был найден археоптерикс, который представлял классическую переходную форму между пресмыкающимися и птицами. Его находках, стала не только подтверждением теории Дарвина, а также знаковым фактом, подтверждающий реальность существования биологической эволюции. С тех пор было найдено большое количество ископаемых форм, которые показывают, что все классы позвоночных животных являются родственными между собой, причем большинство из них — через переходные формы.
С увеличением сведений о таксономическое разнообразие сосудистых растений в начале ХХ в., Начались исследования по поиску их возможного предка. В 1917 г.. Роберт Кидстон и Уильям Генри Ленд обнаружили остатки очень примитивной растения возле поселка Rhynia в Шотландии. Это растение было названо Rhynia. Она сочетает в себе признаки зеленых водорослей и сосудистых растений.
Трактовка понятия
Переходные формы между двумя группами организмов не обязательно потомками одной группы и предком другой. По ископаемыми, как правило, невозможно точно установить является ли определенный организм предком другого. Кроме того, вероятность найти в палеонтологической летописи прямого предка определенной формы чрезвычайно мала. Гораздо больше вероятность обнаружить относительно близких родственников этого предка, которые сходны с ним по строению. Поэтому любая переходная форма автоматически интерпретируется как боковая ветвь эволюции, а не «участок филогенетического ствола».
Переходные формы и таксономия
Эволюционная таксономия оставалась доминирующей формой таксономии течение ХХ в. Выделение таксонов базируется на различных признаках, вследствие чего таксоны изображают в виде ветвей разветвленного эволюционного дерева. Переходные формы рассматриваются как «падающие» между различными группами в плане анатомии, они смесь характеристик от внутренней и внешней клади, что недавно разделилась.
С развитием кладистики в 1990-х гг. Взаимосвязи обычно изображают в виде кладограмы, иллюстрирующую дихотомическое ветвление эволюционных линий. Поэтому в кладистици переходные формы рассматриваются как более ранние ветви дерева, где еще не развились не все черты, характерные для ранее известных потомков на этой ветке. Такие ранние представители группы обычно называют основным таксоном (англ. Basal taxa) или сестринским таксоном (англ. Sister taxa), в зависимости от того, принадлежит ли ископаемый организм к данной клади или нет.
Проблемы выявления и интерпретации
Отсутствие переходных форм между многими группами организмов является предметом критики со стороны креационистов. Однако далеко не каждая переходная форма существует в виде окаменелостей из-за принципиальной неполноты палеонтологической летописи. Неполнота вызвана особенностями процесса фосилизации, то есть перехода в окаменевший состояние. Для образования окаменелости необходимо, чтобы организм, который погиб, был погребен под большим слоем осадочных пород. Из-за очень медленную скорость осадконакопления на суше, сухопутные виды редко переходят в окаменевший состояние и сохраняются. Кроме того, редко удается выявить виды, которые живут в глубинах океана через редкие случаи поднятия на поверхность больших массивов дна. Таким образом, большинство известных ископаемых (а, соответственно, и переходных форм) — это либо виды, обитающие на мелководье, в морях и реках, или наземные виды, которые ведут полуводный образ жизни, или живут у береговой линии. К упомянутым выше проблемам следует добавить чрезвычайно малую (в масштабах планеты) количество палеонтологов, которые осуществляют раскопки.
Переходные формы, как правило, не живут на больших территориях и не существуют в течение большого времени, иначе они были бы персистентный. Этот факт также снижает вероятность фосилизации и последующего обнаружения переходных форм.
Поэтому вероятность обнаружения промежуточных форм чрезвычайно мала.
Примеры среди животных
Древнейшими представителями земноводных считают ихтиостеги. Их считают переходным звеном между кистеперых рыбами и земноводными. Несмотря на то, что в ихтиостеги была пятипалая концовка, адаптированная к жизни на суше, значительную часть жизни они проводили как рыбы, имели хвостовой плавник, боковую линию и некоторые другие признаки рыб.
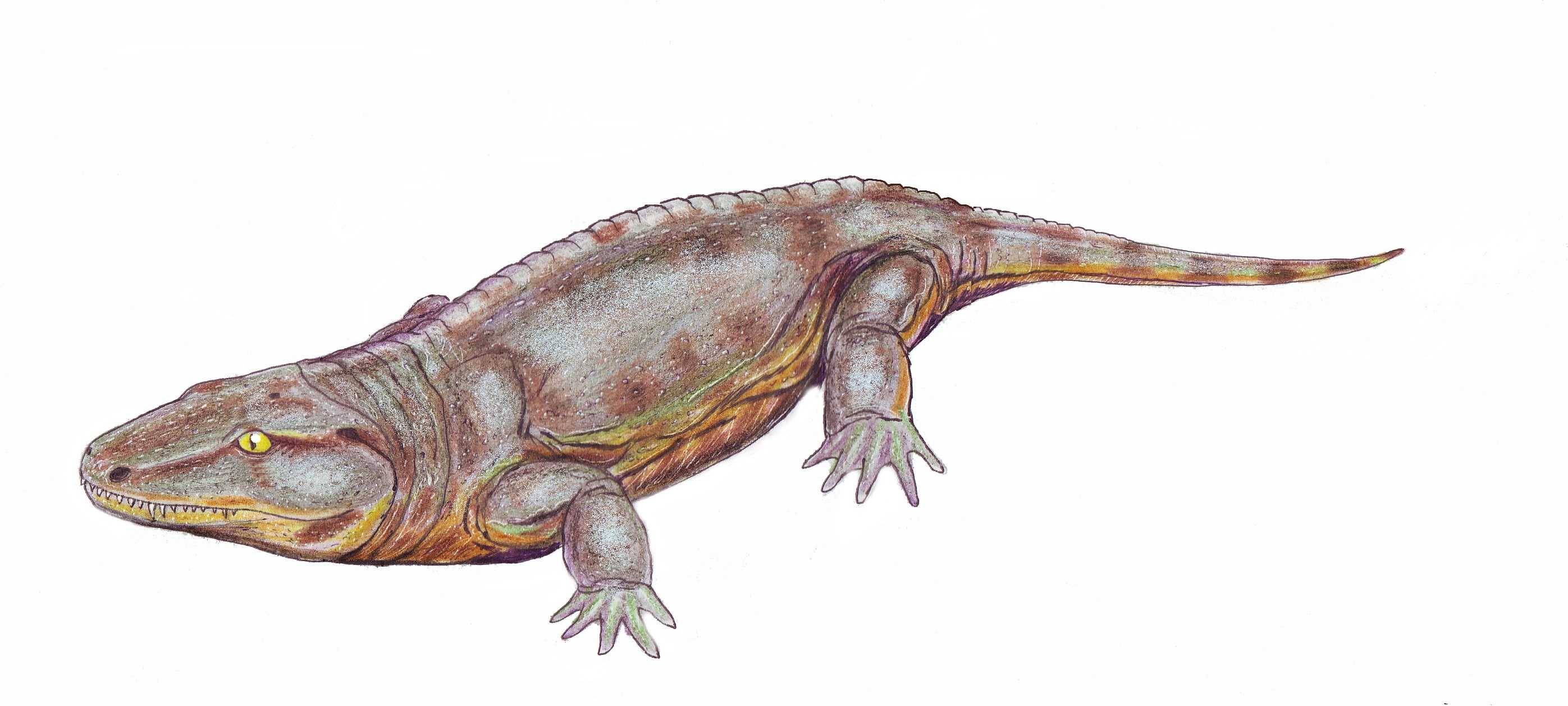
Батрахозавры, существовавшие в каменноугольный и пермский периоды, рассматривают как переходную форму между земноводными и пресмыкающимися. Батрахозавры, хоть и проводили жизнь во взрослой стадии на суше (подобно пресмыкающихся), были тесно связаны с водоемами и сохранили ряд признаков, присущих земноводным, в частности, откладывания икры и развитие личинок в воде, наличие жабр и тому подобное.
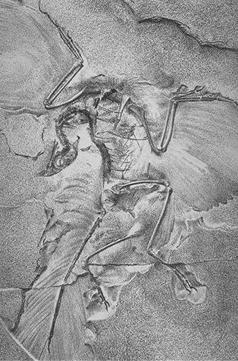
Обнаружено большое количество пресмыкающихся, которые выработали способность летать, часть из них имела перья, поэтому их рассматривают как переходные формы между пресмыкающимися и птицами. Наиболее известен археоптерикс. Он был размером примерно с современную ворону. Формой тела, строением конечностей и наличием оперения подобный современных птиц, возможно, летал. Общим с пресмыкающимися была особое строение таза и ребер, наличие клюва с коническими зубами, по три свободные пальцы на крыльях, подвийноувигнути позвонки, длинный хвост с 20-21 позвонка, кости могло не пневматизовани, грудная кость без киля. Другие известные переходные формы между пресмыкающимися и птицами — протоавис, конфуциусорниса.
Большое количество ископаемых форм звероподобных пресмыкающихся (синапсиды, терапсид, пеликозавров, различных динозавровых и др.), Найденных во многих районах земного шара, существовали в юрский и меловой периоды, сочетающие признаки пресмыкающихся и млекопитающих, раскрывают возможные направления и способы становления различных групп четвероногих , в частности млекопитающих. Например, звероподобный пресмыкающееся из группы терапсид — лиценопс (Lycaenops) по развитию костей ротовой полости, дифференцированием зубов на клыки, резцы, по резцовые зубы и рядом других признаков строения тела напоминает хищных млекопитающих, хотя по другим признакам и образом жизни это были настоящие пресмыкающиеся.
Одной из форм, сохранилась в ископаемом состоянии является амбулоцетус Ambulocetus natans («ходячий кит») — переходная форма между наземными млекопитающими и китообразными, которые являются вторинноводнимы формами. Внешне животное напоминало нечто среднее между крокодилом и дельфином. Кожа должна частично редуцированную шерсть. Животное имело лапы с перепонками; хвост и конечности приспособлены как вспомогательные органы передвижения в воде.
Примеры среди растений
Первые наземные растения из класса риниопсид, семей риниевих и псилофитовых, живших в силуре — девоне, сочетали признаки зеленых водорослей и примитивных форм высших растений. Их тело было безлистным, цилиндрический осевой орган — телом в верхней части дихотомически разветвленным на верхушках с спорангиями. Функцию минерального питания риниопсид выполняли ризоиды.
Ископаемые формы семенных папоротников, которые процветали в конце девона, сочетают в себе признаки папоротников и голосеменных. Они образовывали не только споры (как папоротники), но и семена (как семенах растения). Проводящая ткань их стеблей по строению напоминает древесину голосеменных (саговников).
Другой предшественник семенных растений был идентифицирован из отложений среднего девона. Рункария (Runcaria heinzelinii) существовала около 20 млн лет назад. Это была небольшая растение с радиальной симметрией; имела спорангий, окруженный интегументом и плюской. Рункария демонстрирует путь эволюции растений от споровых к семенных.
Переходные формы в эволюции человека
В наше время найдено большое количество ископаемых останков, которые раскрывают эволюционный путь человека разумного от ее человекообразных предков. К формам, которые в большей или меньшей степени можно отнести к переходным, относятся: сахелантропа, ардипитека, австралопитеки (африканский, афарский и другие), человек умелый, человек работающий, человек прямоходящий, человек-предшественник, гейдельбергский человек и кроманьонцы.
Среди упомянутых форм значительное внимание заслуживают австралопитеки. Австралопитек афарский с точки зрения эволюции находится между современными двуногими людьми и их четвероногими древними предками. Большое количество рис скелета этого австралопитека четко отражают двуногость, причем до такой степени, что некоторые исследователи считают, что это свойство возникла задолго до появления австралопитека афарского. Среди общих черт анатомии, его таз гораздо больше похож на этих костей у человека, чем у обезьян. Края подвздошных костей короче и шире, крестцовая кость широкая и расположена непосредственно позади тазобедренного сустава. Существует явное свидетельство о существовании мест крепления для мышц-разгибателей колена, предусматривает вертикальное положение этого организма. В то время, как таз австралопитека не совсем как у человека (заметно шире, с ориентацией края подвздошных костей наружу), эти особенности указывают на принципиальную перестройку, связанную с хождением на двух ногах. Бедренная кость образует угол в направлении колена. Эта черта позволяет ноге размещаться ближе к средней линии тела и является явным свидетельством привычный характер передвижения на двух ногах. В наше время человек разумный, орангутаны и коаты имеют такие же черты. Ноги австралопитека имели большие пальцы, что делает практически невозможным захват стопой ветвей деревьев. Кроме особенностей локомоции, в австралопитека был также значительно больше мозг, чем у современных шимпанзе и зубы были значительно больше подобными зубов современного человека, чем к обезьянам.
Филогенетические ряды
Филогенетические ряды — ряды ископаемых форм, связанные между собой в процессе эволюции и отражают постепенные изменения их исторического развития.
Были исследованы русским ученым А. Ковалевским и английским Дж. Симпсоном. Они показали, что современные однопалого копытные происходят от древних мелких всеядных животных. Анализ ископаемых лошадей помог установить постепенность процесса эволюции в пределах этой группы животных, в частности, как изменяясь во времени, ископаемые формы приобретали все большего сходства с современными лошадьми. Сравнивая эоценового еогипуса с современным конем, трудно доказать их филогенетическую родство. Однако наличие ряда переходных форм, которые последовательно сменяли друг друга на больших пространствах Евразии и Северной Америки, позволила восстановить филогенетический ряд лошадей и установить направление их эволюционных изменений. Он состоит из ряда следующих форм (в упрощенном виде): Phenacodus — Eohippus — Miohippus — Parahippus — Pliohippus — Equus.
Гильгендорф (1866) описал палеонтологический ряд брюхоногих моллюсков из миоценовых отложений, накопившихся в течение двух миллионов лет в озерных отложениях Штейнгеймського бассейна (Вюртемберг, Германия). Было обнаружено в последовательных слоях 29 различных форм, принадлежащих к ряду планорбис (Planorbis). Древние моллюски имели раковину в виде спираи, а более поздние — в виде турбоспирали. Ряд имел два ответвление. Предполагается, что изменение формы черепашки была вызвана повышением температуры и увеличением содержания карбоната кальция в результате горячих вулканических источников.
Таким образом, филогенетические ряды представляют собой историческую последовательность переходных форм.
В настоящее время известны филогенетические ряды для аммонитов (Вааген, 1869), брюхоногих моллюсков рода живородок (Viviparus) (Неймайром, 1875), носорогов, слонов, верблюдов, парнокопытных и других животных.
Изображения по теме
info-farm.ru
Переходная форма [Промежуточная] — это, что такое, какие, определение, значение, доклад, реферат, конспект, сообщение, вики — WikiWhat
Переходными (промежуточными) формами называются организмы, которые по строению объединяют в себе признаки различных классов. Другими словами, переходные формы выражают родственные связи между представителями животных и растений, относящихся к разным систематическим группам. Материал с сайта http://wikiwhat.ru
Кистеперые рыбы, которые жили в девонском периоде, являются промежуточной формой между рыбами и земноводными. Археоптерикс — промежуточная форма между пресмыкающимися и птицами. Некоторые представители терапсид являются переходными формами между пресмыкающимися и млекопитающими. А семенные папоротники считаются переходными формами между папоротниками и голосеменными растениями. Наличие переходных форм является убедительным доказательством постепенного изменения органического мира в процессе исторического развития.
Картинки (фото, рисунки)

Рис. 63. Промежуточные формы: 1 — ихтиостега; 2 — археоптерикс; 3 — терапсид
 На этой странице материал по темам:
На этой странице материал по темам: Реликтовые виды это доказательства палеонтологии?
Какие высокоорганизованые формы растений животных появились в мезозойской эре
Индивидуальная изменчивость растений
Промежуточная это что
Переходные формы из мира растений
 Вопросы к этой статье:
Вопросы к этой статье: Приведите примеры промежуточных форм, связывающих друг с другом различные группы животных и растений.
wikiwhat.ru
Доказательства эволюции: Переходные формы | Блог unknown hero
В целях ликбеза в рядах "стихийных" антиэволюционистов (т.е. не идейных - неизлечимых, а просто недоучившихся, с которыми не всё ещё потеряно) публикую на Конте отличную книгу "Доказательства эволюции" (2010 г., авторы: Борисов Н. М., Воробьев Ф. Ю., Гиляров А. М., Еськов К. Ю., Журавлев А. Ю., Марков А. В., Оскольский А. А., Петров П. Н., Шипунов А. Б. Под редакцией А.В.Маркова.). Поскольку её объём довольно велик, и мало кто осилит его сразу и целиком, публикую книгу по частям. Каждая из них представляет интерес. Сама книга (в HTML) доступна по ссылке. Желающие читать её в форматах FB2, ePub и др. могут найти соответствующие ссылки при помощи поисковых сервисов.
Группы доказательств:
Переходные формы
Общеизвестное доказательство эволюции — наличие так называемых промежуточных форм (переходных форм), то есть организмов, сочетающих в себе характерные признаки разных видов (или разных таксонов более высокого ранга - родов, семейств и т.д.). Как правило, говоря о промежуточных (или переходных) формах имеют ввиду ископаемые виды, хотя промежуточные виды вовсе не должны непременно вымирать. На основе филогенетического дерева теория эволюции предсказывает, какие промежуточные формы реально существовали (и поэтому могут быть найдены), а какие — нет. В соответствии с научным методом, сбывшиеся предсказания подтверждают теорию. Например, зная строение организмов пресмыкающихся и птиц, можно предсказать некоторые особенности переходной формы между ними. Мы прогнозируем возможность найти останки животных, подобных рептилиям, но с перьями, или останки животных, подобных птицам, но с зубами или с длинными хвостами. При этом мы предсказываем, что не будут найдены переходные формы между птицами и млекопитающими, например — ископаемые млекопитающие с перьями или подобные птицам ископаемые с костями среднего уха как у млекопитающих.
Вскоре после публикации «Происхождения видов» был обнаружен первый скелет археоптерикса — промежуточной формы между рептилиями и птицами. У археоптерикса было развито оперение (типичная птичья черта), а по строению скелета он слабо отличался от динозавров. У него были когти на передних конечностях, зубы и длинный костяной хвост, а характерных «птичьих» особенностей скелета было немного (крючковидные отростки на ребрах, вилочка). Позднее были найдены и другие переходные формы между рептилиями и птицами.
Антиэволюционисты, как правило, имеют крайне превратные представления о том, что должны представлять из себя обнаруживаемые в ископаемой летописи переходные формы. Например, они "требуют", чтобы переходная форма между А и Б была непременно промежуточной между А и Б по всем без исключения признакам. Если какие-то признаки переходной формы ближе к А, а другие - к Б, антиэволюционисты объявляют такую форму "не переходной, а просто мозаичной". Тем самым антиэволюционисты навязывают эволюционной теории такие проверяемые следствия, которые из нее вовсе не следуют. Эволюционная теория не утверждает, что все признаки организмов эволюционируют с абсолютно одинаковой скоростью. Неравномерность темпов эволюции - одно из общих мест современной эволюционной теории. Естественно, в ходе эволюционного перехода от А к Б одни признаки А становятся "как у Б" раньше, другие - позже. Нет никаких причин, почему бы это должно было быть не так. Иными словами, эволюционная теория как раз и предсказывает, что большинство переходных форм должны быть "мозаичными".
Другой часто встречающийся тип неправомерных "требований" антиэволюционистов строится по схеме: "По-вашему утки произошли от крокодилов, ну так предъявите нам крокоутку". Но современные утки не произошли от современных крокодилов. И те, и другие, произошли от общего предка, который жил давно (по-видимому, в пермском периоде, более 250 млн лет назад) и не был ни крокодилом, ни уткой. Он был, если уж на то пошло, древним примитивным архозавром. И от него произошли не только утки и крокодилы, но и все динозавры, а уже затем от динозавров произошли птицы. Общий предок есть у любых двух современных организмов, но этот предок, естественно, не является ни одним из них. Поэтому мы не можем и не должны никому предъявлять крокоуток. Однако, поскольку птицы реально произошли от динозавров, мы имеем возможность предъявить целый ряд самых настоящих птицединозавров (см. ниже).
Еще один способ борьбы антиэволюционистов с переходными формами - это "игра в слова". "Эта форма ближе к А? Значит это просто А. Ближе к Б? Ну тогда это просто Б". Такой эссенциализм поддерживается формальными правилами биологической классификации. Биологи, согласно этим правилам, не могут написать на этикетке "форма, переходная между видом А и видом Б". Максимум, что они могут сделать, это выделить переходную форму в отдельный вид, назвать его, скажем, А1, и подписать внизу мелкими буквами "вид, переходный между А и Б". Но антиэволюционисты не замечают этих приписок и говорят: "Какая же это переходная форма? Это просто вид А1".
Еще один тип антиэволюционистских передергиваний - требование доказать, что данная переходная между А и Б форма действительно является прямым предком Б. Это все равно что требовать строгих доказательств того, что найденный в Африке череп Омо I (см: 195 000 лет назад в Эфиопии жили «анатомически современные» люди) принадлежит самой Митохондриальной Еве, а не ее сестре или более дальней родственнице. Шансы найти в очень неполной палеонтологической летописи чьего-либо прямого предка очень малы, но даже когда таких предков находят, нет способов доказать, что это именно прямой предок, а не его близкий родственник. Реально находимые переходные формы являются формами, близкородственными искомому прямому предку (или его мало изменившимися потомками), требовать большего - значит опять-таки приписывать эволюционной теории не следующие из нее "следствия".
Превращение динозавров в птиц ("орнитизация динозавров-теропод")
Серия блестящих палеонтологических находок, сделанных в последние годы, пролила свет на многие детали эволюционного превращения динозавров в птиц. Как выяснилось, многие хищные динозавры-тероподы в течение юрского и мелового периодов эволюционировали "в птичью сторону". Только одна из этих эволюционных линий дала начало современным птицам и не вымерла 65,5 млн лет назад вместе с остальными динозаврами.
Ниже приведены краткие обзоры нескольких недавних исследований (чтобы узнать подробности, воспользуйтесь гиперссылками):
1) Перья сначала служили для красоты, а для полета пригодились позже. В 2008 году был найден пушистый нелетающий "птицединозаврик" с четырьмя очень длинными перьями на хвосте, как у современных райских птиц. Исследователи полагают, что длинные хвостовые перья служили исключительно «для красоты», то есть для привлечения брачного партнера. Самцы многих современных птиц красуются перед самками длинными хвостовыми перьями, больше ни на что не годными и только мешающими летать, причем самки отдают предпочтение тем женихам, у которых эти перья длиннее. Палеонтологи и раньше предполагали, что у некоторых пернатых динозавров крупные и непригодные для других целей перья могли служить для брачных демонстраций, но такого «явного» случая до сих пор не находили. Это исследование, наряду со многими другими, показало, что происхождение птичьих перьев - типичный пример смены функции органа в эволюции. Перья сначала развились для термоизоляции, и, возможно, для улучшения аэродинамических качеств динозавра при быстром беге; потом начали использоваться для брачных демонстраций, и, наконец, пригодились для планирующего, а затем и для машущего полета.
2) Малый размер птичьих геномов — наследие эпохи динозавров. Американские и британские палеонтологи в 2007 году показали, что по микроструктуре ископаемых костей можно судить о размере генома вымерших четвероногих. Оказалось, что одна из отличительных особенностей современных птиц — малый размер генома — была характерна для предков птиц (динозавров-теропод) с самого начала их истории. Это еще одна «птичья» черта, которая, наряду с перьями и некоторыми особенностями скелета, сформировалась у динозавров задолго до того, как они научились летать.
3) Предки птиц, возможно, были четырехкрылыми. В юрских отложениях провинции Ляонин (Северо-Восточный Китай) в 2009 году найден хорошо сохранившийся экземпляр оперенного "птицединозавра" Anchiornis huxleyi, жившего на несколько миллионов лет раньше археоптерикса. Находка показала, что предки птиц, по-видимому, изначально имели крупные контурные перья, пригодные для полета, не только на передних, но и на задних конечностях. Данная находка - еще один яркий пример сбывшегося эволюционного предсказания. Палеонтологи понимали, что должны были существовать полуптицы-полудинозавры, более древние и при этом более близкие к рептилиям, чем археоптерикс. Однако до сих пор все находки такого рода были более молодыми (эксперименты по "орнитизации" среди динозавров продолжались еще долго после появления настоящих птиц). Теперь, наконец, найден и искомый пернатый "до-археоптериксовый" динозавр.

Anchiornis huxleyi. Видны отпечатки крупных контурных перьев, располагавшихся на всех четырех конечностях и на хвосте. Длина масштабной линейки — 5 см.
4) Как велоцираптор разучился летать. На кости велоцираптора из Монголии в 2007 году были обнаружены специальные бугорки, к которым крепились перья. Стало понятно, что не только мелкие представители теропод имели перья и летали, но и крупные, явно нелетающие динозавры также имели перьевой покров.

На схеме показаны основные ветви эволюции динозавров, приведшие к птицам. Красные прямоугольники отмечают найденные (а не предполагаемые!) признаки перьев у этих рептилий. А цифры показывают 4 события гигантизма. Видно, что рептилии, находящиеся в основании древа, мелкие. Эта схема не учитывает вышеупомянутого четырехкрылого динозавра Anchiornis huxleyi, который был найден лишь в 2009 году, и птицединозавра Epidexipteryx с четырьмя длинными хвостовыми перьями, найденного в 2008 году.
5) В Китае нашли кости гигантского птицеподобного динозавра. В 2007 году китайские палеонтологи сообщили о находке еще одного необычного динозавра, близкого к предполагаемым предкам птиц и получившего название Gigantoraptor erlianensis. В отличие от других птицеподобных динозавров, вес которых редко превышал 40 кг, гигантораптор был настоящим гигантом и весил около полутора тонн.
6) В Китае нашли мохнатого динозавра. В 2009 году был найден небольшой (около 70 см) растительноядный динозавр из группы гетеродонтозавров, живший около 125 млн лет назад. Вместе с костями сохранились пучки длинных нитевидных образований, похожих на волосы, а еще больше — на «протоперья», характерные для некоторых ящеротазовых манирапторных динозавров — предков птиц. Наличие протоперьев у неродственного птицам динозавра показывает, что мохнатые (или оперенные) формы встречались среди динозавров чаще, чем было принято считать.
Другие переходные формы между крупными таксонами
Известно множество других переходных форм, в том числе — от беспозвоночных к рыбам, от рыб к четвероногим, от земноводных к рептилиям, от рептилий к млекопитающим (см. также: List of transitional fossils). Все эти хрестоматийные примеры имело бы смысл разобрать подробно, однако нельзя объять необъятное, поэтому прих
cont.ws
§ 17. Доказательства эволюции
Для обоснования теории эволюции Ч. Дарвин широко использовал многочисленные доказательства из области палеонтологии, биогеографии, морфологии. Впоследствии были получены факты, воссоздающие историю развития органического мира и служащие новыми доказательствами единства происхождения живых организмов и изменяемости видов в природе.
Палеонтологические находки — едва ли не самые убедительные доказательства протекания эволюционного процесса. К ним относятся окаменелости, отпечатки, ископаемые остатки, ископаемые переходные формы, филогенетические ряды, последовательность ископаемых форм. Рассмотрим более подробно некоторые из них.
1. Ископаемые переходные формы — формы организмов, сочетающие признаки более древних и молодых групп.
Среди растений особый интерес представляют псилофиты. Они произошли от водорослей, первыми из растений осуществили переход на сушу и дали начало высшим споровым и семенным растениям.Семенные папоротники — переходная форма между папоротниковидными и голосеменными, а саговниковые — между голосеменными и покрытосеменными.
Среди ископаемых позвоночных можно выделить формы, являющиеся переходными между всеми классами этого подтипа. Например, древнейшая группа кистеперых рыб дала начало первым земноводным —стегоцефалам (рис. 3.15, 3.16). Это было возможно благодаря характерному строению скелета парных плавников кистеперых рыб, имевших анатомические предпосылки для превращения их в пятипалые конечности первичных земноводных. Известны формы, образующие переход между рептилиями и млекопитающими. К ним относятся звероящеры (иностранцевия) (рис. 3.17). А связующим звеном между пресмыкающимися и птицами явилась пер-воптица (археоптерикс) (рис. 3.18).
Наличие переходных форм доказывает существование филогенетических связей между современными и вымершими организмами и помогает в построении естественной системы и родословного древа растительного и животного мира.
2. Палеонтологические ряды — ряды ископаемых форм, связанные друг с другом в процессе эволюции и отражающие ход филогенеза (от греч. phylon — род, племя, genesis — происхождение). Классическим примером применения рядов ископаемых форм для выяснения истории отдельной группы животных является эволюция лошади. Русский ученый В.О. Ковалевский (1842—1883) показал постепенность эволюции лошади, установив, что сменяющие друг друга ископаемые формы приобретали все большее сходство с современными (рис. 3.20).
Современные однопалые животные произошли от мелких пятипалых предков, живших в лесах 60—70 млн лет назад. Изменение климата привело к увеличению площади степей и расселению по ним лошадей. Передвижение на большие расстояния в поиске пищи и при защите от хищников способствовало преобразованию конечностей. Параллельно увеличивались размеры тела, челюстей, усложнялось строение зубов и др.
К настоящему времени известно достаточное количество палеонтологических рядов (хо ботных, хищных, китообразных, носорогов, некоторых групп беспозвоночных), которые доказывают существование эволюционного процесса и возможность происхождения одного вида от другого.
Морфологические доказательства основаны на принципе: глубокое внутреннее сходство организмов может показать родство сравниваемых форм, следовательно, чем больше сходство, тем ближе их родство.
1. Гомология органов. Органы, име ющие сходное строение и общее происхождение, называютсягомологичными. Они занимают одинаковое положение в теле животного, развиваются из сходных зачатков и имеют одинаковый план строения. Типичный пример гомологии — конечности наземных позвоночных животных (рис. 3.21). Так, скелет свободных передних конечнрс-тей у них обязательно имеет плечевую кость, предплечье, состоящее из лучевой и локтевой костей, и кисть (запястье, пясть и фаланги пальцев). Такая же картина гомологии отмечается при сравнении скелета задних конечностей. У лошади грифельные косточки гомологичны пястным косточкам второго и четвертого пальцев других копытных. Очевидно, что у современной лошади эти пальцы исчезли в процессе эволюции.
Доказано, что ядовитые железы змей — гомолог слюнных желез других животных, жало пчелы — гомолог яйцеклада, а сосущий хоботок бабочек — гомолог нижней пары челюстей других насекомых.
Гомологичные органы есть и у растений. Например, усики гороха, колючки кактуса и барбариса — видоизмененные листья.
Установление гомологии органов позволяет найти степень родства между организмами.
2. Аналогия. Аналогичные органы — это органы, имеющие внешнее сход ство и выполняющие одинаковые фун кции, но имеющие разное происхож дение. Эти органы свидетельствуют лишь о сходном направлении приспо соблений организмов, определяемом в
процессе эволюции действием естественного отбора. Наружные жабры головастиков, жабры рыб, многощетинковых кольчатых червей и водных личинок насекомых (например, стрекоз) аналогичны. Бивни моржа (видоизмененные клыки) и бивни слона (разросшиеся резцы) — типичные аналогичные органы, так как их функции сходны. У растений аналогичны колючки барбариса (видоизмененные листья), колючки белой акации (видоизмененные прилистники) и шиповника (развиваются из клеток коры).
Рудименты. Рудиментарными (от лат. rudimentum — зачаток, первоос нова) называются органы, которые закладываются в ходе эмбрионального раз вития, но в дальнейшем перестают развиваться и остаются у взрослых форм в недоразвитом состоянии. Другими словами, рудименты — это органы, утра тившие свои функции. Рудименты — ценнейшие доказательства историчес кого развития органического мира и общности происхождения живых форм. Например, у муравьедов рудиментарны зубы, у человека — ушные мышцы, кожная мускулатура, третье веко, а у змей — конечности (рис. 3.22).
Атавизмы. Появление у отдельных организмов какого-либо вида признаков, которые существовали у отдаленных предков, но были утраче ны в ходе эволюции, называется атавизмом (от лат. atavus — предок). У человека атавизмами являются хвост, волосяной покров на всей повер хности тела, многососковость (рис. 3.23). Среди тысяч однопалых лоша дей встречаются экземпляры с трехпалыми конечностями. Атавизмы ненесут каких-либо функций, важных для вида, но показывают историческую взаимосвязь между вымершими и ныне существующими родственными формами.
Эмбриологические доказатель ства. В первой половине 19 в. русский эмбриолог К.М. Бэр (1792—1876) сформулировал закон зародышевого сходства: чем более ранние стадии индивидуального развития исследуются, тем больше сходства обнаруживается между различными организмами.
Например, на ранних стадиях развития эмбрионы позвоночных не отличаются друг от друга. Только на средних стадиях появляются особенности, характерные для рыб и амфибий, а на более поздних — особенности развития рептилий, птиц и млекопитающих (рис. 3.24). Эта закономерность в развитии зародышей указывает на родство и последовательность расхождения в эволюции этих групп животных.
Глубокая связь между индивидуальным и историческим выражается в биогенетическом законе,установленном во второй половине 19 в. немецкими учеными Э. Геккелем (1834—1919) и Ф. Мюллером (1821—1897). Согласно этому закону каждая особь в своем индивидуальном развитии (онтогенезе) повторяет историю развития своего вида, или онтогенез есть краткое
и быстрое повторение филогенеза. Например, у всех позвоночных животных в онтогенезе закладывается хорда — признак, который был свойствен их отдаленным предкам. У головастиков бесхвостых земноводных развивается хвост, что является повторением признаков их хвостатых предков.
В дальнейшем в биогенетический закон были внесены поправки и дополнения. Особый вклад в выяснение связей онто- и филогенеза внес русский ученый А.Н. Северцов (1866—1936).
Ясно, что за такой короткий срок, как индивидуальное развитие, не могут быть повторены все этапы эволюции. Поэтому повторение стадий исторического развития вида в зародышевом развитии происходит в сжатой форме, с выпадением многих этапов. Вместе с тем зародыши организмов одного вида сходны не со взрослыми формами другого вида, а с их зародышами. Так, жаберные щели у зародыша человека в месячном возрасте сходны с аналогичными у зародыша рыбы, а не взрослой рыбы. Это означает, что в онтогенезе млекопитающие проходят стадии, сходные с зародышами рыб, а не со взрослыми рыбами.
Следует отметить, что еще Ч. Дарвин обратил внимание на явление повторения в онтогенезе черт строения предковых форм.
Все приведенные выше сведения имеют большое значение для доказательства эволюции и для выяснения родственных связей между организмами.
Биогеографические доказательства. Биогеография — это наука о закономерностях современного расселения животных и растений на Земле.
Вы уже знаете из курса физической географии, что современные географические зоны сформировались в ходе исторического развития Земли, в результате действия климатических и геологических факторов. Знаете и о том, что часто сходные природные зоны оказываются заселены различными организмами, а разные зоны — сходными. Найти объяснения этим фактам можно только с позиций эволюции. Например, своеобразие флоры и фауны Австралии объясняется обособлением ее в далеком прошлом, в связи с чем развитие животного и растительного мира происходило в изоляции от других материков. Следовательно, биогеография вносит много доказательств в эволюцию органического мира.
В настоящее время для доказательства эволюционных процессов широко используются методы биохимии и молекулярной биологии, генетики, иммунологии.
Так, изучая состав и последовательность нуклеотидов в нуклеиновых кислотах и аминокислот в белках у разных групп организмов и обнаруживая сходство, можно судить об их родстве.
Биохимия располагает методами исследования, с помощью которых можно выяснить «кровное родство» организмов. При сравнении белков крови учитывается способность организмов в ответ на введение в кровь чужих белков вырабатывать антитела. Эти антитела можно выделить из сыворотки крови и определить, при каком разведении эта сыворотка будет реагировать с сывороткой сравниваемого организма. Такой анализ показал, что ближайшие родственники человека — высшие человекообразные обезьяны, а наиболее дальние из них — лемуры.
Эволюция органического мира на Земле подтверждается множеством фактов из всех областей биологии: палеонтологии (филогенетические ряды, переходные формы), морфологии (гомология, аналогия, рудименты, атавизмы), эмбриологии (закон зародышевого сходства, биогенетический закон), биогеографии и др
studfiles.net
Переходные формы у животных
Разнообразные переходные формы являются красноречивым свидетельством процесса эволюции и в животном мире. Связующие звенья между низшими и высшими типами животных отчасти вымерли и могут быть установлены лишь по ископаемым находкам, отчасти же встречаются еще в настоящее время. Вообще формы организмов, которые отстали от других форм в дальнейшем ходе эволюции, могут быть отлично приспособленными к своим особым условиям существования и поэтому сохранять и впредь свою жизнеспособность. Ведь в противном случае на Земле были бы только цветковые растения и люди (что кажется идилличным только с поверхностной точки зрения).
Поднятый о мая 1952 года ни борт датского экспедиционного корабля «Галатея» в Мексиканском задние Тихого океана с глубины 3590 метров живой экземпляр чрезвычайно примитивного моллюска, который считался вымершим еще с девонского периода, проливает слет на организацию общего прототипа аннелидов и моллюсков.
Среднее место между червями и членистоногими занимает группа перипатидов. Их долгое время считали моллюсками, но теперь относят к червям, хотя они, подобно высшим членистоногим, покрыты защитным хитиновым покровом. Их челюсти и усики возникли из ног.
Переходной группой от раков к паукам служат мечехвосты. Вопрос о переходной форме между беспозвоночными и позвоночными еще спорен. Знаменитый ланцетник, несомненно, является чрезвычайно примитивным предшественником позвоночных животных, имеющих еще сегментное строение наподобие червей. Двоякодышащие рыбы, занимающие место между рыбами и амфибиями, дышат в полноводных реках посредством жабр. В периоды же заиления и пересыхания рек они дышат при помощи плавательного пузыря, снабженного большим количеством кровеносных сосудов.
Соединение носа и ротовой полости в первый раз встречается у двоякодышащих рыб. Признаки, свойственные первичным земноводным и пресмыкающимся, имеющие длину 60—80 см, лаитаназухус и котлассия. И. Л. Ефремов объединил их в особый подкласс батрахозавров. Ближе всего к первобытным млекопитающим стоит относящийся к рептилиям подкласс териодонтов. Эти животные были малого или среднего размера. Они передвигались не ползком и толчками, как пресмыкающиеся, а шагами а. Териодонты имели наружные слуховые органы, и их зубы уже слабо дифференцировались на резцы, клыки и бугорчатые коренные зубы. Носовая и ротовая полость уже была соединена вторичным нёбом.
Во время успешных раскопок в Зольнхофено был найден археоптерикс — ископаемая переходная форма между рептилией и птицей. По-видимому, археоптерикс произошел от живших на деревьях ящеров, которые приобрели способность взбираться вверх прыжками. После переходной стадии прыжка с планирующим полетом выработался настоящий птичий лёт. Однако этот родоначальник птиц величиной с голубя имел еще оперенный хвост пресмыкающегося, когти на передних конечностях и зубастый рот. Таким образом, он ясно выдавал свое происхождение.
Характерна группа и поныне существующих однопроходных животных. Это — примитивные яйцекладущие млекопитающие. Их органы пищеварения, выделения и размножения выходят, как у рептилий и птиц, в общий канал, клоаку. Выделяемый грудными железами секрет, которым питаются детеныши, еще не является настоящим молоком. Переходные формы от высших форм млекопитающих к первобытному человеку будут показаны позднее.
mirznanii.com
Макроэволюция. Основные доказательства эволюции
Согласно синтетической теории эволюции, протекающий в природе эволюционный процесс разделяется на два этапа: микроэволюцию и макроэволюцию.
Макроэволюция включает процессы, приводящие к появлению систематических единиц крупнее вида. Изучая макроэволюцию, современное естествознание накопило ряд научных фактов, доказывающих эволюцию органического мира. Доказательством эволюции может считаться любой научный факт, который доказывает хотя бы одно из следующих положений.
- Единство происхождения жизни (наличие общих признаков у всех живых организмов).
- Родственные связи между современными и вымершими организмами или между организмами в крупной систематической группе (наличие общих признаков у современных и вымерших организмов или у всех организмов в систематической группе).
- Действие движущих сил эволюции (факты, подтверждающие действие естественного отбора).
Доказательства эволюции, добытые и накопленные в рамках определенной науки, составляют одну группу доказательств и называются по названию данной науки. Рассмотрим палеонтологические, эмбриологические, сравнительно-анатомические и молекулярно-генетические примеры доказательств эволюции.
Палеонтологические доказательства эволюции
Палеонтология — наука об ископаемых остатках вымерших организмов. Основателем эволюционной палеонтологии считается русский ученый В. О. Ковалевский. К доказательствам эволюции можно отнести ископаемые переходные формы и филогенетические ряды современных видов.
Ископаемые переходные формы — это вымершие организмы, сочетающие в себе признаки более древних и эволюционно более молодых групп. Они позволяют выявить родственные связи, доказывающие историческое развитие жизни. Такие формы установлены как среди животных, так и среди растений. Переходной формой от кистеперых рыб к древним земноводным — стегоцефалам — является ихтиостега. Эволюционную связь между пресмыкающимися и птицами позволяет установить первоптица (археоптерикс). Связующим звеном между пресмыкающимися и млекопитающими является звероящер из группы терапсид. Среди растений переходной формой от водорослей к высшим споровым являются псилофиты (первые наземные растения). Происхождение голосеменных от папоротниковидных доказывают семенные папоротники, а покрытосеменных от голосеменных — саговниковые.
Филогенетические (от греч. phylon — род, племя, genesis — происхождение) ряды — последовательности ископаемых форм, отражающие историческое развитие современных видов (филогенез). В настоящее время такие ряды известны не только для позвоночных, но и для некоторых групп беспозвоночных животных. Русский палеонтолог В. О. Ковалевский восстановил филогенетический ряд современной лошади.
Эмбриологические доказательства эволюции
Эмбриология — наука, изучающая зародышевое развитие организмов. В рамках данной науки были сформулированы закон зародышевого сходства (К. Бэр) и биогенетический закон (Э. Геккель, Ф. Мюллер), доказывающие эволюцию.
Сравнивая зародышевые стадии представителей разных классов позвоночных животных, натуралист К. Бэр в начале XIX в. сформулировал закон зародышевого сходства: чем более ранние стадии индивидуального развития исследуются, тем больше сходства обнаруживается между разными организмами. Этот закон доказывает родственные связи между организмами в крупной систематической группе.
Во второй половине XIX в. ученые Э. Геккель и Ф. Мюллер установили наличие связи между индивидуальным развитием особей (онтогенезом) и историческим развитием видов (филогенезом). Они сформулировали биогенетический закон: онтогенез есть краткое повторение филогенеза.
Позже биогенетический закон был дополнен А. Н. Северцовым и И. И. Шмальгаузеном. Они показали, что в онтогенезе повторяются не взрослые формы предков, а их зародышевые стадии, причем некоторые из них могут выпадать. Например, у человека зародыш имеет жаберные щели, сходные с таковыми у зародыша рыбы, а не у взрослой особи.
Биогенетический закон доказывает наличие родственных связей между современными организмами и их предками.
Сравнительно-анатомические доказательства эволюции
Сравнительная анатомия изучает строение организмов разных систематических групп в сравнительном плане. К доказательствам эволюции, установленным данной наукой, относятся: гомологичные и аналогичные органы, рудименты, атавизмы.
Гомологичные органы формируются из одинаковых эмбриональных зачатков (на одной генетической основе) и занимают на теле организмов одинаковое положение. Например, передние конечности у разных позвоночных животных могут существенно отличаться в зависимости от выполняемой функции, но все они имеют сходное строение.
Пары гомологичных органов у животных составляют: плавательный пузырь рыб и легкие наземных позвоночных; ядовитые железы змей и слюнные железы других организмов; зубы млекопитающих и чешуя акул; жало пчелы и яйцеклад других насекомых.
Гомологичными органами у растений являются: колючки кактуса и барбариса, усики гороха, ловчие кувшины насекомоядных растений, почечные чешуи, пленчатые редуцированные листья хвоща. Все эти органы являются листьями по происхождению, но выполняют разные функции.
Гомологичные органы позволяют установить родственные связи между организмами и доказывают действие разнонаправленного естественного отбора.
Аналогичные органы формируются из разных эмбриональных зачатков (на разной генетической основе) и занимают на теле и внутри тела организмов неодинаковое положение. Например, разные по происхождению колючки у растений. У животных к аналогичным органам относятся: жабры головастиков, рыб, морских кольчатых червей, личинок стрекоз; бивни моржа и слона; крылья птицы и бабочки; конечности крота и медведки.
Аналогичные органы не позволяют установить родственные связи между организмами, но доказывают действие однонаправленного естественного отбора.
Рудименты (от лат. rudimentum — зачаток) — недоразвитые органы современных организмов, которые были хорошо развиты у их предков. Они постепенно утратили свое значение и сейчас находятся на стадии исчезновения. Рудименты сохраняются в течение всей жизни у всех особей данного вида. Примерами рудиментов являются: недоразвитые глаза у пещерных видов животных и кротов, зачатки крыльев у птицы киви, редуцированные зубы у муравьедов, зачатки задних конечностей у китов и дельфинов, задняя пара крыльев у мух (жужжальца), зачатки тазовых костей у змей. У человека к рудиментам относятся: аппендикс, мышцы ушной раковины, третье веко, копчик.
Рудименты подтверждают наличие родственных связей между современными и вымершими организмами. Они также доказывают действие естественного отбора, удаляющего ненужный признак.
Атавизмы (от лат. atavus — предок) — признаки отдаленных предков, появляющиеся у некоторых современных организмов как отклонение от нормы. Они были утрачены в процессе эволюции. Возникновение атавизмов доказывает, что в генотипах современных организмов сохранились гены предков, отвечающие за эти признаки. Но действие этих генов заблокировано. В случаях, когда блокирование снимается, проявляется признак предков. В отличие от рудиментов атавизмы присутствуют только у отдельных особей. К атавизмам относятся: выраженный волосяной покров на всем теле, развитый хвост и дополнительные пары молочных желез у человека, трехпалые конечности у лошади. Атавизмы доказывают родственные связи между современными и вымершими организмами.
Молекулярно-генетические доказательства эволюции
Молекулярная биология — наука, изучающая процессы жизнедеятельности организмов на молекулярном уровне. Генетика изучает закономерности наследственности и изменчивости организмов. В рамках этих наук было доказано, что у всех организмов наследственная информация хранится в ДНК, состоящей из четырех типов нуклеотидов. Эта информация зашифрована с помощью универсального триплетного кода. Как вы уже знаете, ДНК входит в состав хромосом, количество которых является видовой характеристикой. Расшифровка наследственной информации у всех организмов происходит в процессе транскрипции и трансляции с участием иРНК и тРНК. Все эти факты доказывают единство происхождения жизни и, следовательно, являются доказательствами эволюции.
Доказательства эволюции накоплены в области разных наук. Палеонтология установила наличие ископаемых переходных форм и филогенетические ряды современных видов. В эмбриологии были открыты закон зародышевого сходства и биогенетический закон. К сравнительно-анатомическим доказательствам эволюции относятся: гомологичные и аналогичные органы, рудименты и атавизмы. В рамках молекулярной биологии и генетики получены доказательства единства происхождения жизни.
jbio.ru
Переходная форма - это... Что такое Переходная форма?

 Лондонский экземпляр Археоптерикса, переходного звена между рептилиями и птицами.
Лондонский экземпляр Археоптерикса, переходного звена между рептилиями и птицами. Переходная форма — организм с промежуточным состоянием, обязательно существующим при состоявшемся постепенном переходе от одного биологического типа строения к другому. Переходные формы характеризуются наличием более древних и примитивных (в значении первичных) черт, чем их более поздние родственники, но, в то же время, наличием более прогрессивных (в значении более поздних) черт, чем их предки. Как правило, говоря о промежуточных формах, имеют в виду ископаемые виды, хотя промежуточные виды вовсе не должны непременно вымирать. Известно множество переходных форм, иллюстрирующих происхождение четвероногих от рыб, рептилий от земноводных, птиц от динозавров, млекопитающих от териодонтов, китообразных от наземных млекопитающих, лошадей от пятипалого предка и человека от древних гоминид.
Теория эволюции
Ко времени написания Чарльзом Дарвином «Происхождения видов» науке не были известны переходные формы жизни. Этот факт Дарвин считал одним из самых слабых мест своей теории. Однако уже через два года был обнаружен археоптерикс — переходное звено между рептилиями и птицами.
Переходные формы между двумя группами организмов не обязательно являются потомками одной группы и предками другой. По ископаемым останкам, как правило, невозможно достоверно установить, является ли один организм прямым предком другого. Кроме того, вероятность найти в палеонтологической летописи чьего-нибудь прямого предка очень мала, и по умолчанию среди биологов принято такую возможность вообще не рассматривать. Гораздо выше вероятность обнаружить относительно близких родственников этого предка, схожих с ним по строению. Поэтому любая переходная форма автоматически интерпретируется как боковая ветвь эволюции, а не «участок ствола».[1]
Примеры
 Меритерий, ранний предок слонов, больше похож на свинью, хотя уже имеет маленький хобот и бивни, свойственные хоботным.
Меритерий, ранний предок слонов, больше похож на свинью, хотя уже имеет маленький хобот и бивни, свойственные хоботным. В качестве примеров переходных форм можно привести Амбулоцетуса — «ходячего кита» (переходная форма между китообразными и наземными млекопитающими), Тиктаалика и Ихтиостегу (переходная форма от рыб к земноводным), Мезогиппуса (между ранними лошадиными и современной лошадью).
Переходные формы в эволюции Человека Разумного
 Черепа гоминид, от шимпанзе (A) до человека разумного (N). A — Шимпанзе, B,C — Австралопитек африканский, D,E — Человек умелый , F — Человек рудольфский, G — Человек прямоходящий, H — Человек работающий , I — Гейдельбергский человек, J,K,L — Неандерталец, M — Кроманьонец, N — Человек разумный
Черепа гоминид, от шимпанзе (A) до человека разумного (N). A — Шимпанзе, B,C — Австралопитек африканский, D,E — Человек умелый , F — Человек рудольфский, G — Человек прямоходящий, H — Человек работающий , I — Гейдельбергский человек, J,K,L — Неандерталец, M — Кроманьонец, N — Человек разумный В настоящее время известно множество переходных форм между человеком разумным и его обезьяноподобными предками. Так, например, открыты останки Сахелантропа, Ардипитека, Австралопитеков (африканского, афарского и других), Человека умелого, Человека работающего, Человека прямоходящего, Человека-предшественника, Гейдельбергского человека, Неандертальца и раннего Человека разумного.
Спор о постепенности эволюционных преобразований
Переходные формы являются объектом исследований в споре между градуалистами и пунктуалистами. Градуалисты считают, что эволюция является постепенным, непрерывным процессом. Пунктуалисты, или сторонники теории прерывистого равновесия считают, что виды в течение долгого времени остаются неизменными, а изменения происходят в очень короткий промежуток времени. Переходные формы являются аргументом обеих сторон. Пунктуалисты утверждают, что большинство видов появляются внезапно и на протяжении всей своей истории меняются слабо. Градуалисты отмечают, что среди позвоночных нет ни одного бесспорного примера очень быстрого перехода от одного вида к другому, большинство переходов вообще плохо задокументированы ископаемыми. Также много хорошо описанных внутривидовых изменений. Например, описаны изменения среднекайнозойских кроликов, миоценовых грызунов, гиппарионов, эоценовых парнокопытных, переход между родами эоценовых приматов.[2]
Недостающие звенья
| 1850 |  |
| 1900 |  |
| 1950 | 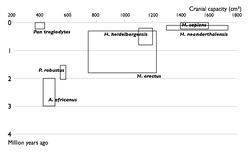 |
| 2002 |  |
| Прогресс в изучении эволюции человека. Увеличение числа известных науке видов гоминин по времени. Каждый вид изображен в виде прямоугольника, показывающего границы, в которых варьировал объём черепа, и место вида в палеонтологической летописи. Видно постепенное заполнение промежутков между видами. | |
Многие переходные формы между живыми организмами пока не найдены. Обычно их называют недостающими звеньями. Редкость переходных форм объясняется неполнотой палеонтологической летописи. Сторонники теории прерывистого равновесия также указывают на быстроту эволюционных преобразований. Однако палеонтологи постоянно находят новые ископаемые, заполняя пробелы (например, в 2004 году был найден Тиктаалик — ещё одно переходное звено между рыбами и земноводными)
Неполнота палеонтологической летописи
Далеко не каждая переходная форма существует в виде окаменелости из-за принципиальной неполноты палеонтологической летописи. Неполнота вызвана особенностями процесса фоссилизации, то есть перехода в окаменелое состояние. Для образования окаменелости необходимо, чтобы погибший организм был погребён под большим слоем осадочных пород. Из-за крайне медленной скорости осадконакопления на суше, чисто сухопутные виды редко переходят в окаменелое состояние и сохраняются. Кроме того, редко удаётся обнаружить виды, живущие в глубинах океана, из-за редкости поднятия на поверхность больших массивов дна. Таким образом, большинство известных ископаемых (а, соответственно, и переходных форм) — это либо виды, обитающие на мелководье, в морях и реках, либо наземные виды, ведущие полуводный образ жизни, либо обитающие недалеко от береговой линии. Закономерности процессов захоронения останков живых существ изучает особая отрасль палеонтологии — тафономия.
Филогенетические ряды
 Филогенетический ряд лошади. Показана реконструкция ископаемых видов, полученных из последовательных слоёв породы. Справа — вид спереди передней левой ноги с выделенным третьим сегментом, а также зуб в продольном сечении[3].
Филогенетический ряд лошади. Показана реконструкция ископаемых видов, полученных из последовательных слоёв породы. Справа — вид спереди передней левой ноги с выделенным третьим сегментом, а также зуб в продольном сечении[3]. Филогенетические ряды − ряды видов, последовательно сменявших друг друга в процессе эволюции различных групп животных и растений.
Впервые были открыты В. О. Ковалевским, который показал, что современные однопалые копытные происходят от древних пятипалых мелких всеядных животных.
Креационизм
Сторонники креационизма заявляют, что переходных форм не обнаружено. Научное сообщество считает подобные заявления ложью и намеренным введением в заблуждение.
См. также
Примечания
Литература
- Дарвин, Чарльз. Происхождение видов путём естественного отбора: В 2 кн. — М.: ТЕРРА - Книжный клуб, 2009. — ISBN 978-5-275-02114-1
- Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х т. — Мир, 1992. — 280 с. — ISBN 5-03-001819-0
Ссылки
dic.academic.ru









