
Большая Энциклопедия Нефти и Газа. Конечный продукт фотосинтеза у растений
Конечные продукты фотосинтеза - Справочник химика 21
Конечные продукты фотосинтеза [c.238]Окисление органических веществ. В результате поглощения СО2 и дальнейших его преобразований в ходе фотосинтеза образуется молекула углевода, которая служит углеродным скелетом для построения всех органических соединений в клетке. Органические вещества, возникшие в процессе фотосинтеза, характеризуются высоким запасом внутренней энергии. Но энергия, аккумулированная в конечных продуктах фотосинтеза — углеводах, жирах, белках,— недоступна для непосредственного использования ее в химических реакциях. Перевод этой потенциальной энергии в активную форму осуществляется в процессе дыхания. Дыхание включает механизмы активации атомоп водорода органического субстрата, освобождения и мобилизации энергии в виде АТФ и генерации различных углеродных скелетов. В процессе дыхания углевод, жиры и белки в реакциях биологического окисления и постепенной перестройки органического скелета отдают спои атомы водорода с образованием восстановленных форм. Последние при окислении в дыхательной цепи освобождают энергию, которая аккумулируется в активной форме в сопряженных реакциях синтеза АТФ. Таким образом, фотосинтез и дыхание — это разли ные, но тесно связанные стороны общего энергообмена. [c.609]
Интересно, что с возрастом увеличивается скорость превращения продукта фиксации СО2 — фосфоглицериновой кислоты — в конечные продукты фотосинтеза. [c.248]Фотосинтез углеводов. Зеленые растения синтезируют углеводы нз углекислого газа и воды при участии солнечной энергии и хлорофилла. Конечным продуктом фотосинтеза растений является крахмал. Процесс фотосинтеза и роль в нем хлорофилла, содержащегося в хлорофилловых зернах, исследованы великим русским ученым К. А. Тимирязевым (1843—1920). [c.168]
В то время как прямая химическая связь между промежуточными продуктами фотосинтеза и субстратами дыхания возможна, но еще не доказана, стимуляция дыхания конечными продуктами фотосинтеза (углеводами) установлена с несомненностью и является вполне естественной, так как известно, что дыхание может стимулироваться и доставляемыми извне сахарами. [c.573]
Прежде всего флуоресценция конкурирует только с первичной фотохимической реакцией, а не со всем процессом фотосинтеза. Скорость фотосинтеза, измеренная по выделению кислорода или поглощению углекислоты, часто определяется не только эффективностью первичного фотопроцесса, но также и скоростью одной или нескольких связанных с этим процессом темновых каталитических реакций. К их числу относятся реакции, которые превращают первичные фотопродукты в стабильные конечные продукты фотосинтеза. Когда эти завершающие реакции слишком слабы, чтобы идти наравне с первичным фотохимическим процессом (что может иметь место, например, в очень сильном свете, или при низких температурах, или в присутствии некоторых ядов), первичные фотопродукты будут накопляться до определенной концентрации и вновь исчезать при обратных реакциях. Вследствие этого квантовый выход фотосинтеза уменьшится, однако на интенсивности флуоресценции это не отразится, так как первичный фотохимический процесс, конкурирующий с флуоресценцией, продолжается с неизменной скоростью. Этим можно объяснить существование светового насыщения в фотосинтезе, без одновременного возрастания выхода флуоресценции (явление, о котором мы упоминали выше). [c.234]
Многие углеводы и другие вещества, образованные при фотосинтезе, подвергаются сложным превращениям в процессе дыхания растений. Глюкоза — конечный продукт фотосинтеза — содержит значительное количество солнечной энергии, заключенной в ее молекуле. [c.11]
Содержание энергии в конечных продуктах фотосинтеза— сахаре и кислороде — известно. Оно равно количеству тепла, образующегося при окислении сахара до углекислоты и воды. Это составляет 112 больших калорий на грамм-атом (атомный вес элемента, выраженный в граммах) углерода. Таково, следовательно, минимальное количество энергии, которое должен дать свет для фотосинтеза. Для того чтобы восстановить молекулу углекислоты до уровня восстановленности углевода, нужно перенести на эту молекулу четыре атома водо- [c.45]
После рассмотрения сравнительно простых реакций образования отдельных моносахаридов необходимо остановиться на весьма сложном, но исключительно важном процессе фотосинтеза. Это нужно сделать не только потому, что конечные продукты фотосинтеза — углеводы [c.204]
Параллельно образованию конечных продуктов фотосинтеза происходит сложная цепь процессов, приводящих к регенерации молекулы акцептора СОг. Последний представляет собой фосфорсодержащее соединение, при образовании которого используется энергия АТФ, образующаяся в процессе фотосинтетического фосфорилирования. [c.174]
Конечные продукты фотосинтеза, гексозофосфаты, переводятся в крахмал для хранения и в сахарозу для транспортировки в другие части растения кроме того, они превращаются в про-, цессе дыхания в различные строительные блоки, необходимые растению. Все эти процессы мы рассмотрим в следующей главе. [c.137]
Подавление фотосинтеза при продолжительной и сильной засухе, особенно если она развивается постепенно и не приво-,дит к резкому падению тургора и водного потенциала (что как раз сплошь и рядом наблюдается в полевых условиях), в значительной степени вызывается не только и даже не столько повреждением самих хлоропластов, которые особенно устойчивы к обезвоживанию [92, 449—451], сколько задержкой оттока из. .листьев ассимилятов [98, 452—455], связанной с уменьшением их потребления вследствие задержки ростовых процессов. Накопление ассимилятов — конечных продуктов фотосинтеза — приводит к перекорму листьев и ингибирующе действует на фотосинтез [9, 456], подобно тому как накопление в избыточном количестве продуктов любой химической реакции начинает тормозить эту реакцию. Таким образом, в условиях засухи продукты фотосинтеза потребляются медленнее, чем они вырабатываются. Об этом свидетельствует и повышенное содержание [c.180]
В итоге за счет полимеризации получается конечный продукт фотосинтеза — гек-соза СбН120б. [c.178]
Крахмал СвНю05)п — полисахарид. Образуется на свету в листьях растений, является конечным продуктом фотосинтеза. В состав К. входят амилоза и амилопек тин. К. дает синее окрашивание с иодом, подвергается гидролизу. Конечным продуктом гидролиза К. является глюкоза [c.72]
Итогом двух фотохимических реакций является создание ассимиляционной силы — НАДФ Н2 и АТФ. Конечные продукты фотосинтеза в этом случае в принципе аналогичны продуктам, образующимся при бескислородном фотосинтезе, за исключением того, что в последнем случае восстановитель находится в форме НАД Н2. [c.289]
Крахмал (СвНюОб) . Крахмал — запасное питательное вещество растений. Он является конечным продуктом фотосинтеза. Крахмал образуется на свету в зеленых частях растений. Далее он подвергается гидролизу образующиеся при этом более простые углеводы переносятся в остальные части растения, где частично идут на построение клеток и тканей или используются как источник энергии, а частично превращаются снова в крахмал, который откладывается в виде запасного материала в клубнях и других частях растений. Крахмальное зерно неоднородно и состоит из двух веществ амилозы и амилопектина. Амилоза представляет собой длинную цепочку из многих остатков глюкозы (от 100 до 1000), сое- [c.180]
Чтобы объяснить насыщение, не сопровождаемое изменениями интенсивности флуоресценции, лимитирующий катализатор следует поместить не между одним из двух реагентов (A Og или A HgO) и светочувствительным комплексом, а между первичными и конечными продуктами фотосинтеза. Другими словами, обратные реакции, вызываемые недостатком катализатора, должны быть скорее вторичными, чем первичными. Обратная реакция такого типа (г) была добавлена в механизмы (28.20) и (28.21). Мы можем постулировать, например, что реакция (28.20г) имеет место потому, что превращение первого фотопродукта AH Og в более устойчивое промежуточное соединение требует катализатора Ев (возможно, мутазы), который имеется в ограниченном количестве. Подобный постулат можно сделать и в отношении действия катализатора Ес, который требуется для первой стадии превращения А НО в свободный кислород. Благодаря симметрии между правой и левой сторонами в схемах на фиг. 194 и 195 ограничение в использовании продуктов окисления будет иметь то же влияние на кинетику процесса в целом, как и ограничение в использовании продуктов восстановления. В первом случае вторичная обратная реакция будет ускоряться накоплением первичного окисленного продукта, А НО во втором случае — накоплением первичного восстановленного продукта, AH Og. [c.467]
Много усилий затрачено для выяснения характера взаимодействия хлорофилла с белками и липидами, однако связь молекулы хлорофилла с этими структурами до сих пор не ясна. Порфирино-вый фрагмент гидрофилен, фитольный гидрофобен. Какой из них осуществляет связь с соответствующей поверхностью белка, до сих пор не установлено. Выяснено только, что структурный белок in vitro проявляет явную склонность к агрегации и к адсорбции на поверхности как молекул липоидов, так и молекул хлорофилла. Ферментные системы, обеспечивающие запасание энергии света в макроэргических связях фосфора АТФ, находятся в тилакоидах у растений и водорослей и в хроматофорах у бактерий. Ассимиляция же СО2 вплоть до образования конечных продуктов фотосинтеза осуществляется с помощью соответствующих ферментов стромы. [c.14]
У некоторых водорослей нет фермента гликолатоксидазы, из-за чего гликолевая кислота становится конечным продуктом фотосинтеза, выделяемым в водоемы. Возможно, это играет большую роль в развитии фитопланктона и бактерий. [c.39]
Интересно, что у ба ктер ий в процессе уов оен ия СО2 синтезируются, главным образом, не углеводы. В то же время у водорослей и высших растений углеводы выступают в роли основных конечных продуктов фотосинтеза. Такое аильное отличие определяется, по-види-мо му, тем, что у аэробных фотосинтезирующих форм растений углеводы могут легко и. по мере надобност и расходоваться в процессах дыхания. При этом происходит и освобождение энергии и образование в цикле Кребса тех соещинений, которые я1вяяются конечными продуктам)И бактериального фотосинтеза (фоторедукции). [c.238]
Изучение оттока ассимилятов из хлоропластов в окружающую их среду in vivo и in vitro заставило изменить принятое раньше мнение о том, что все фосфорилированные продукты усвоение СО не диффундируют в цитоплазму, а выполняют лишь функцию промежуточных звеньев, через которые радиоактивный углерод проскакивает в конечные продукты фотосинтеза. [c.266]
Рассмотрим, почему удобнее выражать эффективность фотосинтеза через квантовый выход или квантовый расход. Этот способ выражения основывается на законе фотохимических эквивалентов Эйнштейна в фотохимической реакции один фотон взаимодействует с одной молекулой. Этот закоп, несомненно, применим для первичных превраш ений в захватывающих центрах. Однако не все первичные электронные переходы ведут к образованию конечных продуктов фотосинтеза, а эти последние, вероятно, сильно отличаются от первичных продуктов. [c.585]
Образование конечных продуктов фотосинтеза - это путь углерода от 3-ФШ до конечных стабильных продуктов. Сначала Ф1К восстанавливается при участии образовавшихся в световой фазе фотосинтеза молекул АТФ и НАДФ Н в фосфогли[1ериновый альдегид (ФГА), а затем полученные триозы путем рада ферментативных превращений образуют конечные продукты фотосинтеза - углеюды или другие соединения. [c.244]
Др(угой путь превращения 3-Ф1К, приводящий к образованию различннх органических кислот и аминокислот, изложен ниже (стр. 25 1 ). Молекулы 3-ФГК и 3-ФГА участвуют таким образом в дальнейших многочисленных ферментативных реакциях, приводящих к образовавию конечных продуктов фотосинтеза и к регенврвдии акцептора С02-рибулозодифосфата. Список важнейших ферментов, принямавщих участие в восстановительном цикле углерода, и катализируемые ИМИ реакции приведены на рис.4В и в табя.40. [c.251]
Из образовавшихся при полном обороте цикла 6 молекул фосфотриоз 5 участвуют в реакциях, приводящих к регенерации акцептора СО . Сюбодной остается одна триоза, которая служит исходным материалом для образования конечных продуктов фотосинтеза. Одна молекула гексозофосфата образуется из 2 фосфотриоз [c.253]
Раньше считали, что конечный продуктами фотосинтеза являются только углеюды, из которых в дальнейшем путем вторичных превращений строятся все разнообразные и сложные вещества,соо-тавлящие растение. [c.254]
Экспериментальные работы, выполненные советскими учеными под руководством А. А. Ничиноровича, приводят к важному выводу, что углеводы не являются единственными конечными продуктами фотосинтеза. В зависимости от условий фотосинтеза образуются не только углеводы, но также белки и некоторые другие соединения. [c.66]
Не останавливаясь подробно на световой фазе фотосинтеза [64], исследованной, главным образом, Кэлвиным и др., следует упомянуть, что важнейшими процессами здесь являются поглощение хлорофиллом квантов света и использование их энергии для синтеза богатых энергий пирофосфатных связей (АТФ, НАДФ-Н2) поглощаемая энергия света используется при разложении воды, кислород которой выделяется в виде О 2 как конечный продукт фотосинтеза, а водород используется для восстановления при участии АТФ и НАДФ-Н фосфоглицериновой кислоты на второй, темновой стадии фотосинтеза. [c.204]
В основе первичных процессов фотосинтеза ППФ лежит сложная совокупность окислительно-восстановительных реакций переноса электрона между компонентами электрон-транспортной цепи ЭТЦ. Наибольший интерес представляют механизмы трех основных стадий трансформации энергии в ННФ поглощение света фотосинтетическими пигментами и миграция энергии электронного возбуждения на РЦ фотосинтеза первичное разделение зарядов и трансформация энергии в РЦ перенос электрона по ЭТЦ и сопряженные с ним процессы, приводящие к образованию первичных стабильных продуктов (НАДФ и АТФ), используемых в дальнейших темновых реакциях фиксации СО2 и образования конечных продуктов фотосинтеза. [c.280]
КИСЛОТЫ транспортируются к хлоропластам, локализованным в-клетках обкладки. Здесь СОг высвобождается в высокой кон--центрации и при сравнительно низком содержании кислорода,, благодаря чему хлоропласты этих клеток могут весьма эффективно фиксировать СОг в виде сахаров через цикл Кальвина — Бенсона. С этой точки зрения С4-фиюсация представляется чем-то вроде насоса, поставляющего СОг для Сз-пути. К этому можно добавить, что само положение клеток обкладки создает условия для передачи конечных продуктов фотосинтеза (в частности, сахарозы) непосредственно в ситовидные трубки флоэмы, по которым эти продукты могут затем транспортироваться в другие части растения. [c.135]
Подсчитаем энергию связей для исходных и конечных продуктов фотосинтеза. В исходных продуктах СО2 и Н2О содержатся две С = 0-связи (190 ккал/моль) и две О—Н-связи (ПО ккал/моль). Итого 190-2+ -f 110-2 = 600 ккал/моль. В конечных продуктах СН2О и [c.44]
Итогом двух фотохимических реакций у цианобактерий является образование ассимиляционной силы — НАДФ1Н2 и АТФ. Как можно видёть, конечные продукты фотосинтеза цианобактерий в принципе аналогичны продуктам, образующимся при фотосинтезе пурпурных и зеленых бакте рий, за исключением того, что у двух последних, групп йосстановитель образуется в форме НАД-Нг. [c.246]
З./Фаза регенерации первичного акцептора диоксида углерода и синтеза конечного продукта фотосинтеза. В результате описанных выше реакций при фиксации трех молекул СО2 и образовании шести молекул восстановленных 3-фосфотриоз пять из них используются затем для регенерации рибулозо-5-фосфата, а один — для синтеза глюкозы. 3-ФГА под действием триозофосфатизомеразы изомеризуется в фосфодиокси-ацетон. При участии альдолазы 3-ФГА и фосфодиоксиацетон конденсируются с образованием фруктозо-1,6-дифосфата, у которого отщепляется один фосфат с помощью фруктозо-1,6-дифосфатазы. В дальнейших реакциях, связанных с регенерацией первичного акцептора СО 2, последовательно принимают участие транскетолаза и альдолаза. Транскетолаза катализирует перенос содержащего два углерода гликолевого альдегида от кетозы на альдозу [c.92]
IV. Стадия синтеза продуктов. Конечными продуктами фотосинтеза считаются в первую очередь сахара и углеводы. Однако установлено, что в ходе фиксации СОг при фотосинтезе образуются также жиры, жирные кислоты, аминокислоты и органические кислоты. Многие детали соответствующих реакций известны, но для нас они опять-таки большого интереса не представляют. Следует лишь отметить, что в разных условиях, различающихся по освещенности, концентрации СОг, Ог и т. п., по-видимому, происходит образование разных конечных продуктов (рис. 6.7). В последнее время синтез конечных продуктов исследуется очень активно, пoqкoлькy понимание механизма реакций и выяснение благоприятствующих им факторов могут в конечном счете позволить нам создавать необходимые условия для того, чтобы растения по нашему желанию синтезировали больше или меньше сахаров, жиров, аминокислот. [c.92]
Крахмал нередко называют типичным конечным продуктом фотосинтеза, и он действительно часто накапливается в хлоропластах в течение дня и потребляется ночью (разд. 8.1 и 9.7). Однако количество накапливающегося крахмала варьирует в широких пределах. Некоторые виды растений, например подснежник (Galanthus nivalis), совсем его ие образуют, а листья ряда основных сельскохозяйственных культур, скажем пшени- [c.152]
chem21.info
Конечные продукты фотосинтеза — КиберПедия
Образовавшиеся в цикле Кальвина С3-продукты (3-фосфоглицери-новая кислота, 3-фосфоглицериновый альдегид и фосфодиоксиацетон), а также фруктозо-6-фосфат не накапливаются в хлоропластах. Они участвуют в синтезе сахарозы, крахмала, аминокислот, органических кислот, липидов и белков как непосредственно в хлоропластах, так и в цитоплазме фотосинтезирующих клеток. Фруктозо-6-фосфат в значительном количестве изомеризуется в глюкозо-6-фосфат, который далее используется для синтеза крахмала. Накопление крахмала может достигать до 90 % от общего объёма хлоропластов.
Синтез сахарозы происходит в цитоплазме, в которую легко диффундируют из хлоропластов фосфоглицериновая кислота, фосфоглицериновый альдегид и фосфодиоксиацетон. Из триоз по уже известным механизмам образуются гексозы, а уже с их участием синтезируются молекулы сахарозы (см. стр…). Большая часть синтезированной в цитоплазме сахарозы транспортируется в акцепторные клетки по флоэме.
В ходе фотосинтеза в цитоплазме инициируются и другие процессы: синтез жирных кислот, восстановление нитратов и сульфатов, образование липидов и белков и др. Для их осуществления необходимы биоэнергетические продукты в виде АТФ и восстановленных динуклеотидов НАДФ×Н и НАД×Н . Но они очень слабо проникают через оболочку хлоропластов. Поэтому для их переноса из хлоропластов в цитоплазму выработаны специальные механизмы.
Перенос в цитоплазму макроэргических связей АТФ осуществляется в результате взаимных превращений фосфоглицериновой кислоты и фосфодиоксиацетона, при этом также происходит перенос восстановительного потенциала НАД×Н (рис. 35). Указанные С3 – продукты легко диффундируют через оболочку хлоропластов. В цитоплазме фосфодиоксиацетон изомеризуется в 3-фосфоглицериновый альдегид, который далее под действием фермента триозофосфатдегидрогеназы фосфорилируется с образованием 1,3-дифосфоглицериновой кислоты. В ходе реакции также образуются восстановленные динуклеотиды НАД×Н. Затем остаток ортофосфорной кислоты с макроэргической связью переносится на АДФ и таким образом осуществляется синтез АТФ. Эту реакцию катализирует фермент фосфоглицераткиназа.
|
Образовавшаяся в цитоплазме фосфоглицериновая кислота далее диффундирует в хлоропласты, включается там в реакции цикла Кальвина и снова превращается в фосфодиоксиацетон, способный вновь перемещаться в цитоплазму и т.д.
Одним из механизмов переноса из хлоропластов в цитоплазму восстановительного потенциала НАДФ×Н является взаимное превращение яблочной и щавелевоуксусной кислот. Щавелевоуксусная кислота легко проникает через оболочку хлоропластов и подвергается превращению в яблочную кислоту с участием НАДФ×Н под действием фермента малатдегидрогеназы. Затем яблочная кислота диффундирует в цитоплазму и там окисляется с участием дегидрогеназы в щавелевоуксусную кислоту. При этом происходит образование НАДФ×Н:
|
После этого щавелевоуксусная кислота вновь диффундирует в хлоропласты и там превращается в яблочную кислоту и т.д. В результате каждого такого превращения в хлоропласте потребляются восстановленные динуклеотиды НАДФ×Н, а в цитоплазме регенерируются.
Выход продуктов фотосинтеза зависит от условий окружающей среды, которые могут ускорять или ингибировать биохимические реакции при фотосинтезе . К важнейшим факторам, прямо влияющим на процесс фотосинтеза, относятся интенсивность и состав солнечной радиации, концентрация в атмосфере СО2 и О2, температура, обеспеченность влагой и элементами питания.
На световую стадию фотосинтеза решающее влияние оказывает интенсивность солнечной радиации. Образование продуктов фотосинтеза начинается при освещённости, составляющей 1% от полного солнечного света, однако максимальная скорость фотосинтеза наблюдается у разных видов растений при освещённости 10-40 тыс. лк, тогда как в ясный солнечный день уровень освещённости может достигать 100 тыс. лк. Чрезмерно высокая интенсивность солнечной радиации может повреждать пигментные системы фотосинтезирующих клеток и ингибировать фотосинтез.
На образование конечных продуктов фотосинтеза существенное влияние оказывает состав солнечной радиации. Если в ней увеличивается доля коротковолнового света (синих лучей), то в фотосинтезирующих органах уже при коротких экспозициях инициируется включение углерода СО2 в неуглеводные продукты – аланин, аспарагиновую, яблочную, лимонную, глутаминовую кислоты, серин, а через несколько минут меченый углерод СО2 обнаруживается в значительном количестве в составе белков. При увеличении доли длинноволнового света (красных лучей) меченый углерод СО2 при коротких экспозициях более интенсивно включается в моносахариды и их фосфорнокислые эфиры, а при 5-минутной и более длительных экспозициях – в состав крахмала.
Кроме того, в опытах отмечено, что скорость фотосинтеза заметно возрастает при добавлении к красному свету небольшого количества синего света (до 20 %), который действует в определённой степени как регулятор фотохимических реакций. Увеличение интенсивности синтеза азотистых веществ под действием синего света также связано с его регуляторным действием на ферменты, катализирующие реакции обмена азотистых веществ. Выяснено, что роль хромофоров у этих ферментов, поглощающих синий свет, выполняют флавиновые группировки.
Темновая стадия фотосинтеза зависит от концентрации СО2. Оптимум концентрации СО2 для С3-растений составляет 0,1-0,3%, тогда как среднее его содержание в атмосфере – 0,03%. При такой концентрации СО2 интенсивность фотосинтеза у С3-растений достигает лишь 50 % от максимальной. Поэтому для тепличных культур разрабатываются специальные технологии выращивания растений в атмосфере с повышенной концентрацией СО2. Как было показано ранее, при пониженных концентрациях СО2 в атмосфере более эффективно осуществляют фотосинтез С4-растения, у которых первичное связывание СО2 катализирует фермент фосфопируваткарбоксилаза, обладающий более высокой карбоксилирующей активностью и не способный инициировать процесс фотодыхания.
У С3-растений интенсивность темновой стадии фотосинтеза также в значительной степени определяется концентрацией О2. При понижении концентрации О2 в окружающей среде до 3-5 % увеличивается выход продуктов фотосинтеза за счёт ослабления оксигеназной реакции, которую катализирует фермент рибулозодифосфаткарбоксилаза в условиях повышенной концентрации О2.
Все ферментативные реакции темновой и световой стадий фотосинтеза зависят от температуры окружающей среды и при её повышении их скорости возрастают как и при любом химическом процессе. Однако при температурах свыше 35°С наблюдаются солнечные ожоги листьев, что приводит к повреждению пигментных и других фотосинтетических структур, в результате чего интенсивность фотосинтеза понижается. Температурный оптимум С3-растений наблюдается при 25°С, а у С4-растений повышается до 35°С.
Обеспеченность растений водой оказывает как прямое, так и косвенное влияние на фотосинтез. Вода непосредственно потребляется в реакциях фотосинтеза и является источником кислорода. С другой стороны, основой жидкой физиологической среды растительных клеток, включая и хлоропласты, являются также молекулы воды и при её недостатке нарушается функционирование мембранных структур и вследствие усиления вязкости жидкой физиологической среды понижаются скорости ферментативных реакций. Кроме того, водный режим растений влияет на работу устьиц, которые регулируют поступление во внутреннюю физиологическую среду СО2 из атмосферы. При дефиците и избытке воды устьица закрываются.
Для формирования и функционирования фотосинтетических структур растений также необходим довольно сложный комплекс различных химических элементов, которые должны содержаться в фотосинтезирующих клетках в оптимальных концентрациях. Недостаток или избыток питательных элементов понижает интенсивность фотосинтеза, оказывая влияние на его световую или темновую стадии, работу каталитических и мембранных структур хлоропластов, взаимодействие компонентов электронтранспортных цепей и др. Роль отдельных химических элементов в процессе фотосинтеза показана при изложении конкретных механизмов фотохимических реакций в данной главе.
Следует также отметить воздействие на фотосинтетический аппарат растений различных химических загрязнителей окружающей среды. Так, например, выяснено, что сернистый газ промышленных выделений даже в малых концентрациях повреждает листья многих растений, в результате чего понижается выход фотосинтетических продуктов и происходит недобор урожая сельскохозяйственных культур.
Некоторые фотосинтезирующие организмы (одноклеточные водоросли, некоторые бактерии) образуют в анаэробных условиях в качестве основного продукта жизнедеятельности молекулярный водород, который может использоваться в энергетике страны как высокоэффективное топливо. В связи с этим проводятся исследования по созданию искусственных биологических систем, обладающих способностью улавливать солнечную энергию и превращать её в химическую энергию молекул водорода.
|
|
Первая искусственная биологическая система (1973 г.) для фотосинтетического получения водорода включала изолированные хлоропласты высших растений, ферредоксин и ферментный комплекс гидрогеназы, выделенный из бактерий.
Фотосинтез у бактерий
Клетки фотосинтезирующих бактерий отличаются по своей структуре от клеток растений и эукариотических водорослей. В них нет хлоропластов и пигменты локализованы в цитоплазмотических мембранах. Главный пигмент большинства фотосинтезирующх бактерий – бактериохлорофилл. У этих бактерий отсутствует белково-пигментный комплекс фотосистемы II, поэтому не происходит фотоокисления воды при фотосинтезе. У них в качестве доноров электронов для реакций фотосинтеза используется не вода, а другие соединения: h3S, Н2, органические вещества. В связи с этим у них функционирует только фотосистема I, а фотосистемы II нет.
Цианобактерии (сине-зелёные водоросли), как и другие фотосинтезирующие бактерии, не имеют хлоропластов, но главный пигмент у них - хлорофилл, который входит в состав белково-пигментных комплексов как фотосистемы I, так фотосистемы II. Поэтому у цианобактерий в качестве донора водорода для фотосинтетических реакций, как и у растений, используются молекулы воды, в результате фотолиза которых выделяется О2.
У галофильных бактерий (Н. halobium) роль светопоглощающего пигмента выполняет родопсиноподобный белок с молекулярной массой 26000, входящий в состав поверхностной мембраны, окружающей цитоплазму. Этот белок получил название бактериородопсина. В нём в качестве активной группировки содержится ретиналь – альдегидная форма витамина А. Пептидные цепи бактериородопсина образуют спиралевидные структуры, вытянутые вдоль длинной оси их молекулы, но расположены перпендикулярно по отношению к плоскости мембраны. На долю бактериородопсина приходится до 75 % от общей массы поверхностной цитоплазматической мембраны.
При поглощении света бактериородопсин инициирует перенос электронов по электронтранспортной цепи, приводящей в действие Н+-помпу, способную перемещать протоны через цитоплазматическую мембрану на её внешнюю поверхность, создавая трансмембранный электрохимический потенциал. В дальнейшем энергия трансмембранного потенциала используется для осуществления ионного транспорта и синтеза АТФ в клетках галофильных бактерий, при этом фотолиза воды не происходит.
По способу питания фотосинтезирующие бактерии подразделяются на две группы – фотоавтотрофные и фотогетеротрофные. Первые способны использовать в качестве источника углерода для синтеза органических веществ СО2 воздуха или другой окружающей среды. Вторые используют в качестве источников углерода различные органические вещества.
К типичным фотоавтотрофным бактериям относятся зелёные и пурпурные серобактерии. В их клетках найдены ферментные системы, которые способны с участием восстановленного ферредоксина катализировать присоединение СО2 к ацетилкоферменту А или сукцинилкоферменту А:
|
Образующиеся в ходе этих реакций продукты могут включаться в различные биохимические превращения, связанные с обеспечением жизнедеятельности рассматриваемых бактерий.
У некоторых пурпурных бактерий при фотосинтезе происходит накопление аспарагиновой кислоты, которая синтезируется из 3-фосфо-глицериновой кислоты, образующейся в результате присоединения СО2 к рибулозо-1,5-дифосфату. Фосфоглицериновая кислота превращается в фосфоенолпировиноградную кислоту, которая, подвергаясь карбоксилированию, образует щавелевоуксусную кислоту. Затем из щавелевоуксусной кислоты в результате реакции переаминирования может синтезироваться аспарагиновая кислота.
|
К фотогетеротрофным относятся галобактерии и некоторые разновидности пурпурных несерных бактерий. Они под действием света способны использовать в качестве субстрата питания для превращения в продукты жизнедеятельности уксусную, пировиноградную кислоты и некоторые другие органические вещества.
9.2. Ассимиляция СО2 за счёт использования энергии
Химических реакций.
Известна большая группа хемоавтотрофных бактерий, которые используют для синтеза органических веществ в качестве источника углерода СО2, а энергию для этих процессов получают в результате окисления неорганических веществ: сероводорода, серы, аммиака, нитратов, водорода, восстановленных форм железа и марганца и некоторых других неорганических соединений. Синтез организмами органических веществ из СО2 за счёт энергии, полученной не от солнечных лучей, а с помощью химических реакций, получил название хемосинтеза. Типичным примером хемоавтотрофных организмов являются бесцветные серобактерии, которые получают энергию для восстановления СО2 в органические вещества в результате окисления сероводорода или серы:
2Н2S + O2 ¾¾® 2h3O + S2 + DG (Thiobacillus)
S2 + 3O2 + 2h3O ¾¾® 2h3SO4 + DG
Высвобождающаяся при окислении указанных веществ энергия используется для синтеза АТФ и других нуклеозидполифосфатов, которые участвуют в синтезе органических веществ, образуемых из СО2. Как было установлено в опытах с меченым углеродом, первичным акцептором СО2 у этих и других хемоавтотрофных бактерий служит рибулозо-1,5-дифосфат и продуктом этой реакции так же, как и в хлоропластах растений, является 3-фосфоглицериновая кислота.Нитрофицирующие бактерии в качестве источника энергии используют восстановленные формы азота в виде аммиака и нитритов, которые они с помощью своих ферментов подвергают окислению кислородом:
2NH₃ + 3O2 ¾¾® 2HNO2 + 2h3O + DG (Nitrosomonas)
2HNO2 + O2 ¾¾® 2HNO3 + DG (Nitrobacter)
Эти бактерии играют важную роль в круговороте азота в биосфере и формировании плодородия почвы, так как от их деятельности зависит процесс нитрификации, обогащающий почву нитратной формой азота.
Хемоавтотрофные бактерии, способные окислять восстановленные формы железа и марганца, по-видимому стали источником отложения железных и марганцевых руд. Они также способны использовать энергию химических реакций окисления для синтеза органических веществ путем ассимиляции СО2.
Частичное усвоение СО2 происходит даже в клетках гетеротрофных организмов. Выявлено несколько типов реакций, в которых происходит связывание СО2 с органическими субстратами. Под действием соответствующих ферментов карбоксилированию подвергаются некоторые кетокислоты, например, пировиноградная кислота:
|
Хорошо известный путь гетеротрофного карбоксилирования – присоединение СО2 к фосфоенолпировиноградной кислоте. Такие реакции изучены у растений и автотрофных бактерий. Роль катализатора в них выполняет фермент фосфопируваткарбоксилаза. Причём фиксация СО2 под действием данного фермента происходит в нефотосинтезирующих клетках.
В клетках растений, микроорганизмов и животных найден фермент фосфопируваткарбоксикиназа, который катализирует присоединение СО2 к фосфоенолпировиноградной кислоте с участием ГДФ, при этом образуется яблочная кислота и ГТФ:
|
К важным реакциям карбоксилирования следует отнести синтез карбамоилфосфата, катализируемый ферментом карбамоилфосфатсинте-тазой. Источником углерода в этой реакции служит бикарбонат-ион (НСО3¯):
|
Образовавшийся карбамоилфосфат далее используется в синтезе пиримидиновых нуклеотидов, а также в реакциях орнитинового цикла (см. «Обмен азотистых веществ»).
С участием ферментов фосфопируваткарбоксилазы и фосфопируваткарбоксикиназы по-видимому осуществляется усвоение СО2, поступившего из почвы, в клетках корней растений. Фосфоенолпировиноградная кислота в них образуется в ходе дыхательных реакций. Продукт реакций гетеротрофного карбоксилирования – щавелевоуксусная кислота может далее включаться в реакции цикла Кребса и вместе с другими продуктами этого цикла поступать в любые акцепторные клетки растительного организма и использоваться в них для синтеза различных органических веществ.
Таким образом, у растений наряду с процессом фотосинтеза существует дополнительный механизм углеродного питания за счёт СО2 почвы, поступающего в нефотосинтезирующие клетки корней. Однако первичный акцептор СО2 для этих реакций - фосфоенолпировиноградная кислота - образуется в результате окисления фотоассимилятов, поступающих в корни из листьев.
Дыхание
Основной источник энергии для поддержания процессов жизнедеятельности у гетеротрофных организмов – это химическая энергия органических веществ, которые они используют в качестве пищи. Несинтезирующие органы растений, а в темноте и способные к фотосинтезу клетки также используют в качестве источника энергии органические вещества, образующиеся как продукты фотосинтеза. Для того чтобы химическая энергия органических веществ могла использоваться для поддержания жизнедеятельности организма, они подвергаются окислению. При этом на каждом этапе окислительного распада веществ выделяется энергия, которая связывается в ходе определённых реакций в виде макроэргических соединений и восстановленных динуклеотидов и может затем использоваться для осуществления эндергонических процессов, связанных с синтезом, транспортом и активированием различных химических веществ. В ходе биологического окисления образуются промежуточные метаболиты, которые затем участвуют в синтезе конечных продуктов, выполняющих в организме те или иные биологические функции (структурные, запасные вещества и др.).
Последовательность биохимических реакций, лежащих в основе биологического окисления, называют дыханием. У растений и других высших организмов в реакциях дыхания участвует кислород, а в качестве продуктов дыхания выделяются СО2 и Н2О. Первые сведения о дыхании растений были получены Я. Ингенхаузом (1778 г.), а к концу 19 века было установлено общее уравнение дыхательного процесса:
С6Н12О6 + 6О2 ¾¾® 6СО2 + 6Н2О + DG, (1)
где С6Н12О6 – моносахариды, образуемые при фотосинтезе.
В дальнейшем благодаря работам В.И. Палладина (1912 г.) было показано, что на первом этапе дыхательных реакций происходит отщепление водорода от молекул сахара и воды и образование промежуточных восстановленных продуктов, а на следующем этапе эти продукты уже окисляются кислородом с образованием воды. Поэтому дыхательный процесс можно представить в две стадии:
С6Н12О6 + 6Н2О ¾® 6СО2 + 24[Н] (2)
24[Н] + 6О2 ¾® 12Н2О (3)
Если из уравнений (2) и (3) составить суммарное уравнение процесса, то мы получим уравнение (1).
В 1921–25 г.г. в опытах по изучению роли фермента цитохромоксидазы О.Г. Варбург и Д. Кейлин установили, что на второй стадии дыхания на кислород переносятся электроны и протоны, которые и участвуют в образовании молекул воды:
О2 + 4ē + 4Н+ ¾® 2Н2О
Отщепление электронов и протонов от молекул сахара и воды, как было выявлено в ходе дальнейших исследований, катализируют ферменты дегидрогеназы и с их участием происходит образование восстановленных динуклеотидов НАД×Н и ФАД×Н2, при окислении которых и осуществляется перенос электронов и протонов на кислород.
По современным представлениям процесс дыхания у растений состоит из трёх стадий: гликолиза, цикла ди- и трикарбоновых кислот и окисления восстановленных динуклеотидов, образующихся на первых двух стадиях дыхания, с участием ферментных комплексов, локализованных во внутренних мембранах митохондрий.
Гликолиз
В процесс гликолиза включаются активированные формы моносахаридов – гексоз, которые подвергаются окислительному распаду с образованием пировиноградной кислоты. Большой вклад в раскрытие механизмов этих реакций внесли известные биохимики Г. Эмбден, О.Ф. Мейергоф и Я.О. Парнас. Гликолиз представляет собой анаэробную стадию дыхания у высших организмов и основной механизм дыхания у анаэробных микроорганизмов. Реакции гликолиза локализованы в жидкой фазе цитоплазмы и хлоропластов. Ферменты, катализирующие эти реакции в цитоплазме, образуют комплексы с актиновыми белками цитоплазматических филаментов и таким образом обеспечивается локализация происходящих превращений.
Активирование гексоз осуществляется путём фосфорилирования с участием АТФ под действием соответствующих киназ и образующиеся фосфаты гексоз изомеризируются во фруктозо-6-фосфат, который уже далее включается в реакции гликолиза. В первой реакции фруктозо-6-фосфат подвергается повторному фосфорилированию под действием фермента фосфофруктокиназы:
(1)
+ АТФ ¾® + АДФ
фруктозо-6-фосфат фруктозо-1,6-дифосфат
Образующийся в этой реакции фруктозо-1,6-дифосфат имеет полярную молекулу, которая легко подвергается расщеплению на следующем этапе превращений.
Фермент фосфофруктокиназа – аллостерический и поэтому подвержен активной регуляции. Его активность ингибируется высокой концентрацией АТФ и лимонной кислоты, образующейся в цикле Кребса. Однако фермент аллостерически активируется АДФ. Следовательно, при высокой обеспеченности клетки энергией, повышающей концентрацию АТФ, происходит замедление первой реакции гликолиза и в целом интенсивности дыхания. А когда энергетический потенциал клетки понижается и возрастает концентрация АДФ, последний аллостерически активирует фермент фосфофруктокиназу, вследствие чего возрастает скорость включения фрутозо-6-фосфата в реакции гликолиза, вызывая повышение интенсивности дыхания, обеспечивающего клетку энергией.
На следующем этапе гликолиза под действием фермента альдолазы происходит расщепление фруктозо-1,6-дифосфата на две триозы – 3-фос-фоглицериновый альдегид и фосфодиоксиацетон:
|
(
Между триозофосфатами устанавливается динамическое равновесие с участием фермента триозофосфатизомеразы:
|
Образовавшийся 3-фосфоглицериновый альдегид далее окисляется в 3-фосфоглицериновую кислоту под действием фермента глицеральдегид-фосфатдегидрогеназы (1.2.1.12), молекулы которого состоят из четырёх полипептидных субъединиц с молекулярными массами по 45000 и содержат в активном центре тиоловые группы (-SН) и окисленные динуклеотидные группировки НАД+. При взаимодействии фермента с 3-фосфоглицери-новым альдегидом образуется фермент-субстратный комплекс, в котором инициирутся возникновение ацилмеркаптановой макроэргичесой связи и восстановление динуклеотидной группировки НАД+ в НАД×Н:
|
|
|
Так как в реакции (5) макроэргическая ацилфосфатная связь возникает в составе субстрата, такой процесс называют субстратным фосфорилированием.
Полученная в реакции (6) 3-фосфоглицериновая кислота под действием фермента фосфоглицератмутазы изомеризуется затем в 2-фосфо-глицериновую кислоту:
|
|
В результате дегидратации в молекуле субстрата происходит перераспределение внутренней энергии и возникает макроэргическая связь.
|
Пируваткиназа аллостерически ингибируется высокой концентрацией АТФ и ацетилкофермента А, который образуется при дальнейшем превращении пировиноградной кислоты в ходе дыхательных реакций.
Продукт реакции (9) енолпировиноградная кислота самопроизвольно подвергается перегруппировке и превращается в более устойчивую кетоформу – пировиноградную кислоту.
|
Учитывая, что из гексозы образуется 2 триозы, в ходе их окисления возникают 2 молекулы пировиноградной кислоты. По этой же причине все реакции гликолиза, начиная с четвертой, при окислении одной молекулы гексозы повторяются дважды. В ходе гликолиза, кроме двух молекул пировиноградной кислоты, синтезируются биоэнергетические продукты в виде четырёх молекул АТФ (в реакциях 6 и 9) и двух молекул НАД×Н (в реакции 4). В связи с тем, что на активирование гексозы затрачивается 2 молекулы АТФ, то окончательный выход АТФ при гликолизе составляет не 4, а 2 молекулы. Кроме того, в реакции 9 происходит дегидратация 2-фосфоглицериновой кислоты, поэтому в расчете на 1 молекулу окисляемой гексозы выделяется 2 молекулы воды. Исходя из этих данных, суммарное уравнение гликолиза принимает следующий вид:
С6Н12О6+2НАД++2Н3РО4+2АДФ ® 2СН3–СО–СООН+2НАД×Н+2Н++ 2АТФ+2Н2О
гексоза пировиноградная
кислота
Конечный продукт гликолиза – пировиноградная кислота представляет собой один из важнейших продуктов дыхания и в целом обмена веществ любого организма. У анаэробных организмов пировиноградная кислота включается в реакции брожения, в ходе которых в зависимости от организма происходит синтез этилового спирта, молочной, уксусной, пропионовой кислоты и некоторых других биохимических продуктов. У растений и других аэробных организмов пировиноградная кислота подвергается окислению до СО2 и Н2О в ходе реакций цикла Кребса. В некоторых органах растений при недостатке кислорода также могут частично происходить реакции брожения, конечным продуктом которых чаще всего является этиловый спирт.
Через образование пировиноградной кислоты, как будет показано далее, осуществляется связь между обменом углеводов, липидов и белков в растительном организме. Из пировиноградной кислоты путём аминирования синтезируется аминокислота аланин, которая участвует в реакциях азотного обмена и синтезе белков. Путём превращения в ацетил-КоА пировиноградная кислота даёт начало синтезу жирных кислот. А с другой стороны, при распаде белков, аминокислот, жирных кислот на определённых этапах образуется пировиноградная кислота, которая может использоваться на синтез углеводов путём обращения реакций гликолиза благодаря тому, что большинство ферментов, катализирующих эти реакции, обладают обратимым действием. Однако некоторые стадии гликолиза необратимы и для их преодоления используются обходные пути.
Первый необратимый этап в ходе превращения пировиноградной кислоты в гексозу – это образование из пировиноградной фосфоенолпирови-
|
|
|
|
Превращение фосфоенолпировиноградной кислоты в 3-фосфо-глицериновый альдегид осуществляется в результате обращения соответствующих реакций гликолиза (8-4). В ходе этих реакций на фосфорилирование 3-фосфоглицериновой кислоты затрачивается АТФ, а на восстановление фер
cyberpedia.su
Конечные продукты фотосинтеза: процесс, растений, фазы, световой, кислород
 Оксигенный (или кислородный) фотосинтез сопровождается выделением кислорода в качестве побочного продукта. В соответствии с этим выделяют бесхлорофилльный и хлорофилльный фотосинтез. Подсчитаем энергию связей для исходных и конечных продуктов фотосинтеза.
Оксигенный (или кислородный) фотосинтез сопровождается выделением кислорода в качестве побочного продукта. В соответствии с этим выделяют бесхлорофилльный и хлорофилльный фотосинтез. Подсчитаем энергию связей для исходных и конечных продуктов фотосинтеза.
Таким образом, в условиях засухи продукты фотосинтеза потребляются медленнее, чем они вырабатываются. У некоторых водорослей нет фермента гликолатоксидазы, из-за чего гликолевая кислота становится конечным продуктом фотосинтеза, выделяемым в водоемы. Возможно, это играет большую роль в развитии фитопланктона и бактерий.
В исходных продуктах СО2 и Н2О содержатся две С = 0-связи (190 ккал/моль) и две О—Н-связи (ПО ккал/моль). Углеводы стоят у истоков всего органического вещества биосферы — это первые продукты фотосинтеза. Oj и воды. Вместе с тем автотрофы осуществляют частичное окисление продуктов фотосинтеза. Фотосинтез является фотоавтотрофным процессом, представляющим собой комплекс реакций поглощения, преобразования и использования световых квантов в разных эндэргонических процессах.
Так, аноксигенный фотосинтез присущ зеленым и пурпурным бактериям. А оксигенный фотосинтез – источник органических веществ и кислорода для всех растений, имеющих хлорофилл, и нуждающихся в доступе атмосферного воздуха для осуществления фотосинтеза.
Фотосинтез и биосфера
На первой стадии кванты света поглощаются фотосинтетическими пигментами, которые переходят в возбужденное состояние и передают энергию к последующим элементам фотосистемы. На фотохимической стадии синтезируются носители энергии клеток – НАДФН и АТФ. Описанные две стадии объединяют понятием световая фаза фотосинтеза.

Благодаря совершенному строению фотосинтетического аппарата зеленых растений, обеспечивается высокая эффективность фотосинтеза. Иногда хлоропласты наблюдаются в клетках стеблей и плодов, однако главным органом фотосинтеза является зеленый лист растения, благодаря особенностям своего строения.

Данные пигменты поглощают кванты видимого света и обеспечивают дальнейшее запасание энергии излучения в виде энергии электрохимического градиента H+ на биологических мембранах. Система бесхлорофилльного фотосинтеза отличается значительной простотой организации, в связи с чем предполагается эволюционно первичным механизмом запасания энергии электромагнитного излучения.
Таким образом, были получены неопровержимые доказательства существования бесхлорофилльного фотосинтеза. При дефиците кислорода помимо бесхлорофилльного фотосинтеза источниками энергии для галобактерий может служить анаэробное нитратное дыхание или сбраживание аргинина и цитруллина.
В качестве экзогенных доноров электронов могут использоваться как органические, так и неорганические восстановители. Фотосинтез — процесс с крайне сложной пространственно-временной организацией. Темновые» реакции или циклы углерода при фотосинтезе. На первом этапе происходит поглощение квантовсветапигментами, их переход в возбуждённое состояние и передача энергии к другим молекулам фотосистемы.
С4-путь фотосинтеза
Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюконеогенез, образование сахаров и крахмала из углекислого газа воздуха. Фотосинтез растений осуществляется в хлоропластах: полуавтономных двухмембранных органеллах, относящихся к классу пластид. На тканевом уровне, фотосинтез у высших растений обеспечивается специализированной тканью — хлоренхимой. В хлоропластах имеются свои ДНК, РНК, рибосомы (70s типа), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих.
Генетика фотосинтеза
Цианобактерии, таким образом, как бы сами являются хлоропластом, и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе. Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно, и хлорофилл за 10−12 сек переходит с него на первое с потерей 100 кДж/моль энергии только в виде теплоты.
С4 фотосинтез практически не сопровождается потерями рибулозо-1,5-бифосфата из цикла Кальвина, поэтому более эффективен. При CAM (англ.Crassulaceae acid metabolism — кислотный метаболизм толстянковых) фотосинтезе происходит разделение ассимиляции CO2 и цикла Кальвина не в пространстве как у С4, а во времени. Этот механизм позволяет максимально экономить воду, однако уступает в эффективности и С4, и С3. Он оправдан при стресстолерантной жизненной стратегии.
Экология фотосинтеза.
Фототрофы обеспечивают конверсию и запасание энергии термоядерных процессов, протекающих на Солнце, в энергию органических молекул. При использовании энергии химических связей органических веществ гетеротрофы высвобождают её в процессах дыхания и брожения. Энергия, получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф), также является запасённой в процессе фотосинтеза. Фотосинтез является основой продуктивности сельско-хозяйственно важных растений.
Хлорофиллы были впервые выделены в 1818 г. П. Ж. Пельтье и Ж. Кавенту. В 1937 г. Роберт Хилл установил, что процесс окисления воды (и выделения кислорода), а также ассимиляции CO2 можно разобщить.
Хлоропласты способны фотосинтезировать в организме слизня в течение нескольких месяцев, что позволяет слизню жить за счёт глюкозы, полученной в результате фотосинтеза. Фотосинтез играет ведущую роль в биосферных процессах, приводя в глобальных масштабах к образованию органического вещества из неорганического. Фотосинтезирующие организмы, используя солнечную энергию в реакциях фотосинтеза, осуществляют связь жизни на Земле со Вселенной и определяют в конечном итоге всю ее сложность и разнообразие.
Процесс фотосинтеза происходит в таких клеточных органеллах, как хлоропласты. В ходе световой стадии фотосинтеза образуются высокоэнергетические продукты: АТФ, служащий в клетке источником энергии, и НАДФ, использующийся как восстановитель. Является первым видимым продуктом фотосинтеза. В 1877 г. В. Пфеффер назвал этот процесс фотосинтезом.
alterguona.ru
конечным продуктом фотосинтеза у зеленых растений является вещество?
учебник то возьми, мозг работать будет, мож фотосинтез подействует.
В результате фотосинтеза растения строят свое тело (ствол, листва, плоды) и выделяют в окружающую среду кислород и влагу (днем) и углекислый газ (ночью) и все эти перечисленные соединения являются каждый сам по себе ВЕЩЕСТВО.
углекислый газ
touch.otvet.mail.ru
Продукты темновой стадии фотосинтеза
Образовавшиеся в процессе темновой стадии вещества делят на первичные, промежуточные и конечные. Раньше считали, что первичным продуктом фотосинтеза является крахмал. Затем, в 40-е годы прошлого века шла дискуссия, где одни ученые в качестве первичных продуктов фотосинтеза считали сахарозу, другие – фруктозу и глюкозу. В 50-е годы, когда стали использовать изотопы (радиоактивный СО2), то показали, что через несколько секунд после попадания света на лист или зеленую клетку водоросли метка у С3-растений появляется в ФГК, а затем в продуктах ее восстановления – фосфорилированных сахарах, составных компонентах цикла Кальвина. С того времени ФГК стала считаться первичным продуктом ассимиляции СО2.
Дальнейший путь превращения первичных продуктов фотосинтеза может быть разным в зависимости от вида растений, их физиологического состояния, условий питания, температуры, освещенности. Все это приводит к образованию конечных продуктов разного состава и качества. Некоторые из этих конечных продуктов (крахмал) образуется в самих хлоропластах.
Крахмал используется для синтеза разных веществ, в том числе сахарозы – главного по количеству сахара, запасаемого в растениях. Сахароза синтезируется в слое цитоплазмы, которая прилагает к хлоропласту.
Качественный состав конечных продуктов фотосинтеза зависит от спектрального состава света. Опыты с выращиванием растений при освещении только красным или только синим светом, показали, что при действии красного света увеличивается синтез крахмала; при действии синего – уменьшается. При облучении синим светом увеличивается проницаемость мембраны хлоропласта для ФГК и неорганического фосфата: ФГК выходит, а неорганический фосфат входит в хлоропласт. В результате соотношение ФГК/Фн в хлоропласте уменьшается. Это подавляет синтез крахмала. ФГК, которая вышла из хлоропласта, используется на синтез амино- и органических кислот. «Фотосинтетическое» происхождение (на основе ФГК из С3-цикла) амино- и органических кислот впервые доказано в СССР в 50-е годы А. А. Ничипоровичем и др. Минеральные элементы также влияют на качественный и количественный состав синтезируемых веществ. Например, у кукурузы (С4-растение) в условиях аммонийного питания образование малата понижается, а аспартата увеличивается.
При фотосинтезе, как мы отмечали, действуют две цепи переноса электронов: циклическое фотофосфорилирование (ФС I) и нециклическое (ФС I и ФС II). Обе системы поставляют АТФ, а ФС II еще и восстановительную силу (НАДФН) для фиксации СО2 и восстановления его до уровня углеводов. В других клеточных реакциях используется АТФ, который образуется главным образом в процессе окислительного фотофосфорилирования.
Окислительное фосфорилирование связано с процессом дыхания и осуществляется при помощи другой цепи переноса электронов, которую мы рассмотрим позже.
Постепенно, т. е. в несколько этапов, должно осуществляться не только разрушение органических молекул (так как это необходимое условие эффективного запасания и использования энергии), но и синтез сложных соединений – белков, нуклеиновых кислот, липидов или полисахаридов. Когда из простых молекул строятся более крупные, этим простым молекулам часто необходима активация; другими словами, им необходима энергия. Поэтому синтез того или иного соединения проходит более сложным путем, чем его распад.
Синтез сахарозы и полисахаридов довольно сложный процесс. Углерод, фиксированный при фотосинтезе, как уже отмечалось, превращается в фотосинтезирующих клетках в два основных углевода: сахарозу и крахмал.
Сахароза. Большая часть углеводов выносится из клетки, вероятно, в виде сахарозы. Сахара транспортируются по растению в виде дисахарида, который состоит из остатков глюкозы и фруктозы.
Сахароза транспортируется в те части растения, где она особенно нужна – к участкам быстрого роста, развивающимся семенам и клубням.

Коротко путь синтеза сахарозы у фотосинтезирующих клеток С3-растений следующий. Фруктоза-6-фосфат под влиянием специфических ферментов превращается в глюкоза-6-фосфат и глюкоза-1-фосфат, причем легко происходит и обратное превращение;

Из этих трех гексозофосфатов строятся затем цепи углеводных молекул, которые используются для транспорта, хранения и в других реакциях синтеза. Для таких превращений гексозофосфаты должны быть активированы. Это обычно достигается их присоединением к нуклеотидам – сложным кольцевым структурам, подобным адениловой кислоте.
Продуктом такой реакции присоединения являются нуклеотидные производные моносахаридов или нуклеотидсахаров. Чаще других встречается уридиндифосфатглюкоза (УДФГл), которая образуется в реакциях между уридинтрифосфатом (УТФ) и глюкоза-1-фосфатом (Гл-1-Ф). Сам УТФ образуется косвенным путем, в результате переноса фосфатной группы от АТФ на УДФ:

Нуклеотиды АТФ и УТФ присутствуют во всех клетках, потому что они используются вместе с другими нуклеотидами в синтезе ДНК и РНК. Сахароза (ГлФр) образуется в реакции между УДФГл и Фр-6-Ф:

Равновесие этой реакции сильно сдвинуто в сторону синтеза сахарозы, что обеспечивает запасание данного дисахарида в значительных концентрациях. Для последующего использования сахароза должна сначала подвергнуться расщеплению (фермент инвертаза катализирует ее гидролиз с образованием свободной глюкозы и фруктозы):

Энергия гликозидной связи в такой реакции растрачивается впустую, распределяясь между двумя молекулами. Поэтому, когда глюкозе и фруктозе предстоит распад в процессе дыхания или участие (в качестве сырья) в синтезе полисахаридов, то они должны сначала снова подвергнуться фосфорилированию за счет АТФ.
Крахмал представляет собой как бы временный запас фиксированного углевода. Он образуется во время наиболее активного периода фотосинтеза.
Длинные полимерные цепи крахмала построены из элементарных звеньев – остатков глюкозы:

Крахмал представлен двумя формами: линейным полимером – амилазой, и разветвленным полимером – амилопектином. Крахмал – основной запасной полисахарид растений. Он нерастворим в воде и откладывается слой за слоем в крахмальных зернах в хлоропластах, или накапливается в лейкопластах в тканях стебля, корней, семян. Он просто обнаруживается, поскольку способен легко окрашиваться йодом в синий цвет. Так как крахмал нерастворим в воде в отличие от сахарозы и гексоз, он не вызывает в клетках осмотического эффекта. Поэтому образование крахмала в клетках листа в периоды интенсивного фотосинтеза, не вызывает его подавления, которое происходит в результате накопления других продуктов фотосинтеза (ассимилятов). В темноте крахмал постепенно снова гидролизуется с образованием глюкозофосфата, который затем превращается в сахарозу, которая транспортируется в другие части растений.
Начальным продуктом для синтеза крахмала служит аденозиндифосфатглюкоза (АДФГл), образующаяся из АТФ и Гл-1-Ф. Молекула крахмала строится путем поэтапного присоединения одного глюкозного остатка к другому в реакции АДФГл с преобразованной глюкозной цепью:

При низком содержании глюкозы крахмал разрушается и переводится в сахарозу. Однако сначала он разрушается до остатков глюкозы и к каждому из них присоединяется остаток фосфорной кислоты, т. е. образуется Г-1-Ф, чем и обеспечивается сохранение энергии связи:
[Глюкозаn+1] + Ф → [Глюкозаn] + Гл-1-Ф.
Затем Гл-1-Ф может использоваться для синтеза сахарозы. В семенах и некоторых других органах, в которых одновременно идет разрушение больших количеств крахмала, он распадается до дисахарида мальтозы под воздействием α-амилазы. Мальтоза затем разрушается до глюкозы:

Из последней может снова синтезироваться сахароза. Для превращения глюкозы в глюкоза-6-фосфат необходима АТФ.
Целлюлоза служит основным компонентом первичной клеточной оболочки и является одним их наиболее распространенных углеводов на Земле. Молекулы целлюлозы построены подобно молекулам крахмала из остатков глюкозы, только в отличие от последнего соединены по другому:

Эти структурные отличия обусловлены тем, что два рассматриваемых полимера глюкозы (глюканы) значительно отличаются по своей природе; крахмал, например, легко переваривается в организме человека, а целлюлоза совсем не переваривается. Основная разница между крахмалом и целлюлозой состоит в том, что 1-й и 4-й углеродные атомы двух соседних остатков глюкозы соединены в крахмале α-связями, а целлюлозе – β-связями.
Отличается от крахмала и реакция синтеза целлюлозы. Роль донора глюкозы выполняет нуклеотидное производное моносахарида – гуаназиндифосфатглюкоза (ГДФГ) и связь между мономерными звеньями относится к β-типу:
ГнДА + АТФ → ГнТФ + АДФ
ГнТФ + Г-1-Ф → ГнДФГ + ФФн
ГнДФГ + [Глюкозаn] → ГнДФ + [Глюкозаn+1] целлюлоза
В некоторых случаях донором глюкозы для синтеза целлюлозы может быть и уридиндифосфатглюкоза.
У высших растений целлюлоза разрушается редко (если не считать разрушений под действием микробов). Два известных исключения из правил касаются клеток в отделительной зоне листа, образованной перед сбрасыванием листьев, и поперечных стенок сосудов ксилемы. В отделительной зоне листа фермент целлюлаза разрушает клеточные стенки, расщепляет образующуюся в них целлюлозу до отдельных мономерных единиц, т. е. до глюкозы. Клеточные оболочки, в конце концов, разрываются, и лист сбрасывается.
Целлюлозные микрофибриллы в клеточной оболочке скреплены при помощи матрикса из смешанных полисахаридных цепей (пентозы). Эти полисахариды синтезируются также из предшественников, нуклеотидсахаридов, главным образом, в диктиосомах. Пузырьки, которые отшнуровываются от диктиосом, в конце концов, сливаются с плазмалеммой и таким образом передают свое содержимое образующейся клеточной оболочке.
Все полисахариды легко переходят один в другой, но синтез их всегда идет через стадию нуклеотидсахаров, тогда как разрушение проходит более прямым путем.
biofile.ru
Первый продукт - фотосинтез - Большая Энциклопедия Нефти и Газа, статья, страница 1
Первый продукт - фотосинтез
Cтраница 1
Первый продукт фотосинтеза, согласно уравнению (3.10), является гидратом формальдегида. Теория ван Ниля не требует, однако, образования при фотосинтезе именно этого соединения, так как она также легко приложима к восстановлению более крупной молекулы ( например, кислоты с карбоксильной группой В СООН), куда С02 включается на первой ступени реакции. [1]
Если бы формальдегид был первым продуктом фотосинтеза [110], то тепловой эффект этого процесса оказался бы близок к 135 ккал на грамм-атом ассимилированного углерода. Неизвестно также, образуется ли при фотосинтезе промежуточный продукт со столь же большим содержанием энергии, как у формальдегида. [2]
Кузина показали, что в качестве первых продуктов фотосинтеза растения образуют уроновые кислоты. Это было подтверждено Е. А. Бойченко, осуществившей восстановление С02 при участии гидрогеназы изолированных хлоропластов в темноте. Эта работа имеет исключительно важное значение, освещая, во-первых, вопрос о природе одного из ферментов, участвующих в переносе водорода на С02, и, во-вторых, доказывая, что реакции восстановительного конца фотосинтеза осуществляются темновым путем. [3]
В настоящее время принимается, что первым продуктом фотосинтеза является фосфоглицериновая кислота, которая затем превращается в сахарозу, крахмал и другие углеводы. В ходе этой реакции используется только часть поглощенной световой энергии, а другая ее часть расходуется на образование фермента - восстановителя - и на выделение кислорода. Углеводы же синтезируются из СОа за счет энергии АТФ при участии восстановителя. [4]
В 1965 г. было обнаружено, что первыми продуктами фотосинтеза у сахарного тростника ( растение тропиков) являются органические кислоты, в состав которых входят 4 атома углерода ( яблочная, щевелевоуксусная и аспарагиновая), а не ЗС-кислота ( фосфоглицериновая), как у большинства растений умеренного климата. С тех пор выявили множество растений, в основном тропических и субтропических, и имеющих порой важное экономическое значение, у которых фотосинтез протекает по такому же пути. [6]
Из работ Кальвина следует, что фосфороглицериновая кислота, являясь одним из первых продуктов фотосинтеза, составляет звено замкнутого цикла превращений, в которых существенную роль играют фосфорилированные промежуточно образующиеся углеводы: рибоза и седогептоза. Этот цикл ведет к образованию щавелево-уксусной, а затем трикарбонсвых кислот. Аминирование их приводит к образованию аминокислот и синтезу белков. Другое ответвление это-го цикла ведет к синтезу жиров и углеводов. [7]
Последнее вещество, по исследованию В. Л. Кретов ича, представляет собой трифруктозан и является, очевидно, первым продуктом фотосинтеза в созревающем зерне. Зерно ржи в незрелом состоянии может содержать до 35 % ле-вулезана. Левулезан вращает плоскость поляризации влево, что не может не оказать влияния на показания поляриметра при определении крахмала. [8]
При этом образуются 2 молекулы ЗС-соединения, а именно 2 молекулы фос-фоглицерата ( ФГ), являющегося первым продуктом фотосинтеза. [10]
Они являются первым продуктом фотосинтеза зеленых растений. [12]
Последняя образуется из углекислого газа, поступающего в листы растений из воздуха. Фосфороглицериновый альдегид является первым продуктом фотосинтеза. Дальнейшие превращения этого альдегида приводят к образованию углеводов. Все эти процессы очень сложны, они протекают под влиянием ферментов. Фотосинтез является единственным процессом в природе, за счет которого органические вещества образуются из неорганических. [13]
Эксперименты показали, что все превращения в растениях происходят очень быстро. Углеводы и белки возникают прямо в зеленых хлоропластах. Ученые уже убедились, что прежнее мнение о первичной реакции фотосинтеза было ошибочным. Первым продуктом фотосинтеза является не формальдегид. В листьях растений образуются гораздо более сложные вещества, а на конечной стадии - простые сахара. [14]
Таким образом, можно сделать вывод, что ФГК является первым стабильным продуктом фиксации углекислоты. Последующие исследования были проведены с целью выяснить природу молекулы акцептора. Простое объяснение, согласно которому двууглеродная молекула акцептирует углекислоту, образуя ФГК, не подтвердилось. Среди первых продуктов фотосинтеза были обнаружены фосфаты С5 - и С7 - сахаров, и этот факт вместе с открытием новых ферментативных превращений указанных соединений ( см. Пентозофосфат-ный путь, стр. [15]
Страницы: 1 2
www.ngpedia.ru
Параграф 27. Фотосинтез
1. Фотосинтез относится к процессам пластического или энергетического обмена? Почему?
Фотосинтез относится к процессам пластического обмена. Так как в процессе фотосинтеза происходит образование органических веществ из углекислого газа и воды.
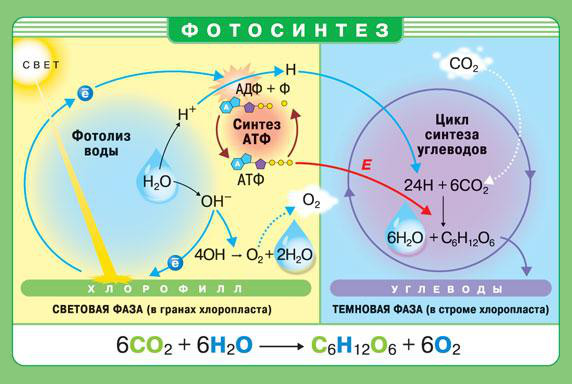
2. В каких органоидах растительной клетки происходит фотосинтез? Что представляет собой фотосистема? Какую функцию выполняют фотосистемы?
У растений и водорослей фотосинтез осуществляется в специальных органоидах — хлоропластах. В мембранах тилакоидов расположены особые пигмент-белковые комплексы — фотосистемы. Существует два типа фотосистем — фотосистема I и фотосистема II. В состав каждой входит светособирающая антенна, образованная молекулами пигментов, реакционный центр и переносчики электронов. Главное различие фотосистем заключается в том, что в состав фотосистемы II входит особый ферментный комплекс, осуществляющий на свету фотолиз воды — расщепление молекул воды с образованием кислорода (О2), электронов и протонов (Н). В фотосистеме I отсутствует подобный ферментный комплекс, и, следовательно, она не способна использовать воду в качестве источника электронов для восстановления своей молекулы-ловушки.
3. Каково значение фотосинтеза на Земле? Почему без фототрофных организмов существование биосферы было бы невозможным?
Уникальность и биологическое значение фотосинтеза определяются тем, что жизнь на нашей планете всем своим существованием обязана этому процессу. Фотосинтез является основным источником питательных веществ для живых организмов, а также единственным поставщиком свободного кислорода на Земле. Из кислорода сформировался и поддерживается озоновый слой, защищающий живые организмы Земли от губительного воздействия коротковолнового ультрафиолетового излучения. Кроме того, благодаря фотосинтезу поддерживается относительно постоянное содержание СО2 в атмосфере.
4. Охарактеризуйте световую и темновую фазы фотосинтеза по плану: 1) место протекания; 2) исходные вещества; 3) происходящие процессы; 4) конечные продукты. Какие продукты световой фазы фотосинтеза используются в темновой фазе?
Световая фаза осуществляется на мембранах тилакоидов и только при наличии света. Реакции темновой фазы протекают в строме хлоропласта и не требуют света, однако для их прохождения необходимы продукты световой фазы. Поэтому темновая фаза идет практически одновременно со световой. Исходные вещества в световой фазе - восстановленный НАДФ, темновая фаза – углекислый газ и восстановленный НАДФ. Таким образом, в ходе световой фазы энергия света поглощается и преобразуется в энергию макроэргических связей АТФ, происходит расщепление воды с выделением кислорода и накопление атомов водорода. Продуктами световой фазы фотосинтеза являются АТФ, восстановленный НАДФ и кислород. Кислород — побочный продукт фотосинтеза, он выделяется в окружающую среду. Во время темновой фазы из окружающей среды в хлоропласты поступает углекислый газ, а в строме хлоропластов происходит его восстановление до органических веществ. Конечными продуктами является глюкоза и кислород.
5. Сравните фотосинтез и аэробное дыхание. Укажите черты сходства и различия.
Различие: в процессе фотосинтеза происходит синтез глюкозы, а в процессе аэробного дыхания ее расщепление. Оба процесса происходят с участием кислорода.
6. Человек за сутки потребляет примерно 430 г кислорода. Дерево средней величины поглощает около 30 кг углекислого газа в год. Сколько деревьев необходимо, чтобы обеспечить одного человека кислородом?
Из расчета, что при потреблении 430 г кислорода выделяется 430 г углекислого газа. В год человек выделяет 430*365=154 800 г в год, что равно 154,8 кг в год. Если 1 дерево поглощает 30 кг углекислого газа в год, то 154,8 кг/30 кг= 5,1 деревьев на одного человека.
7. Исследователи разделили растения пшеницы на две группы и выращивали их в лаборатории в одинаковых условиях, за исключением того, что растения первой группы освещали красным светом, а растения второй группы — зеленым. У растений какой группы фотосинтез протекал более интенсивно? С чем это связано?
Пшеница содержит молекулы хлорофилла, которые поглощают красный и синий лучи спектра и отражают зеленые (поэтому растения имеют зеленую окраску). Значит у той группы растений, которых освещали красными светом фотосинтез протекал более интенсивно, так как хлорофилл может поглощать лучи данного спектра и полученную энергию тратить на образование органических веществ.
8. С помощью какого эксперимента можно доказать, что кислород, выделяющийся при фотосинтезе, образуется именно из молекул воды, а не из молекул углекислого газа или какого-либо другого вещества?
Можно применить метод меченных атомов, когда атомы кислорода в воде будут помечены веществами-метками, а затем данные атомы будут прослеживаться в молекулах кислорода.
resheba.com








