Срочно!!! где образуются лизосомы? как их может использовать клетка? Есть ли в клетках растений лизосомы
Лизосома — Википедия
Материал из Википедии — свободной энциклопедии
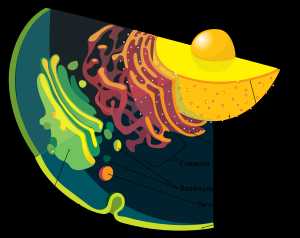 Эндомембранная система эукариотической клетки.
Эндомембранная система эукариотической клетки. 
Лизосо́ма (от греч. λύσις — растворяю и σώμα — тело) — окружённый мембраной клеточный органоид, в полости которого поддерживается кислая среда и находится множество растворимых гидролитических ферментов[1]. Лизосома отвечает за внутриклеточное переваривание макромолекул, в том числе при аутофагии; лизосома способна к секреции своего содержимого в процессе лизосомного экзоцитоза; также лизосома участвует в некоторых внутриклеточных
ru.wikipedia.org
Срочно!!! где образуются лизосомы? как их может использовать клетка?
1.Эукариоты - организмы (все, кроме бактерий, включая цианобактерии) , обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключен в хромосомах. Клетки эукариоты имеют митохондрии, пластиды и другие органоиды. Характерен половой процесс. Представители: грибы, животные, растения. 2. Фагоцитоз - процесс, при котором специальные клетки крови и тканей организма (фагоциты) захватывают и переваривают возбудителей инфекционных заболеваний и отмершие клетки. Осуществляется двумя разновидностями клеток: циркулирующими в крови зернистыми лейкоцитами (гранулоцитами) и тканевыми макрофагами.3. Эндоплазматический ретикулум (ЭПР) или эндоплазматическая сеть (ЭПС) - внутриклеточный органоид эукариотической клетки, представляющий собой разветвлённую систему из окружённых мембраной уплощённых полостей, пузырьков и канальцев. При участии эндоплазматического ретикулума происходит трансляция и транспорт белков, синтез и транспорт липидов и стероидов. Для ЭПС характерно также накопление продуктов синтеза. Эндоплазматический ретикулум принимает участие в том числе и в создании новой ядерной оболочки (например после митоза). Эндоплазматический ретикулум содержит внутриклеточный запас кальция, который является, в частности, медиатором сокращения мышечной клетки. В клетках мышечных волокон расположена особая форма эндоплазматического ретикулума — саркоплазматическая сеть. https://ru.wikipedia.org/wiki/Эндоплазматический_ретикулум#/media/File:Biological_cell.svg4. Фотоси́нтез - процесс преобразования энергии света в энергию химических связей органических веществ на свету фотоавтотрофами при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл и бактериородопсин у бактерий). Процесс фотосинтеза осуществляют зеленые растения. В хлоропластах. 5. Лизосомы - органоиды грибов и животных, отсутствующие в клетках растений. Они представляют собой пузырьки, окруженные мембраной. Образуются в комплексе Гольджи . Содержат набор гидролитических ферментов. С их помощью осуществляется внутриклеточное пищеварение. У растений функции лизосом сравнимы с некоторыми функциями вакуолей .otvet.mail.ru
2.5. Лизосомы. Митохондрии. Пластиды | ВСЕ Сочинения.RU
Вопрос 1. Где формируется лизосома?
Лизосомы — мембранные структуры, содержащие множество активных ферментов, участвующих в расщеплении высокомолекулярных соединений: белков, липидов, углеводов. Лизосомы образуются в комплексе Гольджи, куда поступают и где накапливаются ферменты.
Вопрос 2. Какова функция митохондрий?
Митохондрии — клеточные структуры, покрытые двойной мембраной. На внутренней мембране, имеющей многочисленные выросты, расположено огромное количество ферментов, принимающих участие в синтезе АТФ. Следовательно, главная функция митохондрий — обеспечение клетки энергией за счет синтеза АТФ.
Вопрос 3. Какие виды пластид вы знаете?
Различают три вида пластид — лейкопласты, хромопласты и хлоропласты.
Лейкопласты — бесцветные пластиды, которые располагаются в органах растений, недоступных для солнечного света (например, в корневищах, клубнях). На свету в них образуется хлорофилл.
Хромопласты — пластиды, содержащие желтый, оранжевый, красный и фиолетовый пигменты. Они расположены в основном в плодах и лепестках цветков, что придает этим органам растений соответствующую яркую окраску.
Хлоропласты — зеленые пластиды, содержащие хлорофилл и участвующие и фотосинтезе.
Вопрос 4. Чем отличается каждый вид пластид от другого?
Пластиды разных видов отличаются друг от друга наличием или отсутствием тех или иных пигментов. В лейкопластах пигменты отсутствуют, в хлоропластах содержится зеленый пигмент, а в хромопластах — красный, оранжевый, желтый и фиолетовый пигменты.
Вопрос 5. Почему граны в хлоропласте расположены в шахматном порядке?
Граны в хлоропластах расположены в шахматном порядке для того, чтобы не загораживать друг друга от солнечных лучей. Солнечный свет должен хорошо освещать каждую грану, тогда фотосинтез будет протекать более интенсивно.
Вопрос 6. Что будет, если лизосома в одной из клеток внезапно разрушится?
При внезапном разрыве мембраны, окружающей лизосому, содержащиеся в ней ферменты попадают в цитоплазму и постепенно разрушают всю клетку.
Вопрос 7. В чем сходство митохондрий и пластид?
Во-первых, сходство митохондрий и пластид заключается в том, что они имеют двухмембранное строение.
Во-вторых, эти органоиды содержат собственные молекулы ДНК, поэтому способны самостоятельно размножаться, независимо от деления клетки.
В-третьих, можно отметить, что и в тех и в других синтезируется АТФ (в митохондриях — при расщеплении белков, липидов и углеводов, а в хлоропластах — за счет превращения солнечной энергии в химическую).
2.5. Лизосомы. Митохондрии. Пластиды
5 (99.2%) 25 votesНа этой странице искали :
- где формируется лизосома
- лизосомы формируются на
- где формируются лизосомы
- чем отличается каждый вид пластид от другого
- где образуются лизосомы
Сохрани к себе на стену!
vsesochineniya.ru
Реферат Лизосома
скачатьРеферат на тему:
План:
- Введение
- 1 Распространенность среди царств живой природы
- 2 Признаки лизосом
- 3 Образование лизосом и их типы
- 4 Функции лизосом
- 5 Внутриклеточное пищеварение и участие в обмене веществ
- 6 Аутофагия
- 7 Автолиз
- 8 Клиническое значение. Болезни, связанные с нарушением работы лизосом
Введение
Лизосома — (от греч. λύσις — растворяю и sōma — тело) клеточный органоид размером 0,2 — 0,4 мкм, один из видов везикул. Эти одномембранные органоиды — часть вакуома (эндомембранной системы клетки). Разные виды лизосом могут рассматриваться как отдельные клеточные компартменты.
1. Распространенность среди царств живой природы
Лизосомы были впервые описаны в 1955 году Кристианом де Дювом в животной клетке, а позже были обнаружены и в растительной. У растений к лизосомам по способу образования, а отчасти и по функциям близки вакуоли. Лизосомы есть также у большинства протистов (как с фаготрофным, так и с осмотрофным типом питания) и у грибов. Таким образом, наличие лизосом характерно для клеток всех эукариот. У прокариот лизосомы отсутствуют, так как у них отсутствует фагоцитоз и нет внутриклеточного пищеварения.
2. Признаки лизосом
Один из признаков лизосом — наличие в них ряда ферментов (кислых гидролаз), способных расщеплять белки, углеводы, липиды и нуклеиновые кислоты. К числу ферментов лизосом относятся катепсины (тканевые протеазы), кислая рибонуклеаза, фосфолипаза и др. Кроме того, в лизосомах присутствуют ферменты, которые способны отщеплять от органических молекул сульфатные (сульфатазы) или фосфатные (кислая фосфатаза) группы.
Для лизосом характерна кислая реакция внутренней среды. Обычно рН в лизосомах составляет около 4,5-5 (концентрация протонов на два порядка выше, чем в цитоплазме). Это обеспечивается активным транспортом протонов, который осуществляет встроенный в мембраны лизосом белок-насос протонная АТФаза.
Высокая активность кислой фосфатазы ранее использовалась как один из маркеров лизосом. В настоящее время более надежным маркером считается присутствие специфических мембранных гликопротеидов — LAMP1 и LAMP2. Они присутствуют на мембране лизосом и поздних эндосом, но отсутствуют на мембранах других компартментов вакуома.
3. Образование лизосом и их типы
Лизосомы формируются из пузырьков (везикул), отделяющихся от аппарата Гольджи, и пузырьков (эндосом), в которые попадают вещества при эндоцитозе. В образовании аутолизосом (аутофагосом) принимают участие мембраны эндоплазматического ретикулума. Все белки лизосом синтезируются на «сидячих» рибосомах на внешней стороне мембран эндоплазматического ретикулума и затем проходят через его полость и через аппарат Гольджи.
Лизосомы — гетерогенные органеллы, имеющие разную форму, размеры, ультраструктурные и цитохимические особенности. «Типичные» лизосомы животных клеток обычно имеют размеры 0,1-1 мкм, сферическую или овальную форму. Число лизосом варьирует от одной (крупная вакуоль во многих клетках растений и грибов) до нескольких сотен или тысяч (в клетках животных).
Общепринятой классификации и номенклатуры для разных стадий созревания и типов лизосом нет. Различают первичные и вторичные лизосомы. Первые образуются в области аппарата Гольджи, в них находятся ферменты в неактивном состоянии, вторые же содержат активные ферменты. Обычно ферменты лизосом активируются при понижении рН. Среди лизосом можно также выделить гетеролизосомы (переваривающие материал, поступающий в клетку извне — путем фаго- или пиноцитоза) и аутолизосомы (разрушающие собственные белки или органоиды клетки). Наиболее широко используется следующая классификация лизосом и связанных с ними компартментов:
- Ранняя эндосома — в нее поступают эндоцитозные (пиноцитозные) пузырьки. Из ранней эндосомы рецепторы, отдавшие (из-за пониженного рН) свой груз, возвращаются на наружную мембрану.
- Поздняя эндосома — в нее из ранней эндосомы поступают пузырьки с материалом, поглощенном при пиноцитозе, и пузырьки из аппарата Гольджи с гидролазами. Рецепторы маннозо-6-фосфата возвращаются из поздней эндосомы в аппарат Гольджи.
- Лизосома — в нее из поздней эндосомы поступают пузырьки со смесью гидролаз и перевариваемого материала.
- Фагосома — в нее попадают более крупные частицы (бактерии и т. п.), поглощенные путем фагоцитоза. Фагосомы обычно сливаются с лизосомой.
- Аутофагосома — окруженный двумя мембранами участок цитоплазмы, обычно включающий какие-либо органоиды и образующийся при макроаутофагии. Сливается с лизосомой.
- Мультивезикулярные тельца — обычно окружены одинарной мембраной, содержат внутри более мелкие окруженные одинарной мембраной пузырьки. Образуются в результате процесса, напоминающего микроаутофагию (см. ниже), но содержат материал, полученный извне. В мелких пузырьках обычно остаются и затем подвергаются деградации рецепторы наружной мембраны (например, рецепторы эпидермального фактора роста). По стадии формирования соответствуют ранней эндосоме. Описано образование мультивезикулярных телец, окруженных двумя мембранами, путем отпочковывания от ядерной оболочки.
- Остаточные тельца (телолизосомы) — пузырьки, содержащие непереваренный материал (в частности, липофусцин). В нормальных клетках сливаются с наружной мембраной и путем экзоцитоза покидают клетку. При старении или патологии накапливаются.
4. Функции лизосом
Функциями лизосом являются:
- переваривание захваченных клеткой при эндоцитозе веществ или частиц (бактерий, других клеток)
- аутофагия — уничтожение ненужных клетке структур, например, во время замены старых органоидов новыми, или переваривание белков и других веществ, произведенных внутри самой клетки
- автолиз — самопереваривание клетки, приводящее к ее гибели (иногда этот процесс не является патологическим, а сопровождает развитие организма или дифференцировку некоторых специализированных клеток). Пример: При превращении головастика в лягушку, лизосомы, находящиеся в клетках хвоста, переваривают его: хвост исчезает, а образовавшиеся во время этого процесса вещества всасываются и используются другими клетками тела.
5. Внутриклеточное пищеварение и участие в обмене веществ
У многих протистов и у животных, имеющих внутриклеточное пищеварение, лизосомы участвуют в переваривании пищи, захваченной путем эндоцитоза. При этом лизосомы сливаются с пищеварительными вакуолями. У протистов непереваренные остатки пищи обычно удаляются из клетки при слиянии пищеварительной вакуоли с наружной мембраной.
Многие клетки животных, у которых преобладает полостное пищеварение (например, хордовые) получают питательные вещества из межклеточной жидкости или плазмы крови с помощью пиноцитоза. Эти вещества также вовлекаются в обмен веществ клетки после их переваривания в лизосомах. Хорошо изученный пример такого участия лизосом в обмене веществ — получение клетками холестерина. Холестерин, приносимый кровью в виде ЛПНП, поступает внутрь пиноцитозных везикул после соединения ЛПНП с рецепторами ЛПНП на мембране. Рецепторы возвращаются к мембране из ранней эндосомы, а ЛПНП поступают в лизосомы. После этого ЛПНП перевариваются, а высвободившийся холестерин через мембрану лизосом поступает в цитоплазму.
Косвенно лизосомы участвуют в обмене, обеспечивая десенсибилизацию клеток к воздействию гормонов. При длительном действии гормона на клетку часть рецепторов, связавших гормон, поступают в эндосомы и затем деградируют внутри лизосом. Снижение числа рецепторов понижает чувствительность клетки к гормону.
Для крупных вакуолей растений характерна запасающая функция — в них могут накапливаться ионы, пигменты (например, антоцианы), вторичные метаболиты, белки (в алейроновых зернах эндосперма злаков). Внутри вакуолей (например, в прорастающих семенах) у растений происхдят и процессы переваривания запасенных белков.
6. Аутофагия
Обычно различают два типа аутофагии — микроаутофагия и макроаутофагия. При микроаутофагии, как при образовании мультивезикулярных телец, образуются впячивания мембраны эндосомы или лизосомы, которые затем отделяются в виде внутренних пузырьков, только в них попадают вещества, синтезированные в самой клетке. Таким путем клетка может переваривать белки при нехватке энергии или строительного материала (например, при голодании). Но процессы микроаутофагии происходят и при нормальных условиях и в целом неизбирательны. Иногда в ходе микроаутофагии перевариваются и органоиды; так, у дрожжей описана микроаутофагия пероксисом и частичная микроаутофагия ядер, при которой клетка сохраняет жизнеспособность.
При макроаутофагии участок цитоплазмы (часто содержащий какие-либо органоиды) окружается мембранным компартментом, похожим на цистерну эндоплазматической сети. В результате этот участок оказывается отгорожен от остальной цитоплазмы двумя мембранами. Затем такая аутофагосома сливается с лизосомой, и ее содержимое переваривается. Видимо, макроаутофагия также неизбирательна, хотя часто подчеркивается, что с помощью нее клетка может избавляться от «отслуживших свой срок» органоидов (митохондрий, рибосом и др.).
Третий тип аутофагии — шаперон-зависимая. При этом способе происходит направленный транспорт частично денатурировавших белков из цитоплазмы сквозь мембрану лизосомы в ее полость.
7. Автолиз
Ферменты лизосом нередко высвобождаются при разрушении мембраны лизосомы. Обычно при этом они инактивируются в нейтральной среде цитоплазмы. Однако при одновременном разрушении всех лизосом клетки может произойти ее саморазрушение — автолиз. Различают патологический и обычный автолиз. Распространенный вариант патологического автолиза — посмертный автолиз тканей.
В норме процессы автолиза сопровождают многие явления, связанные с развитием организма и дифференцировкой клеток. Так, аутолиз клеток описывается как механизм разрушения тканей у личинок насекомых при полном превращении, а также при рассасывании хвоста у головастика. Правда, эти описания относятся к периоду, когда различия между апоптозом и некрозом еще не были установлены, и в каждом случае требуется выяснять, не лежит ли на самом деле в основе деградации органа или ткани апоптоз, не связанный с автолизом.
У растений автолизом сопровождается дифференциация клеток, которые функционируют после смерти (например, трахеид или члеников сосудов). Частичный автолиз происходит и при созревании клеток флоэмы- члеников ситовидных трубок.
8. Клиническое значение. Болезни, связанные с нарушением работы лизосом
Иногда из-за неправильной работы лизосом развиваются болезни накопления, при которых ферменты из-за мутаций не работают или работают плохо. Примером болезней накопления может служить амавротическая идиотия при накоплении гликогена.
Разрыв лизосомы и выход в гиалоплазму расщепляющих ферментов сопровождается резким повышением их активности. Такого рода повышение активности ферментов наблюдается, например, в очагах некроза при инфаркте миокарда и при действии излучения.
wreferat.baza-referat.ru
Вакуолярная система клеток растений
Лизосомы, классификация, строение и значение
Лизосомы (греч. “лизео” - растворяю, “сома” - тело). Лизосомы были открыты в 1955 г. французским ученым де Дювом при изучении клеток печени методом фракционного центрифугирования. В дальнейшем применение биохимических, цитохимических и электронно-микроскопических методов позволило детально изучить строение, функционирование и происхождение этих структур. Лизосомы представляют собой пузырьки диаметром от 0,2 до 1 мкм, содержащие различные ферменты. Всего в лизосомах обнаружено около 50 гидролитических ферментов(протеиназы, нуклеазы, глюкозидазы, фосфатазы, липазы и др.). Маркерным ферментом является кислая фосфатаза. Это самые мелкие мембранные органоиды, встречающиеся практически у всех эукариот. Однако частота встречаемости лизосом варьирует в зависимости от типа клеток. Так в тканях животных лизосомы чаще встречаются в клетках, поглощающих белковые и другие вещества. Это клетки кровеносной системы, клетки печени и почек.
Образование лизосом происходит за счет деятельности эндоплазматической сети и комплекса Гольджи. Основная функция их заключается в участии в процессах внутриклеточного расщепления как экзогенных, так и эндогенных биологических макромолекул.
Среди лизосом можно выделить три основных типа: первичные лизосомы, вторичные лизосомы, остаточные тельца.
Первичные лизосомы. В первичных лизосомах содержатся неактивные ферменты. Эти ферменты синтезируются в гранулярной эндоплазматической сети. Затем они поступают в комплекс Гольджи, где упаковываются в мелкие мембранные пузырьки – первичные лизосомы.
Вторичные лизосомы. При соединении первичной лизосомы с фагоцитарными или пиноцитозными вакуолями образуются вторичные лизосомы. При этом содержимое первичной лизосомы сливается с содержимым эндоцитозной вакуоли. В дальнейшем под действием ферментов первичной лизосомы поглощенный материал постепенно расщепляется до мономеров. Мономеры транспортируются через мембрану лизосомы в цитоплазму, где они включаются в метаболизм клетки. Например, при введении в организм мыши чужеродного белка пероксидазы наблюдается накопление его в эндоцитозных вакуолях. С этими вакуолями соединяются первичные лизосомы, обладающие кислой фосфатазой. В образовавшейся новой вакуоли обнаруживается как пероксидазная, так и фосфатазная активность.
Разновидностью вторичных лизосом являются аутолизосомы (аутофагосомы), которые постоянно встречаются в клетках простейших, растений и животных. В отличие от обычных вторичных лизосом в аутолизосомах встречаются фрагменты или даже целые цитоплазматические структуры (митохондрии, пластиды, рибосомы и т. д.). Функциональное значение их заключается в уничтожении дефектных структур клетки. Число аутофагосом возрастает при метаболических стрессах и различных повреждениях клеток.
Остаточные тельца, или телолизосомы. В ряде случаевпереваривание биогенных макромолекул внутри лизосом может идти не до конца. В этом случае в полостях лизосом накапливаются непереваренные продукты. Такие лизосомы называют остаточными тельцами. Судьба остаточных телец может быть различной: одни из них выбрасываются из клетки путем экзоцитоза, другие же остаются в клетках вплоть до их гибели (например, липофусциновые гранулы). Так у простейших остаточные тельца выделяются во внешнюю среду. У человека при старении организма в остаточных тельцах клеток мозга, печени, мышечных волокон накапливается «пигмент старения» - липофусцин. Остаточные тельца содержат меньше гидролитических ферментов, в них происходит уплотнение содержимого, его перестройка. Часто в остаточных тельцах наблюдается вторичная структуризация неперевариваемых липидов, которые образуют слоистые структуры.
Функции лизосом
1. Внутриклеточное пищеварение. Это основная функция лизосом. За эту функцию лизосомы часто называют "пищеварительными станциями" клетки.
2. Изменение клеточных продуктов. Например, благодаря лизосомам, в клетках щитовидной железы происходит преобразование тироглобулина в тироксин.
3. Переваривание дефектных клеточных органоидов. В некоторых случаях лизосомы также могут переварить отдельные органы. Например, исчезновение хвоста у головастика лягушек происходит под действием ферментов лизосом.
Вакуолярная система клеток растений
Вакуоли (от лат. vacuus – пустой)– полости в цитоплазме растительных клеток,ограниченные мембраной и заполненные жидкостью. Вакуоли содержатся в клетках как низших, так и высших растений. В последнем случае они достигают наибольшего развития. Вакуоли образуются в эндоплазматической сети и из пузырьков комплекса Гольджи. В молодых клетках вакуоли представлены системой мелких провакуолей. По мере роста и дифференцировки клетки провакуоли сливаются в одну или несколько крупных центральных вакуолей, занимающих основной объем клетки. В результате цитоплазма с ядром и органоидами оттесняется к периферии клетки. В зависимости от типа и возраста клеток вакуоли занимают от 5 до 95% объема клетки. Чаще всего это величина составляет свыше 50% объема клетки. От цитоплазмы центральные вакуоли отделены тонкой (около 10 нм) одинарной мембраной, которая называется тонопласт. Полость вакуоли заполнена клеточным соком. Клеточный сок представляет собой водный раствор, в котором находятся различные неорганические соли, сахара, органические кислоты и их соли, а также другие низкомолекулярные и высокомолекулярные соединения.
В растительной клетке вакуоли выполняют многообразные функции:
· Регуляция водно-солевого обмена.
· Поддержание тургорного давления в клетке. Тургор – это напряженное состояние клеточной оболочки, создаваемое гидростатическим давлением внутриклеточной жидкости. В сочетании с жесткой клеточной стенкой тургорное давление придает растительным клеткам большую прочность. Это позволяет достигать им больших размеров по сравнению с клетками животных. Все процессы увядания, автолиза и старения сопровождаются снижением тургорного давления.
· Накопление различных веществ. Из органических веществ в вакуолях накапливаются углеводы и белки. Запасание белков особенно характерно для семян. Происходит оно в алейроновых вакуолях, которые после заполнения белками обезвоживаются и превращаются в твердые алейроновые зерна. При прорастании семян эти зерна обводняются и снова превращаются в вакуоли. Из неорганических веществ в вакуолярном соке накапливаются фосфаты калия, натрия, кальция, могут накапливаться соли органических кислот (оксалаты, цитраты и др.) Это придает вакуолярному соку кислую реакцию (рН от 2 до 5). В вакуолях могут накапливаться «красящие» вещества. Так антоцианы придают окраску лепесткам цветков, что способствует привлечению насекомых-опылителей. Возможно также накопление ядовитых веществ, которые предохраняют их от поедания травоядными животными. При повреждении клеток эти вещества высвобождаются из вакуолей и оказывают негативное действие на организм животного.
· Выведение экскреторных веществ. Вакуоли используются не только для накопления различных веществ, но и для экскреции их из клетки. Так выводятся из клетки все водорастворимые метаболиты. Нерастворимые в воде органические компоненты могут превращаться в растворимые глюкозиды, соединяясь с молекулами сахаров.
mykonspekts.ru
Опишите лизосомы, как участников клеточного метаболизма. Какова их биологическая роль в клетке?
Лизосомы (от lysis – расщепление и soma – тело) обнаруживаются у большинства эукариотических клеток, но особенно много их в тех животных клетках, которые обладают способностью к фагоцитозу. Они представляют собой простые мембранные мешочки (стенка мешочка состоит из одинарной мембраны), наполненные гидролитическими (пищеварительными) ферментами-протеазами, нуклеазами, липазами и кислыми фосфатазами. Содержимое лизосом имеет кислую реакцию, и для лизосомных ферментов характерен низкий оптимум рН. Эти ферменты должны быть изолированы от всех остальных клеточных компонентов и структур, иначе они их разрушат. В животных клетках лизосомы обычно имеют округлую форму и диаметр от 0,2 до 0,5 мкм. На электронных микрофотографиях лизосомы представляются гомогенными.
В растительных клетках роль лизосом могут играть крупные центральные вакуоли. Впрочем, иногда в цитоплазме, особенно в погибающих клетках, бывают видны тельца, напоминающие по своему виду лизосомы животных клеток. Большая часть работ, посвященных лизосомам, выполнена на животных клетках.
Заключенные в лизосомах ферменты синтезируются на шероховатом ЭПР и транспортируются к аппарату Гольджи. Позже от него отпочковываются пузырьки Гольджи, содержащие ферменты, подвергшиеся необходимым превращениям. Такие пузырьки называются первичными лизосомами. Они выполняют ряд функций, связанных главным образом с внутриклеточным перевариванием, но иногда и с секрецией пищеварительных ферментов.
Переваривание материалов, поглощенных путем эндоцитоза. С пузырьками или вакуолями, образовавшимися в процессе эндоцитоза, могут сливаться первичные лизосомы. При этом образуются вторичные лизосомы, в которых происходит переваривание материалов, поступивших в клетку путем эндоцитоза. У некоторых простейших, например у амеб, эндоцитоз – это способ поглощения пищи. В других случаях он выполняет защитную функцию, когда, например, специализированные лейкоциты (фагоциты) и макрофаги захватывают и переваривают попавшие в организм бактерии. Вторичную лизосому можно назвать также пищеварительной вакуолью. Продукты переваривания поглощаются и усваиваются цитоплазмой клетки, но часть материала так и остается непереваренной. Вторичная лизосома, содержащая этот непереваренный материал, называется остаточным тельцем. Остаточные тельца направляются обычно к плазматической мембране, н здесь их содержимое выводится наружу (экзоцитоз). В некоторых клетках, в частности в клетках сердечной мышцы и в клетках печени, остаточные тельца сохраняются.
Своеобразную роль играют лизосомы в клетках щитовидной железы, которые под действием тиреотропного гормона (ТТГ) поглощают путем пиноцитоза тиреоглобулин. Образовавшиеся пиноцитозные пузырьки сливаются с первичными лизосомами, и тиреоглобулин подвергается частичному гидролизу для того, чтобы превратиться в активный гормон тироксин. Только после этого лизосомы путем слияния с плазматической мембраной изливают свое содержимое наружу – выделяют этот гормон в кровь.
Автофагия – процесс, посредством которого клетка уничтожает ненужные ей структуры. Сначала эти структуры окружаются одинарной мембраной, отделяющейся обычно от гладкого ЭПР, а затем такой мембранный мешочек с заключенной в нем структурой сливается с первичной лизосомой, в результате чего образуется вторичная лизосома, или автофагическая вакуоль, в которой структура переваривается. Данная последовательность событий входит как составная часть в естественный круговорот цитоплазматических органелл, при котором старые органеллы заменяются новыми. Автофагия наблюдается чаще в клетках, претерпевающих реорганизацию во время дифференцировки.
Выделение ферментов из клетки (экзоцитоз). Иногда ферменты, содержащиеся в первичных лизосомах, выделяются из клетки наружу. Это происходит, например, при замене хряща костной тканью в процессе развития. Аналогичное явление можно наблюдать, когда основное вещество кости разрушается при перестройке костной ткани в ответ на повреждения, при новых нагрузках и т.п. В этом случае ферменты секретируются лизосомами клеток, которые называются остеокластами.
Автолиз – это саморазрушение клетки, наступающее в результате высвобождения содержимого ее лизосом. Именно в связи с этим лизосомы были в свое время метко названы «орудиями самоубийства» (suicide bags). При некоторых процессах дифференцировки автолиз представляет собой нормальное явление; он может распространяться и на всю ткань, как это, например, имеет место при резорбции хвоста головастика во время метаморфоза. Автолиз наступает также после гибели клетки. Иногда он является следствием некоторых лизосомных болезней или результатом повреждения клетки.
34. Каковы основные молекулярные механизмы самосохранения биосистем? Что такое генный код? В чём заключается его универсальность? Укажите основные стадии биосинтеза белков.
Продолжительность жизни организмов ограничена, однако все они обладают способностью непрерывно «поддерживать жизнь», обеспечивая выживание вида. Вид выживает в результате того, что родители передают потомству свои основные признаки, независимо от того, возникло ли потомство в результате полового или бесполого размножения. В поисках причин, обусловливающих такую передачу признаков (наследование), были открыты нуклеиновые кислоты — ДНК (дезоксирибонуклеиновая кислота) и РНК (рибонуклеиновая кислота) В молекулах этих кислот содержится закодированная информация, передающаяся от одного поколения организмов другому.
Нуклеиновые кислоты – высокомолекулярные биологические полимеры (полинуклеотиды), структурной ед является мононуклеотид (сост из : пятиуглеродного сахара; азотистого основания пуринового - А,Г и пиримидинового - Ц,Т/У; фосфорной кислоты).
Ген – участок ДНК, кодирующий определённый пептид (белок).
Генетический код – это соответствие м/д последовательностью нуклеотидов в молекуле ДНК и последовательностью аминокислот в синтезируемом белке.
Свойства и характеристики ген.кода:
-триплетность: каждая аминок-та кодируется тройкой нуклеотидов.
-код универсален: одни и те же триплеты кодируют одни и те же аминокислоты у всех организмов
-код вырожденный: данная аминок-та может кодироваться более чем одним триплетом.
-код неперекрывающийся: один и тот же нуклеотид не может входить в состав двух соседних триплетов
Биосинтез белка.
I. Транскрипция – переписывание последовательности оснований в одном из участков цепи ДНК в комплементарную последовательность оснований мРНК. Протекает внутри ядра (у прокариот), имеет 3 стадии:
1. для инициации транскрипции ДНК необходимо наличие специального участка в ДНК (промотора). Фермент РНК-полимераза связывается с промотором и происходит локальное расщепление двойной спирали ДНК => образуется открытый промоторный участок (фермент действует как застёжка молнии). Одна из цепей является матрицей для образования комплементарной одиночной цепи мРНК.
2. элонгация – удлинение цепи
3. терминация – прекращение роста мРНК происходит на специфических участках ДНК (терминаторах)
Образованная незрелая мРНК (пре-иРНК) подвергается процессингу и сплайсингу
Процессинг: кэпирование 5’-конца пре-иРНК, т.е. к 5’-концу присоединяется 5’-кэп-структура (защищает мРНК от ферментативного расщепления, способствует трансляции иРНК) ; к 3’-концу пре-иРНК присоединяется 3’-poly А – хвост
Сплайсинг – удаление интронов (структурные гены, которые связывают экзоны – гены, несущие информацию о первичной структуре белка)
II. Трансляция – процесс, посредством которого последовательность оснований в молекуле РНК переводится в последовательность аминокислот в полипептидной цепи. Этот процесс происходит на рибосомах. Комплекс мРНК и нескольких рибосом назыв-ся полисомой. Трансляция протекает в направлении 5’ → 3’ концу и имеет3 стадии:
1. Инициация. Считывание информации с мРНК. Первый кодон связывает молекулу аминоацил-тРНК, содержащую комплементарный ему антикодон и несущую первую аминокислоту (обычно это метионин) синтезируемого полипептида. Затем второй кодон присоединяет к себе молекулу аминоацил-тРНК, содержащую комплементарный ему антикодон. Функция рибосомы в том, чтобы удерживать в нужном положении мРНК, тРНК и ферменты до тех пор, пока м/д соседними аминокислотами не образуется пептидная связь.
2. элонгация – удлинение полипептидной цепи
3. терминация – окончание синтеза – происходит по команде стоп-кодона: УАА / УАГ / УГА. На этом этапе полипептидная цепь покидает рибосому, и трансляция завершается.
Биосинтез белка (вариант 2)
Осуществляется на рибосомах. Для передачи информации с ДНК к месту синтеза белка требуется и-РНК. Процесс синтеза на цепи ДНК молекулы и-РНК – транскрипция. Она происходит на небольшом участке, отвечающие определённому гену. При этом часть двойной спирали ДНК раскручивается, обнажая короткий участок одной из цепей, который будет служить матрицей для синтеза и-РНК. Затем вдоль этой цепи движется фермент РНК-полимераза, соединяя между собой нуклеотиды в растущую цепь и-РНК. В результате образуется и-РНК, последовательность нуклеотидов которой является копией последовательности нуклеотидов матрицы. Синтезированная в ядре и-РНК отделяется от ДНК и через поря оболочки ядра поступает в цитоплазму, где прикрепляется к малой субъединице рибосомы. Нить и-РНК проходит между малой и большой субъединицей, а начальная часть вновь синтезированного белка находится в структуре большой субъединицы. Комплекс из и-РНК и нескольких рибосом – полисома. На ней осуществляется синтез белка (трансляция). Она начинается со стартового кодона АУГ. Отсюда рибосома движется вдоль молекулы и-РНК, что сопровождается ростом полипептидной цепи. Число АК в таком белке равно числу триплетов и-РНК. Выстраивание АК осуществляется на рмбосомах при помощи т-РНК. Благодаря определённому расположению комплиментарных нуклеотидов, цепь т-РНК имеет форму листа клевера. В противоположной части молекулы т-РНК располагается антикодон, ответственный за прикрепление к определённому триплету и-РНК. Комплексы аминоацил-т-РНК считывают информацию, закодированную в и-РНК. Этот комплекс с помощью антикодона присоединяется к кодону и-РНК на малой субъединице. Затем к той же рибосоме прикрепляется второй комплекс. В рибосоме оказываются 2 АК и между ними возникает пептидная связь. Первая т-РНК покидает рибосому. Затем к образованному дипептиду присоединяется третья АК, четвёртая АК и до тех пор, пока рибосома не дойдёт до одного из трёх “стоп-кодонов” – УАА, УАГ, УГА. После этого синтез белка прекращается. Синтез идёт непрерывно с большой скоростью.
35. Сравните основные стадии процессов митоза и мейоза. Какова биологическая роль отдельных стадий и процессов в целом?
Митоз (непрямое деление) – основной способ деления эукариотических клеток – деление ядра, которое приводит к образованию двух дочерних ядер, в каждом из которых имеется такой же набор хромосом, что и в родительском ядре. Митоз – непрерывный процесс; делится на 4 стадии: профаза, метафаза, анафаза, телофаза.
В профазе происходит укорочение и утолщение хромосом за счёт их спирализации. В это время хромосомы состоят из двух хроматид, связанных между собой в области центромеры. С утолщением хромосом исчезает ядрышко и распадается ядерная оболочка. Хромосомы свободно лежат в цитоплазме. Центриоли расходятся к полюсам клетки. В конце профазы образуется веретено деления.
В метафазе завершается образование веретена деления. Хромосомы образуют экваториальную пластинку. В метафазе строение хромосом двойное, соединённое в области центромеры.
В анафазе центромеры делятся и из каждой удвоенной хромосомы образуются 2 отдельные идентичные дочерние хромосомы. Разделившись, хромосомы с помощью микротрубочек веретена деления движутся к полюсам клетки. В клетке находятся 2 диплоидных набора хромосом.
В телофазе начинается раскручивание хромосом. Вокруг них формируется ядерная оболочка, ядрышки. Разрушается веретено деления. Происходит разделение цитоплазмы с образованием двух клеток.
Биологическое значение митоза: одинаковое распределение между дочерними клетками молекул ДНК, поэтому обеспечивается образование генетически равноценных клеток. Это обеспечивает эмбриональное развитие и рост организма, восстановление тканей после повреждений. Митоз является основой бесполого размножения организмов.
Мейоз – особый способ деления клеток, в результате которого происходит уменьшение числа хромосом вдвое и переход клеток из диплоидного состояния в гаплоидное. Мейоз включает 2 деления. Перед началом мейоза хромосомы удваиваются.
I деление (редукционное) приводит к образованию гаплоидных клеток из диплоидных. Оно начинается с профазы I, в которой осуществляется спирализация хромосом. Также происходит сближение парных хромосом одинаковыми участками – конъюгация. В результате образуются хромосомные пары – биваленты. При спирализации хроматиды гомологичных хромосом переплетаются между собой; хромосомы обмениваются участками – кроссинговер. После этого хромосомы расходятся. В метафазе I завершается формирование веретена деления. Биваленты устанавливаются в плоскости экватора. В анафазе I гомологичные хромосомы разделяются и расходятся к полюсам. К каждому полюсу отходит гаплоидный набор хромосом, состоящий из двух хроматид. В телофазе I у полюсов веретена собирается гаплоидный набор хромосом, в котором каждый вид хромосом представлен одной хромосомой, состоящей из двух хроматид. Восстанавливается ядерная оболочка и материнская клетка делится на 2 дочерние.
II деление мейоза следует сразу за первым и сходно с митозом. Профаза II непродолжительна. В метафазе II снова образуется веретено деления, хромосомы выстраиваются на экваторе. В анафазе II осуществляется разделение центромер и каждая хроматида становится самостоятельной хромосомой. Дочерние хромосомы направляются к полюсам клетки. В телофазе II происходит деление клеток: из двух гаплоидных клеток образуются 4 гаплоидные дочерние клетки.
Значение мейоза: обеспечение постоянства кареотипа в ряду поколений; обеспечение разнообразия генетического состава гамет. Это обеспечивает появление разнообразного потомства при половом размножении.
41. Дайте понятие биогеохимического цикла. Приведите основные закономерности.
Основных круговоротов веществ в природе два: большой (геологический) и малый (биогеохимический).
Большой круговорот веществ в природе (геологический) обусловлен взаимодействием солнечной энергии с глут бинной энергией Земли и осуществляет перераспределение вещества между биосферой и более глубокими горизонтами Земли.
Осадочные горные породы, образованные за счет выветривания магматических пород, в подвижных зонах земной коры вновь погружаются в зону высоких температур и давлений. Там они переплавляются и образуют магму — источник новых магматических пород. После поднятия этих пород на земную поверхность и действия процессов выветривания вновь происходит трансформация их в новые осадочные породы (рис. 1). Символом круговорота веществ является спираль, а не круг. Это означает, что новый цикл круго-
Рис. 1. Большой круговорот веществ ворота не повторяет в точности старый, а вносит что-то новое, что со временем приводит к весьма значительным изменениям.
Большой круговорот — это и круговорот воды между сушей и океаном через атмосферу. Влага, испарившаяся с поверхности Мирового океана (на что затрачивается почти половина поступающей к поверхности Земли солнечной энергии), переносится на сушу, где выпадает в виде осадков, которые вновь возвращаются в океан в виде поверхностного и подземного стока. Круговорот воды происходит и по более простой схеме: испарение влаги с поверхности океана — конденсация водяного пара — выпадение осадков на эту же водную поверхность океана.
Подсчитано, что в круговороте воды на Земле ежегодно участвует более 500 тыс. км3воды.
Круговорот воды в целом играет основную роль в формировании природных условий на нашей планете. С учетом транс-пирации воды растениями и поглощения ее в биогеохимическом цикле, весь запас воды на Земле распадается и восстанавливается за 2 млн лет (см. рис. 6.10).
Малый круговорот веществ в биосфере (биогеохимический),в отличие от большого, совершается лишь в пределах биосферы. Сущность его в образовании живого вещества из неорганических соединений в процессе фотосинтеза и в превращении органического вещества при разложении вновь в неорганические соединения.
Этот круговорот для жизни биосферы — главный, и он сам является порождением жизни. Изменяясь, рождаясь и умирая, живое вещество поддерживает жизнь на нашей планете, обеспечивая биогеохнмический круговорот веществ.
Главным источником энергии круговорота является солнечная радиация, которая порождает фотосинтез. Эта энергия довольно неравномерно распределяется по поверхности земного шара. Например, на экваторе количество тепла, приходящееся на единицу площади, в три раза больше, чем на архипелаге Шпицберген (80° с.ш). Кроме того, она теряется путем отражения, поглощается почвой, расходуется на транспирацию воды и т. д. (рис. 6,8) а, как мы уже отмечали, на фотосинтез тратится не более 5% от всей энергии, но чаще всего 2—3%.
В ряде экосистем перенос вещества и энергии осуществляется преимущественно посредством трофических цепей.
Такой круговорот обычно называют биологическим . Он предполагает замкнутый цикл веществ, многократно используемый трофической цепью. Безусловно, он может иметь место в водных экосистемах, особенно в планк-
. 6.8. Поступление и распределение солнечной энергии в пределах биосферы Земли
тоне с его интенсивным метаболизмом, но не в наземных экосистемах, за исключением дождевых тропических лесов, где может быть обеспечена передача питательных вешеств «от растения к растению», корни которых на поверхности почвы.
Однако в масштабах всей биосферы такой круговорот невозможен. Здесь действует биогеохимический круговорот, представляющий собой обмен макро- и микроэлементов и простых неорганических веществ (СО„ Н20) с веществом атмосферы, гидросферы и литосферы. Круговорот отдельных веществ В. И. Вернадский назвал бив геохимическими циклами. Суть цикла в следующем: химические элементы, поглощенные организмом, впоследствии его покидают, уходя в абиотическую среду, затем, через какое-то время, снова попадают в живой организм, и т. д. Такие элементы называют биофилъными. Этими циклами и круговоротом в целом обеспечиваются важнейшие функции живого вещества в биосфере. В. И. Вернадский выделяет пять таких функций:
— первая функция — газовая — основные газы атмосферы Земли, азот и кислород, биогенного происхождения, как и все подземные газы — продукт разложения отмершей органики;
— вторая функция — концентрационная — организмы накапливают в своих телах многие химические элементы, среди которых на первом месте стоит углерод, среди металлов — первый кальций, концентраторами кремния являются диатомовые водоросли, йода — водоросли (ламинария), фосфора — скелеты позвоночных животных;
— третья функция — окислительно-восстановительная — организмы, обитающие в водоемах, регулируют кислородный режим и создают условия для растворения или же осаждения ряда металлов (V, Mb, Fe) и неметаллов (S) с переменной валентностью;
— четвертая функция — биохимическая — размножение, рост
перемещение в пространстве жипого вещества;
—пятая функция — биомохилт ческая деятельность челоае» ко —- охватывает все разрастающееся количество веществ земной коры, в том числи таких концентраторов углерода, кнк уголь, нефть, газ и другие, для хозяйственных и бытовых нужд человеке.
В бногеохкмических круговоротах следует' различать дне чисти, как бы два среза: 1)резервный фонд — это огромная масса движущихся веществ, не связанных с организмами; 2) обменный фонд — значительно меньший, но весьма активный, обусловленный прямым обменом биогенным веществом между организмами и их непосредственным окружением. Если же рассматривать биосферу в целом, то в ней можно выделить: 1) круговорот газообразных веществ с резервным фондом в атмосфере и гидросфере (океан) и 2) осадочный цикл с резервным фондом в земной коре (в геологическом круговороте).В связи с этим, следует отметить, лишь одни-вдннствем-пый на Земле процесс, который не тратит, а, наоборот, связывает солнечную энергию и даже накапливает ее — это создание органического вещества в результате фотосинтеза. В связывании и запасании солнечной энергии и заключается основная планетарная функция живого вещества на Земле.
3. Дайте определения и сущность понятий: «начала термодинамики», «качество энергии». В чем заключается понятие энтропии? Что такое «стрела времени» в термодинамических процессах?
Все химические процессы и превращения подчиняются законам термодинамики.
Первое начало термодинамики в формулировке Гельмгольца-Майера: общее количество энергии в изолированной системе остается постоянным. Т.Е. для любого химического процесса общая энергия системы и ее окружения всегда остается постоянной. Энергия не разрушается и не создается, а только переходит из одного вида в другой. Виды энергии: атомная, электрическая, химическая, механическая, тепловая. Согласно 1 закону термодинамики, каждый процесс в природе мог бы протекать так же легко в обратном направлении, как и в прямом. В действительности, природные процессы протекают самопроизвольно только в одном направлении, они необратимы, т.е. их нельзя заставить идти в обратную сторону, не изменив состояние окружающей среды.
В качестве критерия необратимости оказалось пригодным понятие энтропии (S), введенное Клаузиусом в 1859 г. Энтропия – мера неупорядоченности системы. Энтропия – характеристика структурного состояния системы.
Второе начало термодинамики: система и ее окружение, будучи предоставлены самой себе, приближаются обычно к состоянию максимальной неупорядоченности. Т.е., изменение энтропии для изолированной системы всегда положительно: ΔS >0. это значит, что высокоупорядоченные системы легко разрушаются, если на поддержание их упорядоченности не затрачивается энергия.
Таким образом, второе начало термодинамики указывает направление происходящих в изолированных системах процессов: возможны лишь такие процессы, которые приводят к возрастанию хаоса и беспорядка в этих системах, к снижению уровня их организованности. Именно поэтому говорят, что второе начало термодинамики определяет термодинамическую стрелу времени, т.е.однонаправленность развития событий.Для изолированных систем нормальному ходу времени соответствуют процессы, приводящие к увеличению хаоса и беспорядка в этих системах.Все биологические системы подчиняются этим двум законам термодинамики и управляются ими.
Качество энергии характеризует ее работоспособность. Чем дальше состояние системы от термодинамического равновесия, тем выше качество. И наоборот, чем ближе система к равновесию, тем качество ниже и тем меньше мы можем получить от системы энергии. Виды энергии: атомная, электромагнитная, химическая, механическая, тепловая. Этот ряд – ряд последовательной деградации энергии, ее перехода во все более рассеянное, вырожденное состояние.
33. Что такое закономерность образования потока вещества? Опишите критерии: замкнутость, степень замкнутости.
При обсуждении закона (принципа) «энергетической проводимости» уже упоминался второй экологический закон, по Ю. Н. Куражсковскому, «закон сохранения жизни»: жизнь может существовать только в процессе движения через живое тело потока вещества, энергии и информации. Прекращение движения в этом потоке прекращает жизнь». Скорее, это общий диалектический постулат или, уже, действительно закон жизни, а не только экологии, хотя он справедлив и для любых экологических образований и вообще многих природных систем» даже непосредственно не связанных с живым.
Более специфичен для экологических и биоценотических систем закон однонаправленности потока энергии: энергия, получаемая сообществом (экосистемой) и усваиваемая продуцентами, рассеивается или вместе с их биомассой передается консумеятам первого, второго и т. д. порядков, а затем редуцентам с падением потока на каждом трофическом уровне в результате процессов, сопровождающих дыхание. Поскольку в обратный поток (от редуцентов к продуцентам) поступает ничтожное количество изначально вовлеченной энергии (не более 0,25, максимум 0,35%), говорить о «круговороте энергии» нельзя. Существует лишь круговорот веществ, поддерживаемый потоком энергии.
Из закона однонаправленности потока энергии, конечно, с учетом ее рассеивания, а потому «бокового» использования в соседних экосистемах, и того обстоятельства, что энергия приходит в экосистему не только непосредственно от Солнца, но и опосредованно через вещественные абиотические экологические компоненты (атмосферу, воду, субстраты), вытекают многочисленные следствия и прежде всего правило (принцип) экологического дублирования. Однонаправленность потока энергии формирует в экосистемах относительно замкнутый круговорот веществ. Закономерности распространения информации также связаны с потоком энергии и перемещением вещества, но они пока еще мало изучены. Видимо, здесь большую роль играют горизонтальные связи между экосистемами, хотя информационные сети явно гуще и лучше налажены в пределах образований каждого уровня их иерархии. Пока еще поле обобщений в области экоинформатики почти не возделано. Тут непочатый край работы для специалистов.
Эмпирически более ясен вопрос о взаимоотношении энергии, вещества и информации внутри экосистем и отношении этого взаимодействия к их динамическим качествам. В начале 70-х гг. был сформулирован закон внутреннего динамического равновесия, а затем четыре основных следствия из него. Формулировка закона: вещество, энергия, информация и динамические качества отдельных природных систем (в том числе экосистем) и их иерархии взаимосвязаны настолько, что любое изменение одного из этих показателей вызывает сопутствующие функционально-структурные количественные и качественные перемены, сохраняющие общую сумму вещественно-энергетических, информационных и динамических качеств систем, где эти изменения происходят, или в их иерархии.
Действие закона внутреннего динамического равновесия совершенно четко связано с законом однонаправленности потока энергии. Именно ограниченность этого потока и специфические свойства формируют всю массу связей в экосистеме в их разнообразии. Поэтому и соблюдается экологический аналог законов сохранения массы и энергии. Этот закон — одна из основных путеводных нитей в управлении природопользованием, впрочем, нитей, долго игнорировавшихся.
Важные для практики следствия из закона внутреннего динамического равновесия:
Любое изменение среды (вещества, энергии, информации, динамических качеств экосистем) неизбежно приводит к развитию природных цепных реакций, идущих в сторону нейтрализации произведенного изменения или формирования новых природных систем, образование которых при значительных изменениях среды может принять необратимый характер.
Все закономерности саморегуляции ценозов обобщаются в виде принципа стабильности: любая относительно замкнутая биосистема с проходящим через нее потоком энергии в ходе саморегуляции развивается в сторону устойчивого состояния. Этот принцип характерен не только для ценозов -нижнего уровня иерархии, но и для биосферы в целом. Тут важно то, что ценоз стремится к нормальной «энергетической проводимости с помощью механизмов, обобщенно сформулированных в правилах (принципах) экологического дублирования, эквивалентности, подвижного равновесия, продукционной оптимизации и, вероятно, других, еще не открытых исследователями.
Если принцип стабильности справедлив, то парадигма континуума получает еще одно ограничение, а парадигма организма—лишний аргумент своей справедливости. Правда, система может складываться и из ненадежных элементов — правило конструктивной эмерджентности.
infopedia.su
Лизосомы | steelbros.ru - сообщество стальных братюней
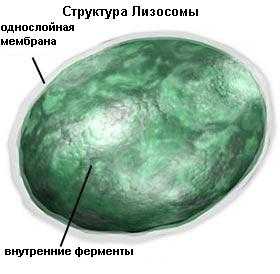
Лизосомы.Лизосомы - мелкие сферические органоиды, стенки которых образованы одинарной мембраной; содержат расщепляющие ферменты. Размер их в среднем 0,2 — 0,4 мкм.
Строение и функции схожи с вакуолью. Образовываются из пузырьков отделяющихся от аппарата Гольджи, а так же из пузырьков при захвате внешнего материала клеткой. В некоторых клетках человека количество лизосом может достигать тысячи.
Виды лизосом и их функции.
1) Ранняя — в нее поступают захваченные клеткой пузырьки. Рецепторы, отдавшие (из-за пониженного рН) свой груз, возвращаются на наружную мембрану.
pH – мера активности йонов водорода. (h3+)
2) Поздняя — в нее из ранней поступают пузырьки с материалом, поглощенном при захвате клеткой, и пузырьки из аппарата Гольджи с расщепляющими веществами.
3) Лизосома — в нее из поздней поступают пузырьки со смесью гидролаз и перевариваемого материала.
4) Фагосома — в нее попадают более крупные частицы (бактерии и т. п.), поглощенные путем фагоцитоза. Фагосомы обычно сливаются с лизосомой.
Фагоцито́з — процесс, при котором специально предназначенные для этого клетки крови и тканей организма захватывают и переваривают возбудителей инфекционных заболеваний и отмершие клетки.
5) Аутофагосома — окруженный двумя мембранами участок цитоплазмы, обычно включающий какие-либо органоиды и образующийся при макроаутофагии. Сливается с лизосомой.
Аутофагия - является одним из основных механизмов для ликвидации поврежденных органелл, долгоживущих и аномальных белков и излишних объёмов цитоплазмы.
6) Мультивезикулярные тельца — обычно окружены одинарной мембраной, содержат внутри более мелкие окруженные одинарной мембраной пузырьки. Образуются в результате процесса, напоминающего микроаутофагию , но содержат материал, полученный извне. В мелких пузырьках обычно остаются и затем подвергаются деградации рецепторы наружной мембраны. По стадии формирования соответствуют ранней лизосоме.
При микроаутофагии образуются впячивания мембраны захваченной клетой среды или лизосомы, которые затем отделяются в виде внутренних пузырьков, только в них попадают вещества, синтезированные в самой клетке. Таким путем клетка может переваривать белки при нехватке энергии или строительного материала (например, при голодании). Но процессы микроаутофагии происходят и при нормальных условиях и в целом неизбирательны. Иногда в ходе микроаутофагии перевариваются и органоиды.
7) Остаточные тельца (телолизосомы) — пузырьки, содержащие непереваренный материал. В нормальных клетках сливаются с наружной мембраной и путем слияния с клеточной мембраной покидают клетку. При старении или патологии накапливаются.
Мембрана лизосом очень прочная и препятствует проникновению собственных ферментов в цитоплазму клетки, но если лизосома повреждается от каких-либо внешних воздействий, то разрушается вся клетка или часть ее.
Надо отметить еще одну важную функцию:автолиз — самопереваривание клетки, приводящее к ее гибели (иногда этот процесс не является патологическим, а сопровождает развитие организма или дифференцировку некоторых специализированных клеток).
Функции лизосом
3д представление
steelbros.ru