1. Корневые системы. Вторичные изменения корня. Оплодотворение у покрытосеменных растений
1. Корневые системы. Вторичные изменения корня.
Подобно побегу , корень способен к ветвлению. В результате образуется корневая система, под которой понимают совокупность корней одного растения. Характер корневой системы определяется соотношением роста главного, боковых и придаточных корней.
Первый корень семенного растения развивается из зародышевого корешка . Он называется главным. У двудольных и голосеменных от главного корня отходят боковые корни первого порядка, в свою очередь дающие начало боковым корням второго порядка и т.д. В результате формируется стержневая или ее разновидность - ветвистая корневая система. Все боковые корни семенных растений закладываются эндогенно, т.е. внутри корня предшествующего порядка, и развиваются из перицикла . У однодольных зародышевый корешок живет относительно короткое время, в силу чего главный корень не развивается. Вместо него при основании побега образуются так называемые придаточные корни (придаточные корни могут возникать также на листьях), более или менее сходные между собой по размерам. Они, в свою очередь, могут давать боковые корни. Корневая система, сформированная подобным образом, получила название мочковатой. Придаточные корни также закладываются эндогенно.
У многих двудольных главный корень также нередко отмирает и заменяется системой придаточных корней, отходящих от корневища.
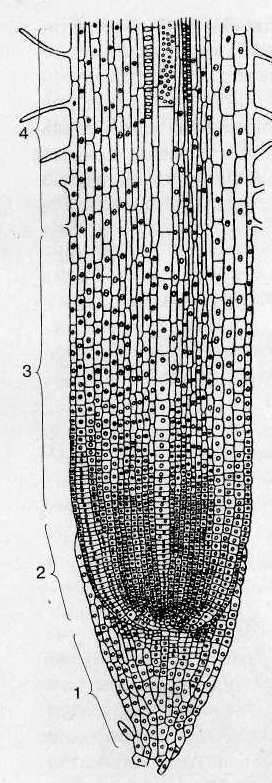
У однодольных и папоротников первичная структура корня сохраняется в течение всей жизни и вторичные ткани не возникают. Иначе обстоит дело с голосеменными и двудольными , у которых в дальнейшем происходят вторичные изменения и в конечном итоге формируется вторичная структура корня, при которой радиальное расположение проводящих тканей заменяется коллатеральным. Образование вторичной структуры корня связано прежде всего с деятельностью камбия , который обеспечивает рост корня в толщину. Камбий возникает из тонкостенных паренхимных клеток в виде разобщенных участков с внутренней стороны тяжей флоэмы между лучами первичной ксилемы . Камбиальную активность вскоре приобретают и некоторые участки перицикла , располагающиеся кнаружи от лучей первичной ксилемы. В результате образуется непрерывный камбиальный слой
2. Опыление и оплодотворение у покрытосеменных растений. Эндосперм и зародыш семени.
Опыление
Опыление - необходимое условие оплодотворения, происходящего в цветке. Суть опыления состоит в переносе пыльцы из пыльников на рыльце пестика.
Различают два принципиально различных типа опыления: самоопыление и перекрестное. При самоопылении пыльца переносится на рыльце пестика в пределах данного цветка или данной особи.
Самоопыление, особенно постоянное, рассматривается как вторичное явление, вызванное неблагоприятными условиями среды, т.е. неблагоприятными для перекрестного опыления; оно выполняет страхующую роль. Постоянное самоопыление трактуется как тупик эволюционного развития. Самоопыление чаще бывает у однолетников с коротким жизненным циклом, растущих в неблагоприятных экологических условиях на сухих и бедных почвах (пастушья сумка, клевер шершавый, клевер скученный). Такой вид опыления позволяет им быстрее восстановить численность вида. Кроме того, самоопыление характерно для растений, где невозможно перекрестное опыление. Этот вид опыления осуществляется, когда тычинки постоянно соприкасаются с пестиком (копытень, седмичник). Чаще самоопыление наблюдается у растений темнохвойного леса (брусника, черника). У кислицы, растущей в темнохвойном лесу, где большая влажность воздуха и почти нет опыляющих насекомых, опыление происходит, когда цветки еще в бутоне, - клейстогамия.
Перекрестное опыление - основной тип опыления цветковых растений. Оно биологически более совершенно. У растений есть специальные устройства морфологического и физиологического характера, предотвращающие или ограничивающие самоопыление и, следовательно, способствующие перекрестному опылению. К ним относится двудомность растений, т. е. разновременное созревание тычинок или пестиков: протандрия - раньше созревает пыльца (колокольчики, гвоздики, сложноцветные), протогиния - раньше созревает рыльце (крестоцветные, розоцветные).
Различают два вида перекрестного опыления: биотическое и абиотическое.
Биотическое опыление: энтомофилия, орнитофилия, хироптерофилия. Абиотическое - анемофилия, гидрофилия. При перекрестном опылении цветки имеют приспособления для опыления агентами живой и неживой природы.
Анемофилия - опыление с помощью ветра. Оно представляет собой адаптацию цветковых растений к неблагоприятным условиям, которые ограничивают возможность биотического опыления. Ветроопыляемые растения цветут до распускания листьев (лещина, береза), их цветки без околоцветника, без запаха и окраски (невзрачные), но имеют перистое рыльце. Цветки собраны в соцветия (сережка, кисть, колос). Тычинки свободно свисающие.
Энтомофилия - опыление с помощью насекомых. Насекомые посещают цветки для сбора пыльцы, нектара, а иногда в поисках убежища, отложения яиц, поиска партнера. Растения для опыления насекомыми имеют ряд приспособлений. Так, мак, пион, шиповник, зверобой образуют много пыльцы и этим привлекают жуков, которые питаются пыльцой. Нектар растения выделяют в разном количестве и в разное время дня, что помогает насекомым находить его в любое время. В привлечении насекомых большое значение имеют запах и окраска растений. Цветовая гамма цветков разнообразна, и восприятие цвета у насекомых отличается от восприятия человека
Оплодотворение
Процессу оплодотворения предшествует прорастание пыльцевого зерна на рыльце. Микроспора еще внутри пыльника прорастает и ее ядро подвергается митотическому делению, в результате которого образуется маленькая репродуктивная клетка и большая вегетативная. Попав на пестик, вегетативная клетка вытягивается в пыльцевую трубку, а ядро генеративной клетки делится и дает две мужские клетки спермии. На следующем этапе пыльцевая трубка проникает в зародышевый мешок, разрывается, и из нее выходят гаметы. Один спермий сливается с яйцеклеткой (зигота диплоидна), другой - со вторичным ядром. Такое оплодотворение называется двойным. Это выдающееся открытие принадлежит русскому ученому С. Г. Навашину. Из оплодотворенной клетки (зиготы) развивается зародыш будущего растения. Из вторичного ядра зародышевого мешка - эндосперм семени, иными словами, семя образуется из семяпочки после оплодотворения. Из покровов семяпочки развивается кожура семени, из зиготы - многоклеточный зародыш, из триплоидной клетки - запасающая ткань эндосперм. Возможно, возникновение в семени запасающей ткани из нуцеллуса (центральной части семяпочки). Эту ткань называют периспермом.
8БИЛЕТ
studfiles.net
Вопрос 14. Спорогенез, гаметогенез и оплодотворение у покрытосеменных растений.
Двойное оплодотворение у покрытосеменных растений.
Пыльцевая трубка проникает в семязачаток. Спермии при помощи пыльцевой трубки переносятся к зародышевому мешку.
Первый спермий сливается с яйцеклеткой (n) с образованием зиготы (2n), впоследствии – зародыша.
Второй спермий сливается с центральной клеткой (2n) с образованием эндосперма (3n), запас питательных веществ.
После оплодотворения из семязачатка образуется семя, а из завязи – плод. Зигота растет, путем митотических делений превращается в многоклеточный зародыш. Кратное увеличение числа хромосом в эндосперме повышает интенсивность обмена веществ и ускоряет рост клеток эндосперма.
Спорогенез — процесс образования спор из спорогенной ткани. Обычно спорогенез предваряется редукционным делением клеток.
Споры образуются путем митоза или мейоза в обычных вегетативных клетках материнского организма или специальных органах - спорангиях и представляют собой микроскопические одноклеточные образования.
При любой форме бесполого размножения - частями тела или спорками - наблюдается увеличение численности особей данного вида без повышения их генетического разнообразия: все особи являются точной копией материнского организма. Новые признаки у таких организмов появляются только в результате мутаций.
Половое размножение существенно отличается от бесполого тем, что в данном случае генотип потомков возникает в результате перекомбинации генов, принадлежащих обоим родителям. Это повышает возможности организмов в приспособлении к меняющимся условиям среды.
Половое размножение характеризуется наличием полового процесса, одним из важнейших этапов которого является слияние половых клеток, или гамет.Гаметы различаются по строению и физиологическим свойствам и делятся на мужские (подвижные - сперматозоиды, неподвижные - спермин) и женские (яйцеклетки).
Образование половых клеток (гаметогенез) у водорослей, многих грибов и высших споровых растений происходит путем митоза или мейоза в специальных органах полового размножения: яйцеклеток - в оогониях или архегониях, сперматозоидов и спермиев - в антеридиях.
Первичные половые клетки делятся путем митоза (период размножения), в результате чего их количество постоянно возрастает. В период роста деление клеток прекращается, и они начинают усиленно расти. При этом будущие яйцеклетки (ооциты) увеличиваются в размерах иногда в сотни и даже в тысячи раз за счет накопления в их цитоплазме запасных питательных веществ в виде желтка. Размеры незрелых мужских гамет (сперматоцитов) увеличиваются незначительно. Затем происходит их мейотическое деление, что приводит к образованию четырех гаплоидных клеток. При сперматогенезе все четыре клетки в дальнейшем превращаются в сперматозоиды.
Вопрос 15. Первый и второй законы Менделя. Гипотеза чистоты гамет.
Закон единообразия гибридов первого поколения: при скрещивании организмов, принадлежащих к разным чистым линиям, все потомство получается одинаковым.
Закон расщепления: при скрещивании гибридов первого поколения между собой в потомстве наблюдается расщепление: примерно ¼ потомков будет иметь рецессивный признак, а ¾ - доминантный.
Гипотеза чистоты гамет. Находящиеся в каждом организме пары альтернативных генов не смешиваются при образовании гамет и по одному переходят в них в чистом виде.
Вопрос 16. Закон независимого комбинирования признаков.
Третий закон Менделя (закон независимого наследования): расщепление по каждой паре признаков происходит независимо от других пар признаков.
Закон выполняется только в том случае, если гены, отвечающие за разные признаки, находятся в негомологичных хромосомах. В основе независимого наследования лежит случайное расхождение хромосом при мейозе.
Вопрос 17. Комплементарность
Комплементарность – признак проявляется при наличии доминантных аллелей в обеих аллельных парах.
Комплементарное взаимодействие бывает двух видов:
КВ обусловлено доминантными генами из разных пар хромосом в каждой из которых не имеет самостоятельного фенотипического проявления.
А – белый; В – белый; А_В_ - пурпурный.
2) КВ обусловлено доминантными генами каждый из которых имеет самостоятельное фенотипическое проявление.
А – светлый; В – голубой; А_В_ - серый; аавв – белый.
Расщепление: 9:3:3:1, 9:3:4, 9:6:1, 9:7.
studfiles.net
Оплодотворение | Биология
Оплодотворение представляет собой процесс слияния женской и мужской гамет – клеток с гаплоидным набором хромосом [94]. Оплодотворенная яйцеклетка носит название зиготы [94]. Зигота диплоидна, так как она образовалась в результате слияния двух гаплоидных гамет. Рис. 94. Последовательные стадии оплодотворения и начала дробления яйцеклетки у животныхВ ядре зиготы все хромосомы вновь становятся парными; в каждой паре гомологичных хромосом одна хромосома отцовская, другая – материнская. Следовательно, диплоидный набор хромосом, характерный для соматических клеток каждого вида организмов, восстанавливается при оплодотворении.
Рис. 94. Последовательные стадии оплодотворения и начала дробления яйцеклетки у животныхВ ядре зиготы все хромосомы вновь становятся парными; в каждой паре гомологичных хромосом одна хромосома отцовская, другая – материнская. Следовательно, диплоидный набор хромосом, характерный для соматических клеток каждого вида организмов, восстанавливается при оплодотворении.
Вскоре после оплодотворения происходит синтез ДНК, хромосомы удваиваются и наступает первое деление ядра зиготы, которое осуществляется путем митоза и представляет собой начало развития зародыша – нового организма.
Развитие женских и мужских гамет и оплодотворение у растений происходит так же, как и у животных, – в женских и мужских половых органах.
Рассмотрим процесс оплодотворения и развития семени у покрытосеменных (цветковых) растений (Вспомните строение и функции цветка из курса ботаники.) В цветке образуются споры (бесполое размножение), возникают гаметы (половой процесс), происходит оплодотворение, в результате которого формируются семя и плод.
У покрытосеменных растений мужские гаметы малоподвижны и называются спермиями [95]. Яйцеклетка неподвижна, и образование ее происходит в зародышевом мешке [95], расположенном в семяпочке. Зародышевый мешок содержит кроме гаплоидной яйцеклетки одну диплоидную клетку, которая участвует в оплодотворении и лежит в центре зародышевого мешка, и несколько других гаплоидных клеток. Рис. 95. Двойное оплодотворение у покрытосеменных растенийСпермии развиваются в пыльцевых зернах пыльников (на тычинках). С помощью пыльцевой трубки спермии переносятся к зародышевому мешку, где происходит процесс оплодотворения [95]. В пыльцевой трубке находятся два спермия. Когда пыльцевая трубка входит в зародышевый мешок, один спермий сливается с яйцеклеткой, образуя диплоидную зиготу, из которой развивается зародыш. Второй спермий сливается с диплоидной центральной клеткой, и в результате возникает новая клетка с триплоидным ядром, т. е. в нем содержится три набора хромосом. Из нее развивается эндосперм семени [95]. Этот универсальный для всех покрытосеменных половой процесс получил название двойного оплодотворения. Он был открыт в 1898 г. С. Г. Навашиным.
Рис. 95. Двойное оплодотворение у покрытосеменных растенийСпермии развиваются в пыльцевых зернах пыльников (на тычинках). С помощью пыльцевой трубки спермии переносятся к зародышевому мешку, где происходит процесс оплодотворения [95]. В пыльцевой трубке находятся два спермия. Когда пыльцевая трубка входит в зародышевый мешок, один спермий сливается с яйцеклеткой, образуя диплоидную зиготу, из которой развивается зародыш. Второй спермий сливается с диплоидной центральной клеткой, и в результате возникает новая клетка с триплоидным ядром, т. е. в нем содержится три набора хромосом. Из нее развивается эндосперм семени [95]. Этот универсальный для всех покрытосеменных половой процесс получил название двойного оплодотворения. Он был открыт в 1898 г. С. Г. Навашиным.
Биологическое значение двойного оплодотворения у покрытосеменных растений заключается в том, что развитие эндосперма начинается только после того, как произойдет оплодотворение яйцеклетки. Триплоидный эндосперм покрытосеменных представляет собой запасной питательный материал для развивающегося зародыша. Кроме того, он включает наследственные задатки материнского и отцовского организмов.
Партеногенез – развитие организма из неоплодотворенного яйца. Партеногенез встречается в природе у многих видов растений и животных. Например, среди растений он известен у одуванчиков, ястребином. Среди животных партеногенез широко распространен у коловраток, солоноватоводных рачков артемий, у ветвистоусых пресноводных рачков дафний, тлей, пчел. В составе пчелиной семьи путем партеногенеза развиваются трутни (самцы).
В чем сущность двойного оплодотворения цветковых растений?
blgy.ru