Отдел Цветковые растения (Magnoliophyta или Angiospermae). Отдел цветковые растения
Отдел Цветковые растения (Magnoliophyta или Angiospermae)
Общая характеристика
Цветковые растения представляют собой самый большой отдел растительного мира, насчитывающий не менее 240 тысяч видов. По числу видов он значительно превосходит все остальные группы высших растений вместе взятые.
Цветковые растения произрастают во всех климатических зонах и в самых разных экологических условиях — от тропических лесов до тундр и полярных пустынь.
Они составляют основную массу растительного вещества биосферы и являются самой важной для человека группой растений. Все важнейшие, культурные растения относятся к цветковым растениям.
Цветковые являются семенными растениями. От голосеменных же они отличаются прежде всего тем, что семязачатки у них заключены в более или менее замкнутую полость завязи (образованную одним или несколькими сросшимися плодолистиками), именно поэтому их чаще всего называют покрытосеменными.
Благодаря тому, что семязачатки заключены в полость завязи, пыльца попадает не непосредственно на микропиле семязачатка, а на рыльце. Наличие рыльца (которое у примитивных форм тянется вдоль шва плодолистика) является характерной особенностью цветковых растений и фактически главным отличием цветка от стробила голосеменных. На этом основании в 1901 г. известный французский ботаник Ван Тигем, предложил называть цветковые растения — рыльцевыми (Stigmateae).
Очень характерны также гаметофиты цветковых растений, которые до крайности редуцированы и миниатюризированы, что позволяет им развиваться значительно, более ускоренными темпами, чем гаметофитам голосеменных. Как женский, так и мужской гаметофиты образуются в результате минимального числа митотических делений. Так, развитие даже относительно более сложного женского гаметофита (так называемого зародышевого мешка) осуществляется путем лишь трех митотических делений. Развитие мужского гаметофита сводится лишь к двум митотическим делениям.
В связи с крайним упрощением гаметофитов цветковых растений и резким сокращением их онтогенеза они утратили гаметангии — антеридии и архегонии. Половое поколение цветковых растений лишилось, таким образом, половых органов. В результате формирование самих гамет (особенно мужских) также ускоряется и они крайне упрощаются.
Одной из характерных особенностей цветковых растений является так называемое двойное оплодотворение, резко отличающих их от всех остальных групп растительного мира. В результате двойного оплодотворения образуется весьма своеобразный триплоидный эндосперм цветковых растений.
Напомним, что в отличие от большинства голосеменных для цветковых характерно наличие в ксилеме наряду с трахеидами сосудов и волокон, которые отсутствуют только у некоторых примитивных групп покрытосеменных. Значительные различия имеются также в строении флоэмы: ситовидные элементы всех цветковых снабжены так называемыми клетками спутницами.
Все это позволяет утверждать, что в целом цветковые растения достигли более высокого уровня эволюционного развития, чем голосеменные. Цветковые представляют собой самую продвинутую в эволюционном отношении группу в мире растений.
Строение цветка
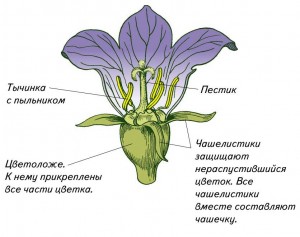
Цветок — это целая система органов, притом система достаточно сложная. Функции цветка многообразны: на разных стадиях его развития в нем происходят микро- и мегаспорогенез, опыление, сложный процесс оплодотворения и не менее сложный процесс образования зародыша и плода.
По современным представлениям, восходящим своими корнями еще к немецким ботаникам А. Брауну и К. Нэгели (конец 19 века), цветок — это укороченный и ограниченный в росте побег, несущий спорофиллы: микроспорофиллы или тычинки и мегаспорофиллы или плодолистики. Основной биологический смысл укорочения побега (как и в случае стробилов голосеменных) заключается в улучшении защиты молодых спорангиев и, особенно, в усовершенствовании процесса опыления.
Часть стебля под цветком, обычно лишенная листьев называется цветоножкой. Иногда имеются видоизмененные листья — прицветники.
Нередко цветоножка сильно укорочена или отсутствует, и тогда цветок называют сидячим.
Цветоножка переходит в укороченную ось цветка, называемую цветоложем. Фактически это типичный укороченный побег, состоящий из тесно сближенных узлов.
На этих узлах и расположены все части цветка. Цветоложе имеет различную форму — выпуклую, плоскую, вогнутую.
Нам будет удобно рассмотреть части цветка, расположенные на выпуклом, коническом цветоложе, двигаясь снизу вверх.
В самой нижней части к цветоложу крепятся чашелистики. В большинстве случаев они служат для защиты цветка, особенно до его распускания в бутоне. Нередко чашелистики становятся лепестковидными и принимают на себя функции лепестков, как у калужницы, ветреницы.
Чашелистики имеют явственную листовую природу: они произошли из верховых вегетативных листьев. Убедительные доказательства тесной морфологической связи чашелистиков с верховыми листьями мы находим, например, у некоторых видов Пиона (Paeonia). Чашелистики здесь как бы продолжают спираль, образуемую вегетативными листьями. При этом листья образуют постепенные переходы к чашелистикам. Это так называемые листовые серии.
Совокупность чашелистиков цветка называется чашечкой (лат. термин calyx). В процессе эволюции чашелистики нередко срастаются между собой, образуя сростнолистную чашечку. Во многих случаях, в частности в простных густых соцветиях, чашечка редуцируется и образует летательный аппарат.
Над чашелистиками размещаются лепестки. В типичном случае их основная функция — привлечение опылителей. Многие современные ученые считают, что в подавляющем большинстве случаев лепестки произошли из тычинок. Классическим примером, иллюстрирующим превращение тычинок в лепестки, является семейство нимфейных. У некоторых представителей этого семейства, например у кувшинки (Nymphaea) в пределах одного и того же цветка наблюдаются постепенные переходы от тычинок к лепесткам. Получается что лепестки — это стерилизовавшиеся тычинки, в которых не происходит развитие микроспор.
Превращение тычинок в лепестки нередко наблюдается в виде отклонений от нормального развития, так называемых уродств. К числу уродств относятся, например махровые формы цветов многих декоративных растений. Таким путем возникли культурные формы розы.
Однако не у всех цветковых растений лепестки имеют бесспорно тычиночное происхождение. Например, у представителей семейства Магнолиевых или у растения Пион лепестки связаны в своем происхождении не с тычинками, а с чашелистиками. Лепестки у этих растений, как и чашелистики, имеют листовое происхождение.
В связи с этим можно различать цветы андропеталы, лепестки которых имеют тычиночную природу, и брантеопеталы — лепестки которых произошли из прицветных листьев — брактей.
Совокупность лепестков цветка называется венчиком (лат. термин corolla). Венчик отличается высокой эволюционной пластичностью, его форма, размеры и окраска имеют очень большое разнообразие. Это связано с биологией опыления. В тоже время у многих, особенно ветроопыляемых растений венчики недоразвиты или вообще отсутствуют.
Также как и чашелистики, лепестки часто срастаются краями, образуя сростнолепестный венчик. Сростнолепестный венчик возник из свободнолепестного независимо в разных линиях эволюции цветковых растений. Таким образом, в процессе эволюции наблюдается общая тенденция к срастанию различных частей цветка.
Чашелистики и лепестки в совокупности составляют околоцветник.
В том случае, когда в цветке имеется как чашечка, так и венчик околоцветник называют двойным.
Если выражены либо только чашечка, либо только венчик, околоцветник называют простым. Соответственно различают простой венчиковидный околоцветник и простой чашевидный околоцветник. Части простого околоцветника, независимо от их природы, называют долями околоцветника.
Следующий круг над околоцветником составляют тычинки.
Как по своей функции, так и по природе тычинки представляют собой разновидность микроспорофиллов.
Самые примитивные формы тычинок, особенно хорошо сохранившиеся у таких растений, как знаменитая дегенерия фиджийская (Degeneria vitiensis), имеют тычинки с широкими трехнервными пластинками, слабо дифференцированными на стерильную и фертильную части.
Такие архаические тычинки снабжены четырьмя попарно сближенными удлиненными микроспорангиями. Они расположены прямо на поверхности пластинки. Примитивные тычинки также у магнолиевых, винтеровых.
В процессе эволюции этот древний тип тычинки, сохранившийся лишь у немногих родов, постепенно дифференцировался на тычиночную нить и пыльник. Пыльник состоит из двух половинок — тек. Теки соединяются между собой посредством связника или коннектива, представляющего продолжение тычиночной нити. Когда связник еще больше редуцируется и почти исчезает, две двухгнездные половинки пыльника соединяются в один четырехгнездный пыльник.
У подавляющего большинства цветковых растений тычинка несет четыре микроспорангия. В зрелом пыльнике часто исчезает перегородка между соседними микроспорангиями и образуется вторично двухгнездный пыльник.
В некоторых специализированных цветках тычинки теряют основную и первоначальную функцию, становятся стерильными и превращаются в стаминодии. Нередко стаминодии представляют промежуточные образования между тычинками и лепестками (как у нимфейных).
Совокупность тычинок в цветке называется андроцеем (от греч. andros — мужчина и oikia — дом). Тычинки могут располагаться в цветке спирально и циклически (по кругу). Спиральный тип расположения тычинок считается более примитивным.
В процессе эволюции цветковых растений обычным явлением считается уменьшение числа тычинок и в некоторых случаях андроцей состоит лишь из одной тычинки.
Нередко также срастаются между собой тычиночные нити, образуя пучок.
Плодолистики
Верхушечную область цветоложа занимают плодолистики. Для вас более знакомо название — пестики. Термин плодолистик явно неудачен, поскольку уже в самом названии органа дана листовая интерпретация его происхождения. Однако этот термин достаточно прочно вошел в литературу.
Возможно, более удачными являются название карпеллы.
Функционально и морфологически плодолистики или карпеллы соответствуют мегаспорофиллам.
В типичном случае плодолистик состоит из следующих структурных частей.
Нижняя расширенная часть плодолистика, содержащая семязачатки, называется завязью. Обычно завязь переходит в длинный вытянутый столбик, который увенчивается рыльцем. Столбик отдельного плодолистика принято называть стилодием, в этом случае термин столбик употребляется только для сросшихся стилодиев.
Наиболее примитивный тип плодолистика сохранился у упомянутого уже рода дегенерия.
Если развернуть плодолистик дегенерии он будет иметь вид листовидной пластинки с тремя жилками. Между средней жилкой и двумя боковыми расположены многочисленные семязачатки. Края наиболее примитивных плодолистиков даже не срастаются. Рыльцевая область у дегенерии протягивается широкой полосой по всей длине плодолистика.
В процессе эволюции рыльцевая область постепенно локализуется в верхней части плодолистика, а верхушка вытягивается в столбик. На первых порах рыльце разделяется еще на так называемые рыльцевые гребни, соответствующие отогнутым краям плодолистика. В конце концов, рыльце локализуется на самой верхушке и становится головчатым.
Типы гинецея
Совокупность плодолистиков в цветке называется гинецеем (от греч. gyne — женщина и oikia — дом). Гинецей цветковых растений эволюционировал от апокарпного (свободного, состоящего из несросшихся плодолистиков) к ценокарпному гинецею, который образован сросшимися плодолистиками.
Однако срастание плодолистиков происходило разными способами. В одном случае, они срастались боковыми стенками, и так образовался синкарпный, многогнездный гинецей. В синкарпном гинецее сохраняется уголковая плацентация семязачатков.
Когда же плодолистики срастались краевыми концами и при этом разворачивались, появлялся паракарпный гинецей. В паракарпном гинецее плодолистики крепятся к стенкам общей вторично-одногнездной завязи и такой способ плацентации называется париетальной.
Лизикарпный гинецей возник из синкарпного в результате лизиса — растворения боковых перегородок. Однако это растворение было не полным, и основания перегородок сохранились в центре лизикарпного гинецея в виде колонки. К этой колонке и крепятся семязачатки. Таким образом, для лизикарпного гинецея характерны вторично одногнездная завязь и колончатая плацентация семязачатков.
Кроме этих трех основных типов различают также псевдомономерный гинецей. Это очень редуцированный вариант синкарпного или парапкарпного гинецея, в котором фертилен и полностью развит лишь один плодолистик. Стерильные плодолистики в псевдомономерном гинецее очень редуцированы и практически не заметны. Типичный псевдомономерный гинецей можно наблюдать у крапивы, конопли, сложноцветных и злаков.В процессе эволюции заметно менялась и завязь цветка.
Наиболее примитивным типом считается верхняя завязь сидящая на коническом цветоложе.
Нектарники
В цветках двудольных и однодольных растений имеются особые железки — нектарники, выделяющие сахаристую жидкость — нектар.
Нектар как выделение привлек внимание ботаников еще со времен Линнея. Линней же и предложил термин nectarium. Нектар служит пищей для многих насекомых, в том числе для пчел, которые перерабатывают его в мед.
Добывая нектар и пыльцу насекомые производят перекрестное опыление цветов. Нектар некоторых тропических и субтропических растений привлекает и птиц. Во всяком случае, известно более 2000 видов растений, опыляемых птицами.
Значение нектарников не ограничивается только этим. Опытами с введением радиоактивных изотопов показано, что нектар используется и самим цветком. Так нектар поступает в область семязачатка, где расходуется на образование зародыша.
Нектар обладает также бактерицидными свойствами.
Основными компонентами нектара являются глюкоза, сахароза и фруктоза в разных соотношениях.
Количество нектара в цветках и концентрация в нем сахаров очень сильно варьируется.
Весьма разнообразны нектарники по величине, форме, размерам, по нахождению и расположению на частях цветка. Чаще всего нектарники возникают из эпидермальных и субэпидермальных клеток, разрастаются в железы и по своему происхождению являются эмергенцами. Нектарники могут развиваться на цветоложе, тычинках, на завязи, на долях околоплодника: лепестках и чашелистиках.
Олигомеризация частей цветка
Если сравнить цветки, характерные для более примитивных семейств, с цветками более продвинутых в эволюционном отношении растений, то можно отметить, что происходит прогрессивное уменьшение всех частей цветка. Уменьшение частей цветка соответствует общему принципу эволюции — закону интеграции гомологов, согласно которому, число однородных частей или гомологов в процессе эволюции уменьшается. Это явление (уменьшение частей цветка) известно под названием олигомеризации.
Олигомеризация является главным, но не единственным направлением эволюции. В некоторых случаях происходил противоположный процесс — полимеризация или увеличение частей цветка. Хорошим примером является семейство кактусовых.
Спиральное и циклическое расположение частей цветка
Примитивные типы цветка характеризуются еще сравнительно длинным цветоложем. В процессе эволюции цветоложе постепенно укорачивается и у наиболее продвинутых групп оно обычно очень короткое.
В связи с укорочением цветоложа и сокращением длины междоузлий части цветка все больше сближаются. Это приводит к переходу первоначально спирального расположения частей цветка к круговому или циклическому. Циклические цветки встречаются гораздо чаще спиральных и характерны для всех сколько-нибудь продвинутых семейств.
www.agrojour.ru
Тема 24. Отдел покрытосеменные, цветковые растения
Основные (ключевые) понятия: покрытосемянность, плод, плодолистик, двойное оплодотворение, пыльца, пыльцевая трубка, семезачаток, типы семезачатков, зародышевый мешок, нуцеллус, цветок, андроцей, гинецей, тычинка, пестик, околоцветник, жизненная форма.
Содержание темы. Характерные признаки покрытосеменных. Важнейшие направления эволюции жизненных форм цветковых растений, систем проводящих тканей и типов стелы, репродуктивных структур-цветков и плодов. Происхождение покрытосеменных, гипотезы и предположения. Теории происхождения цветка и цветковых растений, их критический анализ. Эволюционные ряды признаков покрытосеменных по А Л . Тахтаджяну.
Системы покрытосеменных. Краткий обзор систем Энглера, Кузнецова, Гроссгейма. Обзор системы цветковых растений А.Л. Тахтаджяна. Анализ современных взглядов на классификацию покрытосеменных. Сравнительная характеристика классов двудольные и однодольные.
Класс Двудольные. Характерные особенности, принципы деления на порядки. Порядок магнолиецветные, семейства магнолиевые и дегенериевые: жизненные формы, особенности строения репродуктивных органов, филогенетическое значение.
Порядок лютикоцветные. Семейство лютиковые. Распространение, жизненные формы, краткая ботаническая характеристика. Деление на подсемейства. Важнейшие роды и типичные представители местной флоры. Редкие и нуждающиеся в охране виды на территории Пермской области. Практическое значение семейства.
Порядок макоцветные, характеристика семейства маковые. Порядок розоцветные. Семейство розовые. Распространение, жизненные формы, краткая ботаническая характеристика. Принципы деления на подсемейства. Важнейшие роды и виды дикорастущих и культивируемых растений. Практическое значение семейства.
Порядок бобовоцветные: мимозовые, цезальпиниевые, бобовые, их таксономический статус. Характеристика семейства бобовые ( мотыльковые).
Порядок Зонтикоцветные, важнейшие семейства. Семейство зонтичные. Распространение, жизненные формы, краткая ботаническая характеристика. Важнейшие роды и виды дикорастущих и культивируемых растений. Практическое значение семейства.
Порядок центросеменные, важнейшие семейства - маревые, гвоздичные, кактусовые: распространение, жизненные формы, практическое использование.
Порядок каперцовые, семейство крестоцветные. Распространение, жизненные формы,краткая ботаническая характеристика. Важнейшие роды и виды дикорастущих и культивируемых растений. Охраняемые виды крестоцветных в уральском регионе. Практическое значение семейства.
Порядок мальвоцветные, важнейшие семейства –липовые и мальвовые, их краткая характеристика.
Порядок тыквенные, семейство тыквенные: жизненные формы, краткая ботаническая характеристика,практическое использование.
Порядки березоцветные и букоцветные. Семейства березовые, буковые. Краткая ботаническая характеристика. Важнейшие роды, их лесообразующая роль, использование в озеленении.
Порядок бурачникоцветные, семейство бурачниковые: жизненные формы, краткая ботаническая характеристика. Распространенные роды и виды в составе флоры Пермской области.
Порядок вербеноцветные, семейство губоцветные. Характерные признаки , распространение, практическое значение.
Порядок пасленоцветные, семейство паслёновые: характерные признаки, распространение, практическое значение.
Порядок норичникоцветные, семейство норичниковые: специфика жизненных форм, краткая ботаническая характеристика, типичные представители. Семейства тропических и субтропических областей- акантовые и геснериевые, их краткая характеристика.
Порядок сростнопыльниковые (Астровые), семейство сложноцветные. Характерные признаки и объем семейства, деление на подсемейства и трибы. Участие и роль в растительном покрове. Ведущие по числу видов роды .Охраняемые растения на территории Пермского края. Практическое значение семейства.
Класс Однодольные. Общая характеристика, отличительные особенности. Взгляды на происхождение, важнейшие порядки.
Порядок лилиецветные, семейство лилейные: распространение, жизненные формы, важнейшие подсемейства, их краткая характеристика. Практическое значение семейства.
Порядок чешуецветные, семейство злаки: объем семейства, распространение и роль в сложении растительного покрова, ботаническая характеристика, практическое значение. Наиболее крупные подсемейства, их краткая характеристика, типичные представители.
5.2. Аннотация теоретического курса дисциплины (аннотация представляет собой краткое содержание лекционного курса дисциплины.)
studfiles.net
Цветковые растения - это... Что такое Цветковые растения?
Цветко́вые расте́ния, или Покры́тосеменны́е (лат. Magnoliophyta, или Angiospermae от др.-греч. ἀγγεῖον — сосуд, σπέρμα — семя) — отдел высших растений, отличительной особенностью которых является наличие цветка в качестве органа полового размножения и замкнутого вместилища у семяпочки (а затем и у происшедшего из неё семени, откуда и появилось название покрытосеменные). Ещё одна существенная особенность цветковых растений — двойное оплодотворение.
Покрытосеменные, наряду с голосеменными (Gymnospermae) составляют одну из двух групп семенных растений (Spermatophytae).
В 1999 году Амборелловые (Amborellaceae) были идентифицированы как базальная группа цветковых растений[1].
Объём таксона
По числу видов цветковые растения значительно превосходят все остальные группы высших растений, вместе взятые.
В литературе приходятся разные данные о числе современных видов цветковых растений. В 2009 году была опубликована работа австралийского учёного Чапмена (A. D. Chapman) «Numbers of Living Species in Australia and the World», в которой он даёт обзор мнений по этому вопросу и делает вывод, что по состоянию на 2009 год можно оценить общее число описанных современных видов цветковых растений примерно в 269 тысяч, а общее число современных видов цветковых растений на нашей планете — примерно в 350 тысяч[2].
Число видов покрытосеменных, по данным Angiosperm Phylogeny Website на февраль 2010 года, составляет 271—272 тысячи, число родов — 13350—13400[3].
Число семейств и порядков сильно отличается в зависимости от классификаций; Система классификации APG III (2009) выделяет 414 семейств, которые, в свою очередь, объединены в 59 порядков.
Морфологические особенности
Важнейшая особенность цветковых растений — наличие специализированного генеративного органа — цветка, берущего на себя функции полового размножения и привлечения агентов опыления. Цветковые растения заключают свои семязачатки (семяпочки) в полость завязи, которая образована срастанием когда-то открытого плодолистика. Стенки завязи после оплодотворения разрастаются и видоизменяются, давая образование под названием плод.
В другой группе семенных растений, у голосеменных (Pinophyta, или Gymnospermae), семязачаток не скрыт от опыления, а семена не заключены в истинный плод, но иногда семя могут покрывать мясистые структуры, например, у представителей рода Тис.
Происхождение
 | ||||||||||
| Амборелла волосистоножковая (Amborella trichopoda), эндемик Новой Каледонии, одно из наиболее древних цветковых растений, сохранившихся до наших дней. Отделившись от других цветковых растений примерно 130 миллионов лет назад, амборелла с тех пор изменилась весьма незначительно, что можно объяснить относительной изолированностью экосистемы Новой Каледонии | ||||||||||
Первые остатки покрытосеменных датируются меловым периодом приблизительно 140 миллионов лет назад. Базируясь на современных данных, можно предположить, что предки покрытосеменных и гнетовых дивергировали в триасе (220—202 миллионов лет назад). Первые отпечатки растений с признаками покрытосеменных обнаружены в пластах юрского и раннего мелового периодов (135-65 миллионов лет назад), но это были довольно-таки малочисленные и примитивные формы. Древнейшими покрытосеменными являются растения из группы нимфейных[4]. Следы широкого развития и распространения покрытосеменных появились в палеонтологической летописи в период среднего мела (около 100 миллионов лет назад). Но уже в позднем мелу покрытосеменные оказались доминирующей формой растительной жизни, и в многих фоссилиях узнаются представители современных семейств (например, бук, дуб, клён и магнолия)
Одно из важнейших направлений эволюции растительного царства — приспособление к изменчивым условиям наземной жизни. Цветковые растения являются ярчайшим выражением этой линии и доминируют на земной поверхности в данную эпоху.
К широчайшему географическому разнообразию прибавляется разнообразие форм и способов роста. Банальная ряска, покрывающая поверхность пруда, представляет собой крошечный зелёный побег с простым корешком, вертикально погруженным в воду, и с очень нечёткими листиками и частями стебля. Могучее лесное дерево столетия развивало свою сложную систему стволов и ветвей, покрытых бесчисленными веточками и листвой, а под землёй соответствующую площадь занимает мощная, хорошо развитая корневая система. Между этими двумя крайностями — бесконечные градации: водные и земные травы, ползучие, прямостоящие или карабкающиеся, кусты и деревья, гораздо большее разнообразие, чем в другом отделе семенных растений — Gymnospermae.
Известны многочисленные водные покрытосеменные растения, они в изобилии встречаются в долинах рек и чистых озёрах, в меньшем количестве — в солёных озёрах и морях. Однако такие водные покрытосеменные не являются примитивными формами, а возникли путём приспособления наземного предка к водной среде.
Покрытосеменные
Систематическое положение
Цветковые растения обычно рассматриваются как отдел. Так как эта систематическая категория более высокого ранга, чем семейство, есть определённая свобода в выборе названия. Статья 16 Международного Кодекса Ботанической Номенклатуры позволяет использовать как и традиционные исторические названия, так и название, образованное от рода. Официальное униноминальное название этого таксона — Magnoliophyta, от названия рода Magnolia. Но традиционно укоренились такие имена, как Angiospermae и Anthophyta (цветковые растения).
Классификация
 | Информация в этой статье или некоторых её разделах устарела. Вы можете помочь проекту, обновив её и убрав после этого данный шаблон. |
История понятия
Ботанический термин «Angiospermae» предложил Пауль Герман в 1690 году, термин был составлен из греческих слов ἀγγεῖον (вместилище, сосуд, полость) и σπέρμα (семя). Так Герман называл один из главнейших отделов растительного царства, включавший в себя растения, обладающие заключёнными в капсулы семенами. Gymnospermae же по Герману являлись цветковыми растениями, чей шизокарпичный или единственный цельный плод считались семенами с отсутствующими покровами. Сам термин и его антоним были подхвачены Карлом Линнеем, который использовал их в похожем, но более ограниченном смысле — для названий порядков своего класса Двусильных (Didynamia). В своём современном значении эти термины начали использоваться после того как Роберт Броун в 1827 году установил существование истинно голых семязачатков у Cycadeae и Coniferae, присвоив им название Gymnospermae. С этого времени термин Angiospermae стал использоваться различными авторами, иногда с разными вариациями, для обозначения одной из подгрупп в пределах двудольных растений (разделение «явнобрачных» растений на бессемядольные, однодольные и двудольные получило широкое распространение несколько раньше).
Однако, после того как Вильгельм Хофмейстер описал процессы, происходящие в зародышевом мешке цветковых растений (1851 год), и сопоставил их с оплодотворением тайнобрачных, стало ясно, что Gymnospermae представляют собой группу, совершенно отличную от Angiospermae. В результате понятие «покрытосеменные» постепенно стали рассматривать как синоним понятия «цветковые», и, соответственно, двудольные (Magnoliopsida, или Dicotyledones) и однодольные (Liliopsida, или Monocotyledones) — как подгруппы в составе Angiospermae. В этом значении понятие «покрытосеменные» (Angiospermae) используется и по сей день.
Благодаря постоянному пересмотру взглядов на родство цветковых растений, внутренняя систематика этой группы подвергалась и подвергается изменениям. Две широко используемые, хотя и несколько устаревшие, системы цветковых растений — система Тахтаджяна и система Кронквиста, не отражают филогению таксона. Таким образом, классификация цветковых растений сейчас активно дорабатывается и исправляется.
Отдел цветковых растений традиционно подразделяется на 2 класса — Magnoliopsida (двудольные) от названия рода Magnolia и Liliopsida (однодольные) от названия рода Lilium. Используются также и традиционные названия этих таксонов — Dicotyledones и Monocotyledones. Происхождение этих названий связано с тем, что представители Dicotyledones обычно имеют две семядоли в семени (у некоторых видов может быть одна, три или четыре), тогда как у представителей Monocotyledones семядоля всегда одна.
Имеется целый ряд современных систем классификации цветковых растений. Классы двудольных и однодольных могут подразделяться на подклассы, те, в свою очередь, на порядки (иногда объединяемые в надпорядки), семейства, роды и виды со всеми промежуточными категориями.
Класс Двудольные
| | Информация в этой статье или некоторых её разделах устарела. Вы можете помочь проекту, обновив её и убрав после этого данный шаблон. |
В классе Двудольные (Magnoliopsida, или Dicotyledones) описано 6 подклассов, 128 порядков, 418 семейств, приблизительно 10 000 родов и около 199 000 видов[5] растений.
Выделяют подклассы[источник не указан 931 день]:
Класс Однодольные
| | Информация в этой статье или некоторых её разделах устарела. Вы можете помочь проекту, обновив её и убрав после этого данный шаблон. |
В класс однодольных растений (Liliopsida, или Monocotyledones) включаются 5 подклассов, 37 порядков, около 125 семейств, более 3 000 родов и около 59 000 видов[6] .
Выделяют подклассы[источник не указан 931 день]:
- Подкласс 1. Алисматиды (Alismatidae)
- Подкласс 2. Лилииды (Liliidae)
- Подкласс 3. Коммелиниды (Commelinidae)
- Подкласс 4. Арециды (Arecidae)
- Подкласс 5. Зингибериды (Zingiberidae)
Филогения
Предполагается, что Однодольные эволюционно более прогрессивны, чем двудольные.
Реликтовые представители покрытосеменных
Некоторые из наиболее примитивных представителей ныне живущих двудольных, сохранили архаичное строение проводящей системы, тычинок, пыльцевых зерен, плодолистиков, и других органов.
Часть видов семейства Винтеровые (Winteraceae), роды троходендрон (Trochodendron) и тетрацентрон (Tetracentron), по строению проводящей системы мало чем отличаются от примитивных представителей голосеменных. У них отсутствуют проводящие сосуды во всех органах, их роль выполняют трахеиды.
Другие демонстрируют архаичное строение цветка, в частности примитивные строение тычинок; особенно заметное у таких растений, как дегенерия (Degeneria vitiensis), гальбулимима (Galbulimima) и ряда представителей магнолиевых и винтеровых[7].
Настоящими «живыми ископаемыми» являются виды амборелла (Amborella), дегенерия (Degeneria), австробэйлия (Austrobaileya), буббия (Bubbia), эвпоматия (Eupomatia)[8].
См. также
Примечания
- ↑ Ботаника. Учебник для вузов: в 4 т = Lehrbuch der Botanik für Hochschulen. Begründet von E. Strasburger, F. Noll, H. Schenck, A. F. W. Schimper. / 35. Auflage neubearbeitet von Peter Sitte, Elmar W. Weiler, Joachim W. Kadereit, Andreas Bresinsky, Christian Körner / П. Зитте, Э. В. Вайлер, Й. В. Кадерайт, А. Брезински, К. Кёрнер; на основе учебника Э. Страсбургера [и др.]; пер. с нем. Е. Б. Поспеловой, К. Л. Тарасова, Н. В. Хмелевской. — М.: Издательский центр «Академия», 2007. — Т. 3. Эволюция и систематика / под ред. А. К. Тимонина, И. И. Сидоровой. — С. 15. — 576 с. — 3000 экз. — ISBN 978-5-7695-2741-8 (рус.) ISBN 978-5-7695-2746-3 (Т. 3) (рус.), ISBN 3-8274-1010-X (Elsevier GmbH) — УДК 58(075.8)
- ↑ A. D. Chapman. Numbers of Living Species in Australia and the World. 2nd edition (англ.) (Проверено 29 мая 2010)
- ↑ Angiosperm Phylogeny Website (англ.). Число родов и видов приведено отдельно для каждого порядка. См. также Семейства покрытосеменных (APG III).
- ↑ Происхождение основных групп наземных растений
- ↑ Hamilton, Alan & Hamilton, Patrick (2006), «Plant conservation : an ecosystem approach», London: Earthscan, сс. 2, ISBN 978-1-84407-083-1, <http://books.google.com/?id=P6m0OTheY8sC&printsec=frontcover#PPA2,M1>
- ↑ http://cmsdata.iucn.org/downloads/2008rl_stats_tables_all.xls
- ↑ Фёдоров А. А., Тахтаджян А. Л. и др. Жизнь растений. Том 5(1). Цветковые растения — М.: Просвещение, 1980. — с. 117—127.
- ↑ Смирнов А. В. Мир растений. Рассказы о саксауле, селитрянке, баобабе, березах, кактусах, капусте, банксиях, молочаях и многих других широко известных и редких цветковых растениях. — М.: Молодая гвардия, 1979. — с. 14-18
Литература
- Тахтаджян А. Л. Система и филогения цветковых растений / Академия наук СССР. Ботанический институт имени В. Л. Комарова. — М.—Л.: Наука, 1966. — 611 с. — 4 300 экз.
- Angiosperm Phylogeny Group (2009) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III // Botanical Journal of the Linnean Society : журнал. — Лондон, 2009. — Т. 161. — № 2. — С. 105—121. — DOI:10.1111/j.1095-8339.2009.00996.x
Ссылки
Система классификации APG III (2009)
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| •• Angiosperm Phylogeny Group (в том числе Бремер Б., Бремер К., Ривил Д., Чейз М.) • APG I • APG II (+список) • APG III (+список) •• |
 angiosperms
angiospermsdik.academic.ru
Цветковые - это... Что такое Цветковые?
Цветко́вые расте́ния, покры́тосеменны́е (лат. Magnoliophyta, или Angiospermae) — отдел высших растений, имеющих цветок.
Эта важнейшая группа наземных растений насчитывает свыше 165 порядков, 540 семейств, более 13 000 родов и, вероятно, не менее 250 000 видов. По числу видов цветковые растения значительно превосходят все остальные группы высших растений, вместе взятые. Они составляют одну из двух групп семенных растений.Морфологические особенности
Важнейшая особенность цветковых растений — наличие специализированного генеративного органа — цветка, берущего на себя функции полового размножения и привлечения агентов опыления. Цветковые растения заключают свои семязачатки (семяпочки) в полость завязи, которая образована срастанием когда-то открытого плодолистика. Стенки завязи после оплодотворения разрастаются и видоизменяются, давая образование под названием плод.
В другой группе семенных растений, Голосеменные (Pinophyta, или Gymnospermae), семязачаток не скрыт от опыления, а семена не заключены в истинный плод, но иногда семя могут покрывать мясистые структуры, например, у представителей рода Тис.
Размножение
Происхождение
Первые остатки покрытосеменных датируются Юрским периодом приблизительно 140 миллионов лет назад. Базируясь на современных данных, можно предположить, что предки покрытосеменных и гнетовых дивергировали в триасе (220—202 миллионов лет назад). Первые отпечатки растений с признаками покрытосеменных обнаружены в пластах юрского и раннего мелового периодов (135-65 миллионов лет назад), но это были довольно-таки малочисленные и примитивные формы. Древнейшими покрытосеменными являются растения из группы нимфейных[1]. Следы широкого развития и распространения покрытосеменных появились в палеонтологической летописи в период среднего мела (около 100 миллионов лет назад). Но уже в позднем мелу покрытосеменные оказались доминирующей формой растительной жизни, и в многих фоссилиях узнаются представители современных семейств (например, бук, дуб, клен и магнолия)
Одно из важнейших направлений эволюции растительного царства — приспособление к изменчивым условиям наземной жизни. Цветковые растения являются ярчайшим выражением этой линии и доминируют на земной поверхности в данную эпоху. От полюсов до экватора нет такого участка, где возможна растительная жизнь, но не найдено покрытосеменных.
К широчайшему географическому разнообразию прибавляется разнообразие форм и способов роста. Банальная ряска, покрывающая поверхность пруда, представляет собой крошечный зелёный побег с простым корешком, вертикально погруженным в воду, и с очень нечёткими листиками и частями стебля. Могучее лесное дерево столетия развивало свою сложную систему стволов и ветвей, покрытых бесчисленными веточками и листвой, а под землёй соответствующую площадь занимает мощная, хорошо развитая корневая система. Между этими двумя крайностями — бесконечные градации: водные и земные травы, ползучие, прямостоящие или карабкающиеся, кусты и деревья, гораздо большее разнообразие, чем в другом отделе семенных растений — Gymnospermae.
Известны многочисленные водные покрытосеменные растения, они в изобилии встречаются в долинах рек и чистых озёрах, в меньшем количестве — в солёных озёрах и морях. Однако такие водные покрытосеменные не являются примитивными формами, а возникли путём приспособления наземного предка к водной среде.
Цветковые растения
Систематическое положение
Цветковые растения обычно рассматриваются как отдел. Так как эта систематическая категория более высокого ранга, чем семейство, есть определённая свобода в выборе названия. Статья 16 Международного Кодекса Ботанической Номенклатуры позволяет использовать как и традиционные исторические названия, так и название, образованное от рода. Официальное униноминальное название этого таксона — Magnoliophyta, от названия рода Magnolia. Но традиционно укоренились такие имена, как Angiospermae и Anthophyta (цветковые растения).
Классификация
История понятия
Ботанический термин «Angiospermae» предложил Пауль Герман в 1690 году, термин был составлен из греческих слов ἀγγεῖον (вместилище, сосуд, полость) и σπέρμα (семя). Так Герман называл один из главнейших отделов растительного царства, включавший в себя растения, обладающие заключёнными в капсулы семенами. Gymnospermae же по Герману являлись цветковыми растениями, чей шизокарпичный или единственный цельный плод считались семенами с отсутствующими покровами. Сам термин и его антоним были подхвачены Карлом Линнеем, который использовал их в похожем, но более ограниченном смысле — для названий порядков своего класса Двусильных (Didynamia). В своём современном значении эти термины начали использоваться после того как Роберт Броун в 1827 году установил существование истинно голых семязачатков у Cycadeae и Coniferae, присвоив им название Gymnospermae. С этого времени термин Angiospermae стал использоваться различными авторами, иногда с разными вариациями, для обозначения одной из подгрупп в пределах двудольных растений (разделение «явнобрачных» растений на бессемядольные, однодольные и двудольные получило широкое распространение несколько раньше).
Однако, после того как Вильгельм Хофмейстер описал процессы, происходящие в зародышевом мешке цветковых растений (1851 год), и сопоставил их с оплодотворением тайнобрачных, стало ясно, что Gymnospermae представляют собой группу, совершенно отличную от Angiospermae. В результате понятие «покрытосеменные» постепенно стали рассматривать как синоним понятия «цветковые», и, соответственно, двудольные (Magnoliopsida, или Dicotyledones) и однодольные (Liliopsida, или Monocotyledones) — как подгруппы в составе Angiospermae. В этом значении понятие «покрытосеменные» (Angiospermae) используется и по сей день.
Благодаря постоянному пересмотру взглядов на родство цветковых растений, внутренняя систематика этой группы подвергалась и подвергается изменениям. Две широко используемые, хотя и несколько устаревшие, системы цветковых растений — система Тахтаджяна и система Кронквиста, не отражают филогению таксона. Таким образом, классификация цветковых растений сейчас активно дорабатывается и исправляется.
Отдел цветковых растений традиционно подразделяется на 2 класса — Magnoliopsida (двудольные) от названия рода Magnolia и Liliopsida (однодольные) от названия рода Lilium. Более популярны традиционные названия этих таксонов — Dicotyledones и Monocotyledones. Происхождение этих названий вполне очевидно: Dicotyledones имеют две семядоли в семени, тогда как у Monocotyledones семядоля одна.
Классы двудольных и однодольных, в свою очередь, подразделяются на подклассы, которые делятся на порядки (иногда объединяемые в надпорядки), семейства, роды и виды со всеми промежуточными категориями. Имеется целый ряд современных систем классификации цветковых растений.
Класс Двудольные
В классе Двудольные (Magnoliopsida, или Dicotyledones) описано 8 подклассов, 128 порядков, 418 семейств, приблизительно 10 000 родов и около 190 000 видов двудольных растений. Выделяют подклассы:
- Подкласс 1. Магнолиевые (Magnoliidae)
- Подкласс 2. Гамамелисовые (Hamamelididae)
- Подкласс 3. Гвоздичные (Caryophyllidae)
- Подкласс 4. Диллениевые (Dilleniidae)
- Подкласс 5. Розоцветные (Rosidae)
- Подкласс 6. Сложноцветные (Asteridae)
Класс Однодольные
В класс однодольных растений (Liliopsida, или Monocotyledones) включаются 5 подклассов, 37 порядков, около 120 семейств, 30 000 родов и больше 60 000 видов. Выделяют подклассы:
- Подкласс 1. Частуховые (Alismatidae)
- Подкласс 2. Лилейные (Liliidae)
- Подкласс 3. Коммелиновые (Commelinidae)
- Подкласс 4. Арековые (Arecidae)
- Подкласс 5. Имбирные (Zingiberidae)
Литература
Примечания
Ссылки
Wikimedia Foundation. 2010.
dikc.academic.ru
Цветковые - это... Что такое Цветковые?
Цветко́вые расте́ния, покры́тосеменны́е (лат. Magnoliophyta, или Angiospermae) — отдел высших растений, имеющих цветок.
Эта важнейшая группа наземных растений насчитывает свыше 165 порядков, 540 семейств, более 13 000 родов и, вероятно, не менее 250 000 видов. По числу видов цветковые растения значительно превосходят все остальные группы высших растений, вместе взятые. Они составляют одну из двух групп семенных растений.Морфологические особенности
Важнейшая особенность цветковых растений — наличие специализированного генеративного органа — цветка, берущего на себя функции полового размножения и привлечения агентов опыления. Цветковые растения заключают свои семязачатки (семяпочки) в полость завязи, которая образована срастанием когда-то открытого плодолистика. Стенки завязи после оплодотворения разрастаются и видоизменяются, давая образование под названием плод.
В другой группе семенных растений, Голосеменные (Pinophyta, или Gymnospermae), семязачаток не скрыт от опыления, а семена не заключены в истинный плод, но иногда семя могут покрывать мясистые структуры, например, у представителей рода Тис.
Размножение
Происхождение
Первые остатки покрытосеменных датируются Юрским периодом приблизительно 140 миллионов лет назад. Базируясь на современных данных, можно предположить, что предки покрытосеменных и гнетовых дивергировали в триасе (220—202 миллионов лет назад). Первые отпечатки растений с признаками покрытосеменных обнаружены в пластах юрского и раннего мелового периодов (135-65 миллионов лет назад), но это были довольно-таки малочисленные и примитивные формы. Древнейшими покрытосеменными являются растения из группы нимфейных[1]. Следы широкого развития и распространения покрытосеменных появились в палеонтологической летописи в период среднего мела (около 100 миллионов лет назад). Но уже в позднем мелу покрытосеменные оказались доминирующей формой растительной жизни, и в многих фоссилиях узнаются представители современных семейств (например, бук, дуб, клен и магнолия)
Одно из важнейших направлений эволюции растительного царства — приспособление к изменчивым условиям наземной жизни. Цветковые растения являются ярчайшим выражением этой линии и доминируют на земной поверхности в данную эпоху. От полюсов до экватора нет такого участка, где возможна растительная жизнь, но не найдено покрытосеменных.
К широчайшему географическому разнообразию прибавляется разнообразие форм и способов роста. Банальная ряска, покрывающая поверхность пруда, представляет собой крошечный зелёный побег с простым корешком, вертикально погруженным в воду, и с очень нечёткими листиками и частями стебля. Могучее лесное дерево столетия развивало свою сложную систему стволов и ветвей, покрытых бесчисленными веточками и листвой, а под землёй соответствующую площадь занимает мощная, хорошо развитая корневая система. Между этими двумя крайностями — бесконечные градации: водные и земные травы, ползучие, прямостоящие или карабкающиеся, кусты и деревья, гораздо большее разнообразие, чем в другом отделе семенных растений — Gymnospermae.
Известны многочисленные водные покрытосеменные растения, они в изобилии встречаются в долинах рек и чистых озёрах, в меньшем количестве — в солёных озёрах и морях. Однако такие водные покрытосеменные не являются примитивными формами, а возникли путём приспособления наземного предка к водной среде.
Цветковые растения
Систематическое положение
Цветковые растения обычно рассматриваются как отдел. Так как эта систематическая категория более высокого ранга, чем семейство, есть определённая свобода в выборе названия. Статья 16 Международного Кодекса Ботанической Номенклатуры позволяет использовать как и традиционные исторические названия, так и название, образованное от рода. Официальное униноминальное название этого таксона — Magnoliophyta, от названия рода Magnolia. Но традиционно укоренились такие имена, как Angiospermae и Anthophyta (цветковые растения).
Классификация
История понятия
Ботанический термин «Angiospermae» предложил Пауль Герман в 1690 году, термин был составлен из греческих слов ἀγγεῖον (вместилище, сосуд, полость) и σπέρμα (семя). Так Герман называл один из главнейших отделов растительного царства, включавший в себя растения, обладающие заключёнными в капсулы семенами. Gymnospermae же по Герману являлись цветковыми растениями, чей шизокарпичный или единственный цельный плод считались семенами с отсутствующими покровами. Сам термин и его антоним были подхвачены Карлом Линнеем, который использовал их в похожем, но более ограниченном смысле — для названий порядков своего класса Двусильных (Didynamia). В своём современном значении эти термины начали использоваться после того как Роберт Броун в 1827 году установил существование истинно голых семязачатков у Cycadeae и Coniferae, присвоив им название Gymnospermae. С этого времени термин Angiospermae стал использоваться различными авторами, иногда с разными вариациями, для обозначения одной из подгрупп в пределах двудольных растений (разделение «явнобрачных» растений на бессемядольные, однодольные и двудольные получило широкое распространение несколько раньше).
Однако, после того как Вильгельм Хофмейстер описал процессы, происходящие в зародышевом мешке цветковых растений (1851 год), и сопоставил их с оплодотворением тайнобрачных, стало ясно, что Gymnospermae представляют собой группу, совершенно отличную от Angiospermae. В результате понятие «покрытосеменные» постепенно стали рассматривать как синоним понятия «цветковые», и, соответственно, двудольные (Magnoliopsida, или Dicotyledones) и однодольные (Liliopsida, или Monocotyledones) — как подгруппы в составе Angiospermae. В этом значении понятие «покрытосеменные» (Angiospermae) используется и по сей день.
Благодаря постоянному пересмотру взглядов на родство цветковых растений, внутренняя систематика этой группы подвергалась и подвергается изменениям. Две широко используемые, хотя и несколько устаревшие, системы цветковых растений — система Тахтаджяна и система Кронквиста, не отражают филогению таксона. Таким образом, классификация цветковых растений сейчас активно дорабатывается и исправляется.
Отдел цветковых растений традиционно подразделяется на 2 класса — Magnoliopsida (двудольные) от названия рода Magnolia и Liliopsida (однодольные) от названия рода Lilium. Более популярны традиционные названия этих таксонов — Dicotyledones и Monocotyledones. Происхождение этих названий вполне очевидно: Dicotyledones имеют две семядоли в семени, тогда как у Monocotyledones семядоля одна.
Классы двудольных и однодольных, в свою очередь, подразделяются на подклассы, которые делятся на порядки (иногда объединяемые в надпорядки), семейства, роды и виды со всеми промежуточными категориями. Имеется целый ряд современных систем классификации цветковых растений.
Класс Двудольные
В классе Двудольные (Magnoliopsida, или Dicotyledones) описано 8 подклассов, 128 порядков, 418 семейств, приблизительно 10 000 родов и около 190 000 видов двудольных растений. Выделяют подклассы:
- Подкласс 1. Магнолиевые (Magnoliidae)
- Подкласс 2. Гамамелисовые (Hamamelididae)
- Подкласс 3. Гвоздичные (Caryophyllidae)
- Подкласс 4. Диллениевые (Dilleniidae)
- Подкласс 5. Розоцветные (Rosidae)
- Подкласс 6. Сложноцветные (Asteridae)
Класс Однодольные
В класс однодольных растений (Liliopsida, или Monocotyledones) включаются 5 подклассов, 37 порядков, около 120 семейств, 30 000 родов и больше 60 000 видов. Выделяют подклассы:
- Подкласс 1. Частуховые (Alismatidae)
- Подкласс 2. Лилейные (Liliidae)
- Подкласс 3. Коммелиновые (Commelinidae)
- Подкласс 4. Арековые (Arecidae)
- Подкласс 5. Имбирные (Zingiberidae)
Литература
Примечания
Ссылки
Wikimedia Foundation. 2010.
med.academic.ru