
Четвертичный период (антропоген). Неогеновый период растения
Неогеновый период (неоген)
Неогеновый период является вторым по счету периодом кайнозойской эры. Неоген берет свое начало 23,3 млн. л. н., длится 20 млн. лет, и заканчивается на рубеже текущего четвертичного периода 2,5 млн. л. н. Во время неогенового периода навсегда перестал существовать древний океан Тетис.
Подразделы неогенового периода, его география и климатические особенности
В соответствии с решением Международного союза геологических наук от 2016 г принято подразделять неоген на два отдела – Миоцен, включающий в себя Аквитанский, Бурдигальский, Лангский, Серравальский, Тортонский и Мессинский ярусы, и Плиоцен, подразделяющийся на Занклский и Пьяченцский ярусы.
| Неогеновый период (неоген) | Отделы | Ярусы |
| Миоцен | Аквитанский | |
| Бурдигальский | ||
| Лангский | ||
| Серравальский | ||
| Тортонский | ||
| Мессинский | ||
| Плиоцен | Занклский | |
| Пьяченцский |
В течение неогена американские материки еще дальше отошли от формирующегося евразийского континента, в северо-восточной части Африки произошел ряд разломов, в результате чего появились Красное море и Аравийский полуостров. Образовались и приняли современные очертания такие горные цепи и массивы, как Альпийско-Гималайские, Кордильеры и Анды. Двигавшийся все дальше по направлению к нынешней Европе африканский материк стал причиной появления пиренейских, альпийских, крымских и карпатских горных массивов, горных систем Ирана и Турции. Соединившийся еще в Палеогене с ближневосточной частью будущего Евроазиатского материка дрейфующий континент Индостан продолжал двигаться в северном направлении, в связи с чем обусловил рост Гималайской горной цепи.
Продолжавший все дальше отдаляться от Африканского континента южноамериканский материк, наконец, в неогене натолкнулся на древнюю океаническую тихоокеанскую плиту Наска, что стало причиной образования нынешней горной цепи Анд. В связи с этим горнопреобразовательные процессы в данных регионах продолжаются и по сей день. Также поныне продолжаются начавшиеся в неогене тектонические процессы в зоне восточноазиатского пояса. Здесь идет попеременное поднятие, опускание и изменение островных дуг, сопровождающееся сильной сейсмической активностью и извержениями вулканов. Идет ускоренный процесс накопления толщ обломочных материалов. Именно в эпоху неогена был сформирован и Байкальский грабен, продолжением изгиба которого является нынешнее озеро Байкал.
В результате дальнейшего продвижения Африканского континента на север древнейший океан Тетис разделился на два огромных морских бассейна. И, если южный бассейн, располагавшийся на территории современного Ближнего Востока, имел связь с основным Мировым океаном, то северный (его принято называть Паратетисом) находился в полной изоляции, вследствие чего соленость его с течением времени все больше повышалась. К концу неогенового периода, в результате начавшихся глобальных горообразовательных процессов, Паратетис распался на ряд еще более мелких бассейнов, в результате которых и образовались будущие Черное и Каспийское моря, а также Средиземное море.
В результате того, что основной котлован Средиземного моря был изолирован от внешних океанов, вследствие отсутствия водных притоков около 5 млн. л. н. данный бассейн бывшего Паратетиса почти полностью испарился. Таким образом, на месте нынешнего Средиземного моря во время неогена существовала пересыхающая гигантская «ванна», то есть низменность, заполненная водой лишь частично, и опускающаяся на многие сотни метров относительно уровня Мирового океана.
В какой-то промежуток времени в одном из мест Гибралтарский хребет, соединяющий Африку с Европой и отделяющий воды Атлантики от Средиземноморской впадины, был нарушен, вследствие чего средиземноморский бассейн вновь начал заполняться водой и через несколько десятилетий воды Средиземноморья и Атлантики полностью сравнялись.
Климат неогена
Климат неогена был холоднее по сравнению с предыдущим палеогеном. Случившееся на границе палеогена и неогена всемирное похолодание обусловило возникновение стабильных ледовых поясных щитов, и повсеместный климат стал еще более континентальным. На севере Европейского континента, как и на обширных областях Сибири, проходила полоса умеренного климата, южнее сменяющаяся на субтропики и тропики, но все же имеющая ярко выраженные сезонные изменения температур, из чего следует, что для данных районов были свойственны более или менее суровые зимние времена года, в зависимости от их близкого расположения к экватору. На территории Гренландии уже в то время климат стал арктическим.
На протяжении всего неогена климат становился более суровым, его континентальность стала выражена еще резче, но при всем том он был еще намного теплее, чем сегодняшний, когда, наконец, случившееся в плиоцене (4,5 млн. л. н.) резкое похолодание не привело его на уровень, близкий к настоящему. На рубеже плиоцена и плейстоцена ледовым панцирем были покрыты с юга – большая часть Антарктиды, севера – обширные площади северного моря, включая Гренландию, Патагонию, Исландию, Скандинавию и северные части Сибири.
Осадконакопление неогенового периода
Накопления осадочных пород неогена имеют место по всей территории нынешних континентов, равно как и по дну Мирового океана. Поскольку горнообразовательная и прочая геологическая активность в данном периоде была чрезвычайно распространена по всем континентам, основными отложениями данного времени являются вулканические породы и продукты прочей тектонической деятельности. Также в черте материков преобладали молассовые и песчано-глинистые формации.
Наибольшая по сравнению с предыдущими периодами седиментация в океанах обусловила различные по толщине на территориях экватора и южных и северных широт кремниевые и карбонатовые накопления. Накопление кремния в период неогена шло даже более мощно, чем в меловом периоде, и обусловлено было необычайным рассветом диатомных водорослей. Ближе к континентальным областям имели место терригенные отложения.
Животные неогена
В морях и океанах неогена процветали простейшие фораминиферы и различные радиолярии. Многочисленны были двустворчатые и брюхоногие моллюски, необычайно размножились различные ракообразные, такие, к примеру, как остракоды. Разнообразны были всевозможные мшанки и иглокожие. В крайних северных и южных территориях в связи с похолоданием исчезли, а в центральных областях еще более и по-новому расцвели кораллы, наибольшая часть которых принадлежала к шестилучевым. Все более множились разновидности хордовых – костистых и хрящевых рыб, а также все более росло количество морских китоподобных, дельфиноподобных и тюленеподобных млекопитающих.
Чрезвычайно разнообразна была фауна наземных млекопитающих неогенового периода. В миоцене, когда местами еще сохранялась структура ландшафта палеогена, на большинстве континентов получила развитие анхитериевая фауна. Характерным представителем данной фауны был анхитерий, небольшое млекопитающее по размерам и строению наиболее близкое к нынешним пони. Это и был древний предок современных лошадей. Животные неогена (рис. 1) имели трехпалые конечности, а разнообразие видов анхитериевой фауны поражало. В нее входили предки, как уже было сказано, лошадей, медведей, носорогов, свиней, антилоп, оленей, грызунов, черепах, приматов и пр. Это были как представители лесной фауны, так и жители степей, саванн и редколесий. Они были экологически неоднородны в соответствии с климатическими условиями, в которых вынуждены были обитать, так, к примеру, для более жарких климатических зон более характерными были обезьяны, газели, антилопы, мастодонты и др. Когда в более суровых северных широтах более распространены были разновидности, успевшие обзавестись обильным шерстяным покровом.
Ближе к середине неогена на обширных территориях евроазиатского материка, Северной Америки и африканского континента стала отчетливо выделяться быстропрогрессирующая гиппарионовая фауна. Состояла она из первых древних лошадей, носорогов, хоботных, грызунов, бегемотов, жирафов, оленей, черепах, верблюдов, саблезубых тигров, гиен, первых человекоподобных обезьян и прочих хищников.
Главными представителями той фауны по праву считаются гиппарионы, лошади небольшого размера, все еще имеющие трехпалые конечности, пришедшие на смену анхитериям, и обитающие на открытых степных и саванных просторах. В силу строения своих конечностей данные животные неогена превосходно передвигались как в высоких степных травах, так и по болотным кочкам.

Рис. 1 – Животные и растения неогена
Преобладающими в гиппарионовой фауне были разновидности, обитающие именно на территориях степных, лесостепных и прочих открытых ландшафтов. К концу неогена гиппарионовая фауна почти повсеместно заменила анхитериевую. Состав ее еще больше расширился за счет увеличения численности поголовья таких разновидностей саванно-редколесных древних животных, как антилопы, различные страусиные, верблюдоподобные, жирафы, однопалые лошади.
Поскольку еще в палеогене связь между различными материками нарушилась, в связи с чем представители фауны более не могли мигрировать с континента на континент. Это стало причиной проявления разнородных провинциальных различий. К примеру, южноамериканский континент обильно был заселен различными копытными, грызунами и плосконосыми приматами сумчатого типа. Данная эндемичная фауна была характерна и для австралийского континента.
Растения неогена
В период неогена под воздействием более суровых климатических факторов возникли первые таежные, лесостепные, равнинно-степные и горные растительные ландшафты.
В экваториальных зонах главенствовали влаголюбивые растения. Эти растения неогенового периода состояли из банановых, фикусных, пальмовых, бамбуковых, лавровых, древесно-папоротниковых, вечнозеленых дубов и пр. Ближе к северным и южным широтам леса, вследствие дефицита осадков и сезонных изменений климата, сменялись саваннами.
Ближе к умеренным широтам начинали преобладать широколиственные леса, состоящие из вечнозеленых древесных форм. С приходом засушливого климата неогена тут широко начала формироваться растительность среднеземноморского типа, что было выражено появлением среди общей массы лавровых вечнозеленых лесов таких разновидностей древовидных растений, как самшиты, платаны, различные ореховые, кипарисовые, маслиновые, а также южных разновидностей сосновых и кедровых.
Также немаловажную роль в распределении растительных видов в неогеновый период имел рельеф местности. В предгорных участках имели место тисовые, таксодиумные, папоротниковые. Выше склоны гор покрывали широколиственные леса с ярко выраженным субтропическим уклоном, далее склоны покрывались хвойными лесами, состоящими из сосен, пихт и елей, в дальнейшем сменяющихся редкими тундровыми кустарниками и травянистой порослью, далее переходящей в ледники и круглогодичные снежные покровы. Данный тип растительности называется высокогорной поясностью.
Ближе к полярным областям в лесах исчезают вечнозеленые широколиственные формы. В лесных массивах более начинают преобладать такие голо- и покрытосеменные растения, как секвойя, сосна, ель, ива, ольха, бук береза, клен, различные ореховые и каштановые. Для аридных областей умеренных широт были характерны широкие саванные и степные пространства. Леса здесь больше тяготеют к речным и приморским бассейнам.
Вследствие резкого похолодания, пришедшего в конце неогена на смену всеобщему короткому и незначительному потеплению, стали ярко выражены разделения на такие зональные типы ландшафтов, как таежные, лесостепные и тундровые. На рубеже неогена и четвертичного периода обширные области суши были заняты таежными лесами, как предполагают ученые, в результате похолодания спустившимися с зон высокогорной поясности и расселившимися по огромным территориям бывших таежных степей.
Ближе к четвертичному периоду обширные равнинные местности умеренных поясов преобразились в степные. Все меньше становилось пролесков, и все больше территории равнин покрывались буйной травянистой степной растительностью. В аридных поясах саванны и редколесья сменились засушливыми пустынями и полупустынями.
Полезные ископаемые неогенового периода
Одним из значительных полезных ископаемых неогенового периода является нефть. К неогену относятся нефтяные месторождения Кавказа, Сахалина, Туркмении и Азербайджана, а также румынские, иракские, иранские, аравийские, индонезийские, калифорнийские, мексиканские, колумбийские, аргентинские и прочие нефтяные бассейны.
Многочисленны также повсеместные месторождения горючих газов, бурого угля, гипса, каменной поваренной и калийной солей. В результате интрузии магматических пород в хребты многих горных массивов оказались впаяны медные, мышьяковые, цинковые, свинцовые, молибденовые, сурьмяные, ртутные, висмутовые, вольфрамовые и прочие руды. Также к неогеновым можно отнести месторождения железных руд керченского полуострова и месторождения бокситов в зоне тропического пояса на островах Гвинея, Гана, Суринам, Гайана, Ямайка и пр.
vse-lekcii.ru
Неоген — wiki.web.ru
Неогеновая система (неогеновый период), неоген (от греч. nйos - новый и gйnos - рождение, возраст), - вторая система кайнозойской эратемы, соответствующая второму периоду кайнозойской эры истории Земли; в стратиграфической шкале следует за палеогеновой системой и предшествует четвертичной системе. Начало неогенового периода, по уточнённым данным, определяется радиометрическим методом в 23,5 млн. лет, а конец - в 0,7 млн. лет (по схеме, принятой Межведомственным стратиграфическим комитетом Академии Наук CCCP) или же в 1,8 млн. лет (по решению Международного геологического конгресса в 1948) от современности. Общая продолжительность периода 22 и 23 млн. лет соответственно. Название "неогеновая система" было предложено в 1853 австрийским геологом М. Гёрнесом для выделенных в 1833 английским геологом Ч. Лайелем двух отделов - миоцена и плиоцена.
Подразделения неогеновой системы
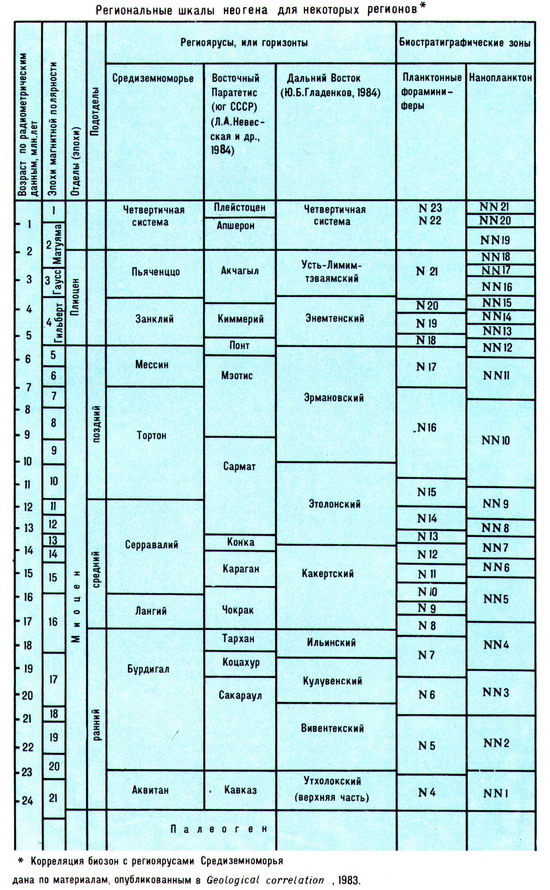
Неоген подразделяется на два отдела: нижний - миоцен и верхний - плиоцен. В дальнейшем эти отделы были разделены на подотделы и ряд ярусов, установленных в области Средиземноморья и некоторое время считавшихся ярусами международной шкалы. Однако после проведённой в 1970-х гг. ревизии ярусы средиземноморской шкалы стали рассматриваться только как региональные. В 1975 на конгрессе Регионального комитета по стратиграфии средиземноморского неогена были приняты три равноценные региоярусные шкалы для Средиземноморья, Западного и Восточного Паратетиса. Одновременно при изучении океанических осадков были разработаны зональные шкалы по планктонным фораминиферам и по нанопланктону, которые используются при корреляции неогена океанических районов, а также при сопоставлении региоярусных шкал отдельных регионов. Таким образом, для неогеновой системы не существует общепринятой международной ярусной шкалы. Региональные ярусы и биозоны, используемые в отдельных регионах, распределяются в пределах отделов и подотделов.
Общая характеристика
Неогеновые отложения широко распространены под покровом четвертичных отложений на всех континентах и на дне океанов. Неогеновый период был одним из наиболее геократических этапов в развитии Земли, особенно его 2-я половина - плиоцен. К концу плиоцена сформировались основные черты современного рельефа и гидросети, завершилось образование многочисленных горных систем - Альп, Карпат, Балкан, Апеннин, Крыма, Кавказа, Гималаев, Кордильер Северной и Южной Америки, островных дуг - Алеутской, Корякско-Камчатской, Японской и др. Усиление поднятий привело к образованию многочисленных внутренних впадин и глубоководных внутренних и окраинных бассейнов. Быстрый рост горных систем сопровождался складко- и шарьяже-образованием и сильной вулканической деятельностью. Для неогенового периода в целом было характерно значительное похолодание климата и образование ледниковых покровов Антарктиды и Гренландии. Снижение температуры привело к резкой дифференциации климата и соответственно ландшафтных зон. На этом общем фоне ухудшения климата наблюдались и отдельные этапы потеплений. После холодного периода 1-й половины раннего миоцена наступило значительное потепление, известное как климатический оптимум 2-й половины раннего - начала среднего миоцена. В это время в высоких широтах появились термофильные элементы среди древесных растений, пресноводных моллюсков и наземных млекопитающих на суше и ряд теплолюбивых форм среди моллюсков, фораминифер и других групп беспозвоночных - в морях.
Начиная со 2-й половины среднего миоцена вновь наступило похолодание и началась аридизация климата, продолжавшаяся в позднем миоцене. Это привело к сокращению лесов и развитию лесостепных и степных пространств. В Антарктиде возникло покровное оледенение. В плиоцене продолжалось похолодание, на фоне которого происходили неоднократные колебания температурного режима. В 1-й половине плиоцена появились покровные ледники Северного полушария. В течение неогенового периода распределение океанов, морей и суши постепенно приближалось к современному, что сопровождалось глобальным (гляцио-эвстатическим) понижением уровня океана с отдельными колебаниями, соответствовавшими масштабам оледенения. В начале миоцена крупное континентальное море - Паратетис, образовавшееся в олигоцене на северной окраине океана Тетис, потеряло связь с бореальными морями, сохранив связь с Тетисом, которая неоднократно прерывалась в миоцене. В середине миоцена Тетис окончательно распался и Средиземное море отделилось от Индо-Пацифики. В позднем миоцене Средиземное море отчленилось от океана в связи с глобальным падением уровня океана, в нём образовались мощные толщи эвапоритов (мессинский кризис солёности). В плиоцене Средиземное море вновь соединилось с Атлантикой, а Паратетис распался, и на его месте сформировались Азово-Черноморский и Каспийский бассейны. Трансгрессии морских бассейнов, захватывавшие окраинные области континентов, отмечались главным образом в 1-й половине миоцена, а в плиоцене морей с нормальной солёностью в пределах современных континентов практически не было. В целом трансгрессии неогеновых морей и осадконакопление на континентах происходили на фоне сложной и быстро меняющейся тектонической и палеогеографической обстановки, что обусловило пестроту фациального состава неогеновых отложений этих областей. Наиболее распространены были континентальные песчано-глинистые и молассовые формации; среди морских отложений основную роль играли песчано-глинистые и карбонатно-обломочные; значительное распространение имели и наземно-вулканогенные формации. Морские карбонатные отложения и эвапориты были относительно мало развиты. Биогенная седиментация в океане стала более интенсивной, чем прежде; оформились три пояса, установленные для современного океана: экваториальный, северный и южный пояса кремненакопления и карбонатонакопления. Кремненакопление в неогене (более мощное, чем в меловое и палеогеновое время) происходило в основном за счёт диатомовых водорослей. В приконтинентальных районах преобладали терригенные отложения.
Растительный и животный мир
В растительном мире неогенового периода основную роль играли те же группы, что и в современную эпоху. На суше господствовали высшие растения, а среди них покрытосеменные и голосеменные, в меньшей степени были распространены папоротники, мохообразные и пр. В умеренных широтах палеогеновые термофильные элементы отмечались только в 1-й половине миоцена, несколько расширив свой ареал в период климатического оптимума конца раннего - начала среднего миоцена. Основной компонент лесной растительности - листопадные формы. В связи с общим похолоданием и аридизацией климата проявилась широтная зональность, оформились все существующие ныне растительные зоны и флористическая область.
Растительность морей и других водоёмов была представлена различными водорослями (диатомовыми, золотистыми, багряными, пиррофитовыми и пр.) и некоторыми высшими растениями.
В Евразии в начале миоцена состав млекопитающих ещё близок к олигоценовому и только во 2-й половине раннего миоцена появились формы миоценового типа - хоботные (гомфотерии, зиголофодоны) и др. В среднем миоцене наряду с хоботными (мастодонты) и лошадиными (анхитерии) распространились бовиды и другие обитатели полуоткрытых и сухих пространств. В позднем миоцене сформировались сообщества степных травянистых равнин - т.н. гиппарионовая фауна (гиппарионы и разнообразные бовиды - газели, палеориксы и др.), существовавшая до конца миоцена - начала плиоцена. В плиоцене на территории Евразии несколько раз происходила смена фаун млекопитающих. На границе плиоцена и плейстоцена (0,7 млн. лет) появились холодолюбивые и перигляциальные формы - мамонт, овцебык, сайга и др. Фауна неогеновых млекопитающих Африки была близка к евразиатской. Млекопитающие Австралии были представлены отрядами сумчатых и однопроходных. В Южной Америке в миоцене основную роль играли сумчатые, неполнозубые (муравьеды, ленивцы, броненосцы), грызуны и некоторые эндемичные копытные. В позднем миоцене произошёл обмен фаунами Северной Америки с Евразией, а в плиоцене отмечена миграция млекопитающих (хищники, лошади и др.) из Северной Америки в Южную.
В неогеновых океанах и морях были распространены представители простейших (фораминиферы, в меньшей степени - радиолярии), брюхоногие и двустворчатые моллюски, ракообразные (остракоды и другие группы), мшанки, иглокожие (морские ежи), кишечнополостные (шестилучевые кораллы), а из хордовых - костистые и хрящевые рыбы и водные млекопитающие.
Полезные ископаемые
С неогеновыми отложениями связаны многочисленные полезные ископаемые. Из осадочных полезных ископаемых наиболее важны месторождения нефти и газа в предгорных и межгорных прогибах Ближнего и Среднего Востока, Калифорнии, Аляски, Японии и т.д. В бывшем CCCP к нефтегазоносным регионам, отвечающим крупным предгорным прогибам, относятся Прикарпатский, Азово-Кубанский, Терско-Каспийский; межгорным впадинам - Закарпатский, Восточно-Черноморский, Южно-Каспийский и Ферганский; внутрискладчатым депрессиям - Сахалино-Охотский, Южно-Охотский и Анадырский. Достаточно многочисленны в неогеновых отложениях месторождения бурых углей и лигнитов, несколько реже отмечаются и каменные угли. На территории Pоссии разрабатываются месторождения Дальневосточной, Колымской, Центрально-Якутской, Амурской, Прибайкальской угленосных провинций и областей, Южно-Уральского буроугольного бассейна и др. На территории зарубежной Европы известны буроугольные месторождения в Германии, Болгарии, Венгрии, Румынии, Чехии, Югославии и др. В Азии наиболее крупные залежи угля связаны с Анатолийским буроугольным бассейном, известны месторождения угля также в Индии, Китае, Юго-восточной Азии. Значительными запасами обладают угольные месторождения Северной и Южной Америки и Австралии (бассейны Латроб-Валли и др.). Отмечены месторождения серы, связанные главным образом с эвапоритовыми формациями (Предкарпатье, Апеннинский полуостров, Сицилия), а также залежи солей (Предкарпатье, Закарпатье, Закавказье, Средняя Азия и др.). Образовались россыпные месторождения титана, олова, ильменита, рутила, циркона и др., многие бокситовые месторождения тропического пояса (Ямайка, Гайана, Суринам, Гана, Гвинея). Значительны также месторождения бентонитовых и палыгорскитовых глин, алунитов, перлитов, каолинитов, галлуазита. В качестве строительных материалов, керамического и цементного сырья используются известняки, кварцевые пески, песчаники, диатомиты, глины.
С интрузивными и эффузивными породами связаны многочисленные и разнообразные месторождения руд ртути, олова, свинца, цинка, сурьмы и других элементов, образующие местами рудные пояса (полиметаллический пояс Перу, золотоносный пояс Эквадора, оловоносный и меденосные пояса Боливии, медно-серебряные месторождения Центральной Америки, медистые песчаники Таджикской депрессии, полиметаллические месторождения и ртутные рудные провинции Альпийского складчатого пояса, в т.ч. Закарпатья). В северо-западной и западной частях Тихоокеанского побережья, в т.ч. на Корякском нагорье, Камчатке, Курильских островах, Сахалине, распространены касситерит-сульфидные, колчеданные, ртутные, сурьмяные рудные провинции, месторождения руд свинца, цинка, меди, золота, серебра, молибдена, мышьяка, хрома и других металлов. Области интенсивного вулканизма и магматизма перспективны для поисков руд цветных металлов. В океанических областях песчано-глинистые шельфовые отложения во многих районах нефтегазоносны: Мексиканский залив, Карибское море, Гвинейский залив, Средиземное, Красное, Берингово моря, залив Аляска, Японское и Южно-Китайское моря, шельфы Чили, Перу, Эквадора, Калифорнии. На северо-восточном шельфе Австралии продуктивная толща приурочена к миоценовым рифам. Большие перспективы связаны с железомарганцевыми конкрециями, содержащими многие ценные металлы (никель, медь, кобальт и др.), и фосфоритами. Фосфориты и фосфатизированные осадки и породы распространены как на шельфах и в верхних частях континентальных склонов, так и в пелагических областях, преимущественно на подводных горах. Часть обогащенных фосфоритами пород имеет миоценовый возраст - шельфы северо-западной, юго-западной и южной Африки, восточных и западных берегов Северной Америки, Новой Зеландии, подводные горы Тихого, Индийского и Атлантического океанов.
Литература
- Горная энциклопедия, в 5 т. М., изд-во "Советская энциклопедия", 1987, гл. ред. Е.А. Козловский
wiki.web.ru
Мир дикой природы на wwlife.ru
Оглавление |
|
1. |
|
1.1. |
|
2. |
|
2.1. |
|
2.2. |
|
2.3. |
|
2.4. |
|
2.5. |
|
2.6. |
|
2.7. |
|
2.8. |
|
2.9. |
|
2.10. |
|
2.11. |
|
2.12. |
|
2.12.1. |
|
2.12.2. |
|
2.13. |
|
2.12.2. Животный мир неогенового периода (23,0 - 2,588 млн. лет назад)
Неогеновый период (23,0 - 2,588 млн.лет назад).
 Рис. 2.12.1. НеогенНеогеновый период (в переводе - новорожденный рис. 2.12.1) подразделяется на два отдела миоцен (23,0 - 5,33 млн. лет назад) и плиоцен (5,33 - 2,588 млн. лет назад). Климат неогенового периода был довольно теплым, и влажным, однако несколько более прохладным по сравнению с климатом палеогенового периода. В конце неогена он постепенно приобретает современные черты.
Рис. 2.12.1. НеогенНеогеновый период (в переводе - новорожденный рис. 2.12.1) подразделяется на два отдела миоцен (23,0 - 5,33 млн. лет назад) и плиоцен (5,33 - 2,588 млн. лет назад). Климат неогенового периода был довольно теплым, и влажным, однако несколько более прохладным по сравнению с климатом палеогенового периода. В конце неогена он постепенно приобретает современные черты.
Среди морских беспозвоночных в это время преобладают двустворчатые и брюхоногие моллюски, морские ежи. Мшанки и кораллы на юге Европы образуют рифы. Прослеживаются арктические зоогеографические провинции: северная, включавшая Англию, Нидерланды и Бельгию, южная - Чили, Патагонию и Новую Зеландию.
Сильно распространилась солоноватоводная фауна. Ее представители населяли большие мелководные моря, образовавшиеся на материках в результате наступления неогенового моря. В этой фауне совершенно отсутствуют кораллы, морские ежи и звезды. Моллюски по количеству родов и видов значительно уступают моллюскам, населявшим океан с нормальной соленостью. Однако по численности особей они во много раз превосходят океанских. Раковины небольших по размерам солоноватоводных моллюсков буквально переполняют отложения этих морей. Рыбы уже совершенно не отличаются от современных.
") Рис. 2.12.2. Махайроды (Machairodus)Похожим на современный становится и сухопутный органический мир. Примитивных креодонтов вытесняют медведи, гиены, куницы, собаки, барсуки. Будучи более подвижными и имея более сложную организацию, они приспособились к разнообразным условиям жизни, перехватывали добычу у креодонтов и сумчатых хищников, а иногда и питались ими.
Рис. 2.12.2. Махайроды (Machairodus)Похожим на современный становится и сухопутный органический мир. Примитивных креодонтов вытесняют медведи, гиены, куницы, собаки, барсуки. Будучи более подвижными и имея более сложную организацию, они приспособились к разнообразным условиям жизни, перехватывали добычу у креодонтов и сумчатых хищников, а иногда и питались ими.
Наряду с видами, которые, несколько изменившись, дожили до нашего времени, появились и виды хищников, которые вымерли в неогене. К ним в первую очередь относится саблезубый тигр (рис. 2.12.2).
У потомков палеогеоновых мерикгиппусов - гиппарионов были уже такие зубы, как у современной лошади. Их небольшие боковые копыта не касались земли. Копыта же на средних пальцах делались все более крупными и широкими. Они хорошо удерживали животных на твердой почве, давали им возможность разрывать снег, чтобы извлечь из-под него корм, а так же защищаться от хищников.
") Рис. 2.12.3. Мегатерий (Megatherium)Наряду с североамериканским центром развития лошадей существовал и европейский. Однако в Европе древние лошади вымерли в начале олигоцена, не оставив потомков. Вероятнее всего они были истреблены многочисленными хищниками. В Америке же древние лошади продолжали развиваться. Впоследствии они дали настоящих лошадей, которые через Берингов перешеек проникли в Европу и Азию. В Америке лошади вымерли еще в начале плейстоцена, а крупные табуны современных мустангов, свободно пасущиеся в американских прериях, являются далекими потомками лошадей, привезенных испанскими колонизаторами. Таким образом, произошел своеобразный обмен лошадьми между Новым Светом и Старым Светом.
Рис. 2.12.3. Мегатерий (Megatherium)Наряду с североамериканским центром развития лошадей существовал и европейский. Однако в Европе древние лошади вымерли в начале олигоцена, не оставив потомков. Вероятнее всего они были истреблены многочисленными хищниками. В Америке же древние лошади продолжали развиваться. Впоследствии они дали настоящих лошадей, которые через Берингов перешеек проникли в Европу и Азию. В Америке лошади вымерли еще в начале плейстоцена, а крупные табуны современных мустангов, свободно пасущиеся в американских прериях, являются далекими потомками лошадей, привезенных испанскими колонизаторами. Таким образом, произошел своеобразный обмен лошадьми между Новым Светом и Старым Светом.
В Южной Америке обитали гигантские ленивцы - мегатерии (до 8 м в длину рис. 2.12.3). Становясь на задние лапы, они объедали листья деревьев. Мегатерии имели толстый хвост, низкий череп с небольшим мозгом. Их передние лапы были намного короче задних. Будучи неповоротливыми, они становились легкой добычей для хищников и поэтому полностью вымерли, не оставив потомков.
Изменение климатических условий привело к образованию обширных степей, что благоприятствовало развитию копытных. От маленьких безрогих оленей, живших на болотистой почве, произошли многочисленные парнокопытные - антилопы, козлы, бизоны, бараны, газели, чьи прочные копыта были хорошо приспособлены к быстрому бегу в степях. Когда парнокопытных развелось такое количество, что начала ощущаться нехватка пищи, часть их освоила новые места обитания: скалы, лесостепи, пустыни. От живших в Африке жирафообразных безгорбых верблюдов произошли настоящие верблюды, заселившие пустыни и полупустыни Европы и Азии. Горб с питательными веществами позволял верблюдам длительное время обходиться без воды и пищи.
 Рис. 2.12.4. МегалоцерасЛеса заселяли настоящие олени, из которых одни виды встречаются и в наши дни, а другие, например мегалоцерасы (рис. 2.12.4), которые были в полтора раза крупнее обычных оленей, полностью вымерли.
Рис. 2.12.4. МегалоцерасЛеса заселяли настоящие олени, из которых одни виды встречаются и в наши дни, а другие, например мегалоцерасы (рис. 2.12.4), которые были в полтора раза крупнее обычных оленей, полностью вымерли.
В лесостепных зонах обитали жирафы, вблизи озер и болот - бегемоты, свиньи, тапиры. В густых кустарниковых зарослях жили носороги, муравьеды.
Среди хоботных появляются мастодонты с прямыми длинными клыками и настоящие слоны.
На деревьях живут лемуры, обезьяны, человекообразные обезьяны. Некоторые лемуры перешли к наземному образу жизни. Передвигались они на задних ногах. Достигали 1,5 м в высоту. Питались главным образом плодами и насекомыми.
") Рис. 2.12.5. Динорнис (Dinornis)Жившая в Новой Зеландии гигантская птица динорнис достигала 3,5 м в высоту (рис. 2.12.5). Голова и крылья у динорниса были маленькими, клюв недоразвитым. Он передвигался по земле на длинных сильных ногах. Дожил динорнис до четвертичного периода и, очевидно, был истреблен человеком.
Рис. 2.12.5. Динорнис (Dinornis)Жившая в Новой Зеландии гигантская птица динорнис достигала 3,5 м в высоту (рис. 2.12.5). Голова и крылья у динорниса были маленькими, клюв недоразвитым. Он передвигался по земле на длинных сильных ногах. Дожил динорнис до четвертичного периода и, очевидно, был истреблен человеком.
В неогеновый период появляются дельфины, тюлени, моржи - виды, живущие и в современных условиях.
В начале неогенового периода в Европе и Азии было много хищных животных: собак, саблезубых тигров, гиен. Среди травоядных преобладали мастодонты, олени, однорогие носороги.
В Северной Америке хищные были представлены собаками и саблезубыми тиграми, а травоядные - титанотериями, лошадьми и оленями.
Южная Америка была несколько изолированной от Северной. Представителями ее фауны были сумчатые, мегатерии, ленивцы, броненосцы, широконосые обезьяны.
В верхнемиоценовый период между Северной Америкой и Евразией происходит обмен фауной. Много животных переселилось с материка на материк. Северную Америку заселяют мастодонты, носороги, хищники, а в Европу и Азию переселяются лошади.
С началом лигоцена в Азии, Африке и Европе расселяются безрогие носороги, мастодонты, антилопы, газели, свиньи, тапиры, жирафы, саблезубые тигры, медведи. Однако во второй половине плиоцена климат на Земле сделался прохладным, и такие животные, как мастодонты, тапиры, жирафы, переселяются на юг, и на их месте появляются быки, бизоны, олени, медведи.
В плиоцене связь между Америкой и Азией прервалась. Одновременно возобновилась связь между Северной и Южной Америкой. Североамериканская фауна переселилась в Южную Америку и постепенно вытеснила ее фауну. Из местной фауны остались лишь броненосцы, ленивцы и муравьеды, распространились медведи, ламы, свиньи, олени, собачьи, кошачьи.
Австралия была изолирована от других материков. Следовательно, значительных изменений в фауне там не произошло.[1]
2.12.1. Животный мир миоценовой эпохи (25 - 5 млн. лет назад)
В миоцене, с началом закрытия восточного Тетиса, изменения уровня моря определяли возможность интенсивных межконтинентальных фаунистических обменов между Евразией, Африкой, Индийским субконтинентом и восточносредиземноморской областью. В пределах Евразийской зоогеографической области сформировалась единая Европейско-Сибирская подобласть с южной границей в Азии приблизительно по 40oсеверной широты, ставшая гигантской ареной эволюции многих групп млекопитающих в миоцене.
") Рис. 2.12.1.1. Анхитерий (Anchitherium)В конце раннего миоцена, впервые после длительного периода взаимной изоляции, происходил значительный обмен между Африкой и Евразией. Главным эффектом этого обмена было изменение африканской фауны: в ее составе появились кошачьи, собачьи, куньи, носороги, халикотерии, свиньи. Вероятно, в это же время из Африки на Индийский субконтинент эмигрировали хоботные. Первые мастодонты вселились в Европу и Северную Азию в раннем миоцене (MN3). В это же время в Евразию из Северной Америки иммигрировали анхитерии (рис. 2.12.1.1). Наиболее значительным событием в истории раннемиоценовых грызунов Европы была иммиграция азиатских по происхождению хомяков современного облика (Eumyarion, Democricetodon, Fahlbuschia, Megacricetodon, Cricetodon (рис. 2.12.1.2), Lartetomys) в среднем орлеании. С их появлением эомиидно-глиридная ассоциация трансформировалась в крицетидно-глиридную, причем во многих комплексах доминирующей группой были именно Cricetidae. В таком виде европейская фауна грызунов просуществовала вплоть до раннего валлезия (MN9). В среднем орлеании в Африку из Евразии проникли первые полорогие и жирафовые.
Рис. 2.12.1.1. Анхитерий (Anchitherium)В конце раннего миоцена, впервые после длительного периода взаимной изоляции, происходил значительный обмен между Африкой и Евразией. Главным эффектом этого обмена было изменение африканской фауны: в ее составе появились кошачьи, собачьи, куньи, носороги, халикотерии, свиньи. Вероятно, в это же время из Африки на Индийский субконтинент эмигрировали хоботные. Первые мастодонты вселились в Европу и Северную Азию в раннем миоцене (MN3). В это же время в Евразию из Северной Америки иммигрировали анхитерии (рис. 2.12.1.1). Наиболее значительным событием в истории раннемиоценовых грызунов Европы была иммиграция азиатских по происхождению хомяков современного облика (Eumyarion, Democricetodon, Fahlbuschia, Megacricetodon, Cricetodon (рис. 2.12.1.2), Lartetomys) в среднем орлеании. С их появлением эомиидно-глиридная ассоциация трансформировалась в крицетидно-глиридную, причем во многих комплексах доминирующей группой были именно Cricetidae. В таком виде европейская фауна грызунов просуществовала вплоть до раннего валлезия (MN9). В среднем орлеании в Африку из Евразии проникли первые полорогие и жирафовые.
 Рис. 2.12.1.2. Cricetodon soriaeМиоцен – время вымирания креодонтов. Однако в Азии в среднем миоцене еще присутствуют 2 рода: самый крупный представитель Proviverrinae - Dissopsalis (размером с шакала) и очень крупный представитель Hyaenodontinae - Hyainailouros (размером с тигра). Интересно отметить, что эти оба рода общие для Азии и Африки, а Hyainailouros присутствует еще и в Европе. Это подтверждает обмен фауны креодонтов между Азией и Африкой через Европу в олигоцене. Вымирание креодонтов совпадает с адаптивной радиациейCarnivora, которые являются конкурентной группой. Вымиранию креодонтов, вероятно, способствовали этологические особенности группы. Морфофункциональный анализ мозга Neohyaenodon horridus (Hyaenodontini) показал сильное морфологическое развитие вентро-каудальной зоны височной доли мозга (Савельев, Лавров, 2001). Следствием этого является повышение контроля за спонтанной активностью и агрессией. Этот крупный хищник был менее агрессивным, чем современные крупные представители отряда хищных (Carnivora).
Рис. 2.12.1.2. Cricetodon soriaeМиоцен – время вымирания креодонтов. Однако в Азии в среднем миоцене еще присутствуют 2 рода: самый крупный представитель Proviverrinae - Dissopsalis (размером с шакала) и очень крупный представитель Hyaenodontinae - Hyainailouros (размером с тигра). Интересно отметить, что эти оба рода общие для Азии и Африки, а Hyainailouros присутствует еще и в Европе. Это подтверждает обмен фауны креодонтов между Азией и Африкой через Европу в олигоцене. Вымирание креодонтов совпадает с адаптивной радиациейCarnivora, которые являются конкурентной группой. Вымиранию креодонтов, вероятно, способствовали этологические особенности группы. Морфофункциональный анализ мозга Neohyaenodon horridus (Hyaenodontini) показал сильное морфологическое развитие вентро-каудальной зоны височной доли мозга (Савельев, Лавров, 2001). Следствием этого является повышение контроля за спонтанной активностью и агрессией. Этот крупный хищник был менее агрессивным, чем современные крупные представители отряда хищных (Carnivora).
 Рис. 2.12.1.3. Pliocyon преследует верблюдовС конца раннего миоцена уже можно четко обосновать провинциальное биогеографическое подразделение евразиатских и североафриканских териофаун. Для фаун млекопитающих всех европейских провинций в целом было характерно преобладание сообществ закрытых лесных обстановок, явно субтропических в южных районах. Суб-Паратетисная провинция характеризовалась наличием гигантского “экологического коридора” открытых ландшафтов на территории от восточносредиземноморской Европы на западе через Малую Азию, Ближний Восток и северную Аравию до Афганистана на востоке и Китая на северо-востоке. Вероятно, именно через нее осуществлялись основные фаунистические обмены между Евразией и Африкой.
Рис. 2.12.1.3. Pliocyon преследует верблюдовС конца раннего миоцена уже можно четко обосновать провинциальное биогеографическое подразделение евразиатских и североафриканских териофаун. Для фаун млекопитающих всех европейских провинций в целом было характерно преобладание сообществ закрытых лесных обстановок, явно субтропических в южных районах. Суб-Паратетисная провинция характеризовалась наличием гигантского “экологического коридора” открытых ландшафтов на территории от восточносредиземноморской Европы на западе через Малую Азию, Ближний Восток и северную Аравию до Афганистана на востоке и Китая на северо-востоке. Вероятно, именно через нее осуществлялись основные фаунистические обмены между Евразией и Африкой.
Взаимное влияние столкнувшихся в Евразии и Африке в миоцене различных по области происхождения экологических конкурентов вызвало бурную эволюцию ряда групп (ежовых, землеройковых, кротовых, пищуховых, беличьих, тушканчикообразных, хомякообразных и многих других грызунов, антропоидных приматов, носороговых, жвачных, хоботных, куньих, собачьих, гиеновых, кошачьих и др.) и вымирание последних “палеогеновых” реликтов (ежей-амфехинин, корнезубых зайцеобразных, аплодонтовых-просциурин, ктенодактилид-татаромиин, тахиориктоидид, гигантских носорогов, кайнотериев, креодонтов, амфиционид (рис. 2.12.1.3) и др.). К концу миоцена сформировались почти все группировки млекопитающих надродового ранга, а также очень многие из рецентных родов. В добавление к ним существовало множество впоследствии вымерших форм, так что с точки зрения териологии миоцен по праву можно считать эпохой великого разнообразия, в формировании которого одна из ведущих ролей принадлежала территории Азии. [2]
 Рис. 2.12.1. 4. Зубы - большой белой акулы и кархарадонаОдна из крупнейших среди современных акул — большая белая акула, достигающая длины 10м, кажется карликом в сравнении с одним из своих предков, кархародоном мегалодоном. Эта рыба обитала на Земле с миоценовой по плейстоценовую эпоху. И хотя кархародон мегалодон известен нам лишь по ископаемым зубам, ученые все же смогли воссоздать его внешний облик. Взгляните на фотографии двух зубов (рис. 2.12.1.4) изображенных в натуральную величину. Тот, что слева, принадлежит взрослой большой белой акуле. Другой же — ископаемый зуб кархародона. Ученые сопоставили оба зуба и, основываясь на известных им размерах большой белой акулы, прикинули, что длина кархародона от носа до кончика хвоста должна была составлять не менее 20 м (рис. 2.12.1.5).
Рис. 2.12.1. 4. Зубы - большой белой акулы и кархарадонаОдна из крупнейших среди современных акул — большая белая акула, достигающая длины 10м, кажется карликом в сравнении с одним из своих предков, кархародоном мегалодоном. Эта рыба обитала на Земле с миоценовой по плейстоценовую эпоху. И хотя кархародон мегалодон известен нам лишь по ископаемым зубам, ученые все же смогли воссоздать его внешний облик. Взгляните на фотографии двух зубов (рис. 2.12.1.4) изображенных в натуральную величину. Тот, что слева, принадлежит взрослой большой белой акуле. Другой же — ископаемый зуб кархародона. Ученые сопоставили оба зуба и, основываясь на известных им размерах большой белой акулы, прикинули, что длина кархародона от носа до кончика хвоста должна была составлять не менее 20 м (рис. 2.12.1.5).
 Рис. 2.12.1.5. КархарадонРаспространение степей и постепенное исчезновение лесов в миоцене сильно повлияли на рост численности популяций травоядных млекопитающих. Меньше деревьев — меньше энергии, которую они "отдают" животным.
Рис. 2.12.1.5. КархарадонРаспространение степей и постепенное исчезновение лесов в миоцене сильно повлияли на рост численности популяций травоядных млекопитающих. Меньше деревьев — меньше энергии, которую они "отдают" животным.
Представьте себе на минуту обычное дерево. Большую часть своей энергии оно расходует на создание для себя опоры. С этой целью у него развивается специальный твердый ствол и множество ветвей.
В умеренном климате животные за год могут употребить в пищу лишь очень незначительную часть съедобного вещества, вырабатываемого деревом.
На листопадных деревьях (которые ежегодно сбрасывают листву) листья вообще растут всего шесть месяцев в году, а плоды и семена (за исключением орехов) появляются только на несколько недель. Значит, леса умеренного пояса могут за год прокормить лишь ограниченное число животных.
Резкое увеличение количества травы на суше в миоценовую эпоху означало, по сути, возникновение принципиально нового источника пищи. Однако поначалу травоядным млекопитающим было непросто воспользоваться ресурсами этой бездонной кладовой. Некоторые их виды даже вымерли, поскольку не смогли приспособиться к травяному рациону. Млекопитающим, зубы которых были рассчитаны на пережевывание мягкой листвы, оказалось нелегко перейти на такой жесткий и волокнистый корм, как трава. Для таких животных травяная "диета" означала постоянное усиленное жевание, и их зубы быстро изнашивались, создавая им серьезнейшие проблемы. Ведь млекопитающие не обладают неограниченным запасом зубов, а беззубые челюсти означают для них неминуемую голодную смерть.
Несмотря на все это, в миоцене млекопитающие стали гораздо многочисленнее и разнообразнее, среди них возникло множество травоядных. К этому времени "жвачный" желудок превратился в идеальный механизм для переваривания травы. В результате в миоцене произошел своего рода взрыв, породивший новые виды травоядных, способных "жевать жвачку". Жвачные животные могут набивать себе животы громадным количеством пищи, переварить которую можно и позднее. При этом если на жвачное животное нападет хищник, оно может удрать от него, унося с собой запас корма на несколько дней вперед. Оказавшись в безопасности, животное может без лишней спешки заняться перевариванием съеденного.
В ходе этой своеобразной "жвачной революции" резко возросла численность предков нынешних антилоп, буйволов, оленей, жирафов и овец. В Северной Америке обитали антилопообразные вилороги — их причудливые рога росли у них на кончике носа.
 Рис. 2.12.1.6. МеригиппусЧтобы совладать с травяным кормом, прежние зубы травоядных претерпели два основных изменения:Во-первых, узор из бугорков на жевательной поверхности зубов значительно усложнился, и они стали самозатачивающимися. Теперь на зубах, по мере стирания, образовывались ряды прочных эмалевых ребер, которые оставались острыми всю жизнь животного.
Рис. 2.12.1.6. МеригиппусЧтобы совладать с травяным кормом, прежние зубы травоядных претерпели два основных изменения:Во-первых, узор из бугорков на жевательной поверхности зубов значительно усложнился, и они стали самозатачивающимися. Теперь на зубах, по мере стирания, образовывались ряды прочных эмалевых ребер, которые оставались острыми всю жизнь животного.
Во-вторых, у отдельных зубов развились гораздо более широкие коронки (та часть зуба, что выдается над десной) и они сделались крупнее.Эти новшества, как и появление корневых отверстий (которые обеспечивали приток крови к растущей части зуба), способствовали тому, что зубы животных стали расти всю их жизнь. Теперь постоянное трение зубов друг о друга больше уже не приводило к их преждевременному износу.
Необходимость приспособиться к преимущественно травяному корму - не единственная проблема, возникшая перед новыми миоценовыми травоядными. Не менее серьезные проблемы были связаны с жизнью на открытых равнинах, где травоядные постоянно находились на виду у хищников.
Чтобы выжить в подобных условиях, животные нуждались в хорошем круговом зрении, дающем широкий или даже 180-градусный обзор (при таком обзоре глаза располагаются по обеим сторонам головы, и животные видят и то, что впереди, и то, что сзади, оставаясь неподвижными). Такое зрение позволяло травоядным вовремя замечать опасность, грозящую с любого направления, что, в свою очередь, улучшало взаимодействие в стаде.
Чтобы стадо имело больше шансов уцелеть на открытой местности, в нем выработались специальные методы несения, "караульной службы" (когда отдельные животные смотрят по диагонали через все стадо), а также более совершенные системы сигналов и обмена информацией внутри стада.
") Рис. 2.12.1.7. Мастадонт (Mammutidae)Длинные ноги также ценное подспорье при жизни на равнине. Они позволяют животному держать голову высоко над землей и тем самым лучше обозревать окрестности. Ну и, разумеется, длинные ноги служат своему хозяину незаменимым средством спасения в случае опасности. Поэтому в течение миоцена конечности травоядных животных постепенно приспособились к быстрому бегу.
Рис. 2.12.1.7. Мастадонт (Mammutidae)Длинные ноги также ценное подспорье при жизни на равнине. Они позволяют животному держать голову высоко над землей и тем самым лучше обозревать окрестности. Ну и, разумеется, длинные ноги служат своему хозяину незаменимым средством спасения в случае опасности. Поэтому в течение миоцена конечности травоядных животных постепенно приспособились к быстрому бегу.
Кости нижней части ноги со временем удлинились, а кости верхней, напротив, стали короче. Главные мышцы, ответственные за движение конечностей, также укоротились и располагались теперь выше, ближе к лопаткам и подвздошным костям, где они соединялись с туловищем животного.
При таком строении мускулатуры животное могло совершать длинные прыжки, затрачивая при этом минимум энергии.
Ноги травоядного, обитающего на равнине, весят очень мало и не обладают большой силой. Однако они идеально приспособлены для бегства от хищника и позволяют животному покрывать большие расстояния на высокой скорости, стоит ему только войти в надлежащий ритм бега.
") Рис. 2.12.1.8. Дейнотерий (Deinotherium)На протяжении миоцена лошади продолжали увеличиваться в размерах. Меригиппус (рис. 2.12.1.6) по величине сравнялся с нынешним пони. Средний палец на каждой его ноге был непропорционально большим по сравнению с таким же пальцем у его предшественников, и можно сказать, что меригиппус всю свою жизнь ходил на цыпочках,— точнее, не ходил, а очень быстро бегал. Многочисленные бугорки на коренных зубах помогали ему пережевывать жесткую траву. Прежде лошади были лесными жителями и питались нежной и сочной листвой. Однако к началу миоцена они приспособились к жизни на открытых равнинах.
Рис. 2.12.1.8. Дейнотерий (Deinotherium)На протяжении миоцена лошади продолжали увеличиваться в размерах. Меригиппус (рис. 2.12.1.6) по величине сравнялся с нынешним пони. Средний палец на каждой его ноге был непропорционально большим по сравнению с таким же пальцем у его предшественников, и можно сказать, что меригиппус всю свою жизнь ходил на цыпочках,— точнее, не ходил, а очень быстро бегал. Многочисленные бугорки на коренных зубах помогали ему пережевывать жесткую траву. Прежде лошади были лесными жителями и питались нежной и сочной листвой. Однако к началу миоцена они приспособились к жизни на открытых равнинах.
Теперь слоны стали куда больше похожи на их современных представителей. Мастодонт (рис. 2.12.1.7) — его называют платибелодон ("лопатобивневый") — прокладывал себе дорогу сквозь заросли подобно тяжелому бульдозеру. Из его нижней челюсти торчали широкие лопатообразные клыки, которыми он выкапывал из почвы разные растения. Дейнотерий (рис. 2.12.1.8) был намного крупнее (около 4 м в плече). Изогнутыми клыками нижней челюсти он, возможно, поддевал съедобные коренья, как большими вилами.
") Рис. 2.12.1.9. Халикотерии (Chalicothenidae)Мы можем получить некоторое представление о миоценовой экосистеме, если обратимся к ее современному аналогу — восточноафриканским саваннам. Травянистая равнина обеспечивает разными видами корма разнообразных животных. В саваннах Восточной Африки зебры объедают грубые верхушки трав, а антилопы гну их поросшие листьями центральные части. Газели же отыскивают богатые протеином семена и побеги у самой земли. Бородавочники часто опускаются на колени, чтобы дотянуться до самой короткой травы или выкопать из земли съедобные луковицы и клубни. Есть в саваннах и травоядные, которые находят корм выше уровня самых высоких трав. К примеру, черный носорог питается древесной корой, тонкими ветками и листвой, а вот слон ест и траву, и листья деревьев, зачастую поглощая за день до 250 кг растительности. Ну а жирафу рост позволяет избегать какой бы то ни было конкуренции, поскольку он может обрывать веточки и листву на высоте 6 м от земли. Таким образом, различные виды растительноядных животных не претендуют на пищевые ресурсы друг друга, и корма здесь хватает на всех. Возможно, так же было и во времена миоцена: различные виды существовали за счет различных частей экосистемы.
Рис. 2.12.1.9. Халикотерии (Chalicothenidae)Мы можем получить некоторое представление о миоценовой экосистеме, если обратимся к ее современному аналогу — восточноафриканским саваннам. Травянистая равнина обеспечивает разными видами корма разнообразных животных. В саваннах Восточной Африки зебры объедают грубые верхушки трав, а антилопы гну их поросшие листьями центральные части. Газели же отыскивают богатые протеином семена и побеги у самой земли. Бородавочники часто опускаются на колени, чтобы дотянуться до самой короткой травы или выкопать из земли съедобные луковицы и клубни. Есть в саваннах и травоядные, которые находят корм выше уровня самых высоких трав. К примеру, черный носорог питается древесной корой, тонкими ветками и листвой, а вот слон ест и траву, и листья деревьев, зачастую поглощая за день до 250 кг растительности. Ну а жирафу рост позволяет избегать какой бы то ни было конкуренции, поскольку он может обрывать веточки и листву на высоте 6 м от земли. Таким образом, различные виды растительноядных животных не претендуют на пищевые ресурсы друг друга, и корма здесь хватает на всех. Возможно, так же было и во времена миоцена: различные виды существовали за счет различных частей экосистемы.
") Рис. 2.12.1.10. Эгиптопитек (Egiptopitek)В тот же период появились и другие новоселы. В начале миоцена ряды птиц пополнились новыми видами попугаев, пеликанов, голубей и дятлов. Чуть позже к ним присоединились первые вороны и соколы. Бурно эволюционировали такие новые млекопитающие, как мыши, крысы, морские свинки и дикобразы. Появилась и странная группа лошадеобразных животных — их называют халикотериями (рис. 2.12.1.9). Большими когтями, напоминающими копыта, они выкапывали съедобные коренья.
Рис. 2.12.1.10. Эгиптопитек (Egiptopitek)В тот же период появились и другие новоселы. В начале миоцена ряды птиц пополнились новыми видами попугаев, пеликанов, голубей и дятлов. Чуть позже к ним присоединились первые вороны и соколы. Бурно эволюционировали такие новые млекопитающие, как мыши, крысы, морские свинки и дикобразы. Появилась и странная группа лошадеобразных животных — их называют халикотериями (рис. 2.12.1.9). Большими когтями, напоминающими копыта, они выкапывали съедобные коренья.
Отныне животные могли свободно перемещаться из Африки в Европу или Азию и обратно. Вскоре возникло своего рода двустороннее движение, при котором слоны мигрировали из Африки в Евразию и Северную Америку, а кошки, буйволы, жирафы и свиньи путешествовали в обратном направлении.
Первые приматы были маленькими зверьками, похожими на землероек. Они появились на Земле около 65 млн лет назад. Эволюция продолжила работу в этом направлении, и к середине олигоцена сформировались две основные группы приматов — обезьяны Нового Света (в Южной Америке) и обезьяны Старого Света (в Африке и Азии).
") Рис. 2.12.1.11. Дриопитек (Dryopithecus)Вскоре от африканской ветви произошла еще одна группа обезьян, ставшая родоначальницей человекообразных обезьян и в конечном итоге — человека. Мозг человекообразных обезьян крупнее, чем у прочих их сородичей. Кроме того, у них нет хвоста, а длинные и сильные руки отлично приспособлены для лазанья по деревьям и перепрыгивания с ветки на ветку.
Рис. 2.12.1.11. Дриопитек (Dryopithecus)Вскоре от африканской ветви произошла еще одна группа обезьян, ставшая родоначальницей человекообразных обезьян и в конечном итоге — человека. Мозг человекообразных обезьян крупнее, чем у прочих их сородичей. Кроме того, у них нет хвоста, а длинные и сильные руки отлично приспособлены для лазанья по деревьям и перепрыгивания с ветки на ветку.
В руки ученых попали ископаемые останки небольшой человекообразной обезьяны, которую назвали эгиптопитеком ("египетской обезьяной"). Она обитала в Африке в олигоценовую эпоху, около 27 млн лет назад (рис. 2.12.1.10).
Никто не может с уверенностью утверждать, что именно эгиптопитек был родоначальником современных человекообразных обезьян, но это вовсе не исключено.
Вскоре после начала миоцена (около 24 млн лет назад) появилась и другая, более высокоразвитая человекообразная обезьяна — дриопитек (рис. 2.12.1.11), похожий на нынешних шимпанзе. Эти животные быстро перебрались со своей африканской родины, через сухопутные "мосты" в Европу и Азию. По всей видимости, дриопитек ходил на двух ногах, однако бегал и лазил по деревьям при помощи всех четырех конечностей. Возможно, он даже переносил в руках пищу. Итак, история человечества должна была вот-вот начаться. [3]
Животный мир неогена. Животный мир миоцена.
А.С.Антоненко
wwlife.ru
Неогеновая система (период), неоген
 НЕОГЕНОВАЯ СИСТЕМА (ПЕРИОД), неоген (от греч. neos — новый и genos — рождение, возраст), — вторая система кайнозойской эратемы, соответствующая второму периоду кайнозойской эры истории Земли; в стратиграфической шкале следует за палеогеновой системой и предшествует четвертичной системе. Начало неогенового периода, по уточнённым данным, определяется радиометрическим методом в 23,5 млн. лет, а конец — в 0,7 млн. лет (по схеме, принятой Межведомственным стратиграфическим комитетом Академии Наук CCCP) или же в 1,8 млн. лет (по решению Международного геологического конгресса в 1948) от современности. Общая продолжительность периода 22 и 23 млн. лет соответственно. Название "неогеновая система" было предложено в 1853 австрийским геологом М. Гёрнесом для выделенных в 1833 английским геологом Ч. Лайелем двух отделов — миоцена и плиоцена.
НЕОГЕНОВАЯ СИСТЕМА (ПЕРИОД), неоген (от греч. neos — новый и genos — рождение, возраст), — вторая система кайнозойской эратемы, соответствующая второму периоду кайнозойской эры истории Земли; в стратиграфической шкале следует за палеогеновой системой и предшествует четвертичной системе. Начало неогенового периода, по уточнённым данным, определяется радиометрическим методом в 23,5 млн. лет, а конец — в 0,7 млн. лет (по схеме, принятой Межведомственным стратиграфическим комитетом Академии Наук CCCP) или же в 1,8 млн. лет (по решению Международного геологического конгресса в 1948) от современности. Общая продолжительность периода 22 и 23 млн. лет соответственно. Название "неогеновая система" было предложено в 1853 австрийским геологом М. Гёрнесом для выделенных в 1833 английским геологом Ч. Лайелем двух отделов — миоцена и плиоцена.
Подразделения неогеновой системы подразделяется на два отдела: нижний — миоцен и верхний — плиоцен. В дальнейшем эти отделы были разделены на подотделы и ряд ярусов, установленных в области Средиземноморья и некоторое время считавшихся ярусами международной шкалы. Однако после проведённой в 1970-х гг. ревизии ярусы средиземноморской шкалы стали рассматриваться только как региональные. В 1975 на конгрессе Регионального комитета по стратиграфии средиземноморского неогена были приняты три равноценные региоярусные шкалы для Средиземноморья, Западного и Восточного Паратетиса. Одновременно при изучении океанических осадков были разработаны зональные шкалы по планктонным фораминиферам и по нанопланктону, которые используются при корреляции неогена океанических районов, а также при сопоставлении региоярусных шкал отдельных регионов. Таким образом, для неогеновой системы не существует общепринятой международной ярусной шкалы. Региональные ярусы и биозоны, используемые в отдельных регионах, распределяются в пределах отделов и подотделов (табл.).
Общая характеристика. Неогеновые отложения широко распространены под покровом четвертичных отложений на всех континентах и на дне океанов. Неогеновый период был одним из наиболее геократических этапов в развитии Земли, особенно его 2-я половина — плиоцен. К концу плиоцена сформировались основные черты современного рельефа и гидросети, завершилось образование многочисленных горных систем — Альп, Карпат, Балкан, Апеннин, Крыма, Кавказа, Гималаев, Кордильер Северной и Южной Америки, островных дуг — Алеутской, Корякско-Камчатской, Японской и др. Усиление поднятий привело к образованию многочисленных внутренних впадин и глубоководных внутренних и окраинных бассейнов. Быстрый рост горных систем сопровождался складко- и шарьяже-образованием и сильной вулканической деятельностью. Для неогенового периода в целом было характерно значительное похолодание климата и образование ледниковых покровов Антарктиды и Гренландии. Снижение температуры привело к резкой дифференциации климата и соответственно ландшафтных зон. На этом общем фоне ухудшения климата наблюдались и отдельные этапы потеплений. После холодного периода 1-й половины раннего миоцена наступило значительное потепление, известное как климатический оптимум 2-й половины раннего — начала среднего миоцена. В это время в высоких широтах появились термофильные элементы среди древесных растений, пресноводных моллюсков и наземных млекопитающих на суше и ряд теплолюбивых форм среди моллюсков, фораминифер и других групп беспозвоночных — в морях.
Начиная со 2-й половины среднего миоцена вновь наступило похолодание и началась аридизация климата, продолжавшаяся в позднем миоцене. Это привело к сокращению лесов и развитию лесостепных и степных пространств. В Антарктиде возникло покровное оледенение. В плиоцене продолжалось похолодание, на фоне которого происходили неоднократные колебания температурного режима. В 1-й половине плиоцена появились покровные ледники Северного полушария. В течение неогенового периода распределение океанов, морей и суши постепенно приближалось к современному, что сопровождалось глобальным (гляцио-эвстатическим) понижением уровня океана с отдельными колебаниями, соответствовавшими масштабам оледенения. В начале миоцена крупное континентальное море — Паратетис, образовавшееся в олигоцене на северной окраине океана Тетис, потеряло связь с бореальными морями, сохранив связь с Тетисом, которая неоднократно прерывалась в миоцене. В середине миоцена Тетис окончательно распался и Средиземное море отделилось от Индо-Пацифики. В позднем миоцене Средиземное море отчленилось от океана в связи с глобальным падением уровня океана, в нём образовались мощные толщи эвапоритов (мессинский кризис солёности). В плиоцене Средиземное море вновь соединилось с Атлантикой, а Паратетис распался, и на его месте сформировались Азово-Черноморский и Каспийский бассейны. Трансгрессии морских бассейнов, захватывавшие окраинные области континентов, отмечались главным образом в 1-й половине миоцена, а в плиоцене морей с нормальной солёностью в пределах современных континентов практически не было. В целом трансгрессии неогеновых морей и осадконакопление на континентах происходили на фоне сложной и быстро меняющейся тектонической и палеогеографической обстановки, что обусловило пестроту фациального состава неогеновых отложений этих областей. Наиболее распространены были континентальные песчано-глинистые и молассовые формации; среди морских отложений основную роль играли песчано-глинистые и карбонатно-обломочные; значительное распространение имели и наземно-вулканогенные формации. Морские карбонатные отложения и эвапориты были относительно мало развиты. Биогенная седиментация в океане стала более интенсивной, чем прежде; оформились три пояса, установленные для современного океана: экваториальный, северный и южный пояса кремненакопления и карбонатонакопления. Кремненакопление в неогене (более мощное, чем в меловое и палеогеновое время) происходило в основном за счёт диатомовых водорослей. В приконтинентальных районах преобладали терригенные отложения.
Органический мир. В растительном мире неогенового периода основную роль играли те же группы, что и в современную эпоху. На суше господствовали высшие растения, а среди них покрытосеменные и голосеменные, в меньшей степени были распространены папоротники, мохообразные и пр. В умеренных широтах палеогеновые термофильные элементы отмечались только в 1-й половине миоцена, несколько расширив свой ареал в период климатического оптимума конца раннего — начала среднего миоцена. Основной компонент лесной растительности — листопадные формы. В связи с общим похолоданием и аридизацией климата проявилась широтная зональность, оформились все существующие ныне растительные зоны и флористическая область.
Растительность морей и других водоёмов была представлена различными водорослями (диатомовыми, золотистыми, багряными, пиррофитовыми и пр.) и некоторыми высшими растениями.
В Евразии в начале миоцена состав млекопитающих ещё близок к олигоценовому и только во 2-й половине раннего миоцена появились формы миоценового типа — хоботные (гомфотерии, зиголофодоны) и др. В среднем миоцене наряду с хоботными (мастодонты) и лошадиными (анхитерии) распространились бовиды и другие обитатели полуоткрытых и сухих пространств. В позднем миоцене сформировались сообщества степных травянистых равнин — т.н. гиппарионовая фауна (гиппарионы и разнообразные бовиды — газели, палеориксы и др.), существовавшая до конца миоцена — начала плиоцена. В плиоцене на территории Евразии несколько раз происходила смена фаун млекопитающих. На границе плиоцена и плейстоцена (0,7 млн. лет) появились холодолюбивые и перигляциальные формы — мамонт, овцебык, сайга и др. Фауна неогеновых млекопитающих Африки была близка к евразиатской. Млекопитающие Австралии были представлены отрядами сумчатых и однопроходных. В Южной Америке в миоцене основную роль играли сумчатые, неполнозубые (муравьеды, ленивцы, броненосцы), грызуны и некоторые эндемичные копытные. В позднем миоцене произошёл обмен фаунами Северной Америки с Евразией, а в плиоцене отмечена миграция млекопитающих (хищники, лошади и др.) из Северной Америки в Южную.
В неогеновых океанах и морях были распространены представители простейших (фораминиферы, в меньшей степени — радиолярии), брюхоногие и двустворчатые моллюски, ракообразные (остракоды и другие группы), мшанки, иглокожие (морские ежи), кишечнополостные (шестилучевые кораллы), а из хордовых — костистые и хрящевые рыбы и водные млекопитающие.
Полезные ископаемые. С неогеновыми отложениями связаны многочисленные полезные ископаемые. Из осадочных полезных ископаемых наиболее важны месторождения нефти и газа в предгорных и межгорных прогибах Ближнего и Среднего Востока, Калифорнии, Аляски, Японии и т.д. В CCCP к нефтегазоносным регионам, отвечающим крупным предгорным прогибам, относятся Прикарпатский, Азово-Кубанский, Терско-Каспийский; межгорным впадинам — Закарпатский, Восточно-Черноморский, Южно-Каспийский и Ферганский; внутрискладчатым депрессиям — Сахалино-Охотский, Южно-Охотский и Анадырский. Достаточно многочисленны в неогеновых отложениях месторождения бурых углей и лигнитов, несколько реже отмечаются и каменные угли. Так, на территории CCCP разрабатываются месторождения Дальневосточной, Колымской, Центрально-Якутской, Амурской, Прибайкальской угленосных провинций и областей, Закарпатского и Южно-Уральского буроугольных бассейнов и др. На территории зарубежной Европы известны буроугольные месторождения в ГДР, Болгарии, Венгрии, Румынии, Чехословакии, Югославии и др. В Азии наиболее крупные залежи угля связаны с Анатолийским буроугольным бассейном, известны месторождения угля также в Индии, Китае, Юго-восточной Азии. Значительными запасами обладают угольные месторождения Северной и Южной Америки и Австралии (бассейны Латроб-Валли и др.). Отмечены месторождения серы, связанные главным образом с эвапоритовыми формациями (Предкарпатье, Апеннинский полуостров, Сицилия), а также залежи солей (Предкарпатье, Закарпатье, Закавказье, Средняя Азия и др.). Образовались россыпные месторождения титана, олова, ильменита, рутила, циркона и др., многие бокситовые месторождения тропического пояса (Ямайка, Гайана, Суринам, Гана, Гвинея). Значительны также месторождения бентонитовых и палыгорскитовых глин, алунитов, перлитов, каолинитов, галлуазита. В качестве строительных материалов, керамического и цементного сырья используются известняки, кварцевые пески, песчаники, диатомиты, глины.
С интрузивными и эффузивными породами связаны многочисленные и разнообразные месторождения руд ртути, олова, свинца, цинка, сурьмы и других элементов, образующие местами рудные пояса (полиметаллический пояс Перу, золотоносный пояс Эквадора, оловоносный и меденосные пояса Боливии, медно-серебряные месторождения Центральной Америки, медистые песчаники Таджикской депрессии, полиметаллические месторождения и ртутные рудные провинции Альпийского складчатого пояса, в т.ч. Закарпатья). В северо-западной и западной частях Тихоокеанского побережья, в т.ч. на Корякском нагорье, Камчатке, Курильских островах, Сахалине, распространены касситерит-сульфидные, колчеданные, ртутные, сурьмяные рудные провинции, месторождения руд свинца, цинка, меди, золота, серебра, молибдена, мышьяка, хрома и других металлов. Области интенсивного вулканизма и магматизма перспективны для поисков руд цветных металлов. В океанических областях песчано-глинистые шельфовые отложения во многих районах нефтегазоносны: Мексиканский залив, Карибское море, Гвинейский залив, Средиземное, Красное, Берингово моря, залив Аляска, Японское и Южно-Китайское моря, шельфы Чили, Перу, Эквадора, Калифорнии. На северо-восточном шельфе Австралии продуктивная толща приурочена к миоценовым рифам. Большие перспективы связаны с Железомарганцевыми конкрециями, содержащими многие ценные металлы (никель, медь, кобальт и др.), и фосфоритами. Фосфориты и фосфатизированные осадки и породы распространены как на шельфах и в верхних частях континентальных склонов, так и в пелагических областях, преимущественно на подводных горах. Часть обогащенных фосфоритами пород имеет миоценовый возраст — шельфы северо-западной, юго-западной и южной Африки, восточных и западных берегов Северной Америки, Новой Зеландии, подводные горы Тихого, Индийского и Атлантического океанов.
www.mining-enc.ru
Четвертичный период (антропоген)
Четвертичный период (антропоген) берет свое начало 2,6 млн. л. н. и длится по сей день. За время, которое длится данный временной отрезок, случились три главные вещи:
- планета вступила в новую ледниковую эпоху, в ходе которого резкие похолодания чередовались с потеплениями;
- материки приняли свои окончательные нынешние очертания, сформировался современный рельеф;
- на планете появился человек разумный.
Подразделы антропогена, геологические изменения, климат
Почти всю протяженность антропогена занимает отдел Плейстоцен, который по международным меркам стратиграфии принято делить на Гелазский, Калабрийский, Средний и Верхний ярусы, и Голоцен, берущий свое начало чуть больше 11 тыс. л. н. и длящийся и по сей день.
| Четвертичный период (антропоген) | Отделы | Ярусы |
| Плейстоцен | Гелазский | |
| Калабрийский | ||
| Средний | ||
| Верхний | ||
| Голоцен |
В основе своей материки в нынешнем обличье сложились уже задолго до начала четвертичного периода, но именно в данный промежуток времени многие молодые горные цепи обрели нынешний вид. Береговая линия материков приняла нынешние очертания, а за счет попеременно наступавших и отступавших ледников сформировались крайние северные континентальные архипелаги, такие как канадский, о-ва Шпицберген, Исландия, Новая земля и пр. В ходе попеременных оледенений в некоторые промежутки времени уровень Мирового океана понижался до 100 метров.
Отступая, гигантские ледники антропогена оставляли за собой след из глубоких морен. В периоды самых максимальных оледенений общая площадь ледников превышала нынешние более чем втрое. Таким образом, можно сказать, что большие части Северной Америки, Европы и нынешней России была погребена под ледовыми толщами.
Стоит сказать, что нынешняя ледниковая эра в истории земли не первая. На протяжении нескольких миллиардов лет длилась первая историческая ледниковая эпоха, бравшая начало 1,5 млрд. л. н. в раннем протерозое. После продолжительного тепла на планету вновь обрушилось 270-миллионолетнее похолодание. Случилось это 900 млн. л. н. в позднем протерозое. Далее имело место еще одно значительное обледенение, длившееся на протяжении 230 млн. л. н. в палеозое (460 – 230 млн. л. н.). И вот теперь планета переживает очередное похолодание, начало которого принято относить к 65 млн. л н. Оно постепенно набирало силу и еще не известно, пережила ли кайнозойская глобальная ледниковая эпоха свой апогей низких температур.
")
Рис. 1 – Антропоген (четвертичный период)
За время текущей ледниковой эпохи случилось великое множество потеплений и похолоданий, и как утверждают ученые, на данном промежутке времени Земля переживает стадию потепления. По их подсчетам последнее похолодание сменилось потеплением от 15 до 10 тыс. л н. Во времена сильнейших оледенений плейстоцена линия ледников опускалась от 1500 до 1700 км южнее нынешней черты.
Климат антропогена был подвержен многократным колебаниям. В те времена, когда ледники наступали, климатические зоны сужались и отступали ближе к экватору, и, наоборот, в периоды потеплений и массового таяния ледников, умеренный пояс распространялся до самых северных континентальных окраин и, как следствие, ширились и остальные климатические пояса.
Осадконакопление четвертичного периода
На осадконакопление четвертичного периода наложила свой отпечаток быстрая изменчивость литологических составляющих и генезиса. Осадки в четвертичном периоде накапливались повсеместно, но ввиду сложного строения разрезов их достаточно сложно выделить. Скорость накопления отложений антропогена была слишком высока, но за неимением давления отложения до сих пор имеют достаточно рыхлую структуру. Условия залегания также нетипичны. Если типичным считается последовательное напластовывание, то здесь более подходит термин «прислонение» к нижним и более древним отложениям. Материковым зонам более характерны континентальные отложения, такие как ледниковые, водные и эоловые. Для морей более типичны вулканические, органогенные, тригенные и хемогенные осадки.
Животные четвертичного периода
Среди беспозвоночных в плейстоцене четвертичного периода необычайно развились всевозможные улитки и прочие сухопутные моллюски. Подводный же мир был во многом схож с предыдущим неогеном. Начинал приобретать схожесть с настоящим и мир насекомых, но самым интересным метаморфозам был подвержен мир млекопитающих.
С начала антропогена широкое распространение получили разновидности слоноподобных. В начале плейстоцена они заселяли огромные территории Евразийского континента. Некоторые их виды в высоту достигали 4 м в холке. Все чаще в северных частях континентов стали появляться виды слонов, покрытых длинной шерстью. К середине плейстоцена мамонтовые были уже самыми обычными и наиболее распространенными представителями северных тундровых широт. Мигрировав по льдам Берингового пролива в один из очередных отрезков похолодания на Аляску, мамонты расплодились и на всей части североамериканского континента. Как считается, мамонты произошли от трогонтериевых слонов, на границе неогена и плейстоцена, широко распространенных в степных широтах.
В южных широтах, как Северной Америки, так и Евразии широко были распространены другие виды слоновых. Среди прочих особо выделялись гигантские мастодонты. Что характерно, эти представители слоновьих на территории евразийского материка полностью вымерли к концу плейстоцена, в то время, как на американском континенте они благополучно пережили все этапы оледенения Земли.
Среди прочих гигантов четвертичного периода также выделялись носороги. Их шерстистые разновидности заселяли тундростепи раннего и среднего антропогена наряду с мамонтами.
Многочисленны были животные четвертичного периода из разряда лошадиных. Что характерно, древний потомок лошадей был родом из североамериканской части Пангеи. После раскола материка и прекращения миграции животных между американским и евроазиатским сегментами лошадиные полностью вымерли на североамериканском материке, и эволюционировали лишь те их виды, которые успели мигрировать на евроазиатский континент. Впоследствии они вновь появились в Америке лишь благодаря человеку.
Наряду с лошадиными, в больших количествах населявшими европейско-азиатские саванны, в периоды потеплений антропогена активность проявляли и гиппопотамы. В больших количествах их останки были найдены на островах Великобритании. Многочисленны были и различные парнокопытные разновидности оленей, самым распространенным из которых был ирландский большерогий. В размахе его рога порой достигали до 3 метров.
В четвертичном периоде появились первые козлиные, среди которых наиболее многочисленными были горные разновидности. Появились первые туры, прародители домашних быков. На степных просторах паслись огромные пастбища всевозможных косуль, зубров, овцебыков, южнее появились первые разновидности верблюжьих.
Также, наряду с травоядными, развивался и отряд хищников. К примеру, разнообразных медвежьих можно было встретить как на заснеженных участках северных широт, так и в тундровых лесах. Многие из них обитали и южнее, спускаясь до степной полосы умеренных широт. Множество из них, населявших пещеры ледникового плейстоцена, не смогли выжить в холодных условиях Арктики того времени, но, так или иначе, многие их разновидности благополучно дожили и до сегодняшних дней.
Многочисленны были в северных районах такие смертоносные хищники антропогена (рис. 2), как саблезубые тигры, и пещерные львы, которые были значительно массивнее и крупнее и опаснее своих современных сородичей. Нередко эти опасные хищники становились темами творчества древних наскальных художников.

Рис. 2 – Хищники четвертичного периода
Также в числе прочих фауну четвертичного периода представляли и прочие разнообразные виды, такие как гиены, волки, лисы, еноты, росомахи и пр. Имело место и большое количество грызунов в лице леммингов, сусликов, бобров различных разновидностей, вплоть до гигантских Trognotherium cuvieri.
Большим разнообразием отличалось и царство пернатых, среди которых выделялись как летающие, так и нелетающие разновидности.
К концу плейстоцена вымерло множество разновидностей млекопитающих, ранее населявших тундростепи. К таким млекопитающим четвертичного периода можно отнести:
- на территории Южной Америки – броненосца тетикуруса, гигантскую саблезубую кошку смилодон, копытных макраухений, ленивцев мегатерий и пр.;
- на территории Северной Америки – последних представителей птиц-тиранов или фороракосов – титанисов Уоллера, многих представителей копытных, таких как американские лошади, верблюды, степные пека́ри, олени, быки и вилорогие антилопы;
- на территории тундростепей Евразии, Аляски и Канады – мамонтов, шерстистых носорогов, большерогих оленей, пещерных львов и медведей.
В голоцене вымерли такие птичьи нелетающие разновидности, как дронты, эпиорнисы, исчезла из морских глубин гигантское тюленеподобное стелларова корова.
Растения антропогена
Климат плейстоцена с постоянными чередованиями ледниковых и межледниковых промежутков оказывал неблагоприятное воздействие на растения антропогена, произраставшие в северных континентальных широтах. Климатический жизненный барьер с наступлением похолоданий порой был вынужден смещаться до черты 40° с. ш., а местами и еще ниже. На протяжении последних двух с лишним миллионов лет растительность вынуждена попеременно то отступать до вышеозначенных широт, то вновь разрастаться вплоть до берегов Северного Ледовитого океана. В результате похолодания многие теплолюбивые растения, которые вели свой род еще со времен триаса, были обречены на вымирание. С исчезновением многих разновидностей трав, кустарников и прочих растений связано и вымирание многих видов животных антропогена. Поэтому не стоит всю вину за исчезновение таких видов, как тот же мамонт, возлагать целиком и полностью на плечи древних людей.
В ледниковые эпохи четвертичного периода южнее оконечности ледников обретали существование три полосы растительности – тундровая, степная и таежная. Тундра была покрыта мхами и лишайниками, южнее начинали расти карликовые березы, полярные ивы, альпийские серебрянки. Для тундры также типичны были азалии, камнеломы, ясколки и пр. Степная зона была полна всевозможными травами и низкими кустарниковыми. Но ближе к югу кое-где встречались и пролески, состоящие из ивовых и березняка. Таежные леса антропогена составляли в основном из сосен и елей, которые ближе к югу смешивались с березами, осинами и другими листопадными лиственными.
В межледниковые эпохи состав флоры четвертичного периода значительно менялся. Оттесненные южнее ледниками, на свои места возвращались заросли таких цветковых и кустарниковых, как лилии, рододендроны и розы. Но понемногу к приближению голоцена межледниковая растительность все более скудела за счет постоянных вынужденных миграций. Редки ныне стали многие ореховые и тисовые, раннее образовывавшие огромные лесные массивы. В самые теплые промежутки межледниковий центрально-европейскую территорию сплошь покрывали широколиственные леса, состоящие из дуба, буков, лип, кленов, граба, ясеня, боярышника и некоторых ореховых.
В местах, где межледниковые миграции растений не были затруднены горными хребтами и морями все же сохранились и образчики древней растительности триасового периода. Например, на территории Северной Америки, где миграции не были затруднены как в случае с горными массивами Европы, а также Средиземным морем, до сих пор в некоторых районах произрастают магнолии, лилиодендроны, таксодиумы и сосны веймутова (Pinus strobus).
Намного южнее растительность не претерпела определенных различий с предыдущим неогеновым периодом.
Становление человека в период антропогена
Предки нынешних людей появились еще в конце неогена 5 млн. л. н. Ими были произошедшие от одной из ветвей гоминид австралопитеки, причем их останки найдены лишь на африканском континенте, что дает повод сказать, что прародина всего человечества является именно Африкой. Теплый климат и бурная растительность данных мест способствовала все большему эволюционному развитию австралопитеков, пока, наконец, первые из них на рубеже четвертичного периода не освоили примитивные виды орудий труда. Следующей ветвью развития человека умелого (Homo habilis) стали архантропы, прямые предки современных людей, которые во второй половине плейстоцена активно стали расселяться по всем материкам. Одним из известнейших ответвлений архантропов являются питекантропы, останки которых археологи находят почти повсеместно. В районе 400-350 тыс. л. н. стали появляться первые переходные формы древних людей от архантропов к палеоантропам, к числу которых можно отнести неандертальцев, которые впоследствии вымерли, не выдержав конкуренции со стороны кроманьонцев. Хотя, как считают некоторые ученые, эти два вида просто смешались друг с другом. Далее палеоантропы развились в неоантропов, которые уже мало отличались от современных людей. Произошло это в районе 40-35 тыс. л. н. В частности кроманьонцы и являли собой первых представителей неоантропов.

Рис. 3 – Становление человека в период антропогена
Постепенно люди осваивали все более сложные орудия труда. 13 тыс. л. н. появились лук и стрелы, после люди научились обжигать горшки и обзавелись первыми предметами из керамики. Они начали заниматься земледелием, скотоводством. 5 тыс. л. н. появились первые изделия из бронзы и меди, а где-то между 3 и 2,5 тыс. л. н. началась эпоха железа.
С этого времени совершенствование орудий труда пошло гораздо быстрее, в средние века началось развитие науки и техники, которые в настоящее время достигли уровня, позволившего людям развить такие науки, как генетика и генная инженерия.
Полезные ископаемые четвертичного периода
Четвертичные отложения содержат в себе множество различных ископаемых. Россыпные отложения в черте горных кряжей и зонах тектонической активности богаты золотом, алмазами, касситеритом, ильменитом и пр. Отложения, формирующиеся во влажных тропических зонах и представляющие собой коры выветривания, содержат запасы бокситов, марганца и никеля, а также таких нерудных строительных материалов, как суглинки, глины, галечник, песчаник, известняк. Также многочисленны накопления бурых углей, попадаются месторождения природного газа, диатомитов, солей, бобовых железных руд, сапропелей и пр. Также в вулканических районах можно встретить залежи серы и марганца. Многочисленны и повсеместны торфяные осадочные накопления.
В пластах четвертичного периода содержится огромное количество пресных подземных вод, в их недрах берут свое начало некоторые термальные источники, также в наше время интенсивно используются различные лечебные грязи, сформировавшиеся в антропогене.
vse-lekcii.ru
Неогеновый период
НЕОГЕНОВЫЙ ПЕРИОД (неоген), второй период кайнозойской эры. Длился ок. 23 млн. лет. Начался ок. 25 млн. лет назад, завершился 2 млн. лет назад. Неоген разделяют на 2 эпохи: продолжительный миоцен и короткий плиоцен. Для миоцена характерна активная вулканическая деятельность. Огромные площади суши покрывались толстыми потоками лавы. Завершалось формирование новых гор альпийской системы – Кордильер, Анд, Кавказа, Карпат, Гималаев. Все материки приобрели современные границы, но разделялись морскими проливами. Северная Америка с Азией несколько раз соединялась «мостами» суши. К югу от Европы простирались внутренние моря Тетис и Паратетис, связывавшие Атлантику с Индийским океаном (эта связь закрылась только в конце плиоцена, тогда же Северная Америка соединилась с Южной). Климат почти всё время был тёплым и влажным, но в конце плиоцена началось похолодание, на полюсах образовались шапки льда.
Флора в миоцене приближалась к современной. Открытые ландшафты быстро заселялись травянистыми растениями. На этой основе происходила бурная эволюция копытных и грызунов, а вслед за ними – новых хищников: кошек, собак, медведей, енотов, куниц и др. В Африке в начале миоцена появились человекообразные обезьяны, к его концу среди них выделились первые гоминиды (австралопитековые), а в конце плиоцена появились первые представители рода «Гомо» (человек). В изолированной Австралии продолжали процветать сумчатые, а в Южной Америке наряду с сумчатыми эволюционировали широконосые обезьяны и несколько эндемичных отрядов плацентарных млекопитающих (неполнозубые, литоптерны и др.). Хищных млекопитающих в Южной Америке не было, и их место заняли птицы – утратившие полёт фороракосы, родственные дрофам и журавлям. Миоценовая эпоха была временем наибольшего процветания животного и растительного сообществ. По палеонтологическим данным, ей свойственно самое значительное биоразнообразие за всю историю кайнозойской эры.
sbio.info
Неоген на сайте Игоря Гаршина
 |
Продолжительность неогена – чуть больше 20 миллионов лет. Эпохи: миоцен (23,0 [~1000 прецессий Земли назад] - 5,33 м.л.н.), плиоцен (5,33 - 2,588 м.л.н.). |
Господство млекопитающих и птиц.
Разделы страницы о неогене:
- Обзоры и статьи о живом мире неогена
- Миоцен (23,0 - 5,3 млн. лет назад)
- Плиоцен (5,3 - 2,6 млн. лет назад)
Обзоры и статьи о живом мире неогена
Неогеновый период, или неоген,- один из самых коротких периодов в истории Земли (абс. возраст 25+2; 1,5+0,5 млн. лет). В развитии органического мира значительных изменений не происходило.
Геодинамика и климат неогена
Неоген был периодом преимущественного поднятия значительных участков земной коры, периодом активного формирования континентальных образований. Поднятия в конце олигоцена привели к тому, что в пределах платформ Северного полушария в неогене установился континентальный режим.
В конце неогенового периода климат посуровел настолько, что появились ледники (а Антарктида покрылась льдом полностью). Началось тяжёлое испытание жизни на планете, ставшее одним из переломных моментов в её истории. Тогда-то и появилось человечество.
Мир беспозвоночных в неогене
Мир морских беспозвоночных очень близок к современному. На суше из беспозвоночных господствуют различные группы членистоногих, в первую очередь насекомые. Их состав даже на родовом уровне почти не меняется в течение периода.
Эволюция позвоночных в неогене
Среди позвоночных перемены также невелики. Никаких крупных изменений на отрядном и семейственном уровнях ни на рубеже палеогена и неогена, ни в неогене не происходило. Можно указать два отряда костистых рыб, появившихся в миоцене, но это монотипические группы, известные только из миоцена. Появляются два новых семейства бесхвостых амфибий, а одно палеогеновое, возможно, вымирает. Небольшие изменения произошли на семейственном уровне в подклассе черепах.
Из позвоночных безусловно господствуют плацентарные млекопитающие. Происходило вымирание некоторых групп, особенно связанных с влажными лесами и болотистыми пространствами; что объясняется постепенным изменением климата в сторону более сухого, возникновением значительного пространств-а стеnей. Общее число семейств к концу периода уменьшается до 119 (Решетов, Трофимов, 1979).
Млекопитающие осваивают моря и воздух – возникают киты ["древние киты" - еще в палеогене, но были они тогда "земноводными"?] и рукокрылые. Плацентарные оттесняют на периферию остальных млекопитающих. Фауна этого периода становиться очень похожей на современную. Но есть и отличия – еще существуют мастодонты, гиппарионы, саблезубые тигры. Крупные нелетающие птицы играют большую роль, особенно в изолированных, островных экосистемах. /p>
Идет быстрая эволюция хоботных и лошадиных. К неогену относится знаменитая «гиппарионовая фауна», в это время происходит окончательное становление рода собственно лошадей. Уже известны медведи, собаки, гиенавые и некоторые другие группы, характернрrе для настоящего времени. Исключительно важно дальнейшее развитие приматов, в том числе и человекообразных, а также постепенное становление в конце периода предков человека. В морях появились ластоногие, продолжалось развитие китообразных.
Происходит значительная дифференциация фауны разных частей света, что связано с теми изменениями конфигурации материков, о которых было сказано выше. Самая богатая фауна известна из Евразии. В Северной Америке, Южной Америке, Австралии развитие шло своими путями, хотя периодически происходил некоторый обмен группами млекопитающих между Евразией и Северной Америкой, Северной и Южной Америкой благодаря появлению кратковременных мостов, соединявших эти материки.
Тем не менее, в Южной Америке развивались свои группы сумчатых, неполнозубых, копытных, широконосых обезьян. В Австралии, не вступавшей в контакт с другими-материками, продолжали существовать только сумчатые и однопроходные.
Растительный мир неогена
В растительном мире происходило постепенное оттеснение теплолюбивых флор в более низкие широты и смена их флорами умеренного климата на большей части материков северного полушария. Как сказано выше, значительное место занимали обширные травянистые покровы, прекрасно приспособленные к разным неблагоприятным условиям.
Миоцен (23,7 - 5,3 млн. лет назад)
Миоцен приходится на середину третичного периода и характеризуется увеличением площади пастбищ по всему земному шару и возникновением большинства современных групп млекопитающих. Похолодание климата Северной Евразии продолжилось.
Для миоцена характерна активная вулканическая деятельность. Огромные площади суши покрывались толстыми потоками лавы. Завершалось формирование новых гор альпийской системы – Кордильер, Анд, Кавказа, Карпат, Гималаев. Все материки приобрели современные границы, но разделялись морскими проливами. Северная Америка с Азией несколько раз соединялась «мостами» суши. К югу от Европы простирались внутренние моря Тетис и Паратетис, связывавшие Атлантику с Индийским океаном (эта связь закрылась только в конце плиоцена, тогда же Северная Америка соединилась с Южной). Климат почти всё время был тёплым и влажным, но в конце плиоцена началось похолодание, на полюсах образовались шапки льда.
Флора в миоцене приближалась к современной. Открытые ландшафты быстро заселялись травянистыми растениями. На этой основе происходила бурная эволюция копытных и грызунов, а вслед за ними – новых хищников: кошек, собак, медведей, енотов, куниц и др. В Африке в начале миоцена появились человекообразные обезьяны, к его концу среди них выделились первые гоминиды (австралопитековые), а в конце плиоцена появились первые представители рода «Гомо» (человек). В изолированной Австралии продолжали процветать сумчатые, а в Южной Америке наряду с сумчатыми эволюционировали широконосые обезьяны и несколько эндемичных отрядов плацентарных млекопитающих (неполнозубые, литоптерны и др.). Хищных млекопитающих в Южной Америке не было [а ягуары?], и их место заняли птицы – утратившие полёт фороракосы, родственные дрофам и журавлям. Миоценовая эпоха была временем наибольшего процветания животного и растительного сообществ. По палеонтологическим данным, ей свойственно самое значительное биоразнообразие за всю историю кайнозойской эры.
- История Земли в миоцене.
- Океанская и климатическая эволюция в миоцене.
Плиоцен (5,3 - 1,6 млн. лет назад)
В период плиоцена климат Северного полушария становился постепенно более сухим и холодным. В конце плиоцена возник Гренландский ледниковый щит и началось оледенение на континентах Северного полушария, расширилось и оледенение континентов Южного полушария.
Растительность стала более холодостойкой, увеличилась площадь степных ассоциаций.
До конца плиоцена продолжала существовать гиппарионовая фауна (названа по преобладающему виду гиппарионов; включала также предков носорогов, мастодонтов, жирафов, антилоп и других копытных, некоторых хищных, грызунов, обезьян, а также страусов, некоторых птиц и других позвоночных), но ее вытеснили к концу плиоцена настоящие лошади, слоны и др.
В плиоцене возникли австралопитеки и первые люди.
|
Ключевые слова для поиска сведений о неогеновом периоде: На русском языке: неогеновый период, неоген, миоцен, плиоцен, глобальное похолодание, оледенение континентов; На английском языке: Neogene. |
|
www.garshin.ru










