Справочник химика 21. Паренхима ткань растений
Структурные ткани
Структурные ткани
Клетки многоклеточных организмов образуют ткани – системы сходных по строению и функциям клеток и связанных с ними межклеточных веществ. Ткани интегрируются в более крупные функциональные единицы, называемые органами. Внутренние органы характерны для животных; здесь они входят в состав систем органов (дыхательной, нервной и пр.). Подобная специализация, с одной стороны, улучшает работу организма в целом, а с другой требует повышения степени координации и интеграции различных тканей и органов.
Наука, изучающая ткани, называется гистологией. Обычно она оперирует с тонкими срезами тканей, иногда окрашенных, изучая их при помощи светового микроскопа.
Растительные ткани можно условно разделить на структурные ткани, состоящие из клеток одного вида, и проводящие ткани, состоящие из клеток разных видов. К первым относятся паренхима, колленхима и склеренхима, ко вторым – ксилема и флоэма.
 1 |
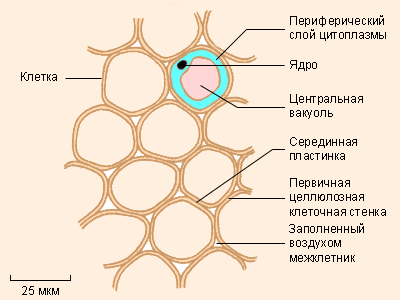
Паренхимные клетки |
 2 |
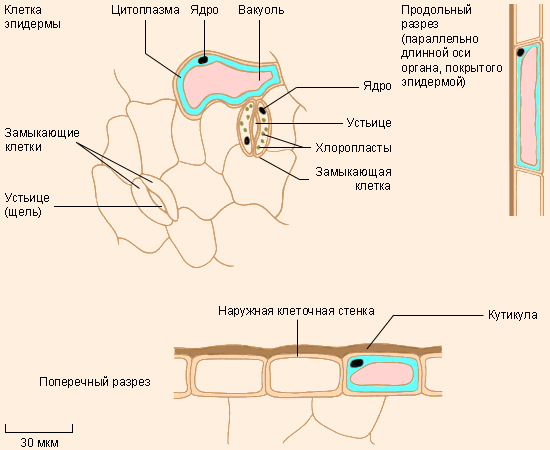
Эпидерма |
Клетки паренхимы заполняют пространство между более специализированными тканями. Они составляют основную массу стеблей и корней. Осмотическое давление дает возможность паренхиме служить опорой тем органам, в которых она находится; в засушливые периоды эти клетки теряют воду, и растение увядает. По стенкам клеток паренхимы перемещаются вода и соли; в некоторых органах эти клетки запасают питательные вещества.
В некоторых частях растения паренхима видоизменяется, становясь более специализированной. Так, эпидерма – это тонкая покровная ткань, состоящая из одного слоя клеток, покрывающая целиком всё первичное тело растения. Её основная функция – защита растения от высыхания и проникновения микроорганизмов; этому помогает кутикула – плёнка из воскообразного вещества кутина, покрывающая внешнюю поверхность эпидермы.
Как правило, эпидерму составляют уплощённые тонкие клетки, плотно прилегающие друг к другу, но среди них на определённом расстоянии друг от друга встречаются особые – замыкающие – клетки. Они всегда располагаются парами, и между ними имеется небольшое отверстие, называемое устьицем, которое играет важную роль в газообмене. Иногда клетки эпидермы образуют волоски и жгутики, которые создают дополнительную поверхность для всасывания питательных веществ, служат опорой, отпугивают животных (например, у крапивы) или наоборот при помощи специальных клейких веществ ловят насекомых. Мезофилл – ещё один тип паренхиматозной ткани, располагающийся между двумя слоями эпидермиса листа и осуществляющий фотосинтез. Эндодерма окружает проводящую ткань растений. Её можно рассматривать как самый внутренний слой коры. Наконец, клетки перицикла, находящиеся между эндодермой и проводящей тканью, участвуют в процессе роста растений.
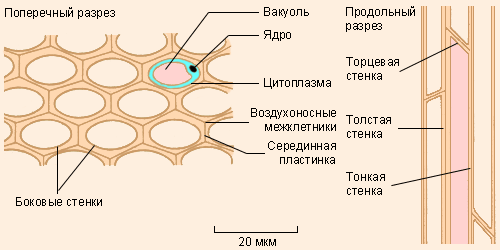
Колленхиму составляют вытянутые в направлении длинной оси органа клетки, в которых содержится большое количество целлюлозы. Эта ткань играет важную роль, обеспечивая органам дополнительную опору; при этом клетки колленхимы, оставаясь живыми, способны растягиваться, не мешая расти другим клеткам.
 3 |
Колленхима |
 4 |
Склеренхима |
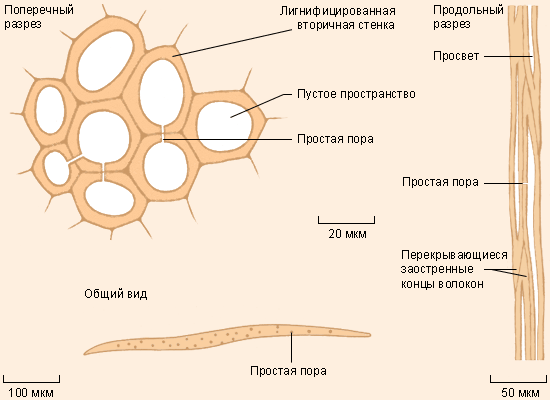
Склеренхима находится, в основном, в коре, сердцевине и плодах. Её мёртвые клетки окружены лигнином – веществом с повышенной прочностью на растяжение и изгиб. Переплетающиеся волокна, которые образует склеренхима, ещё более усиливают опору. Склеренхима – важная опорная ткань деревьев и кустарников; она образуется уже после того, как заканчивается вытягивание живых клеток, которые она окружает.
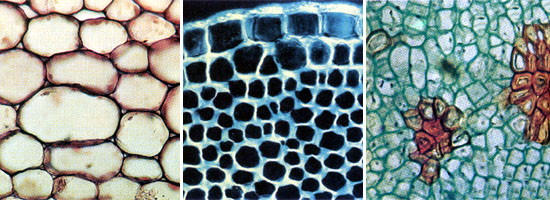 5 |
Слева направо: паренхима в листьях травы, колленхима в молодых побегах бузины, склеренхима (коричневые группы клеток) в мякоти груши |
www.ebio.ru
Паренхимная ткань паренхима растений - Справочник химика 21
В некоторых частях растения паренхимные клетки, видоизменяясь, становятся более специализированными. Мы перечислим здесь некоторые из тканей, которые могут рассматриваться как модифицированная паренхима. [c.222]Фотосинтетическая деятельность клеток мезофилла обогащает ткани листа сахарами и другими продуктами фотосинтеза. В результате возрастает функциональная активность проводящих пучков. Теоретически существуют два способа транспорта ассимилятов к проводящим пучкам по симпласту (через плазмодесмы и цитоплазму последовательного ряда клеток) и по апопласту (по клеточным стенкам). Однако у многих видов растений между клетками мезофилла и флоэмы плазмодесмы развиты крайне слабо или совсем отсутствуют. В апопласте листовой пластинки может находиться около 1/5 сахаров, содержащихся в листе, и значительная доля свободных аминокислот. Клетки листовой паренхимы сравнительно легко выделяют ассимиляты в наружную среду и относительно слабо их поглощают. Клетки флоэмных окончаний, напротив, способны усиленно поглощать из внешних растворов сахара и аминокислоты против концентрационного градиента с помощью энергозависимых переносчиков. Есть основание считать, что, выходя из паренхимных клеток листа в клеточные стенки, сахароза расщепляется находящейся там инвертаз ой на гексозы (фруктозу и глюкозу), которые в проводящих ч ах в овь образуют сахарозу. [c.104]
По-видимому, всякая живая растительная ткань, состоящая из содержащих ядра клеток, при ее изоляции и помещении в подходящую культуральную среду может образовывать делящийся, недифференцированный каллус. Однако получить каллус первый раз из нового растительного материала часто довольно трудно, так как потребности в питательных веществах у тканей, взятых от различных видов растений или даже из разных частей одного растения, очень различны. В общем оказалось легче культивировать исходно незеленые, паренхимные ткани, такие, как паренхима флоэмы или сердцевины. Получать зеленые, фотосинтезирующие каллусы из содержащих хлоропласты клеток листьев научились гораздо позже. [c.236]МЕЗОФИЛЛ (СМ. ТАКЖЕ РИС. 7.3. И 7.4). Эта выполняющая ткань располагается между двумя слоями эпидермиса листа (рис. 6.1) и состоит из модифицированных паренхимных клеток, осуществляющих фотосинтез. Фотосинтетическую паренхиму иногда называют хлоренхимой. Цитоплазма клеток хлоренхимы содержит большое число хлоропластов, в которых и протекают реакции фотосинтеза. У двудольных растений мезофилл состоит из двух четко различающихся слоев верхний слой составляет палисадная паренхима, клетки которой имеют столбчатую форму, а нижний — губчатая паренхима с клетками неправильной формы, содержащими меньше хлоропластов. Фотосинтез вдет главным образом в палисадной паренхиме, а воздухоносные межклетники губчатой паренхимы обеспечивают интенсивный газообмен. [c.224]
Как мы ужо знаем из вводной главы предыдущего тома Кизни растений , паренхима в древесине существует, как правило, в виде двух модификаций. Группы живых клеток, собранных в горизоиталт.пые (радиальные) полосы, называются д р е is с с и н и ы Л1 и или к с н-.II е м н ы м и лучами. Паренхимные клетки, собранные в вертикальные тяжи, тянущиеся вдоль стебля, образуют древесин и у ю, или т я ж е в у ю, паренхиму. Вся система живых запасающих клеток образует единую, интегрированную систему, отдельные звенья которой обычно 6o,itee или меиее соприкасаются на том или ином уровне. На т[)ехмсрной блок-дн-аграмме (рис. 1) хороню видно расположение обоих типов запасающих тканей. [c.14]
chem21.info
Паренхимная ткань паренхима - Справочник химика 21
Микроскопия. На поперечном срезе видно характерное для корня преобладание тонкостенной паренхимной ткани. В коре находятся многочисленные тангентально вытянутые группы лубяных волокон, расположенные прерывистыми концентрическими поясами. Более мелкие группы волокон разбросаны в древесине. Волокна толщиной 10—35 мкм со слабоутолщенными, неодревесневшими или слабоодревесневшими стенками и большим просветом. Сосуды и трахеиды расположены небольшими группами. Сердцевинные лучи одно-, реже двухрядные. В паренхиме видны многочисленные крупные клетки со слизью, находящиеся как в коре, так и в древесине. В воде слизь растворяется, клетки становятся бесцветными и кажутся пустыми. Клетки паренхимы заполнены крахмальными зернами, местами встречаются мелкие друзы оксалата кальция. [c.344] Вторым анатомическим элементом являются паренхимные клетки, образующие живую ткань - паренхиму, главным образом лучевую. Ее клетки образуют сердцевинные лучи - ряды из паренхимных клеток, идущие горизонтально по радиусам ствола. Лучи могут состоять только из паренхимных клеток (гомогенные лучи), либо содержать кроме паренхимных клеток горизонтальные лучевые трахеиды (гетерогенные лучи). У хвойных пород сердцевинные лучи узкие, однорядные (за исключением лучей, в которых образуются горизонтальные смоляные ходы - см. ниже), а их высота по числу клеток колеблется. Горизонтальные (лучевые) трахеиды по размерам близки к паренхимным клеткам, но малочисленны по сравнению с последними. В отличие от паренхимных клеток лучевые трахеиды имеют окаймленные поры (см. ниже). Сердцевинные лучи проводят растворы питательных веществ в горизонтальном направлении. В древесине некоторых хвойных пород в небольших количествах (например, у сосны 0,5%) содержится вертикальная (осевая, или тяжевая) паренхима, ряды клеток которой проходят вдоль ствола. [c.200]Древесина всех хвойных пород состоит на 90—95% (по объему) из трахеид — длинных клеток с крупными окаймленными порами. В древесине хвойных совершенно отсутствуют сосуды. Живая паренхимная ткань представлена здесь паренхимой сердцевинных лучей. Древесная паренхима имеется в небольшом количестве и у немногих хвойных. В древесине некоторых хвойных пород имеются смоляные ходы. В древесине сосны, ели и лиственницы вертикальные смоляные ходы расположены вдоль ствола среди трахеид, а горизонтальные, поперечные,— среди паренхимы более широких сердцевинных лучей. Смоляные ходы имеются также в коре молодых стеблей и ветвей и в хвое. Хвоя может служить местом отложения запасных веществ на зиму, что в некоторой степени компенсирует недостаток древесной паренхимы. [c.39]
В некоторых частях растения паренхимные клетки, видоизменяясь, становятся более специализированными. Мы перечислим здесь некоторые из тканей, которые могут рассматриваться как модифицированная паренхима. [c.222]
Микроскопия. На поперечном срезе видно характерное для корня преобладание тонкостенной паренхимной ткани. В коре находятся многочисленные тангентально вытянутые группы лубяных волокон, расположенные прерывистыми концентрическими поясами. Более мелкие группы волокон разбросаны в древесине. Волокна толщиной 10—35 жо1 со слабо утолщенными неодревеспевшими или слабо одревесневшими стенками и большим просветом. Сосуды и трахеиды расположены небольшими группами. Сердцевинные лучи одно-, реже двухрядные. В паренхиме видны многочисленные крупные клетки со слизью, находящиеся как в коре, так и в древесине. В воде слизь растворяется, клетки становятся бесцветными и кажутся пустыми. Раствор метиленового синего окрашивает клетки со слизью в голубой цвет. Для проведения двойного окрашивания срез помещают в раствор хлорида окисного железа на 20 минут, раствор удаляют фильтровальной бумагой, прибавляют спиртовой раствор метиленового синего и промывают водой. Слизистые клетки окрашиваются в желтый, волокна — в синий, сосуды — в зеленый цвет клетки паренхимы остаются бесцветными. [c.579]
Фотосинтетическая деятельность клеток мезофилла обогащает ткани листа сахарами и другими продуктами фотосинтеза. В результате возрастает функциональная активность проводящих пучков. Теоретически существуют два способа транспорта ассимилятов к проводящим пучкам по симпласту (через плазмодесмы и цитоплазму последовательного ряда клеток) и по апопласту (по клеточным стенкам). Однако у многих видов растений между клетками мезофилла и флоэмы плазмодесмы развиты крайне слабо или совсем отсутствуют. В апопласте листовой пластинки может находиться около 1/5 сахаров, содержащихся в листе, и значительная доля свободных аминокислот. Клетки листовой паренхимы сравнительно легко выделяют ассимиляты в наружную среду и относительно слабо их поглощают. Клетки флоэмных окончаний, напротив, способны усиленно поглощать из внешних растворов сахара и аминокислоты против концентрационного градиента с помощью энергозависимых переносчиков. Есть основание считать, что, выходя из паренхимных клеток листа в клеточные стенки, сахароза расщепляется находящейся там инвертаз ой на гексозы (фруктозу и глюкозу), которые в проводящих ч ах в овь образуют сахарозу. [c.104]
Исследователи [46] пытались также выяснить фазовое состояние ксилоуронидов в клеточных стенках паренхимных тканей древесины березы. Для этой цели из измельченной древесины березы (Betula verru osa) удаляли лигнин хлоритом натрия в уксуснокислой среде в присутствии водного раствора этанола. Полученную мацерированную холоцеллюлозу промывали 50%-ным этанолом на металлической сетке с размерами отверстий 40 мк. В этих условиях через отверстия сетки отмывались только клетки древесной паренхимы, содержащие зерна крахмала. Крахмал удаляли обработкой клеток 0,1 %-ным раствором а-амидазы в 0,01М фосфатном буфере при 40° С в течение 8 ч. Количественный гидролиз полученных паренхимных клеток дал следующие результаты (%) галактозы 3,6, глюкозы 35,7, маннозы 2,0, арабинозы 0,9, ксилозы 57,8. Таким образом, в клеточных стенках паренхимных тканей древесины березы после удаления лигнина обнаружено более 70% [c.323]
Внутренние эфирномасличные вместилища — железы внутренней секреции и выделительные ходы — образуются преимущественно в результате расслоения клеток внутренних паренхимных тканей (схизогенный способ) или же путем растворения клеток паренхимы (лизигенный способ). [c.16]
По-видимому, всякая живая растительная ткань, состоящая из содержащих ядра клеток, при ее изоляции и помещении в подходящую культуральную среду может образовывать делящийся, недифференцированный каллус. Однако получить каллус первый раз из нового растительного материала часто довольно трудно, так как потребности в питательных веществах у тканей, взятых от различных видов растений или даже из разных частей одного растения, очень различны. В общем оказалось легче культивировать исходно незеленые, паренхимные ткани, такие, как паренхима флоэмы или сердцевины. Получать зеленые, фотосинтезирующие каллусы из содержащих хлоропласты клеток листьев научились гораздо позже. [c.236]
Второй вид тканей луба - лубяная (флоэмная) паренхима, выполняющая проводящие и запасающие функции и составляющая основную массу тканей луба. Паренхимные клетки с тонкими нелигнифицированными стенками образуют лубяные (флоэмные) лучи, являющиеся продолжением сердцевинных лучей ксилемы, и вертикальную лубяную паренхиму. В лубяных лучах некоторых пород (например, пихты) имеются горизонтальные смоляные ходы. [c.205]
Микроскопия. На поперечном срезе корня видна покровная ткань, состоящая из 1—2 слоев округлых клеток эпидермиса с тонкими опробковевшими оболочками. Первичная кора состоит из крупных, тангентально вытянутых клеток с неравномерно утолщенными оболочками. Эндодерма хорошо выражена, клеточные оболочки ее окрашиваются от судана 1И в оранжево-красный цвет. Вторичная кора значительно уже первичной и состоит из мелких клеток — проводящих элементов луба и более крупных клеток лубяной паренхимы. Камбиальная зона слабо выражена. В древесине корня сосуды разного диаметра, располагаются без особого порядка, сердцевинные лучи незаметны. В паренхимных клетках коры и древесины содержатся капли жирного масла изредка встречаются мелкие крахмальные зерна. [c.363]
На рис. 11 цифрами 5 и 6 обозначены паренхимные клетки. Описаин1,1е до этого времени элементы 2, 3, 4, 7, 8. 9 являю гся прозенхимными, т. е. они после образования из камбия быстро теряют свой протопласт и в различной степени выполняют проводящие и механические функции. Продольная паренхима 5 и веретенообразная паренхима 6, как и другие элементы, изображенные на рис. И, завершают свой онтогенез в год своего образования, но по какой-то причине сохраняют свои протопласты, пока являются частью заболони. В противопололсность прозенхиме древесины древесная паренхима некоторое время продолжает жить и функционирует, главным образом, как ткань, выполняющая запасные функции. [c.59]
МЕЗОФИЛЛ (СМ. ТАКЖЕ РИС. 7.3. И 7.4). Эта выполняющая ткань располагается между двумя слоями эпидермиса листа (рис. 6.1) и состоит из модифицированных паренхимных клеток, осуществляющих фотосинтез. Фотосинтетическую паренхиму иногда называют хлоренхимой. Цитоплазма клеток хлоренхимы содержит большое число хлоропластов, в которых и протекают реакции фотосинтеза. У двудольных растений мезофилл состоит из двух четко различающихся слоев верхний слой составляет палисадная паренхима, клетки которой имеют столбчатую форму, а нижний — губчатая паренхима с клетками неправильной формы, содержащими меньше хлоропластов. Фотосинтез вдет главным образом в палисадной паренхиме, а воздухоносные межклетники губчатой паренхимы обеспечивают интенсивный газообмен. [c.224]
Микроскопия. Строение корневища и листовых черешков на поперечных срезах, в основном, одинаково. За эпидермисом следует гиподерма, состоящая из многих рядов клеток с толстыми стенками бурого цвета. Основная ткань состоит из рыхло расположенных клеток тонкостенной паренхимы, образующих большие межклетные пространства. В межклетниках встречаются одноклеточные шаровидные или овальные железки на тонкой ножке с желтовато-зеленым содержимым. Паренхимные клетки заполнены мелкими зернами крахмала. Проводящие пучки (столбы) отделены от основной ткани эндодермой, состоящей из одного ряда односторонне утолщенных клеток бурого цвета. Пучки концентрические, центроксилемные. Ксилема состоит из крупных трахеид, между которыми расположены паренхимные клетки. [c.595]
Как мы ужо знаем из вводной главы предыдущего тома Кизни растений , паренхима в древесине существует, как правило, в виде двух модификаций. Группы живых клеток, собранных в горизоиталт.пые (радиальные) полосы, называются д р е is с с и н и ы Л1 и или к с н-.II е м н ы м и лучами. Паренхимные клетки, собранные в вертикальные тяжи, тянущиеся вдоль стебля, образуют древесин и у ю, или т я ж е в у ю, паренхиму. Вся система живых запасающих клеток образует единую, интегрированную систему, отдельные звенья которой обычно 6o,itee или меиее соприкасаются на том или ином уровне. На т[)ехмсрной блок-дн-аграмме (рис. 1) хороню видно расположение обоих типов запасающих тканей. [c.14]
chem21.info
Паренхима - ТКАНИ РАСТЕНИЙ - ВЫСШИЕ РАСТЕНИЯ - РАСТЕНИЯ
РАСТЕНИЯ
ВЫСШИЕ РАСТЕНИЯ
ТКАНИ РАСТЕНИЙ
Паренхима
Паренхима представляет собой целую группу более или менее специализированных тканей, которые заполняют пространства внутри тела растения между проводящими и механическими тканями. Обычно клетки паренхимы имеют округлую, реже вытянутую форму (рис. 160). Характерно наличие развитых межклетников. Пространства между клетками совместно образуют важную транспортную систему - апопласт. Кроме того, межклетники образуют своеобразную «систему вентиляции» растения. Через устьица или чечевички они связаны с атмосферным воздухом и обеспечивают оптимальный газовый состав внутри растения. Особенно актуальными развитые межклетники становятся у растений, которые произрастают на заболоченной почве, где нормальный газообмен затруднен. Такую паренхиму часто называют аэренхимой.
Поскольку элементы паренхимы заполняют промежутки между другими тканями, они выполняют также функцию опоры. Учитывая, что все клетки паренхимы являются живыми, у них нет таких толстых оболочек, как, например, у склеренхимы. Поэтому механические свойства обеспечиваются тургором. В засушливое время содержание воды падает, что приводит к плазмолизу и завяданию растения.


Ассимиляционная паренхима образована тонкостенными клетками с обильными межклетниками. Большую часть цитоплазмы занимают многочисленные хлоропласты, поэтому этот тип паренхимы называют также хлоренхимой. Хлоропласты в ней располагаются не беспорядочно, а выстраиваются вдоль стенки (которая для увеличения поверхности может образовывать инвагинации), не затеняя друг друга. Именно в ассимиляционной паренхиме происходят реакции фотосинтеза, которые обеспечивают растение органическими веществами и энергией, а также делают возможным существование всех живых организмов на планете Земля.
Ассимиляционные ткани представлены только в освещенных частях растения, причем от окружающей среды они отделены прозрачной эпидермой. Как только на смену эпидерме приходят непрозрачные вторичные покровные ткани, ассимиляционная паренхима исчезает.
Запасающая паренхима служит вместилищем органических веществ, которые временно не используются растительным организмом. В принципе любая клетка с живым протопластом способна откладывать органические вещества в виде различного рода включений, однако некоторые клетки на этом специализируются. Богатые энергией соединения откладываются только в вегетационный период, а расходуются в период покоя и при подготовке к очередной вегетации. Поэтому становится понятным, что запасные вещества в вегетативных органах откладываются только у многолетних растений, для однолетних это неактуально.
Вместилищем запасов могут быть как обычные органы (побег, корень), так и специализированные - корневища, клубни, луковицы и др. Кроме того, все без исключения семенные растения запасают энергетически ценные вещества в семенах (в семядолях или эндосперме). Многие растения, произрастающие в засушливых условиях, запасают не только органические вещества, но и воду. Например, алоэ запасает воду в своих мясистых листьях, кактусы - в побегах.
www.compendium.su