Видеоматериалы. Занятие № 15 «Как поступает и удерживается вода в растениях на что расходуется». Как поступает вода в растение
| Еще в 1864 г. Ю. Сакс установил, что поступление воды через корневые системы зависит от температуры. С понижением температуры скорость поступления воды резко сокращается. Это может оказать заметное влияние на растительный организм, особенно в осенний период, когда испарение идет еще достаточно интенсивно, а поступление воды задерживается из-за пониженной температуры почвы. В результате растения завядают и даже могут погибнуть от обезвоживания Причин, по которым понижение температуры вызывает уменьшение поступления воды, по-видимому, несколько: 1) повышается вязкость воды и, как следствие, снижается ее подвижность; 2) уменьшается проницаемость цитоплазмы для поды; 3) тормозится рост корней; 4) уменьшается скорость всех метаболических процессов. Последнее обстоятельство, по-видимому, должно сказаться косвенно, через уменьшение поступления солей и, как следствие, торможение работы нижнего концевого двигателя (корневого давления). Торможение поступления йоды в корневую систему при действии пониженной температуры можно легко продемонстрировать в простом опыте. Так, если положить кусок льда на поверхность почвы в сосуде с растением, то в условиях интенсивной транспирации растение через два часа обнаружит признаки завядания. Если затем удалить лед, то растение снова приобретет тургор. Снижение аэрации почвы также тормозит поступление воды. Это можно наблюдать, когда после сильного дождя все промежутки почвы заполнены водой и вместе с тем на солнце при сильном испарении растения завядают. Это связано С тем, что все условия, снижающие метаболизм (недостаток кислорода, избыток С02, дыхательные яды) снижают поступление ионов и, как следствие, уменьшают поступление воды. Вместе с тем исследования показали, что особенно резкое подавление поступления воды происходит при увеличении содержания С02. Возможно, избыток углекислого газа помимо ингибирования дыхания, повышает вязкость воды, снижает проницаемость цитоплазмы. Большое значение имеет содержание воды в почве, а также концентрация почвенного раствора. Естественно, вода поступает в корень только тогда, когда водный потенциал корня ниже (более отрицателен) водного потенциала почвы. В том случае, если почвенный раствор имеет более отрицательный осмотический потенциал, вода не только не будет поступать в корень, но будет выделяться из него. Особенное значение это имеет для засоленных почв. Именно поэтому растения, растущие на этих почвах (галофиты), имеют резко отрицательный осмотический потенциал. Всякое уменьшение влажности почвы снижает поступление воды. Чем меньше воды в почве, тем с большей силой она удерживается и тем меньше ее водный потенциал. Для того чтобы в растение поступила вода, должен существовать градиент водного потенциала в системе почва — растение — атмосфера. Надо также учитывать, что уменьшение содержания воды в корне затрудняет ее дальнейшее продвижение к сосудам ксилемы. Это объясняется тем, что при уменьшении содержания воды сопротивление ее передвижению по клеткам корня растет. Подсыхающие клеточные оболочки оказывают значительное сопротивление передвижению воды. Сопротивление корневой системы передвижению воды при се недостатке настолько велико, что, по мнению Н. А. Максимова, это может снижать транспирацию, т. е. являться средством для ее регуляции. С физиологической точки зрения удобно выделить следующие формы почвенной влаги, различающиеся по степени доступности для растений. Гравитационная вода заполняет крупные промежутки между частицами почвы, она хорошо доступна растениям. Водный потенциал этой формы воды зависит от осмотической концентрации и составляет -0,1 бар. Однако, как правило, она легко стекает в нижние горизонты под влиянием силы тяжести, вследствие чего бывает в почве лишь после дождей. Капиллярная вода заполняет капиллярные поры в почве. Эта вода также хорошо доступна для растений, удерживается в капиллярах силами поверхностного натяжения и поэтому не только не стекает вниз, но и поднимается вверх от грунтовых вод . Пленочная вода окружает коллоидные частицы почвы. Вода из периферических слоев гидратационных оболочек может поглощаться клетками корня. Вместе с тем чем ближе к коллоидным частицам располагаются молекулы воды, тем с большей силой они удерживаются и, как следствие, менее доступны для растений. Гигроскопическая вода адсорбируется сухой почвой при помещении ее в атмосферу с относительной влажностью воздуха 95%. Количество воды (в %), при котором растение впадает в устойчивое завядание, называют коэффициентом завядания или влажностью завядания. Влажность, при которой наступает завядание на данной почве, зависит от ряда причин. Считается, что растения завядают в тот момент, когда вода в почве перестает передвигаться. Однако было показано, что если завядание начинается при одной и той же влажности, то промежуток времени от завядания до гибели (интервал завядания) у растений может быть резко различным. Так, для растений бобов интервал завядания составляет несколько суток, тогда как для проса — несколько недель. Это, естественно, сказывается на устойчивости растений к засухе. Вместе с тем влажность, при которой наступает завядание, в большой степени зависит от скорости транспирации, а также от осмотического потенциала клеток растения. Так, при одной и той же влажности почвы завядание начинается позднее у растений с меньшим (более отрицательным) осмотическим потенциалом. Большое значение при определении показателя влажности завядания также имеет тип почвы. Одно и то же растение завядает на черноземе при значительно более высокой влажности, чем на подзоле. Это связано с тем, что черноземные почвы характеризуются более тонким механическим составом. Влажность завядания еще не дает представления о количестве воды, полностью недоступной растению, поскольку при завядании какое-то количество воды продолжает поступать в растительный организм. В связи с этим было введено понятие «мертвый запас» влаги в почве — это количество воды, полностью недоступное растению. Мертвый запас зависит исключительно от типа почвы, ее механического состава, что видно из следующей формулы: q = % песка (0,01) + % пыли (0,12) + % глины (0,57), где q — мертвый запас. Чем больше глинистых частиц в почве, тем больше мертвый запас влаги. Количество доступной для растительного организма воды представляет разность между полевой влагоемкостью (максимальное количество воды, удерживаемое почвой в капиллярах) и мертвым запасом. В связи со сказанным перегнойные суглинистые почвы обладают не только наибольшим мертвым запасом, но и наибольшим запасом доступной влаги. Так, если полевая влагоемкость черноземных почв составляет 40%, мертвый запас — 25%, то количество доступной воды равно 25%, тогда как у песка попевая влагоемкость — 15%, мертвый запас — 5%, количество доступной воды всего 10%. |
fizrast.ru
Видеоматериалы. Занятие № 15 «Как поступает и удерживается вода в растениях на что расходуется»
Кинопособие содержит методический аппарат, обеспечивающий помощь учителю на всех этапах проведения уроков.
Кинопособие « Экология растений» 58 занятий
Актуальность:
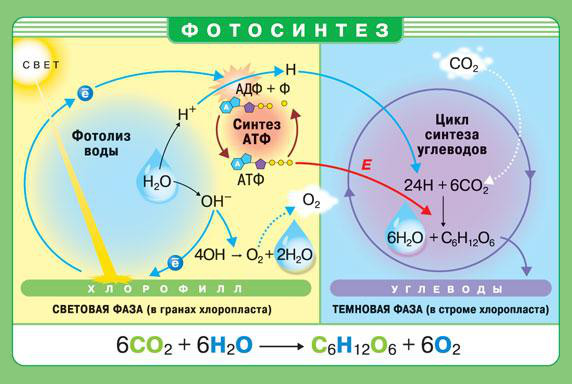
В клетках растения очень много воды. Она содержится в цитоплазме, ядре, пластидах, пропитывает клеточную оболочку, много ее в вакуоли, где находится клеточный сок.
Клетки растений содержат 85—90% воды. Как поступает вода в растения?
Тип учебного занятия; изучения и первичного закрепления новых знаний
Дидактическая цель; создать условия для осознания и осмысления блока новой учебной информации.
Формы проведения занятия; лекция, учебный фильм
Основные понятия
Внекорневая подкормка. Эпифиты. Воздушные корни. Суккуленты
Корневые волоски, корневые системы. Кутикула, пробка
Вопросы для обсуждения
1.Какими путями вода может поступать в растения?
2.Как растения удерживают воду?
3.На что расходуют воду растения?
4.Какое значение имеет испарение воды для растенийц?
Как поступает вода в растения. Растения, погруженные в воду, поглощают ее всей поверхностью тела. Вода легко проходит через тонкие клеточные оболочки.
Корни наземных растений всасывают воду из почвы корневыми волосками. Это основной путь поступления воды в растения. В дождевых тропических лесах на деревьях растут эпифиты (фитон” — растение). Среди них много красивых орхидей, у некоторых есть воздушные корни. Их особая поверхностная ткань может поглощать влагу из воздуха.
В небольшом количестве воду поглощают листья во время дождя. На этом основана внекорневая подкормка растений. Их опрыскивают питательным раствором, проникающим внутрь листа, если кутикула на листьях тонкая.
В пустынях Средней Азии встречаются высокие травянистые растения ферула и дорема. Листья их имеют большие влагалища, размером и формой напоминающие чайные чашки (рис.1). Во время дождя в них накапливается вода. Она поступает внутрь растения через тонкие оболочки клеток стебля. Собравшуюся в этих “чашках” дождевую воду иногда пьют копытные животные пустынь джайраны. Поэтому в народе эти растения называют “джайраньи чашки”.
Рис. 1. Листья ферулы с чашевидными Рис. 2. Очиток едкий — листовой суккулент
Как растения удерживают воду. Вы знаете, что на побегах всех наземных растений есть покровная ткань — кожица. Ее клетки расположены очень плотно. Снаружи они покрыты тонкой пленочкой — кутикулой, почти не пропускающей воду. Испарение происходит в основном через устьичные щели. На многолетних побегах кожица заменяется пробкой. Кожица и пробка ограничивают испарение воды и помогают растениям удерживать воду внутри тела.
Наверняка вы видели комнатные растения с сочными, мясистыми листьями и стеблями: алоэ, кактусы, толстянки и др. Такие растения называют суккулентами, что означает сочный.
Растут они преимущественно в пустынях, где воды мало, поэтому растения должны экономно ее расходовать. У них очень толстая кутикула, а часто поверх нее и восковой налет. В клетках суккулентов содержится много слизистых веществ, которые прочно удерживают воду
В средней полосе России на сухих, сильно прогреваемых склонах растет небольшое растение очиток едкий (рис. 2), а в сосняках на сухой песчаной почве — молодило побегоносное. Эти листовые суккуленты, чтобы высушить для гербария, предварительно ошпаривают кипятком, иначе живые листья будут прочно удерживать воду и не засохнут. У этих растений при сушке могут раскрыться и зацвести бутоны
Расход воды растениями. Поступившая в растения вода в процессе жизнедеятельности непрерывно расходуется, причем больше всего — на испарение. Взрослая береза за сутки испаряет в среднем 75 л воды, бук — 100, а липа — 200 л. Если большое ведро воды вмещает 10 литров, то, значит, эти деревья ежедневно испаряют от 7 до 20 ведер воды.
Вода расходуется на процесс фотосинтеза, на рост растений. Много воды потребляют растения, когда у них, как говорят, наливаются плоды. Это относится не только к таким растениям, как яблоня, груша, виноград, но и к хлебным злакам.
Источники информации: А.М. Былова, Н.И. Шорина, 1999 (209стр.)
ВНЕКОРНЕВЫЕ ПОДКОРМКИ-ПОЧЕМУ ОНИ НЕОБХОДИМЫ?
Эпифиты
Воздушные корни, размножение!"
Черенкование (воздушные корни) Шалфея
Суккуленты давайте разберемся какие они бывают
Корень
xn--j1ahfl.xn--p1ai
| Вода, поступившая в клетки корня под влиянием разности водных потенциалов, которые возникают благодаря транспирации и корневого давления, передвигается до проводящих элементов ксилемы. Согласно современным представлениям, вода в корневой системе может перемещаться в радиальном направлении тремя путями: апопластическим, симпластическим, трансмембранным. Еще в 1932 г. немецкий физиолог Э. Мюнх высказал мнение о существовании в корневой системе двух относительно независимых друг от друга объемов, по которым передвигается вода,— апопласта и симпласта. При транспорте по апопласту вода передвигается по клеточным стенкам, не проходя через мембраны. При симпластном транспорте вода проникает в клетку через полупроницаемую мембрану и далее перемещается по протопластам клеток, которые соединены между собой многочисленными плазмодесмами. При трансмембранном транспорте вода перетекает через клетки и при этом проходит, по крайней мере, две плазматические мембраны. Уже обсуждалось, что в последнее время много внимания уделяется аквапоринам — мембранным белкам, образующим в мембранах специализированные водные каналы и определяющим проницаемость для воды. Эксперименты показали, что передвижение воды по коре корня идет главным образом по апопласту, где она встречает меньшее сопротивление, и лишь частично по симпласту (С. Френч). Апопластный путь прерывается в эндодерме в связи с наличием поясков Каспари. Вместе с тем в апикальной части суберинизация отсутствует, поэтому вода легко проникает через эндодерму. Кроме того, в суберинизированных частях корня вода может проходить через пропускные клетки. Сказанное показывает, что для транспорта в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану клеток эндодермы. Таким образом, мы имеем дело как бы с осмометром, у которого полупроницаемая мембрана расположена в клетках эндодермы. Вода устремляется через эту мембрану. В сторону меньшего (более отрицательного) водного потенциала. Далее вода поступает в сосуды ксилемы. По вопросу о причинах, вызывающих секрецию воды в сосуды ксилемы, имеются различные суждения. Согласно гипотезе Крафтса, это следствие выброса солей в сосуды ксилемы, в результате чего там создается повышенная их концентрация, и водный потенциал становится более отрицательным. Предполагается, что в результате активного поступления соли накапливаются в клетках корня. Однако интенсивность дыхания в клетках, окружающих сосуды ксилемы (перицикл), очень низкая, и они не удерживают соли, которые благодаря этому десорбируются в сосуды. Транспорт воды в корне зависит от интенсивности процесса дыхания. При помещении растений в условия, тормозящие дыхание корней (низкая температура, анаэробиоз или наличие дыхательных ядов), они транспортируют меньше воды. Предполагают, что это может быть связано с инактивированием аквапоринов. Торможение транспорта воды в корнях в аэробных условиях, возможно, объясняет факт завядания растений в переувлажненной почве. Дальнейшее передвижение воды идет по сосудистой системе корня, стебля и листа. Проводящие элементы ксилемы состоят из сосудов и трахеид. Опыты с кольцеванием показали, что восходящий ток воды по растению движется в основном по ксилеме. В проводящих элементах ксилемы вода встречает незначительное сопротивление, что, естественно, облегчает передвижение воды на большие расстояния. Правда, некоторое количество воды передвигается и вне сосудистой системы. Однако по сравнению с ксилемой сопротивление движению воды других тканей значительно больше. Это приводит к тому, что вне ксилемы движется всего от 1 до 10% общего потока воды. Из сосудов стебля вода попадает в сосуды листа. Вода движется из стебля через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся все более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Именно поэтому густота жилкования листа считается одним из важнейших признаков ксероморфной структуры — отличительной чертой растений, устойчивых к засухе. Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. В связи с этим, как только в силу процесса транспирации возникает ненасыщенность водой клеточных стенок паренхимных клеток, она сейчас же передается внутрь клетки, водный потенциал которой падает. Вода передвигается от клетки к клетке благодаря градиенту водного потенциала. По-видимому, передвижение воды от клетки к клетке в листовой паренхиме идет не по симпласту, а в основном по клеточным стенкам, где сопротивление значительно меньше. По сосудам вода движется благодаря создающемуся в силу транспирации градиенту водного потенциала, градиенту свободной энергии (от системы с большей свободой энергии к системе с меньшей). Можно привести примерное распределение водных потенциалов, которое и вызывает передвижение воды: водный потенциал почвы (—0,5 бар), корня (—2 бар), стебля (—5 бар), листьев (—15 бар), воздуха при относительной влажности 50% (-1000 бар). Однако получены экспериментальные данные, которые не позволяют рассматривать силу транспирации как единственную, обуславливающую восходящий ток воды по растению. Так, показано, что восходящий ток воды может осуществляться и при отсутствии транспирации. К этому же выводу приводят опыты, показывающие ритмическое секретирование устьичными клетками жидкой воды, а также зависимость передвижения воды от эндогенной энергии, поставляемых процессом дыхания. Это позволяет считать, что движущая сила транспорта воды в растении является суммой двух весьма различных по своей природе составляющих, условно названных метаболической и осмотической. Об этом уже упоминалось при рассмотрении вопроса о корневом давлении. Осмотическая составляющая представлена в корнях сугубо осмотическими явлениями, в стебле и листьях — гидростатической тягой, создаваемой градиентом водного потенциала в системе почва — растение — атмосфера. Температурный коэффициент Q10 этой составляющей близок к единице. Именно такая величина Q10 характерна для простых физических процессов. В то же время Q10 процесса транспорта воды значительно выше. Столь высокий температурный коэффициент свойственен сложным процессам цепного характера. Очевидно, транспорт воды за счет метаболической составляющей относится к разряду именно таких процессов. Согласно имеющимся экспериментальным данным, в формировании метаболической составляющей непосредственное участие принимают контрактильные системы паренхимных клеток (т. е. белки цитоскелета). Они могут играть роль сфинктеров, регулирующих просветы водных каналов (открывание—закрывание) в плазмодесмах. Благодаря их работе индуцируются ритмические микроколебания внутриклеточного (внутрисимпластного) гидростатического давления, которые внешне выражаются в короткопериодных (1—3 мин) автоколебаниях поступления воды в клетку (симпласт, ткань, орган) и водоотдачи (т. е. микропульсациях). Принципиально важное значение имеет факт противофазное автоколебаний водопоглощения и водоотдачи. Это свидетельствует о том, что процесс транспорта воды состоит из двух последовательных, ритмично чередующихся, относительно самостоятельных, хотя и тесно взаимосвязанных фаз: фазы сокращения, во время которой вода выделяется (выталкивается, секретируется) и последующей фазы расслабления, во время которой вода поглощается. Выделение воды вызывает падение тургора и водного потенциала в целом, создавая предпосылку для поглощения следующей порции воды, вновь приводящего к возрастанию водного потенциала вплоть до того, что он из отрицательного становится положительным. После этого происходит новое сокращение. Именно фаза сокращения происходит с участием контрактильных систем и требует затраты энергии. Таким образом, вода поглощается и выделяется по градиенту водного потенциала, а не против него, т. е. согласно данной схеме, транспорт воды в термодинамическом понимании является пассивным. Возникающие за счет ритмической деятельности внутриклеточного сократительного аппарата микроколебания гидростатического давления паренхимных клеток являются механизмом, создающим локальные градиенты водного потенциала на пути водного тока и тем самым регулирующим скорость этого тока. Именно благодаря этому формируется метаболическая составляющая движущей силы транспорта воды в растении, играющая решающую роль в общей системе эндогенной регуляции. Под влиянием ингибиторов контрактильных систем или окислительного фосфорилирования (т. е. при нарушении энергоснабжения) противофазность исчезает, автоколебания затухают и транспорт воды тормозится (В.Н. Жолкевич). Необходимо также отметить роль сил сцепления для обеспечения поднятия воды в растении на высоту более 100 м. Теория сцепления была выдвинута русским ученым Е.Ф. Вотчалом и английским физиологом Е. Диксоном. Для лучшего понимания рассмотрим следующий опыт. В чашку с ртутью помещают заполненную водой трубку, которая заканчивается воронкой из пористого фарфора. Вся система лишена пузырьков воздуха. По мере испарения воды ртуть поднимается по трубке. При этом высота подъема ртути превышает 760 мм. Это объясняется наличием сил сцепления между молекулами воды и ртути, которые в полной мере проявляются при отсутствии воздуха. Сходное положение, только еще более ярко выраженное, имеется в сосудах у растений. Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту, значительно большую 10 м. Расчеты показали, что благодаря наличию сродства между молекулами воды силы сцепления достигают величины -30 бар. Это такая сила, которая позволяет поднять воду на высоту, равную 120 м, без разрыва водных нитей, что примерно составляет максимальную высоту деревьев. Силы сцепления существуют и между водой и стенками сосудов — адгезия. Стенки проводящих элементов ксилемы эластичны. В силу этих двух обстоятельств даже при недостатке воды связь между молекулами воды и стенками сосудов не нарушается. Это подтверждается исследованиями по изменению толщины стебля травянистых растений. Определения показали, что в полуденные часы толщина стебля растений уменьшается. Если перерезать стебель, то сосуды сразу расширяются и воздух поступает в них. Из этого опыта видно, что при сильном испарении сосуды сужаются и это приводит к появлению отрицательного давления. Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды. Таким образом, при нормальном водоснабжении создается непрерывность воды в системе почва — растение — атмосфера. В том случае, если в отдельные членики сосудов попадает воздух (эмболия), они, по-видимому, выключаются из общего тока проведения воды. Таков путь воды по растению и его основные движущие силы. Современные методы исследования позволяют определить скорость передвижения воды по растению. Скорость передвижения воды определяется разностью водных потенциалов в начале и конце пути, а также сопротивлением, которое она встречает. Скорость движения воды в течение суток изменяется и в дневные часы она значительно больше. При этом разные виды растений отличаются по скорости передвижения воды. Если скорость передвижения у хвойных пород обычно 0,5—1,2 м/ч, то у лиственных она значительно выше. У дуба, например, скорость передвижения составляет 27— 40 м/ч. Скорость передвижения воды мало зависит от напряженности обмена веществ. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды. Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды скорее могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды. |
fizrast.ru
Поступление воды в растение, корневая система как орган поглощения воды; корневое давление, его значение и зависимость от действия внешних факторов.
Поступление воды через корневую систему сокращается с понижением температуры. Это происходит по следующим причинам: 1) повышается вязкость воды, и поэтому снижается ее подвижность; 2) уменьшается проницаемость протоплазмы для воды; 3) тормозится рост корней; 4) уменьшается скорость метаболических процессов. Поступление воды снижается при ухудшении аэрации почвы. Это можно наблюдать, когда после сильного дождя почва залита водой, но при ярком солнце из-за сильного испарения рас-
тения завядают. Большое значение имеет концентрация почвенного раствора. Вода поступает в корень только тогда, когда водный потенциал корня меньше водного потенциала почвы. Если почвенный раствор имеет более отрицательный потенциал, вода будет не поступать в корень, а выходить из него.
Для нормального функционирования клетки растения должны быть насыщены водой. Состояние насыщения поддерживается с помощью двух взаимосвязанных процессов: поступления и выделения (расходования) воды, которые и составляют водный обмен растений. Соотношение между этими процессами называют водным балансом.
Растение выделяет воду в жидком и парообразном состояниях. Физиологический процесс испарения воды наземными органами растений получил, как уже отмечалось, название транспирации, выделение воды в капельножидком состоянии – гуттации.
Жизнь возникла в мировом океане. С выходом растений на сушу появилась опасность их гибели от высыхания. Почему? Концентрация водяных паров в воздухе, даже в условиях влажного климата, всегда меньше, чем в межклетниках листа, поэтому неизбежно происходит движение воды из тела растения в окружающую среду, при этом диффузия молекул воды от поверхности листа в воздух происходит в 1 500 раз быстрее, чем их поступление из почвы в корень.
Большая потеря воды растением обусловлена ее большой листовой поверхностью. Одно растение кукурузы расходует за вегетативный период 200 литров воды, а сахарный тростник – в два раза больше. Растения вынуждены формировать большую листовую поверхность, чтобы получать необходимое количество СО2 для фотосинтеза, несмотря на его малое содержание (0,045 %) в атмосфере.
Например, 8-летняя яблоня может ассимилировать за день до 50 г СО2 при интенсивном фотосинтезе. Это количество СО2 она поглощает из 300 000 литров воздуха.
Необходимость расходовать воду в больших количествах, связано еще с тем, что поглощение солнечных лучей растениями должно привести к повышению температуры, которая может вызвать коагуляцию белков. Испаряя воду, растение понижает температуру своего тела.
Для большинства растений подсыхание смертельно, поэтому расход воды должен соотноситься с приходом. Только мхи и лишайники могут выдерживать длительное отсутствие воды и переносить это время в состоянии высыхания.
Поэтому, как только растения вышли из воды на сушу, начался долгий процесс по выработке приспособлений для поддержания водного баланса: во-первых, для быстрого поступления, во-вторых, для уменьшения траты наземными органами; в-третьих, для экономически выгодного транспорта по стеблю.
При анализе соотношения между поступлением и расходованием воды возможны три случая: поступление больше расходования, равно или меньше. В последнем случае возникает водный дефицит. В полдень водный дефицит может достигать 5–10 и даже 25 %. Одно из условий нормального функционирования наземных растений – поддержание условий без длительного и глубокого водного дефицита. Для этого необходима хорошо развитая корневая система, обеспечивающая поступление воды с большой скоростью.
Поступление воды из почвы в корень. Поступление воды из почвы представляет собой более сложный процесс, чем простое всасывание воды растением, опущенным в какой-либо водный раствор корнями. В почве мы имеем ряд сил, противодействующих этому всасыванию, которые можно назвать водоудерживающими силами.
Почва – многофазная система, состоящая из четырех основных компонентов: твердых минеральных частиц, органического вещества (гумуса), почвенного раствора и почвенного воздуха.
Минеральные частицы и гумус образуют почвенную структуру, вода и воздух заполняют полости этой структуры.
Способность почвы удерживать воду зависит от ее состава и свойств. Определенное количество воды входит в состав минеральных компонентов почвы и недоступна растениям (гигроскопическая вода). Разнообразные глинистые минералы и гетерогенные гумусовые вещества, которые представляют собой коллоиды, могут также удерживать значительное количество гидратационной воды. Такая вода условно называется связанной (клеточной) и тоже труднодоступна. Вода, находящаяся в капиллярах почвы (капиллярная вода), легко всасывается корневыми волосками и поверхностными клетками сосущей зоны корня. Такая вода считается свободной.
Кроме того, в почве мы имеем дело с раствором, а не с чистой водой; поэтому сам раствор имеет сосущую силу. При равновесии концентраций в почве и в сосудах ксилемы в общем случае поступление воды должно остановиться.
Чтобы вычислить количество недоступной почвенной влаги поступают следующим образом. Растения выращивают в сосуде с непроницаемыми для воды стенками (стеклянный или металлический) и после того, как растения хорошо разовьются, почву перестают поливать и оставляют в притемненном месте до завядания растения. Завядание указывает, что доставка воды в корни прекратилась. То количество воды, которое остается в почве к моменту завядания, и будет недоступной для растения (мертвый запас). Количество недоступной воды получило название коэффициента завядания или влажности завядания. Следовательно, коэффициент завядания характеризует влажность, при которой устойчивое завядание только начинается.
В этот момент в почве имеется еще некоторое количество воды, хоть и малое, но в какой-то степени доступное для растения. В этих условиях даже сильно подвядшее растение оживает, если его начать поливать. Полная гибель наступает только при влажности значительно ниже, чем коэффициент завядания.
Таким образом, влажность завядания представляет собой нижнею границу того интервала влажности, при котором возможен рост растения.
При поступлении воды в сухую почву она впитывается очень быстро. Затем, скорость просачивания воды в нижние горизонты становится медленной. Когда скорость нисходящего тока воды резко понижается, влажность почвы достигает уровня, который называется полевой влагоемкостью.
Под доступной для растения почвенной влагой имеется ввиду количество воды, которое накапливается в почве от уровня влажности устойчивого завядания до полевой влагоемкости.
Радиальный транспорт воды.Чтобы перейти к разговору о радиальном транспорте воды, необходимо коротко вспомнить общие черты строения корней, (хотя в деталях строение корней в разных видах растений варьирует).
В обоих типах растений имеется центральный цилиндр (стела), в котором находятся сосудистые пучки и (стелярная) паренхима. При помощи сосудистых пучков (флоэма) поддерживается связь между разными частями растения и местами синтеза углеводов. По сосудистым пучкам (ксилема) поступает из корня в побег вода и растворенные вещества. Если перемещаться к периферии, то далее идет перецикл – однорядный слой клеток. Затем идет эндодерма – тоже однорядный слой клеток, особенно хорошо приспособленный к тому, чтобы служить барьером, который отделяет проводящие ткани от коры. После того, как растяжение клеток заканчивается, в радиальных и поперечных оболочках эндодермы выявляется прослойка материала, которая дает положительную реакцию на лигнин. Это так называемый поясок Каспари. Пояски Каспари сильно ограничивают движение воды, ионов в клеточных оболочках. Таким образом, они эффективно блокируют апопластический транспорт.
Большую часть корней занимают клетки коры. На их долю (вместе с эпидермисом) приходится 86–90 % площади. Клетки коры вытянуты параллельно главной оси, тонкий слой цитоплазмы (1–5 мкм) окружает в них центральную вакуоль, которая занимает ~ 90 % объема протопласта. Вторичные изменения в оболочках клетки коры сводятся к отложению целлюлозы; только у некоторых видов происходит одревеснение. Вообще, оболочки сохраняют проницаемость. В них имеются многочисленные поры, которые могут быть заполнены в зависимости от условий воздухом или водой.
По периферии корня расположен эпидермис (ризодермис). Это наружная ткань корня, состоящая из вытянутых плотно упакованных клеток. Оболочки этих клеток могут со временем претерпевать изменения, связанные с отложением кутина – водоотталкивающего вещества.
Внешние тангенциальные оболочки клеток ризодермиса способны сильно растягиваться и образовывать трубковидные выросты, так называемые корневые волоски. Отметим, что клетки, которые могут образовывать корневые волоски имеют название трихобластов, а которые не способны к образованию – атрихобластов.
Итак, вода, попавшая в корневой волосок или другую клетку корня с помощью одного из механизмов, о которых мы говорили, движется к сосудам ксилемы, т. е. происходит радиальное перемещение воды по тканям корня.
Движение воды в корневой волосок или другую клетку начинается с взаимодействия с оболочкой клеток, затем вода может пройти через плазмалемму и транспортироваться из протопласта одной клетки в протопласт другой через плазмодесмы. В этом случае вода движется по симпласту. Однако вода могла бы и не заходить в симпласт, а оставаться в клеточной оболочке и двигаться по ним через ткани корня к центральному цилиндру. Это апопластический путь. Но по апопласту вода может двигаться только до эндодермы (до поясков Каспари). Поэтому путь воды по апопласту тут прекращается. Для дальнейшего передвижения она должна обязательно войти в симпласт.
Первым барьерную функцию эндодермы выявил Д. Пристли в 1920 г. В последнее время эта функция была неоднократно подтверждена.
Необходимость переключения движения воды с апопластного на симпластный путь имеет большое значение, так как процесс движения по живой протоплазме клетка может сама регулировать в отличие от перемещения воды по клеточным стенкам.
Для поступления воды в корень необходимо чтобы водный потенциал ризодермиса был меньше, чем водный потенциал почвенного раствора; в этом случае вода начнет поступать в клетки.
Почву можно рассматривать как резервуар, количество воды, в котором то увеличивается, то уменьшается. Если после дождя почва находится в состоянии полевой влагоемкости, то ее водный потенциал близок к нулю; вода легко поступает в корни. По мере подсыхания почвы ее водный потенциал снижается.
Водный потенциал максимален в почве, несколько меньший в клетках корня и самый низкий в клетках, прилегающих к эпидермису листа.
Первая причина возникновения градиента водного потенциала – активное поступление солей и их активное перемещение из одной клетки в другую. Вторая – живые клетки паренхимы центрального цилиндра выделяют растворимые органические и минеральные вещества в сосуды и таким образом поддерживают градиент водного потенциала в сосудах. Поступая в сосуды ксилемы, они образуют тут осмотический градиент.
Химический потенциал воды, в которой находятся эти ионы ниже потенциала воды в разбавленном солевом растворе внешней среды. Поэтому вода должна перемещаться по градиенту химического потенциала в ксилему корня.
Сегодня, выдвинута гипотеза, которая имеет под собой экспериментальную основу, что в корнях есть специализированный механизм активного накачивания воды (водная помпа), работа которой не зависит от поступления ионов.
Действительно, для поступления воды в корень необходима энергия, поэтому этот процесс зависит от скорости аэробного дыхания корней – основного источника АТФ. Таким образом, одним из главных факторов, обуславливающих поступление воды, является О2. Отсюда условия, способные подавить дыхание, резко уменьшают поступление воды. В качестве примера рассмотрим следующее явление: прошел сильный дождь, в низком месте собралось много воды, а растение завядает. Почему? Избыток воды в почве вытеснил из нее воздух, поступление кислорода в корень затруднено, дыхание подавлено. Сразу же наблюдается торможение поступления воды в корневую систему. Этот пример объясняет, почему на затопленных водой почвах растения развиваются плохо и даже гибнут.
Затопление приводит не только к уменьшению количества О2, но и к увеличению концентрации СО2 в почве, который повреждает мембраны корневых волосков; наблюдается снижение поступления воды, что подтверждается торможением выделения пасоки.
Поступление воды в корень зависит и от температуры почвы. Например, на холодных болотистых почвах, несмотря на большое количество воды, растения испытывают недостаток воды, поскольку при низких температурах подавляется дыхание и нарушается снабжение корней энергией. У растений в этих условиях начинает формироваться ксероморфная структура (мелкие клетки, много устьиц и т. д.), которая характерна для сухих мест.
Состояние растений, при котором вода не может поступать, несмотря на ее большое количество в окружающей среде, получило название физиологической засухи.
Различные полютанты также могут влиять на поступление воды, в частности через подавление дыхания.
Необходимость поглощать большое количество воды даже в условиях ее лимита, например, в условиях засухи, приводит к тому, что растение формирует огромную корневую систему. В результате корни проникают в почву на большую глубину. У пшеницы длина корней достигает 90 см, у люцерны – 120 см. Если считать, что глубина пахотного горизонта составляет 20–25 см, то большая часть корневой системы размещается ниже. Размер корневой системы характеризуется не только глубиной их проникновения в почву. Большое значение имеет общая поверхность.
Вода в почве двигается очень медленно: на протяжении месяца она продиффундирует не больше, чем на 30 см. Перемещение кончика корня в почве опережает движение воды. Таким образом, не вода движется к корню, а корень к воде в процессе роста. Рост является первой и важнейшей особенностью корней как органа, поглощающего воду.
В засушливых условиях формируется корневая система в 3–4 раза большая, чем во влажных.
Разветвление и быстрый рост помогают корню двигаться к воде, но, с другой стороны, вода – необходимое условие роста. Как видно уже не на клеточном, а на организменном уровне, мы сталкиваемся с примером обратной связи, которая лежит в основе регуляторных процессов.
Как орган, поглощающий воду, корень владеет еще одним важным свойством – положительным гидротропизмом, это значит, что при недостатке воды растущие части корней выгибаются в стороны более влажных участков почвы.
Таким образом, корневая система представляет собой специализированный орган поступления воды.
Рост корней обычно опережает рост наземных органов. Это очень важная особенность, связанная с тем, что корень должен обеспечить потребности в воде формирующего растения.
Однако клетка любого органа, которая не насыщена водой, тоже может поглощать воду, как только она будет приведена с ней в соприкосновение. Поэтому и листья, особенно подвядшие, при погружении в воду довольно энергично ее всасывают; с другой стороны, несмотря на кутикулу, вода может поступать через поверхность листа. Показано, что только сухая кутикула почти непроницаема для воды; при смачивании она набухает и делается проницаемой, поэтому смоченные дождем или росой листья могут поглощать до 25 % падающей на них воды. Это имеет практическое значение при орошении растений дождеванием.
Дальний транспорт воды.Говоря о перемещении воды по растению выделяют транспорт в тканях одного органа, который называют ближним (радиальный), и транспорт между отдельными органами, который называют дальним. Между ними есть существенная разница. Ближний транспорт идет по неспециализированным тканям, а для дальнего в растениях имеются специальные проводящие ткани. Таким образом, путь, который проходит вода от корневого волоска до испаряющей клетки листа, распадается на две части: различные по протяженности, строению и физиологическим признакам. Первая часть состоит из живых клеток и имеет малые размеры (миллиметры или доли миллиметра). Это два коротких участка – один – в корне, от его поверхности с корневыми волосками до сосудов, которые находятся в его центральном цилиндре; второй – в листе, от сосудов, входящих в состав проводящего пучка, и испаряющей воду в межклетники хлоренхимы. Вторая часть пути – это сосуды, трахеиды, которые представляют собой мертвые трубки. У травянистых растений их длина составляет несколько сантиметров, а у деревьев достигает нескольких метров и даже десятков метров.
Вода и минеральные элементы доставляются к каждой клетки надземной части растения благодаря восходящему току по ксилеме. Существует также нисходящий флоэмный ток растворов от листьев к корням. Направленный вниз флоэмный ток формируется в клетках мезофилла листьев, где часть воды, которая пришла с ксилемным током, с клеточных оболочек мезофилла переходит во флоэмные окончания.
Вода с клеток листа и непосредственно с сосудов ксилемы поступает во флоэму по осмотическому градиенту, возникающему из-за накопленных в клетках флоэмы сахаров и других органических соединений, образующихся в процессе фотосинтеза.
Нисходящий флоэмный ток доставляет органические вещества тканям корня, где они используются в метаболизме. В корнях окончания проводящих пучков элементов флоэмы, как и в листе, расположены вблизи элементов ксилемы, и вода вновь по осмотическому градиенту поступает в ксилему и движется вверх. Таким образом, происходит обмен воды в проводящей системе корней и листьев (как бы круговорот).
Ток воды по сосудам ксилемы приводит к тому, что при перерезании стебля какого-нибудь растения на небольшом расстоянии от почвы через некоторое время с конца сосудов начинает выделяться сок, который называют пасокой. Это явление получило название «плача растений».
Силу, которая подымает пасоку вверх по сосудам, назвали корневым давлением. Корневое давление можно измерить если надетую на перерезанный стебель трубку соединить с манометром. Величина корневого давления непостоянна. В оптимальных условиях она составляет 2–3 бара. При определенных условиях достигается равновесие между количеством выделенной пасоки и количеством поступившей воды, поэтому корневое давление, или количество выделенной пасоки, может отражать поглотительную способность корней. Таким образом, активными двигателями начального восходящего водного тока (корневого давления) являются живые клетки, которые прилегают к нижнему концу проводящей системы растений – это клетки паренхимы корней – нижний концевой двигатель водного потока.
Механизм корневого давления, как считают, основывается на действии сократительных белков, функцию которых, как считают, выполняют микрофибриллы Ф-белков.
В какой-то степени доказательством активного выхода воды может служить гуттация.
Однако если бы растение постоянно не теряло воду в результате транспирации, то клетки корневых волосков быстро бы насытились водой, и ее поступление прекратилось. Поэтому, одной из причин возникновения градиента водного потенциала – испарение воды надземными органами.
Чем интенсивней клетки листьев испаряют воду, тем быстрее она будет поступать в клетки корней и быстрей транспортироваться вверх по растению. Потеря молекул воды в верхней части водного столба в результате испарения заставляет воду течь по сосудам ксилемы вверх для ликвидации потери. Это вызванное транспирацией движение воды получило название транспирационного тока. Он, в свою очередь, обуславливает поступление воды из почвы в растение то же по градиенту водного потенциала. Из-за транспирации водный потенциал в верхней части растения ниже, чем у основания.
Активными двигателями водного тока, обусловленного транспирацией, являются живые клетки, которые прилегают к верхнему концу всей проводящей системы растения – клетки паренхимы листа. Они были названы верхним концевым двигателем водного тока.
Механизм работы верхнего концевого двигателя несложный и основан на следующем. Атмосфера обычно недонасыщена водяными парами, поэтому имеет отрицательный водный потенциал. При относительной влажности воздуха 90 % он составляет 140 бар. У большинства растений водный потенциал листьев колеблется от 1 до 30 бар.
По причине большой разности водных потенциалов происходит транспирация. Уменьшение количества воды в паренхимной клетке листа вызывает снижение активности воды в ней и уменьшение водного потенциала.
Водный дефицит постепенно от клетки к клетке достигает корней, и активность воды в них снижается. В этом случае вода и поступает из почвы в корень. Таким образом, можно сделать вывод, что перемещение воды по растению, как и поступление ее в корень, главным образом, связано с градиентом водного потенциала в системе почва-растение-воздух. Этот градиент будет тем больше, чем больше воды будут терять клетки листа, т. е. чем сильней транспирация.
Работают два двигателя неодинаково. В среднем верхний концевой двигатель развивает силу в 10–15 бар и даже больше, а нижний 2–3 бара. Отсюда видно, что главная роль в водном обмене принадлежит верхнему двигателю. Однако, при отсутствие листьев у деревьев зимой и ранней весной, или после сухого периода главную роль в передвижении воды выполняет нижний двигатель. Большую роль в поднятии воды по растению нижний концевой двигатель должен иметь в условиях большей влажности воздуха, когда транспирация минимальна.
Для верхнего концевого двигателя источник энергии – солнце, это означает, что поглощаемая листом лучистая энергия используется для испарения.
Для нижнего концевого двигателя – источник энергии дыхание. Энергия молекул АТФ, синтезируемая во время дыхания клеток корня, расходуется на транспорт ионов в клетке, т. е. на создание градиента водного потенциала. Регуляторная роль корневого давления в водообмене растений схематически представлена на рис. 4.12.
Таким образом, верхний концевой двигатель представляет собой автоматически работающий механизм, который тем сильней присасывает воду, чем быстрей ее расходует.
Работой верхнего и нижнего концевых двигателей без труда можно объяснить поднятие воды на несколько десятков сантиметров, пусть метров. А как объяснить поднятие воды на десятки метров, а секвойя достигает высоты 140 метров? Сосуды, по которым двигается вода на большей части своего пути, представляют собой мертвые трубки. Они не могут развивать силы для поднятия воды. Ответить на этот вопрос помогает теория сцепления, которую предложил английский исследователь Г. Диксон в 1921 году. В соответствие с этой теорией в сосудах образуется непрерывные нити, проходящие от клеток паренхимы корня до клеток паренхимы листьев. Сила, которая заставляет молекулы воды идти друг за другом, была названа силой сцепления (когезия). Непрерывные водяные нити образуются за счет водородных связей. Однако, водяные нити сцеплены и со стенками сосудов (адгезия) с силой 300–350 бар. Все это позволяет нижнему и верхнему концевым двигателям поднимать воду по стволу на высоту 140 м.
После появления этой теории анатомы не раз обращали внимание исследователей на образование пузырьков воздуха, которые должны нарушать сцепление между молекулами воды в сосудах. Однако в случае временного исключения какого-нибудь сосуда вода движется по запасным путям (другим сосудам) или апопласту, а воздушные пузырьки постепенно рассасываются при участии живых клеток.
Передвижение воды из корня в лист по мертвым сосудам, оказывающим минимальное сопротивление водному току, представляет собой одну из находок природы, которая заключается в следующем. Клетки сосудов и трахеид вытянуты в длину, в них отсутствует живое содержимое, внутри они пустые, т. е. они представляют собой простые трубки. Одревесневшие вторичные клеточные оболочки достаточно крепкие на разрыв, способные выдержать большую разность давлений, возникающих при подъеме воды к вершине больших деревьев. Торцевые, а иногда и боковые стенки члеников сосудов, перфорированы; сосуды, которые состоят из соединенных концами члеников, образуют длинные трубки, по которым вода с минеральными веществами легко проходит. В трахеидах нет перфораций, и вода, чтобы попасть из одной трахеиды в другую, должна пройти через их торцевые стенки; но трахеиды очень длинные клетки, а поэтому и это строение тоже очень хорошо приспособлено для проведения воды.
Выход в процессе эволюции растений на сушу, крона которых располагается довольно далеко от земли, стал возможным благодаря образованию высокоспециализированной проводящей системы. Значение этого приспособления подчеркивается и названием самих растений – сосудистые.
Кроме разницы в механизмах действия существует полная согласованность в работе двух концевых двигателей. На действие любого фактора среды, способного подавить работу нижнего двигателя, лист отвечает активацией транспирации, и наоборот. Это биологически очень важное приспособление, хотя на первый взгляд, оно выглядит парадоксально: поступление воды ухудшается, а лист на этот неблагоприятный фактор отвечает не подавлением, а, наоборот, увеличением транспирации. Увеличение транспирации в данном случае имеет своей целью стимулировать поступление воды в корень.
Сейчас существует взгляд, что в растении имеется особая регуляторная система – гидродинамическая. Под ее контролем находятся водный режим, поддержание водного гомеостаза, а также, некоторые другие функции, в частности, фотосинтез. Гидродинамическая регуляторная система очень чувствительна. Она приходит в действие при очень маленькой потере воды листом (0,06 % от исходного количества) и предотвращает более сильное обезвоживание в дальнейшем.
Передача сигнала к листу происходит через сплошной водный поток, а восприятие – устичным и фотосинтетическим аппаратом. Рецептором сдвигов почвенных условий, тормозящих поступление воды, являются, скорее всего, мембраны эндодермальных клеток корня.
Гидродинамическая регуляторная система позволяет растению очень быстро реагировать на внешние изменения, потенциально неблагоприятные для водного гомеостаза.
Наземные растения стоят перед сложной дилеммой: с одной стороны, они должны обладать достаточно развитой поверхностью, чтобы эффективно поглощать солнечный свет и СО2, а с другой стороны, по мере увеличения поверхности увеличиваются потери воды. Эту проблему растения решают разными способами: во-первых, поступление воды увеличивается за счет роста корней и развития гипертрофированной поглотительной поверхности. Во-вторых, потеря воды становится медленнее из-за того, что мезофильные клетки отделены от окружающей среды кутикулой, содержащей воск. В-третьих, противоречие между необходимостью поглощать большее количество СО2 и одновременно уменьшать количество испаряемой воды растения решают с помощью осциляторного механизма.
Пути ближнего и дальнего транспорта, механизм передвижения воды по растению (градиент водного потенциала, движущие силы восходящего тока воды в растении, верхний и нижний концевые двигатели, процессы адгезии и когезии).
Ближний транспорт — это передвижение ионов, метаболитов и воды между клетками и тканями (в отличие от мембранного транспорта в каждой клетке). Дальний транспорт — передвижение веществ между органами в целом растении.
Транспорт веществ в растении осуществляется по любым тканям и по проводящим пучкам, специализированным для этой цели. В свою очередь передвижение воды и растворенных веществ по любым тканям может происходить: а) по клеточным стенкам, т. е. по апопласту, б) по цитоплазме клеток, соединенных друг с другом плазмодесмами, т. е. по симпласту, в) возможно, по эндоплазматическому ретикулуму с участием плазмодесм.
Передвижение воды и веществ по проводящим пучкам включает в себя транспорт по ксилеме («восходящий ток» — от корней к органам побега) и по флоэме («нисходящий ток» — от листьев к зонам потребления питательных веществ или отложения их в запас). По флоэме транспортируются метаболиты и при мобилизации запасных веществ.
Радиальный транспорт. Путем диффузии и обменных процессов ионы поступают в клеточные стенки ризодермы и затем через коровую паренхиму перемещаются к проводящим пучкам. Это передвижение происходит как по клеточным стенкам — апопласту, так и по симпласту.
Перемещение ионов по апопласту происходит за счет диффузии и обменной адсорбции по градиенту концентрации и ускоряется током воды. Движение минеральных веществ по симпласту осуществляется благодаря движению цитоплазмы, а между клетками — по плазмодесмам. Направленному движению по симпласту способствуют градиенты концентрации веществ. Эти градиенты возникают вследствие того, что поступившие в клетку вещества включаются в процессы метаболизма и концентрация их снижается.
Диффузия ионов и молекул по кажущемуся свободному пространству клеток прерывается на уровне эндодермы. Единственный путь дальнейшего передвижения веществ через эндодерму — транспорт по симпласту, что обеспечивается метаболическим контролем поступления веществ. Существование в эндодерме пропускных клеток, в которых пояски Каспари недоразвиты или отсутствуют, позволяет незначительной части поглощенных веществ избежать метаболического контроля.
Симпластический транспорт является основным для многих ионов. При этом активной метаболизации подвергаются соединения, содержащие азот, углерод, фосфор, в меньшей степени — серу, кальций, хлор. Другие ионы метаболическому контролю практически не подвергаются.
Существенную роль в симпластическом транспорте веществ играют вакуоли. В определенной степени они конкурируют с сосудами ксилемы за поглощенные вещества и таким образом выполняют роль регулятора поступления веществ в сосуды. Этот процесс зависит от степени насыщения вакуолярного сока растворенными веществами. При снижении концентрации веществ в цитоплазме они могут вновь выходить из вакуолей, представляя, таким образом, запасной фонд питательных веществ. Поглощение ионов вакуолями снижает концентрацию их в симпласте и обеспечивает создание градиента концентрации, необходимого для транспорта их по симпласту.
Каким образом ионы поступают в мертвые сосуды ксилемы, т. е. как осуществляется ее загрузка?
Ксилемный сок представляет собой раствор, в основном состоящий из неорганических веществ. Однако в пасоке, вытекающей из ксилемы пенька при удалении верхней час стебля, обнаружены также различные азотистые соединен (аминокислоты, амиды, алкалоиды и др.), органические кислоты, фосфорорганические эфиры, соединения, содержащие серу, некоторое количество сахаров и многоатомных спиртов а также фитогормоны. В ксилемном соке могут содержать и более сложные вещества, попадающие сюда из вакуолей и цитоплазмы трахеальных элементов, заканчивающих свое развитие.
Ксилемный сок по составу резко отличается от вакуолярго. Например, содержание иона К+ в вакуолях эпикотиля гороха достигает 55 — 78 ммоль/л, а в ксилемном соке — лишь 2—4 ммоль/л.
Загрузка ксилемы наиболее интенсивно происходит в зоне корневых волосков. В этой зоне функционируют один или два насоса. Основной сое локализован в плазмалемме клеток ризодермы и коровой паренхимы. Он обусловлен работой Н+-помп, в качестве которых выступают Н+-АТРазы и протонперенсщие редокс-цепи. В этой части корня катионы и анионы из клеточных стенок поступают в цитоплазму. Через клетки эндодермы с поясками Каспари вода и минеральные соли проходят только по симпласту. В паренхимных клетках пучка, непосредственно примыкающих к трахеидам или сосудам, функционирует второй насос, выделяющий минеральные вещества, которые через поры в стенках трахеальных элементов попадают в их полости. Благодаря активной работе двух насосов в трахеидах и сосудах увеличивается осмотический потенциал и, следовательно, сосущая сила.
Читайте также:
lektsia.info