
Циркадные ритмы растений и их значение. Циркадные ритмы растений
Чем обусловлено проявление внутренних ритмов роста и развития растений
Как и все живое, растения имеют свои «биологические часы», управляющие их жизнью. На точность этих часов влияет много разных факторов Главный фактор, воздействующий на жизнь растений, это космическое излучение.
Ученые ставили опыты с растениями, выращивая их в очень глубоких пещерах. Растения были помещены в специальные камеры, куда не могли попасть лучи из космоса. В результате этих опытов ученые сделали следующий вывод: на большой глубине, куда нет доступа солнечному излучению, у растений теряется ориентировка во времени.

Суточные ритмы у растений
Наследственность растений за многие поколения закрепила циклические ритмы. Если нарушить освещение, влажность, температуру, то требуется какой-то период, чтобы ритмы эти изменились. «Циркадные» ритмы – так назвали данные процессы. На латинском языке это означает – ритмы, близкие к суточному периоду. «Цирка» — около, «диес» — день. Жизнь растений полностью у власти этих суточных ритмов. Наблюдая за тем, как раскрываются венчики цветов, можно точно определить время.
Первым встречает рассвет козлобородник, раскрывает свои желтенькие звездочки. Почти одновременно с ним расцветают голубенькие цветки цикория. Ранней пташкой являются и цветки шиповника, лилейника.
Только окрасят небо первые лучи солнца, тут же раскрывается множество цветов, показывая свою яркую окраску и свежесть. К шести утра просыпаются алые маки, желтые одуванчики, вьюнки, лен, цветки картофеля.
Стоит лучикам солнца залить поля, луга, реки, как на водяной глади распускается белоснежная лилия. Клумбы алеют и золотятся ноготками, чернобривцами, распускаются граммофоны ипомеи.
К середине дня всеми цветами радуги пестрят луга, степи, сады от огромного количества распустившихся цветов. Но после полудня постепенно краски тускнеют, и растения начинают закрываться. Цветы, которые раньше распустились, первыми и готовятся ко сну. Это — одуванчики, лютики, цикорий, козлобородник, ястребинка. К 17-ти часам складываются цветки у лилии, а шиповник цветет, пока солнце не зайдет.
Растения не только распознают смену дня и ночи, они приспособились к нужной для них длительности дня. Есть растения короткодневные и те, которые расцветают во время летнего солнцестояния.
Самым пунктуальным растением в мире является вечнозеленый кустарник, произрастающий в Малайзии. Его название — симпоха. Этот кустарник цветет ежедневно. Цветки распускаются друг за другом, постепенно набухая, и полностью раскрываясь за час перед восходом солнца. К шестнадцати часам цветок опадает. На 36-ой день после опадения лепестков, утром, ровно в три часа – раскрывается плод симпохи. И так всю жизнь. А живет кустарник иногда и до ста лет.

Почему растения способны выбирать именно то или иное время для цветения?
Растения используют изменение в спектральном составе солнечного света, чтобы определить длительность дня. За это отвечает уникальный пигмент фитохром. Эти пигменты содержатся в растениях в ничтожно маленьких количествах. Своим строением они сходны с желчными пигментами у животных и водорослей. Пигменты голубого цвета, а в листьях хлорофилл и каротиноиды их маскируют. Фитохром существует в двух формах. Они способны превращаться из одной формы пигмента в другую, что зависит от времени суток. Клетки растений способны считывать данную информацию и соблюдать четко биологические ритмы.
Периодичность цветения вырабатывалась у растений постепенно, она приспособилась и синхронно существует с ритмом жизни насекомых, опыляющих их. Цветки отдают нектар, пыльцу, а насекомые не совершают лишних полетов, тем самым экономят свою энергию. Такие растения имеют яркие, броские венчики. Они открываются, когда насекомые начинают летать, а с наступлением вечера закрываются. Тем самым защищают пыльцу, тычинки от увлажнения росой, переохлаждения или осадков.
Многие растения цветут в вечернее время или ночью. Они белого цвета или имеют сильный аромат, что является ориентиром для насекомых, их опыляющих. После захода солнца наполняет ароматом воздух ночная фиалка, благоухают цветки табака, ночной красавицы (мирабилис).
Зная, что влияет на ритмы жизни у растений, научились искусственно создавать условия для выгонки весенних цветов к определенным праздникам. Южные растения теперь могут расти в северных районах, а северные растения – в южных. Можно успешно бороться с сорняками. Для этого надо изучить ритм развития сорняков и определить ту фазу, когда они будут уязвимы для гербицидов, а растения, требующие защиты, меньше всего будут чувствительны к гербицидам.
sadiogorod24.ru
Циркадные ритмы растений и их значение
Для растений характерна суточная периодичность, проявляющаяся в движении листьев, раскрывании цветков, активности выделения пасоки и нектара, изменении скорости и частоты митозов и т. д.
Такая эндогенная, суточная (циркадная) ритмика свойственна растениям даже в темноте, когда они уже не подвергаются действию света («физиологические часы»). Э. Бюннинг (1961) полагает, что у растений существует временная память. Так, цветки ряда видов начинают раскрываться незадолго до рассвета, как будто «зная», что через несколько часов взойдет солнце. Это у них повторяется и в темноте. В целом «растения, подобно животным, правильно измеряют время суток, по крайней мере длительность ночи» (Э. Бюннинг, 1961). Сейчас многие особенности жизнедеятельности связывают с суточной ритмикой, особенно много фактов накоплено при изучении суточной периодичности движения листьев. Для большей части изученных процессов продолжительность периода циркадных ритмов составляет 22—28 ч.
Доказаны наследственная обусловленность (Н. Mletzko, J. Mletzko, 1977) суточной ритмики и ее приспособительное значение. Так, сорта сои, возделываемые в экваториальной зоне, отличаются меньшей амплитудой эндогенного суточного ритма от сортов, возделываемых вне указанной зоны. На примере дикорастущих злаков показана устойчивость суточной ритмики цветения, что позволяет использовать ее как таксономический признак. Суточная ритмика цветения выполняет важную биологическую роль, она ускоряет дифференциацию популяций, создавая барьеры изоляции.
Сходная суточная ритмика цветения видов (Elytrigia) нередко задерживает микроэволюционные процессы полиморфных видов (Е. repens), уменьшая морфологические различия благодаря естественной гибридизации разных видов. Опыты по скрещиванию сортов о неодинаковыми эндогенными ритмами показали их наследование по промежуточному типу в F1. В то же время наследственные эндогенные ритмы могут перестроиться в онтогенезе. Однако с возникновением типичных условий они вновь восстанавливаются, что показано в опытах по изучению влияния различной длины дня на движение листьев фасоли. Оказалось, что на непрерывном освещении, как и при 3—4-часовой длине дня, листья фасоли неактивны. Движение листьев восстанавливалось при сокращении светового и увеличении темнового интервала времени. Так, при 16-часовом дне листья стали опускаться на свету и подниматься в темноте на 5-й день. При 12-часовом и еще более коротком дне листья на свету не опускались, а в темноте поднимались со значительным опережением, достигавшим нескольких часов.
В данном опыте движение листьев определялось условиями. Наследственно закреплены не сами никтинастические движения, а способность растений приспосабливать свой суточный метаболизм к привычным для вида световым и другим условиям. На примере тока пасоки в ксилеме древесных растений обнаружено, что он меняется в зависимости от условий. Динамическую регуляцию биоритмов рассматривают как один из универсальных адаптационных механизмов, ведущих к перестройке жизнедеятельности. Например, расход воды у растений фасоли при 24-часовом освещении оказывается в 2—3 раза меньше по сравнению с растениями, находящимися на 14-часовом освещении. В условиях непрерывного освещения транспирация носила постоянный характер, в то время как в условиях 14-часового освещения она возрастала от момента включения света до середины дня с последующим падением. При этом в середине дня интенсивность транспирации была в 6—7 раз выше, чем при непрерывном освещении.
У некоторых грибов и водорослей высвобождение спор из спорангия представляет собой процесс с четкой суточной периодичностью, регулируемый эндогенно. Свечение одноклеточной водоросли Gonyaulaxpolyedra связано с эндогенными ритмами накопления люциферина и люциферазы ночью. У этой водоросли обнаружены также эндогенные ритмы деления клеток (В. М. Sweeney, 1969).
Суточный ритм физиологических функций — один из основных и наиболее значимых для растений. Он обусловлен чередованием дня и ночи, связан с изменением организации мембран (D. Th. Vanden, 1979) и отражает приспособление растений к вращению Земли и Луны вокруг оси. В ходе эволюции растений менялись процессы, которыми управляют часы и фоторецепторы самих часов, но сам принцип измерения времени на основе эндогенного ритма природа сохранила. В естественных условиях растения «измеряют» не длину дня, а длину ночи. Так, никтофильная перилла зацветает только тогда, когда находится в темноте не менее 9 ч. Эволюционное значение актиноритмизма состояло не в том, чтобы ускорить развитие и спасти растения от осенних заморозков, а в том, «поры замедлить развитие, накопить значительную массу листьев и лишь потом мобилизовать развитый фотосинтетический аппарат на воспроизведение потомства.
Суточные ритмы регулируются с участием фитохромов (Р660 и Р730). У длиннодневных растений уровень Р730 на свету низкий; а в темноте высокий. После включения света содержание Р730 некоторое время остается высоким. У короткодневных растений Р730 регулирует ход биологических часов, а биологические часы — динамику фитохрома.
Для уточнения значения циркадных ритмов представляет интерес выяснение их связи с фотопериодической реакцией. Есть целый ряд доказательств нарушения циркадных ритмов, когда ночной период искусственно прерывался вспышкой света (в течение минуты). При этом у длиннодневных растений даже при коротком дне наступает; зацветание. Прерывание же световой фазы «темновым интервалом» не дает никакого эффекта ввиду того, что решающее значение имеет длина ночи. Отсчет фотопериодического времени идет с участием физиологических часов, где начало освещения или затемнения действует как датчик времени для синхронизации фаз. Кульминационная точка фотофильной фазы у коротко- и длиннодневных растений разная.
Циркадные ритмы у растений, несомненно, имеют приспособительное значение: амплитуда их колебания расширяет норму реакции организма в онтогенезе. Они играют важную роль и в сезонных циклах.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Циркадный ритм | Info-Farm.RU
Циркадные (циркадианного) ритмы (от лат. Circa — около, кругом и лат. Dies — день) — циклические колебания интенсивности различных биологических процессов, связанные со сменой дня и ночи. Несмотря на связь с внешними стимулами, циркадные ритмы имеют эндогенное происхождение, представляя, таким образом, «внутренние часы» организма. Циркадные ритмы присутствуют в таких организмов как цианобактерии, водоросли, грибы, растения, животные. Период циркадных ритмов обычно близок к 24 часов.
Три основные особенности циркадных ритмов
- Ритм сохраняется при постоянных условиях и имеет период близок к 24 часов.
- Ритм может быть синхронизирован под действием внешнего освещения.
- Ритм не зависит от температуры, пока она изменяется в диапазоне, пригодном для жизнедеятельности.
История открытия
Впервые об изменении положения листьев в течение дня в тамаринда (Tamarindus indicus) вспоминает андростен, описывая походы Александра Македонского.
В новое время в 1729 году французский астроном Жан-Жак ДьОрту где Марен сообщил о ежедневных движения листьев у мимозы стыдливой (Mimosa pudica). Эти движения повторялись с определенной периодичностью даже если растения помещались в темноту, где отсутствуют такие внешние стимулы как свет, что позволило предположить эндогенное происхождение биологических ритмов, к которым были приурочены движения листьев растения. Где Мейрон предположил, что эти ритмы могут иметь что-то общее с чередованием сна и бодрствования у человека.
Декандоль в 1832 году определил, что период, с которым растения мимозы осуществляют данные листовые движения, несколько короче чем продолжительность суток и составляет примерно 22-23 часов.
В 1880 году Чарльз Дарвин и его сын Фрэнсис предположили наследственную природу циркадных ритмов. Это предположение было подтверждено опытами, во время которых скрещивались растения разных сортов фасоли, периоды циркадных ритмов которых различались. У гибридов длина периода отличалась от длины периода у обоих родителей. Эндогенная природа циркадных ритмов была окончательно подтверждена в 1984 году во время опытов с грибами вида нейроспоры густая (Neurospora crassa), проведенными в космосе. Эти опыты показали независимость круглосуточного ритма от геофизических сигналов, связанных с вращением Земли вокруг своей оси.
Циркадные ритмы цианобактерий
Простейшие циркадные ритмы обнаружены у цианобактерий. Цианобактерии (сине-зеленые водоросли) — монофилетическая группа (имеют одного общего предка) фотоавтотрофной (питаются за счет фотосинтеза, энергии солнца) бактерий. Это одна из древнейших и самых групп в мире прокариот. Различные представители группы сильно отличаются друг от друга как морфологически, так и генетически, их можно обнаружить почти в любом, доступном для света, среде обитания. Жизненные циклы так же имеют разную продолжительность в различных представителей: от нескольких часов до нескольких тысяч лет между делениями (у некоторых видов, проживающих на олиготрофных, бедных средах).
Впервые наличие циркадные ритмов у цианобактерий было продемонстрировано при изучении процессов кислородно-чувствительной фиксации азота и фотосинтеза с выделением кислорода. В этих процессах была показана суточная ритмичность. Об этом в частности говорили данные электронной микроскопии, с помощью которой изучали количество и размер тех или иных запасающих гранул в клетках. Позже было обнаружено, что и другие процессы в клетках (например, поглощение аминокислот) происходят в рамках циркадного ритма, удовлетворяя трем основным положениям циркадных ритмов.
Кроме того, оказалось, что ритмично изменяется вся экспрессия генов в клетках цианобактерий. Были проведены опыты, в которых гены биолюминесцентных (тех, светящиеся) белков встраивались в геном цианобактерий под случайные бактериальные промоторы. Во всех полученных штаммов наблюдалась подобная картина циркадных изменений интенсивности люминесценции.
Важность синхронизации внутреннего ритма с экзогенным фактором освещенности для цианобактерий было показано в ряде опытов. Например, было показано, что бактерии с нарушенной синхронизацией циркадных ритмов медленнее растут в условиях смены дня и ночи, тем самым, проигрывая бактериям с хорошо отлаженными и настроенным внутренним часам. Кроме того, как уже говорилось, цианобактерии размножаются только в «ночной» период, который определяется их внутренним часам, в частности защищает ДНК, реплицируется, от повреждений кислородными радикалами, которые образуются под действием ультрафиолетовых лучей солнца.
Циркадные ритмы растений
Циркадные ритмы растений связанные со сменой дня и ночи и важные для адаптации растений к суточных колебаний таких параметров как температура, освещение, влажность. Растения существуют в постоянно меняющемся мире, поэтому циркадные ритмы важны для того, чтобы растение могло дать надлежащий ответ на абиотический стресс. Изменение положения листьев в течение суток — лишь один из многих ритмических процессов у растений. В течение суток колеблются такие параметры как активность ферментов, интенсивность газообмена и фотосинтетическая активность.
В способности растений распознавать чередование дня и ночи решающую роль играет фитохромна система. Примером работы такой системы является ритм цветения у растения Pharbitis nil. Цветение у этого растения зависит от длины светового дня: если день короче определенного интервала, то растение цветет, если дольше — вегетуе.
В течение суток условия освещения меняются из-за того, что солнце находится под разными углами к горизонту, и соответственно изменяется спектральный состав света, воспринимается различными фитохромом которые возбуждаются светом с различной длиной волны. Так, вечером в спектре много дальних красных лучей, которые активизируют только фитохром А, давая растению сигнал о приближении ночи. Получив этот сигнал, растение принимает соответствующие меры. Важность фитохромов для температурной адаптации было выяснено во время опытов с трансгенными осинами Populus tremula, в которых продукция фитохрома А была повышенной. Растениям постоянно «казалось», что они получают свет высокой интенсивности, и таким образом они не могли адаптироваться к суточных колебаний температуры и страдали от ночных заморозков.
При исследовании суточных ритмов в арабидопсиса было также показано фотопериодичность работы трех генов CO, FKF1 и G1. Ген constans участвует в определении времени цветения. Синтез продукта гена CO запускается комплексом из белков FKF1 и G1. В этом комплексе продукт гена FKF1 играет роль фоторецептора. Синтез белка CO запускается через 4:00 после начала освещения и останавливается в темноте. Синтезированный белок за ночь разрушается и таким образом необходима для цветения растения концентрация белка достигается только в условиях длинного летнего дня.
Циркадные ритмы у животных
Практически все животные приспосабливают свои физиологические и поведенческие процессы к суточным колебаниям абиотических параметров. Примером циркадного ритма у животных есть цикл сон-бодрствование. У человека и у других животных существуют внутренние часы, которые идут даже при отсутствии внешних стимулов, которые могут дать информацию о времени суток. Исследование молекулярно-биологической природы этих часов началось около 30 лет назад. Конопка и Бензер, работавших в Калифорнийском технологическом институте выявили три мутантных линии дрозофил, циркадные ритмы которых отличались от циркадных ритмов мушек дикого типа. Дальнейший анализ показал, что у мутантов мутации затрагивали аллели одного локуса, который был назван исследователями per (от period). При отсутствии нормальных сигналов окружающей среды период круглосуточной активности у мушек дикого типа составлял 24 часа, в мутантов per-s — 19 часов, в мутантов per-1 — 29 часов, в мутантов per-0 вообще не наблюдалось никакого ритма. Впоследствии было обнаружено, что продукты генов per есть во многих клетках дрозофил, которые участвуют в становлении циркадного ритма насекомые. Более того, у мушек дикого типа наблюдаются циркадные колебания в количестве per мРНК и белка Per в то время как у мушек per0, у которых нет циркадного ритма такой цикличности, экспрессии не наблюдается.
Циркадные ритмы и цикл сон — бодрствование у человека
Периоды сна и бодрствования у человека меняются с циркадных периодичностью. При исследовании связи периодичности сна и бодрствования с внешними стимулами изучалось изменение продолжительности периода данных колебаний у человека. При отсутствии таких стимулов как свет, позволяет человеку судить о времени суток, подопытные все равно ложились спать и просыпались в обычное время; таким образом, период ритма сон — бодрствование не менялся и в течение некоторого времени оставался равным 24 часам, правда через некоторое время он увеличился до 36 часов. Когда испытуемые вернулись в нормальные условия, то 24-часовой цикл был восстановлен. Таким образом, у человека и у многих других животных есть внутренние часы, которые идут даже в отсутствии внешних сигналов.
Одним из самых распространенных внешних сигналов свет. У человека рецепторы, находящиеся в сетчатке, реагируют на свет и посылают сигнал в супрахиазмальне ядро. Дальнейшее распространение сигнала приводит к выработке гормонов, регулирующих циркадные активность организма. Однако при этом такие органы как сердце, печень, почки имеют свои «внутренние часы» и могут выбиваться из ритма, устанавливается супрахиазматическим ядром. Сигнал, поступающий в шишковидную железу, вызывает синтез и выделение в кровоток нейрогормона, что вызывает сон — мелатонина (N-ацетил-5-метокситриптамин). У пожилых людей выделяется меньше мелатонина, что, вероятно, объясняет, почему старики чаще страдают бессонницей. Большая часть исследователей считает, что супрахиазматическом ядро отвечает за циркадные ритмы и за колебания параметров, связанных с циклом сон — бодрствование, таких как температура тела, давление и продукция мочи.
Заболевания, связанные с нарушением циркадного ритма
У взрослых во время сна уменьшается продукция мочи в связи с увеличением содержания АДГ в крови. У некоторых детей и взрослых, в которых цикличность колебаний содержания вазопрессина возбуждено, уменьшение продукции мочи в ночное время не происходит, что приводит к неконтролируемому мочеиспускания. Такое заболевание как смертельное наследственное бессонницы заканчивается летальным исходом и связано с врожденными дефектами нейронов супрахиазматического ядра. Интересно, что подобные симптомы возникают при болезни Крейцфельда-Якоба, когда поражаются клетки того же супрахиазматического ядра.
Международная классификация нарушений
Диссомнии
А. Внутренние нарушения сна:
- Психофизиологическая инсомния
- Идиопатическая инсомния
- Нарколепсия
- Синдром сонных апноэ (ночных апноэ)
- Синдром подергивания конечностей (ночная миоклония)
- Синдром беспокойных ног
Б. Внешние нарушения сна:
- Неадекватная гигиена сна
- Высотная инсомния
- Нарушение сна, вызванные употреблением алкоголя и медикаментов
В. Расстройства циркадного ритма сна
- Нарушение регулярности цикла сон — бодрствование
- Нарушение сна при сменной работе
- Синдром позднего засыпания
- Синдром раннего засыпания
Парасомнии
А. Нарушение пробуждения:
- Пробуждение с спутанной сознанием
- Снохождения
- Ночные страхи
Б. Нарушение перехода сон — бодрствование:
- Говорения во сне
- Ночные судороги ног
В. парасомнии, обычно связанные с фазой быстрого сна:
- Кошмарные сновидения
- Сонный паралич
- Нарушение эрекции полового члена, связанные со сном
- Болезненные эрекции, связанные со сном
Г. Другие парасомнии:
- Сонный бруксизм (скрежет зубами)
- Сонный энурез
Изображения по теме
info-farm.ru
9. Адаптивные биологические ритмы. Циркадный и цирканный ритмы. Фотопериодизм.
Одно из фундаментальных свойств живой природы – цикличность большинства происходящих в ней процессов. Вся жизнь на Земле, от клетки до биосферы, подчинена определенным ритмам. Природные ритмы для любого организма можно разделить на внутренние (связанные с его собственной жизнедеятельностью) и внешние (циклические изменения в окружающей среде).
Внутренние циклы. Внутренние циклы– это прежде всего физиологические ритмы организма. Ни один физиологический процесс не осуществляется непрерывно. Ритмичность обнаружена в процессах синтеза ДНК и РНК в клетках, в сборке белков, в работе ферментов, деятельности митохондрий. Определенному ритму подчиняются деление клеток, сокращение мышц, работа желез внутренней секреции, биение сердца, дыхание, возбудимость нервной системы, т. е. работа всех клеток, органов и тканей организма. При этом каждая система имеет свой собственный период. Изменить этот период действием факторов внешней среды можно лишь в узких пределах, а для некоторых процессов совсем нельзя. Такую ритмику называютэндогенной.
Все внутренние ритмы организма соподчинены, интегрированы в целостную систему и в конечном счете выступают как общая периодичность поведения организма. Ритмически осуществляя свои физиологические функции, организм как бы отсчитывает время. И для внешних, и для внутренних ритмов наступление очередной фазы зависит прежде всего от времени. Поэтому время выступает как один из важнейших экологических факторов, на который должны реагировать живые организмы, приспосабливаясь к внешним циклическим изменениям природы.
Внешние ритмы. Основныевнешние ритмы имеют геофизическую природу, так как связаны с вращением Земли относительно Солнца и Луны относительно Земли. Под влиянием этого вращения множество экологических факторов на нашей планете, в особенности световой режим, температура, давление и влажность воздуха, атмосферное электромагнитное поле, океанические приливы и отливы и др., закономерно изменяются. Кроме того, на живую природу воздействуют и такие космические ритмы, как периодические изменения солнечной активности (рис. 58). Для Солнца характерен 11-летний и целый ряд других циклов. Изменения солнечной радиации существенно влияют на климат нашей планеты. Кроме циклического воздействия абиотических факторов, внешними ритмами для любого организма являются также закономерные изменения активности и поведения других живых существ.
Целый ряд изменений в жизнедеятельности организмов совпадает по периоду с внешними, геофизическими циклами. Это так называемые адаптивные биологические ритмы–суточные (циркадные), приливно-отливные, равные лунному месяцу, годичные (цырканные). Благодаря им самые важные биологические функции организма, такие, как питание, рост, размножение, совпадают с наиболее благоприятным для этого временем суток или года.
Адаптивные биологические ритмы возникли как приспособление физиологии живых существ к регулярным экологическим изменениям во внешней среде. Этим они отличаются от чисто физиологических ритмов, которые поддерживают непрерывную жизнедеятельность организмов – дыхания, кровообращения, деления клеток и т. д.
5.1. Суточный ритм
Суточный ритм обнаружен у разнообразных организмов, от одноклеточных до человека. У человека отмечено свыше 100 физиологических функций, затронутых суточной периодичностью: сон и бодрствование, изменение температуры тела, ритма сердечных сокращений, глубины и частоты дыхания, объема и химического состава мочи, потоотделения, мышечной и умственной работоспособности и т. п. У амеб в течение суток изменяются темпы деления. У некоторых растений к определенному времени приурочены открывание и закрывание цветков, поднятие и опускание листьев, максимальная интенсивность дыхания, скорость роста колеоптиля и т. д. (рис. 59).
По смене периодов сна и бодрствования животных делят на дневных иночных. Ярко выражена дневная активность, например, у домашних кур, большинства воробьиных птиц, сусликов, муравьев, стрекоз. Типично ночные животные – ежи, летучие мыши, совы, кабаны, большинство кошачьих, травяные лягушки, тараканы и многие другие. Животные обычно активны в наиболее благоприятное для них время суток. Конкретные типы активности достаточно разнообразны (рис. 60). Некоторые виды имеют приблизительно одинаковую активность как днем, так и ночью, с чередованием коротких периодов бодрствования и покоя. Такой ритм называютполифазным (многие землеройки, ряд хищных и др.).
У ряда животных суточные изменения затрагивают преимущественно двигательную активность и не сопровождаются существенными отклонениями физиологических функций (например, у грызунов). Наиболее яркие примеры физиологических сдвигов в течение суток дают летучие мыши. Летом в период дневного покоя многие из них ведут себя как пойкилотермные животные. Температура их тела в это время почти равна температуре среды; пульс, дыхание, возбудимость органов чувств резко понижены. Чтобы взлететь, потревоженная мышь долго разогревается за счет химической теплопродукции. Вечером и ночью – это типичные гомойотермные млекопитающие с высокой температурой тела, активными и точными движениями, быстрой реакцией на добычу и врагов.
У одних видов периоды активности приурочены к определенному времени суток (рис. 61), у других могут сдвигаться в зависимости от обстановки (рис. 62). Так, открывание цветков шафрана зависит от температуры, соцветий одуванчика – от освещенности: в пасмурный день корзинки не раскрываются. Активность пустынных мокриц или жуков-чернотелок сдвигается на разное время суток в зависимости от температуры и влажности на поверхности почвы. Они выходят из норок либо рано утром и вечером (двухфазный цикл), либо только ночью(однофазный), либо в течение всего дня.
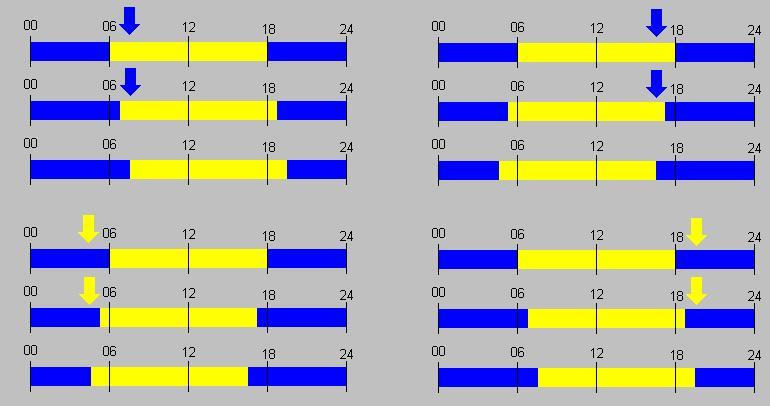
Отличить эндогенные суточные ритмы от экзогенных, т. е. навязываемых внешней средой, можно в эксперименте. У многих видов при полном постоянстве внешних условий (температуры, освещенности, влажности и т. п.) продолжают длительное время сохраняться циклы, близкие по периоду к суточному. У дрозофил, например, такой эндогенный ритм прослеживается в течение десятков поколений. Таким образом, суточная цикличность жизнедеятельности переходит во врожденные, генетические свойства вида. Такие эндогенные ритмы получили итшштициркадных (от лат. circa – около и dies – день, сутки), так как длительность их неодинакова у разных особей одного вида, слегка отличаясь от среднего, 24-часового периода. При снятии внешней ритмики дня и ночи эндогенный ритм становится свобод-нотекущим и через некоторое время перестает совпадать с суточными изменениями (рис. 63).
Летяги, для которых характерна сумеречная активность, просыпаются вечером синхронно, в строго определенный час. В эксперименте, находясь в полной темноте, они сохраняют околосуточный ритм. Однако одни особи начинают свой «день» на несколько минут раньше; другие – на несколько минут позже обычного суточного цикла. Если, например, цир-кадиый ритм короче суточного на 15 мин, то для такого зверька через три дня расхождение во времени с внешним ритмом составит 45 мин, через 10 дней – уже 2,5 ч и т. д. Поэтому все летяги через несколько суток просыпаются и начинают двигаться в совершенно разное время, хотя каждая сохраняет постоянство своего цикла. При восстановлении смены дня и ночи сои и бодрствование зверьков вновь синхронизируются. Таким образом, внешний (-уточный цикл регулирует продолжительность врожденных циркадных ритмов, согласуя их с изменением среды.
У человека циркадные ритмы изучались в различных ситуациях: в пещерах, герметических камерах, подводных плаваниях и т. п. Обнаружилось, что в отклонениях от суточного цикла у человека большую роль играют типологические особенности нервной системы. Циркадные ритмы могут быть различны даже у членов одной и той же семьи.
Известный стереотип поведения, обусловленный циркадным ритмом, облегчает существование организмов при суточных изменениях среды. Однако при расселении животных и растений и попадании их в географические условия с иной ритмикой дня и ночи слишком прочный стереотип может стать неблагоприятным. Поэтому расселительные возможности ряда видов ограничены глубоким закреплением их циркадных ритмов. Так, например, серые крысы отличаются от черных значительно большей пластичностью суточного цикла. У черных крыс он почти не поддается перестройке, и вид имеет ограниченный ареал, тогда как серые крысы распространились практически по всему миру.
У большинства видов перестройка циркадного ритма возможна. Обычно она происходит не сразу, а захватывает несколько циклов и сопровождается рядом нарушений в физиологическом состоянии организма. Например, у людей, совершающих перелеты на значительные расстояния в широтном направлении, наступает десинхронизация их физиологического ритма с местным астрономическим временем. Организм сначала продолжает функционировать по-старому, а затем начинает перестраиваться. При этом чувствуется повышенная усталость, недомогание, желание спать днем и бодрствовать ночью. Адаптивный период продолжается от нескольких дней до двух недель.
Десинхронизация ритмов представляет важную медицинскую проблему в организации ночной и сменной работы лиц ряда профессий, в космических полетах, подводных плаваниях, работах под землей и т. п.
Циркадные и суточные ритмы лежат в основе способности организма чувствовать время. Эту способность живых существ называют «биологическими часами».
Ряду высокоорганизованных животных присуща сложная врожденная способность использовать ориентацию во времени для ориентации в пространстве. Птицы при длительных перелетах также постоянно корректируют направление по отношению к Солнцу или поляризованному свету неба, учитывая время суток. «Биологические часы» живых организмов ориентируют их не только в суточном цикле, но и в более сложных геофизических циклах изменений природы.
5.3. Годичные ритмы
Годичные ритмы – одни из наиболее универсальных в живой природе. Закономерные изменения физических условий в течение года вызвали в эволюции видов множество самых разнообразных адаптаций к этой периодичности. Наиболее важные из них связаны с размножением, ростом, миграциями и переживанием неблагоприятных периодов года. У видов с коротким жизненным циклом годовой ритм закономерно проявляется в ряду поколений (например, цикломорфоз у дафний и коловраток).
Сезонные изменения представляют собой глубокие сдвиги в физиологии и поведении организмов, затрагивающие их морфологию и особенности жизненного цикла (рис. 65). Приспособительный характер этих изменений очевиден: благодаря им такой ответственный момент в жизни вида, как появление потомства, оказывается приуроченным к наиболее благоприятному времени года, а переживание критических периодов происходит в наиболее устойчивом состоянии.
Чем резче сезонные изменения внешней среды, тем сильнее выражена годовая периодичность жизнедеятельности организмов. Осенний листопад, различные диапаузы, спячка, запасание жиров, сезонные линьки, миграции и т. п. развиты преимущественно в странах умеренного и холодного климатов, тогда как у обитателей тропиков сезонная периодичность в жизненных циклах выражена менее резко.
Годичные ритмы у многих видов эндогенны. Такие ритмы называются цирканными (лат.annus– год). Особенно это относится к циклам размножения. Так, животные Южного полушария, содержащиеся в зоопарках Северного, размножаются чаще всего зимой или осенью, в сроки, соответствующие весне и лету на их родине. Австралийские страусы в заповеднике Аскания-Нова откладывали яйца зимой прямо на снег. Собака динго приносит щенков в декабре, когда в Австралии конец весны. С устойчивостью сроков размножения в годовом цикле приходится считаться при интродукции и акклиматизации видов.
Сильные оттепели зимой, заморозки летом обычно не нарушают сезонных изменений у растений и животных. Вместе с тем далеко не всегда точность протекания годового цикла имеет эндогенную природу. Например, семена ряда растений прорастают в строго определенное время года даже после экспериментально вызванного состояния полного анабиоза, который должен нарушить «отсчет времени» в организме. Следовательно, прорастание стимулируют какие-то изменения среды, связанные с геофизическими циклами.
В настоящее время интенсивно изучается реакция организмов на слабые геоэлектромагнитные поля, а также атмосферные приливы и отливы, которые закономерно меняются в циклах вращения Земли. Показано, что интенсивность ряда биологических процессов коррелирует с колебаниями этих тонких показателей состояния атмосферы в течение года, как, например, двигательная активность насекомых, скорость потребления кислорода клубнями картофеля и др.
Таким образом, наступление очередного этапа годичного цикла у живых организмов происходит частично в результате эндогенной ритмики, а частично вызывается колебаниями внешних факторов среды. Примечательно то, что годовая периодичность зависит не от непосредственно действующих на организм мощных экологических факторов (температуры, влажности и др.), которые подвержены сильной погодной изменчивости, а от второстепенных для жизнедеятельности свойств среды, которые, однако, очень закономерно изменяются в течение года. Приспособительный смысл этого явления в том, что кратковременные перемены погодных условий, их возможные значительные отклонения от нормы не меняют биологического ритма организмов, который остается синхронизованным с общим ходом изменений в природе в течение года.
Одним из наиболее точно и регулярно изменяющихся факторов среды является длина светового дня, ритм чередования темного и светлого периодов суток. Именно этот фактор служит большинству живых организмов для ориентации во времени года.
5.4. Фотопериодизм
Реакция организмов на сезонные изменения длины дня получила название фотопериодизма. Его проявление зависит не от интенсивности освещения, а только от ритма чередования темного и светлого периодов суток.
Фотопериодическая реакция живых организмов имеет большое приспособительное значение, так как для подготовки к переживанию неблагоприятных условий или, наоборот, к наиболее интенсивной жизнедеятельности требуется довольно значительное время. Способность реагировать на изменение длины дня обеспечивает заблаговременные физиологические перестройки и пригнанность цикла к сезонным сменам условий. Ритм дня и ночи выступает как сигнал предстоящих изменений климатических факторов, обладающих сильным непосредственным воздействием на живой организм (температуры, влажности и др.). В отличие от других экологических факторов ритм освещения влияет лишь на те особенности физиологии, морфологии и поведения организмов, которые являются сезонными приспособлениями в их жизненном цикле. Образно говоря, фотопериодизм – это реакция организма на будущность.
Хотя фотопериодизм встречается во всех крупных систематических группах, он свойствен далеко не всем видам. Существует много видов с нейтральной фотопериодической реакцией, у которых физиологические перестройки в цикле развития не зависят от длины дня. У таких видов либо развиты другие способы регулирования жизненного цикла (например, озимость у растений), либо они не нуждаются в точном его регулировании. Например, там, где нет резко выраженных сезонных изменений, большинство видов не обладает фотопериодизмом. Цветение, плодоношение и отмирание листьев у многих тропических деревьев растянуто во времени, и на дереве одновременно встречаются и цветки и плоды. В умеренном климате виды, успевающие быстро завершить жизненный цикл и практически не встречающиеся в активном состоянии в неблагоприятные сезоны года, также не проявляют фотопериодических реакций, например многие многолетние эфемероидные растения и эфемеры.
Различают два типа фотопериодической реакции: короткодневный идлиннодневный. Известно, что длина светового дня, кроме времени года, зависит от географического положения местности. Короткодневные виды живут и произрастают в основном в низких широтах, а длиннодневные – в умеренных и высоких. У видов с обширными ареалами северные особи могут отличаться по типу фотопериодизма от южных. Таким образом, тип фотопериодизма – это экологическая, а не систематическая особенность вида.
У длиннодневных растений и животных увеличивающиеся весенний и раннелетний дни стимулируют ростовые процессы и подготовку к размножению. Укорачивающиеся дни второй половины лета и осени вызывают торможение роста и подготовку к зиме (рис. 66). Так, морозостойкость клевера и люцерны гораздо выше при выращивании растений на коротком дне, чем на длинном. У деревьев, растущих в городах близ уличных фонарей, осенний день оказывается удлиненным, в результате у них задерживается листопад и они чаще подвергаются обморожению.
Как показали исследования, короткодневные растения особенно чувствительны к фотопериоду, так как длина дня на их родине меняется в течение года мало, а сезонные климатические изменения могут быть очень значительными. Тропические виды фотопериодическая реакция подготавливает к сухому и дождливому сезонам. Некоторые сорта риса в Шри-Ланке, где общее годовое изменение длины дня составляет не более часа, улавливают даже ничтожную разницу в световом ритме, что определяет время их цветения.
Фотопериодизм насекомых может быть не только прямым, но и опосредованным. Например, у капустной корневой мухи зимняя диапауза возникает через воздействие качества пищи, которое изменяется в зависимости от физиологического состояния растения.
Длина светлого периода суток, обеспечивающая переход в очередную фазу развития, получила название критической длины дня для этой фазы. По мере повышения географической широты критическая длина дня возрастает. Например, переход в диапаузу яблоневой листовертки на широте 32° происходит при продолжительности светлого периода суток, равной 14 ч, 44° – 16 ч, 52° – 18 ч. Критическая длина дня часто служит препятствием для широтного передвижения растений и животных, для их интродукции.
Фотопериодизм растений и животных – наследственно закрепленное, генетически обусловленное свойство. Однако фотопериодическая реакция проявляется лишь при определенном воздействии других факторов среды, например в определенном интервале температур. При некотором сочетании экологических условий возможно естественное расселение видов в несвойственные им широты, несмотря на тип фотопериодизма. Так, в высокогорных притропических районах много растений длинного дня, выходцев из районов умеренного климата.
Для практических целей длину светового дня изменяют при выращивании культур в закрытом грунте, управляя продолжительностью освещения, увеличивают яйценоскость кур, регулируют размножение пушных зверей.
Средние многолетние сроки развития организмов определяются прежде всего климатом местности, именно к ним и приспособлены реакции фотопериодизма. Отклонения от этих сроков обусловливаются погодной обстановкой. При изменении погодных условий сроки прохождения отдельных фаз могут в определенных пределах изменяться. Это особенно сильно проявляется у растений и пойкилотермных животных. Так, растения, не набравшие необходимой суммы эффективных температур, не могут зацвести даже в условиях фотопериода, стимулирующих переход в генеративное состояние. Например, в Подмосковье береза зацветает в среднем 8 мая при накоплении суммы эффективных температур 75 °C. Однако в годовых отклонениях сроки ее зацветания изменяются от 19 апреля до 28 мая. Гомойотермные животные отвечают на особенности погоды изменением поведения, сроков гнездования, миграций.
Изучением закономерностей сезонного развития природы занимается особая прикладная отрасль экологии – фенология (дословный перевод с греческого – наука о явлениях).
Согласно биоклиматическому закону Хопкинса, выведенному им применительно к условиям Северной Америки, сроки наступления различных сезонных явлений (фенодат) различаются в среднем на 4 дня на каждый градус широты, на каждые 5° долготы и на 120 м высоты над уровнем моря, т. е. чем севернее, восточнее и выше местность, тем позже наступление весны и раньше – осени. Кроме того, фенологические даты зависят от местных условий (рельефа, экспозиции, удаленности от моря и т. п.). На территории Европы сроки наступления сезонных событий изменяются на каждый градус широты не на 4, а на 3 дня. Соединяя на карте точки с одинаковыми фенодатами, получают изолинии, отражающие фронт продвижения весны и наступления очередных сезонных явлений. Это имеет большое значение для планирования многих хозяйственных мероприятий, в частности сельскохозяйственных работ.
studfiles.net
Циркадные ритмы Википедия
 Некоторые показатели циркадного (24-часового) биологического ритма человека
Некоторые показатели циркадного (24-часового) биологического ритма человека Цирка́дные (циркадианные) ритмы (от лат. circa «около, кругом» + dies «день») — циклические колебания интенсивности различных биологических процессов, связанные со сменой дня и ночи. Период циркадных ритмов обычно близок к 24 часам.
Несмотря на связь с внешними стимулами, циркадные ритмы имеют эндогенное происхождение, представляя, таким образом, биологические часы организма.[1]
Циркадные ритмы присутствуют у таких организмов как цианобактерии[2], грибы, растения, животные.
История открытия[ | код]
Впервые об изменении положения листьев в течение дня у тамаринда (Tamarindus indicus) упоминает описывавший походы Александра Македонского Андростен.
В Новое время в 1729 году французский астроном Жан-Жак де Меран сообщил о ежедневных движениях листьев у мимозы стыдливой (Mimosa pudica). Эти движения повторялись с определённой периодичностью даже если растения помещались в темноту, где отсутствовали такие внешние стимулы как свет, что позволило предположить эндогенное происхождение биологических ритмов, к которым были приурочены движения листьев растения. Де Мейрен предположил, что эти ритмы могут иметь что-то общее с чередованием сна и бодрствования у человека.
Декандоль в 1834 году определил, что период, с которыми растения мимозы совершают данные листовые движения, короче длины суток и составляет примерно 22-23 часа.
В 1880 году Чарльз Дарвин и его сын Фрэнсис сделали предположение о наследственной природе циркадных ритмов. Предположение о наследственной природе циркадных ритмов было подтверждено окончательно опытами, во время которых скрещивались растения фасоли, периоды циркадных ритмов которых различались. У гибридов длина периода отличалась от длины периода у обоих родителей.
Эндогенная природа циркадных ритмов была окончательно подтверждена в 1984 году во время опытов с грибами вида Нейроспора густая (Neurospora crassa), проведёнными в космосе. Эти опыты показали независимость околосуточных ритмов от геофизических сигналов, связанных с вращением Земли вокруг своей оси.
В 1970-е годы Сеймур Бензер и его ученик Рональд Конопка изучали, можно ли идентифицировать гены, которые контролируют циркадный ритм у плодовых мух. Они продемонстрировали, что мутации неизвестного гена нарушают циркадные часы мух. Неизвестный ген получил название ген периода — Per (от англ. period).
В 1984 году Джеффри Холл и Майкл Росбаш, работающие в тесном сотрудничестве в Брандейском университете в Бостоне, и Майкл Янг из Рокфеллеровского университета в Нью-Йорке смогли выделить ген Per. Затем Джеффри Холл и Майкл Росбаш обнаружили, что белок PER, кодируемый геном Per, накапливается в течение ночи и деградирует в течение дня. Таким образом, уровень белка PER колеблется в течение суток синхронно с циркадным ритмом. Учёные предположили, что белок PER блокирует активность гена Per. Они обосновали, что с помощью ингибирующей петли обратной связи белок может препятствовать своему собственному синтезу и тем самым регулировать собственный уровень в непрерывном циклическом ритме. Однако, чтобы блокировать активность гена Per, белок PER, который продуцируется в цитоплазме, должен был каким-то образом достигнуть клеточного ядра, где расположен генетический материал, — этот вопрос оставался невыясненным.
В 1994 году Майкл Янг обнаружил второй «часовой ген» циркадного ритма, timeless, кодирующий белок TIM, который требовался для нормального циркадного ритма. Майкл Янг показал, что когда белок TIM связан с белком PER, оба белка могут проникать в ядро клетки, где они блокируют активность гена Per, таким образом замыкая ингибирующую петлю обратной связи. Майкл Янг идентифицировал ещё один ген, doubletime, кодирующий белок DBT, который задерживал накопление белка PER. Совместное действие обнаруженных генов обеспечило понимание, как корректируется циркадный ритм для более точного соответствия 24-часовому циклу.
В последующие годы были выяснены другие молекулярные компоненты механизма, объясняющие его стабильность и функционирование. Были определены дополнительные белки, необходимые для активации гена Per, а также механизм, посредством которого свет может синхронизировать цикл.
В 2017 году Джеффри Холл, Майкл Росбаш и Майкл Янг были удостоены Нобелевской премии за открытие молекулярных механизмов, контролирующих циркадный ритм
ru-wiki.ru
Циркадные ритмы Википедия
Некоторые показатели циркадного (24-часового) биологического ритма человека Цирка́дные (циркадианные) ритмы (от лат. circa «около, кругом» + dies «день») — циклические колебания интенсивности различных биологических процессов, связанные со сменой дня и ночи. Период циркадных ритмов обычно близок к 24 часам.
Несмотря на связь с внешними стимулами, циркадные ритмы имеют эндогенное происхождение, представляя, таким образом, биологические часы организма.[1]
Циркадные ритмы присутствуют у таких организмов как цианобактерии[2], грибы, растения, животные.
История открытия
Впервые об изменении положения листьев в течение дня у тамаринда (Tamarindus indicus) упоминает описывавший походы Александра Македонского Андростен.
В Новое время в 1729 году французский астроном Жан-Жак де Меран сообщил о ежедневных движениях листьев у мимозы стыдливой (Mimosa pudica). Эти движения повторялись с определённой периодичностью даже если растения помещались в темноту, где отсутствовали такие внешние стимулы как свет, что позволило предположить эндогенное происхождение биологических ритмов, к которым были приурочены движения листьев растения. Де Мейрен предположил, что эти ритмы могут иметь что-то общее с чередованием сна и бодрствования у человека.
Декандоль в 1834 году определил, что период, с которыми растения мимозы совершают данные листовые движения, короче длины суток и составляет примерно 22-23 часа.
В 1880 году Чарльз Дарвин и его сын Фрэнсис сделали предположение о наследственной природе циркадных ритмов. Предположение о наследственной природе циркадных ритмов было подтверждено окончательно опытами, во время которых скрещивались растения фасоли, периоды циркадных ритмов которых различались. У гибридов длина периода отличалась от длины периода у обоих родителей.
Эндогенная природа циркадных ритмов была окончательно подтверждена в 1984 году во время опытов с грибами вида Нейроспора густая (Neurospora crassa), проведёнными в космосе. Эти опыты показали независимость околосуточных ритмов от геофизических сигналов, связанных с вращением Земли вокруг своей оси.
В 1970-е годы Сеймур Бензер и его ученик Рональд Конопка изучали, можно ли идентифицировать гены, которые контролируют циркадный ритм у плодовых мух. Они продемонстрировали, что мутации неизвестного гена нарушают циркадные часы мух. Неизвестный ген получил название ген периода — Per (от англ. period).
В 1984 году Джеффри Холл и Майкл Росбаш, работающие в тесном сотрудничестве в Брандейском университете в Бостоне, и Майкл Янг из Рокфеллеровского университета в Нью-Йорке смогли выделить ген Per. Затем Джеффри Холл и Майкл Росбаш обнаружили, что белок PER, кодируемый геном Per, накапливается в течение ночи и деградирует в течение дня. Таким образом, уровень белка PER колеблется в течение суток синхронно с циркадным ритмом. Учёные предположили, что белок PER блокирует активность гена Per. Они обосновали, что с помощью ингибирующей петли обратной связи белок может препятствовать своему собственному синтезу и тем самым регулировать собственный уровень в непрерывном циклическом ритме. Однако, чтобы блокировать активность гена Per, белок PER, который продуцируется в цитоплазме, должен был каким-то образом достигнуть клеточного ядра, где расположен генетический материал, — этот вопрос оставался невыясненным.
В 1994 году Майкл Янг обнаружил второй «часовой ген» циркадного ритма, timeless, кодирующий белок TIM, который требовался для нормального циркадного ритма. Майкл Янг показал, что когда белок TIM связан с белком PER, оба белка могут проникать в ядро клетки, где они блокируют активность гена Per, таким образом замыкая ингибирующую петлю обратной связи. Майкл Янг идентифицировал ещё один ген, doubletime, кодирующий белок DBT, который задерживал накопление белка PER. Совместное действие обнаруженных генов обеспечило понимание, как корректируется циркадный ритм для более точного соответствия 24-часовому циклу.
В последующие годы были выяснены другие молекулярные компоненты механизма, объясняющие его стабильность и функционирование. Были определены дополнительные белки, необходимые для активации гена Per, а также механизм, посредством которого свет может синхронизировать цикл.
В 2017 году Джеффри Холл, Майкл Росбаш и Майкл Янг были удостоены Нобелевской премии за открытие молекулярных механизмов, контролирующих циркадный ритм[3].
Циркадные ритмы растений
Циркадные ритмы растений связаны со сменой дня и ночи и важны для адаптации растений к суточным колебаниям таких параметров как температура, освещение, влажность. Растения существуют в постоянно меняющемся мире, поэтому циркадные ритмы важны для того, чтобы растение могло дать надлежащий ответ на абиотический стресс. Изменение положения листьев в течение суток — лишь один из многих ритмических процессов у растений. В течение суток колеблются такие параметры, как активность ферментов, интенсивность газообмена и фотосинтетическая активность.
В способности растений распознавать чередование дня и ночи играет роль фитохромная система. Примером работы такой системы является ритм цветения у растения ‘’Pharbitis nil’’. Цветение у этого растения зависит от длины светового дня: если день короче определённого интервала, то растение цветет, если длиннее — вегетирует. В течение суток условия освещения меняются из-за того, что солнце находится под разными углами к горизонту, и соответственно меняется спектральный состав света, что воспринимается различными фитохромами которые возбуждаются светом с разной длиной волны. Так, вечером в спектре много дальних красных лучей, которые активизируют только фитохром А, давая растению сигнал о приближении ночи. Получив этот сигнал, растение принимает соответствующие меры. Важность фитохромов для температурной адаптации была выяснена во время опытов с трансгенными осинами ‘’Populus tremula’’, у которых продукция фитохрома А была повышена. Растениям постоянно «казалось», что они получают свет высокой интенсивности, и таким образом не могли адаптироваться к суточным колебаниям температуры и страдали от ночных заморозков.
При исследовании суточных ритмов у арабидопсис была также показана фотопериодичность работы трёх генов для белков CO, FKF1 и G1. Ген constans участвует в определении времени цветения. Синтез продукта гена — белка CO запускается комплексом из белков FKF1 и G1. В этом комплексе продукт гена FKF1 играет роль фоторецептора. Синтез белка CO запускается через 4 часа после начала освещения и останавливается в темноте. Синтезированный белок за ночь разрушается, и таким образом необходимая для цветения растения концентрация белка достигается только в условиях долгого летнего дня.
Циркадные ритмы у животных
Практически все животные приспосабливают свои физиологические и поведенческие процессы к суточным колебаниям абиотических параметров. Примером циркадного ритма у животных является цикл сон-бодрствование. У человека и у других животных существуют внутренние часы, которые идут даже в отсутствие внешних стимулов, они дают информацию о времени суток. Исследование молекулярно-биологической природы этих часов началось в 1960-х — 1970-х годах[4]. Сеймур Бензер и Рональд Конопка, работавшие в Калифорнийском технологическом институте, обнаружили три мутантные линии дрозофил, циркадные ритмы которых отличались от циркадных ритмов мушек дикого типа. Дальнейший анализ показал, что у мутантов изменения затрагивали аллели одного локуса, который был назван исследователями per (от period).
В отсутствие нормальных сигналов окружающей среды период околосуточной активности у мушек дикого типа составлял 24 часа, у мутантов per-s — 19 часов (short period[5]), у мутантов per-l — 29 часов (long period[5]), у мутантов per-0 вообще не наблюдалось никакого ритма. Впоследствии было обнаружено, что продукты генов per есть во многих клетках дрозофил, участвующих в продукции циркадного ритма насекомого. Более того, у мушек дикого типа наблюдаются циркадные колебания в концентрации матричной РНК (мРНК) гена per[4] и белка PER (по существующей в молекулярной биологии номенклатуре гены обозначаются строчными, а их белковые продукты — прописными буквами[6]), в то время как у мушек per-0, у которых нет циркадного ритма, экспрессия генов не наблюдается.
У млекопитающих главными генами, лежащими в основе циркадианного молекулярного осциллятора супрахиазматического ядра гипоталамуса, являются гены mPer1 и mPer2 («m» означает «mammalian», то есть period-ген млекопитающих). Экспрессия mPer1 и mPer2 регулируется транскрипционными факторами CLOCK и BMAL1. Гетеромеры CLOCK/BMAL1 связываются с промоторами генов mPer1 и mPer2, что инициирует их транскрипцию. Образующиеся в результате этого мРНК транслируются в цитоплазме клеток супрахиазматического ядра в белки mPER1 и mPER2. Эти белки проникают в ядра клеток и, будучи теперь уже связанными с белками mCRY1 и mCRY2, подавляют транскрипцию генов mPer1 и mPer2, связываясь с CLOCK/BMAL1-белками. Таким образом, по механизму отрицательной обратной связи формируется чередование подъёмов и спадов продукции мРНК, а затем и самих белков mPER1 и mPER2 с фазой, равной приблизительно 24 ч. Этот цикл подстраивается под ритм освещенности[7].
Существует несколько дополнительных молекулярных циклов, регулирующих циклическую экспрессию генов mPer1 и mPer2. Белок BMAL1 тоже синтезируется циклически, и его продукция находится в противофазе с ритмом экспрессии генов mPer1 и mPer2. Транскрипция гена Bmal1 индуцируется белком mPER2 и тормозится белком REV-ERBα. В промоторах генов Cry1 и Cry2 содержится та же нуклеотидная последовательность (Е-box), что и в промоторах генов mPer1 и mPer2, поэтому транскрипция генов Cry1 и Cry2 позитивно регулируется комплексом CLOCK/BMAL1. То же самое справедливо и для транскрипции гена Rev-Erbα[7].
Осцилляции, генерируемые на уровне этих генов и белковых продуктов их экспрессии, амплифицируются и распространяются за пределы циркадианного ритмоводителя супрахиазматического ядра по всему организму. Например, ген вазопрессина, одного из нейромедиаторов супрахиазматического ядра, также имеет промотор, содержащий E-box, в результате чего его экспрессия регулируется напрямую вышеописанным циркадианным осциллятором. За счёт вазопрессина циркадианный сигнал передаётся в другие отделы нервной системы. Другие нейромедиаторные системы, находящиеся под контролем супрахиазматического ядра — глутамат- и ГАМК-эргические, пептидергические и моноаминергические системы. Также имеется нейрогуморальный путь распространения циркадианного сигнала по всему организму с вовлечением эпифизарного гормона мелатонина[7].
Циркадные ритмы и цикл сон-бодрствование у человека
Эндогенная продолжительность циркадного ритма
Первые эксперименты по изоляции людей от источников времени (таких как часы и солнечный свет) привели к тому, что у подопытных вырабатывался 25-часовой циркадный ритм. Ошибка эксперимента состояла в том, что подопытным разрешалось включать и выключать свет по своему усмотрению. Чрезмерное использование искусственного освещения перед сном приводило к увеличению продолжительности ритма. Новое, более корректно проведенное, исследование показало, что длина эндогенного циркадного ритма составляет в среднем 24 часа 11 минут[8][9].
Синхронизация циркадного ритма с внешним миром
Одним из наиболее эффективных внешних сигналов (по сравнению с другими факторами[10]), поддерживающих 24-часовый цикл, является свет. Воздействие света в ранние утренние часы способствует опережению ритма и более раннему засыпанию, а в поздние вечерние часы — к задержке ритма и более позднему засыпанию. За этот механизм отвечают специальные клетки сетчатки глаза (ipRGC), которые напрямую посылают сигнал в супрахиазматическое ядро — центр контроля циркадных ритмов[11]. О важности этого механизма говорит то, что примерно половина полностью слепых людей имеют синдром не-24-часового периода сна-бодрствования. Их циркадный ритм не может подстроиться к 24-м часам и они засыпают каждый день все позже и позже[12].
Супрахиазмальное ядро управляет выработкой гормонов, регулирующих циркадную активность организма. Однако при этом отдельные органы, такие как сердце, печень, почки, имеют свои «внутренние часы» и могут «выбиваться из ритма», устанавливаемого супрахиазматическим ядром. Сигнал, поступающий в шишковидную железу, вызывает синтез и выделение в кровоток вызывающего сон нейрогормона мелатонина. У пожилых людей выделяется меньше мелатонина, что, вероятно, объясняет, почему они чаще страдают бессонницей. Большая часть исследователей полагает, что супрахиазматическое ядро отвечает за циркадные ритмы и за колебания параметров, связанных с циклом сон-бодрствование, например температуры тела, кровяного давления и диуреза [источник не указан 79 дней]
Проявления циркадианного ритма у человека
Некоторые авторы описывают суточные ритмы работы внутренних органов человека.[13] Поскольку статьи с такими сведениями практически или вовсе не цитируются в научной литературе, их ценность сомнительна.
Хронотип
Циркадная система человека имеет индивидуальные отличия. Наиболее ярким их проявлением служит хронотип. Он бывает ранним («жаворонки»), промежуточным («голуби») и поздним («совы»). Люди, относящиеся к раннему хронотипу, ложатся спать и просыпаются в среднем на два часа раньше «сов» и достигают пика интеллектуальной и физической активности в первой половине дня. У людей, относящихся к позднему хронотипу, максимум умственной и физической работоспособности приходится на вторую половину дня. Среди мужчин и двадцатилетней молодежи преобладают «совы», а дети и пожилые люди чаще «жаворонки»[14].
Нарушения циркадного ритма человека
С нарушениями циркадного ритма тесно связаны нарушения сна — джетлаг, расстройство, связанное со сменным графиком работы, бессонница выходного дня и т. п.[15] Также с нарушением циркадного ритма связывают такие нарушения сна, как[16][11]:
У взрослых во время сна уменьшается продукция мочи в связи с увеличением содержания вазопрессина в крови. У некоторых детей и взрослых, у которых цикличность колебаний содержания вазопрессина нарушена, уменьшение продукции мочи в ночное время не происходит, что приводит к неконтролируемому мочеиспусканию.[источник не указан 80 дней]
Такое заболевание как фатальная семейная бессонница заканчивается летальным исходом и связана с врождёнными дефектами нейронов супрахиазматического ядра. Любопытным является то, что подобные симптомы возникают при болезни Крейтцфельдта-Якоба, когда поражаются клетки того же супрахиазматического ядра.[источник не указан 80 дней]
Примечания
- ↑ Красавин В.А., Лебедев А.Н., Бодров В.А., Луговой Л.А., Доскин В.А., Лаврентьева Н.А., Романов Ю.А. Биологические ритмы // Большая медицинская энциклопедия : в 30 т. / гл. ред. Б.В. Петровский. — 3 изд. — Москва : Советская энциклопедия, 1976. — Т. 3. Беклемишев - Валидол. — 584 с. — 150 000 экз.
- ↑ Опыт показал связь циркадного ритма и клеточного деления. — 23 марта 2010
- ↑ Нобелевская премия по физиологии и медицине 2017
- ↑ 1 2 Химия и Жизнь - О летнем времени и биологических часах. www.hij.ru. Проверено 23 февраля 2017.
- ↑ 1 2 Влияние генов на поведение мушки-дрозофилы. biofile.ru. Проверено 23 февраля 2017.
- ↑ Хронобиология - Стр 20. StudFiles. Проверено 23 февраля 2017.
- ↑ 1 2 3 Разыграев А.В., Керкешко Г.О., Арутюнян А.В. Пути циркадианного контроля продукции гонадотропин- рилизинг-гормона // Журнал акушерства и женских болезней. — 2011. — Т. LX, вып. 2. — С. 88—98. — ISSN 1684-0461.
- ↑ Charles A. Czeisler, Jeanne F. Duffy, Theresa L. Shanahan, Emery N. Brown, Jude F. Mitchell, David W. Rimmer, Joseph M. Ronda, Edward J. Silva, James S. Allan, Jonathan S. Emens, Derk-Jan Dijk, Richard E. Kronauer Stability, Precision, and Near-24-Hour Period of the Human Circadian Pacemaker // Science : журнал. — 1999. — 25 июня (т. 284, № 5423). — DOI:10.1126/science.284.5423.2177. — PMID 10381883.
- ↑ William J. Cromie Human Biological Clock Set Back an Hour // The Harvard Gazette : сайт. — 1999. — 15 июля.
- ↑ Биологические ритмы. medbookaide.ru. Проверено 18 марта 2016.
- ↑ 1 2 Кельмансон И. А. Экологические и клинико-биологические аспекты нарушений циркадианных ритмов сон-бодрствование у детей и подростков // Междисциплинарный научный и прикладной журнал "Биосфера" : журнал. — 2015. — Т. 7, № 1.
- ↑ R L Sack, A J Lewy, M L Blood, L D Keith, H Nakagawa Circadian rhythm abnormalities in totally blind people: incidence and clinical significance // The Journal of Clinical Endocrinology & Metabolism : журнал. — 1992. — 1 июля (т. 75, № 1). — С. 1127—134. — DOI:10.1210/jcem.75.1.1619000. — PMID 1619000.
- ↑ Земскова Ю.А. Биоритмы и часы работы внутренних органов // Наука и современность : журнал. — 2014. — № 27. — С. 31—35.
- ↑ Химия и Жизнь - Часовые пояса с точки зрения хронобиологии. www.hij.ru. Проверено 16 марта 2016.
- ↑ Комплекс статей по циркадным нарушениям ритма (популярные, научные)
- ↑ Международная классификация болезней 10-го пересмотра. ICD10data.com. Проверено 11 апреля 2018.
Литература
- Purves D. et al (2004). ‘’Neuroscience’’. Sinauer Associates, Inc. Publishers Sunderland, Massachusetts U.S.A
- Алехина Н. Д. и др (2005) ‘’Физиология растений’’. М.: Издательский центр «Академия»
- McClung C. (2006). ‘’Plant Circadian rhythms’’
- Zehring W. A., Wheeler D. A., Reddy P., Konopka R. J., Kyriacou C. P., Rosbash M., and Hall J. C. (1984). P-element transformation with period locus DNA restores rhythmicity to mutant, arrhythmic Drosophila melanogaster. Cell 39, 369—376.
- Bargiello T. A., Jackson F. R., and Young M. W. (1984). Restoration of circadian behavioural rhythms by gene transfer in Drosophila. Nature 312, 752—754.
- Siwicki K. K., Eastman C., Petersen G., Rosbash M., Hall J. C. (1988). Antibodies to the period gene product of Drosophila reveal diverse tissue distribution and rhythmic changes in the visual system. Neuron 1, 141—150.
- Hardin P. E., Hall J. C., and Rosbash M. (1990). Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels. Nature 343, 536—540.
- Liu X., Zwiebel L. J., Hinton D., Benzer S., Hall J. C., and Rosbash M. (1992). The period gene encodes a predominantly nuclear protein in adult Drosophila. J Neurosci 12, 2735—2744.
- Vosshall L. B., Price J. L., Sehgal A., Saez L., and Young M. W. (1994). Block in nuclear localization of period protein by a second clock mutation, timeless. Science 263, 1606—1609.
- Price J. L., Blau J., Rothenfluh A., Abodeely M., Kloss B., and Young M. W. (1998). double-time is a novel Drosophila clock gene that regulates PERIOD protein accumulation. Cell 94, 83-95.
Ссылки
wikiredia.ru
Циркадный ритм — WiKi
Некоторые показатели циркадного (24-часового) биологического ритма человека Циркадные (циркадианные) ритмы (от лат. circa — около, кругом и лат. dies — день) — циклические колебания интенсивности различных биологических процессов, связанные со сменой дня и ночи. Несмотря на связь с внешними стимулами, циркадные ритмы имеют эндогенное происхождение, представляя, таким образом, биологические часы организма. Циркадные ритмы присутствуют у таких организмов как цианобактерии[1], грибы, растения, животные. Период циркадных ритмов обычно близок к 24 часам.
История открытия
Впервые об изменении положения листьев в течение дня у тамаринда (Tamarindus indicus) упоминает описывавший походы Александра Македонского Андростен.
В Новое время в 1729 году французский астроном Жан-Жак де Меран сообщил о ежедневных движениях листьев у мимозы стыдливой (Mimosa pudica). Эти движения повторялись с определённой периодичностью даже если растения помещались в темноту, где отсутствовали такие внешние стимулы как свет, что позволило предположить эндогенное происхождение биологических ритмов, к которым были приурочены движения листьев растения. Де Мейрен предположил, что эти ритмы могут иметь что-то общее с чередованием сна и бодрствования у человека.
Декандоль в 1834 году определил, что период, с которыми растения мимозы совершают данные листовые движения, короче длины суток и составляет примерно 22-23 часа.
В 1880 году Чарльз Дарвин и его сын Фрэнсис сделали предположение о наследственной природе циркадных ритмов. Предположение о наследственной природе циркадных ритмов было подтверждено окончательно опытами, во время которых скрещивались растения фасоли, периоды циркадных ритмов которых различались. У гибридов длина периода отличалась от длины периода у обоих родителей.
Эндогенная природа циркадных ритмов была окончательно подтверждена в 1984 году во время опытов с грибами вида Нейроспора густая (Neurospora crassa), проведёнными в космосе. Эти опыты показали независимость околосуточных ритмов от геофизических сигналов, связанных с вращением Земли вокруг своей оси.
В 1970-е годы Сеймур Бензер и его ученик Рональд Конопка изучали, можно ли идентифицировать гены, которые контролируют циркадный ритм у плодовых мух. Они продемонстрировали, что мутации неизвестного гена нарушают циркадные часы мух. Неизвестный ген получил название ген периода — Per (от англ. period).
В 1984 году Джеффри Холл и Майкл Росбаш, работающие в тесном сотрудничестве в Брандейском университете в Бостоне, и Майкл Янг из Рокфеллеровского университета в Нью-Йорке смогли выделить ген Per. Затем Джеффри Холл и Майкл Росбаш обнаружили, что белок PER, кодируемый геном Per, накапливается в течение ночи и деградирует в течение дня. Таким образом, уровень белка PER колеблется в течение суток синхронно с циркадным ритмом. Учёные предположили, что белок PER блокирует активность гена Per. Они обосновали, что с помощью ингибирующей петли обратной связи белок может препятствовать своему собственному синтезу и тем самым регулировать собственный уровень в непрерывном циклическом ритме. Однако, чтобы блокировать активность гена Per, белок PER, который продуцируется в цитоплазме, должен был каким-то образом достигнуть клеточного ядра, где расположен генетический материал, — этот вопрос оставался невыясненным.
В 1994 году Майкл Янг обнаружил второй «часовой ген» циркадного ритма, timeless, кодирующий белок TIM, который требовался для нормального циркадного ритма. Майкл Янг показал, что когда белок TIM связан с белком PER, оба белка могут проникать в ядро клетки, где они блокируют активность гена Per, таким образом замыкая ингибирующую петлю обратной связи. Майкл Янг идентифицировал ещё один ген, doubletime, кодирующий белок DBT, который задерживал накопление белка PER. Совместное действие обнаруженных генов обеспечило понимание, как корректируется циркадный ритм для более точного соответствия 24-часовому циклу.
В последующие годы были выяснены другие молекулярные компоненты механизма, объясняющие его стабильность и функционирование. Были определены дополнительные белки, необходимые для активации гена Per, а также механизм, посредством которого свет может синхронизировать цикл.
В 2017 году Джеффри Холл, Майкл Росбаш и Майкл Янг были удостоены Нобелевской премии за открытие молекулярных механизмов, контролирующих циркадный ритм[2].
Циркадные ритмы растений
Циркадные ритмы растений связаны со сменой дня и ночи и важны для адаптации растений к суточным колебаниям таких параметров как температура, освещение, влажность. Растения существуют в постоянно меняющемся мире, поэтому циркадные ритмы важны для того, чтобы растение могло дать надлежащий ответ на абиотический стресс. Изменение положения листьев в течение суток — лишь один из многих ритмических процессов у растений. В течение суток колеблются такие параметры, как активность ферментов, интенсивность газообмена и фотосинтетическая активность.
В способности растений распознавать чередование дня и ночи играет роль фитохромная система. Примером работы такой системы является ритм цветения у растения ‘’Pharbitis nil’’. Цветение у этого растения зависит от длины светового дня: если день короче определённого интервала, то растение цветет, если длиннее — вегетирует. В течение суток условия освещения меняются из-за того, что солнце находится под разными углами к горизонту, и соответственно меняется спектральный состав света, что воспринимается различными фитохромами которые возбуждаются светом с разной длиной волны. Так, вечером в спектре много дальних красных лучей, которые активизируют только фитохром А, давая растению сигнал о приближении ночи. Получив этот сигнал, растение принимает соответствующие меры. Важность фитохромов для температурной адаптации была выяснена во время опытов с трансгенными осинами ‘’Populus tremula’’, у которых продукция фитохрома А была повышена. Растениям постоянно «казалось», что они получают свет высокой интенсивности, и таким образом не могли адаптироваться к суточным колебаниям температуры и страдали от ночных заморозков.
При исследовании суточных ритмов у арабидопсис была также показана фотопериодичность работы трёх генов для белков CO, FKF1 и G1. Ген constans участвует в определении времени цветения. Синтез продукта гена — белка CO запускается комплексом из белков FKF1 и G1. В этом комплексе продукт гена FKF1 играет роль фоторецептора. Синтез белка CO запускается через 4 часа после начала освещения и останавливается в темноте. Синтезированный белок за ночь разрушается, и таким образом необходимая для цветения растения концентрация белка достигается только в условиях долгого летнего дня.
Циркадные ритмы у животных
Практически все животные приспосабливают свои физиологические и поведенческие процессы к суточным колебаниям абиотических параметров. Примером циркадного ритма у животных является цикл сон-бодрствование. У человека и у других животных существуют внутренние часы, которые идут даже в отсутствие внешних стимулов, они дают информацию о времени суток. Исследование молекулярно-биологической природы этих часов началось в 1960-х — 1970-х годах[3]. Сеймур Бензер и Рональд Конопка, работавшие в Калифорнийском технологическом институте, обнаружили три мутантные линии дрозофил, циркадные ритмы которых отличались от циркадных ритмов мушек дикого типа. Дальнейший анализ показал, что у мутантов изменения затрагивали аллели одного локуса, который был назван исследователями per (от period).
В отсутствие нормальных сигналов окружающей среды период околосуточной активности у мушек дикого типа составлял 24 часа, у мутантов per-s — 19 часов (short period[4]), у мутантов per-l — 29 часов (long period[4]), у мутантов per-0 вообще не наблюдалось никакого ритма. Впоследствии было обнаружено, что продукты генов per есть во многих клетках дрозофил, участвующих в продукции циркадного ритма насекомого. Более того, у мушек дикого типа наблюдаются циркадные колебания в концентрации матричной РНК (мРНК) гена per[3] и белка PER (по существующей в молекулярной биологии номенклатуре гены обозначаются строчными, а их белковые продукты — прописными буквами[5]), в то время как у мушек per-0, у которых нет циркадного ритма, экспрессия генов не наблюдается.
У млекопитающих главными генами, лежащими в основе циркадианного молекулярного осциллятора супрахиазматического ядра гипоталамуса, являются гены mPer1 и mPer2 («m» означает «mammalian», то есть period-ген млекопитающих). Экспрессия mPer1 и mPer2 регулируется транскрипционными факторами CLOCK и BMAL1. Гетеромеры CLOCK/BMAL1 связываются с промоторами генов mPer1 и mPer2, что инициирует их транскрипцию. Образующиеся в результате этого мРНК транслируются в цитоплазме клеток супрахиазматического ядра в белки mPER1 и mPER2. Эти белки проникают в ядра клеток и, будучи теперь уже связанными с белками mCRY1 и mCRY2, подавляют транскрипцию генов mPer1 и mPer2, связываясь с CLOCK/BMAL1-белками. Таким образом, по механизму отрицательной обратной связи формируется чередование подъёмов и спадов продукции мРНК, а затем и самих белков mPER1 и mPER2 с фазой, равной приблизительно 24 ч. Этот цикл подстраивается под ритм освещенности[6].
Существует несколько дополнительных молекулярных циклов, регулирующих циклическую экспрессию генов mPer1 и mPer2. Белок BMAL1 тоже синтезируется циклически, и его продукция находится в противофазе с ритмом экспрессии генов mPer1 и mPer2. Транскрипция гена Bmal1 индуцируется белком mPER2 и тормозится белком REV-ERBα. В промоторах генов Cry1 и Cry2 содержится та же нуклеотидная последовательность (Е-box), что и в промоторах генов mPer1 и mPer2, поэтому транскрипция генов Cry1 и Cry2 позитивно регулируется комплексом CLOCK/BMAL1. То же самое справедливо и для транскрипции гена Rev-Erbα[6].
Осцилляции, генерируемые на уровне этих генов и белковых продуктов их экспрессии, амплифицируются и распространяются за пределы циркадианного ритмоводителя супрахиазматического ядра по всему организму. Например, ген вазопрессина, одного из нейромедиаторов супрахиазматического ядра, также имеет промотор, содержащий E-box, в результате чего его экспрессия регулируется напрямую вышеописанным циркадианным осциллятором. За счёт вазопрессина циркадианный сигнал передаётся в другие отделы нервной системы. Другие нейромедиаторные системы, находящиеся под контролем супрахиазматического ядра — глутамат- и ГАМК-эргические, пептидергические и моноаминергические системы. Также имеется нейрогуморальный путь распространения циркадианного сигнала по всему организму с вовлечением эпифизарного гормона мелатонина[6].
Циркадные ритмы и цикл сон-бодрствование у человека
Эндогенная продолжительность циркадного ритма
Первые эксперименты по изоляции людей от источников времени (таких как часы и солнечный свет) привели к тому, что у подопытных вырабатывался 25-часовой циркадный ритм. Ошибка эксперимента состояла в том, что подопытным разрешалось включать и выключать свет по своему усмотрению. Чрезмерное использование искуственного освещения перед сном приводило к увеличению продолжительности ритма. Новое, более корректно проведенное, исследование показало, что длина эндогенного циркадного ритма составляет в среднем 24 часа 11 минут[7][8].
Синхронизация циркадного ритма с внешним миром
Одним из наиболее эффективных внешних сигналов (по сравнению с другими факторами[9]), поддерживающих 24-часовый цикл, является свет. Воздействие света в ранние утренние часы способствует опережению ритма и более раннему засыпанию, а в поздние вечерние часы — к задержке ритма и более позднему засыпанию. За этот механизм отвечают специальные клетки сетчатки глаза (ipRGC), которые напрямую посылают сигнал в супрахиазматическое ядро — центр контроля циркадных ритмов[10]. О важности этого механизма говорит то, что примерно половина полностью слепых людей имеют синдром не-24-часового периода сна-бодрствования. Их циркадный ритм не может подстроиться к 24-м часам и они засыпают каждый день все позже и позже[11].
Супрахиазмальное ядро управляет выработкой гормонов, регулирующих циркадную активность организма. Однако при этом отдельные органы, такие как сердце, печень, почки, имеют свои «внутренние часы» и могут «выбиваться из ритма», устанавливаемого супрахиазматическим ядром. Сигнал, поступающий в шишковидную железу, вызывает синтез и выделение в кровоток вызывающего сон нейрогормона мелатонина. У пожилых людей выделяется меньше мелатонина, что, вероятно, объясняет, почему они чаще страдают бессонницей. Большая часть исследователей полагает, что супрахиазматическое ядро отвечает за циркадные ритмы и за колебания параметров, связанных с циклом сон-бодрствование, например температуры тела, кровяного давления и диуреза [источник не указан 25 дней]
Проявления циркадианного ритма у человека
Некоторые авторы описывают суточные ритмы работы внутренних органов человека.[12] Поскольку статьи с такими сведениями практически или вовсе не цитируются в научной литературе, их ценность сомнительна.
Хронотип
Циркадная система человека имеет индивидуальные отличия. Наиболее ярким их проявлением служит хронотип. Он бывает ранним («жаворонки»), промежуточным («голуби») и поздним («совы»). Люди, относящиеся к раннему хронотипу, ложатся спать и просыпаются в среднем на два часа раньше «сов» и достигают пика интеллектуальной и физической активности в первой половине дня. У людей, относящихся к позднему хронотипу, максимум умственной и физической работоспособности приходится на вторую половину дня. Среди мужчин и двадцатилетней молодежи преобладают «совы», а дети и пожилые люди чаще «жаворонки»[13].
Нарушения циркадного ритма человека
С нарушениями циркадного ритма тесно связаны нарушения сна — джетлаг, расстройство, связанное со сменным графиком работы, бессонница выходного дня и т. п.[14] Также с нарушением циркадного ритма связывают такие нарушения сна, как[15][10]:
- синдром задержки фазы сна — характеризуется поздним засыпанием и пробуждением с невозможностью сместить время сна на более ранние часы;
- синдром опережения фазы сна — характеризуется слишком ранним наступлением сонливости и ранним пробуждением;
- синдром не-24-часового сна-бодрствования — характеризуется ежедневным смещением времени засыпания и пробуждения на более позднее время;
- нерегулярный ритм сна-бодрствования — характеризуется отсутствием стабильного ритма сна-бодрствования; пациенты спят несколько раз в сутки в разное время.
У взрослых во время сна уменьшается продукция мочи в связи с увеличением содержания вазопрессина в крови. У некоторых детей и взрослых, у которых цикличность колебаний содержания вазопрессина нарушена, уменьшение продукции мочи в ночное время не происходит, что приводит к неконтролируемому мочеиспусканию.[источник не указан 26 дней]
Такое заболевание как фатальная семейная бессонница заканчивается летальным исходом и связана с врождёнными дефектами нейронов супрахиазматического ядра. Любопытным является то, что подобные симптомы возникают при болезни Крейтцфельдта-Якоба, когда поражаются клетки того же супрахиазматического ядра.[источник не указан 26 дней]
Примечания
- ↑ Опыт показал связь циркадного ритма и клеточного деления. — 23 марта 2010
- ↑ Нобелевская премия по физиологии и медицине 2017
- ↑ 1 2 Химия и Жизнь - О летнем времени и биологических часах. www.hij.ru. Проверено 23 февраля 2017.
- ↑ 1 2 Влияние генов на поведение мушки-дрозофилы. biofile.ru. Проверено 23 февраля 2017.
- ↑ Хронобиология - Стр 20. StudFiles. Проверено 23 февраля 2017.
- ↑ 1 2 3 Разыграев А.В., Керкешко Г.О., Арутюнян А.В. Пути циркадианного контроля продукции гонадотропин- рилизинг-гормона // Журнал акушерства и женских болезней. — 2011. — Т. LX, вып. 2. — С. 88-98. — ISSN 1684-0461.
- ↑ Charles A. Czeisler, Jeanne F. Duffy, Theresa L. Shanahan, Emery N. Brown, Jude F. Mitchell, David W. Rimmer, Joseph M. Ronda, Edward J. Silva, James S. Allan, Jonathan S. Emens, Derk-Jan Dijk, Richard E. Kronauer Stability, Precision, and Near-24-Hour Period of the Human Circadian Pacemaker // Science : журнал. — 1999. — 25 июня (т. 284, № 5423). — DOI:10.1126/science.284.5423.2177. — PMID 10381883.
- ↑ William J. Cromie Human Biological Clock Set Back an Hour // The Harvard Gazette : сайт. — 1999. — 15 июля.
- ↑ Биологические ритмы. medbookaide.ru. Проверено 18 марта 2016.
- ↑ 1 2 Кельмансон И. А. Экологические и клинико-биологические аспекты нарушений циркадианных ритмов сон-бодрствование у детей и подростков // Междисциплинарный научный и прикладной журнал "Биосфера" : журнал. — 2015. — Т. 7, № 1.
- ↑ R L Sack, A J Lewy, M L Blood, L D Keith, H Nakagawa Circadian rhythm abnormalities in totally blind people: incidence and clinical significance // The Journal of Clinical Endocrinology & Metabolism : журнал. — 1992. — 1 июля (т. 75, № 1). — С. 1127-134. — DOI:10.1210/jcem.75.1.1619000. — PMID 1619000.
- ↑ Земскова Ю.А. Биоритмы и часы работы внутренних органов // Наука и современность : журнал. — 2014. — № 27. — С. 31-35.
- ↑ Химия и Жизнь - Часовые пояса с точки зрения хронобиологии. www.hij.ru. Проверено 16 марта 2016.
- ↑ Комплекс статей по циркадным нарушениям ритма (популярные, научные)
- ↑ Международная классификация болезней 10-го пересмотра. ICD10data.com. Проверено 11 апреля 2018.
Литература
- Purves D. et al (2004). ‘’Neuroscience’’. Sinauer Associates, Inc. Publishers Sunderland, Massachusetts U.S.A
- Алехина Н. Д. и др (2005) ‘’Физиология растений’’. М.: Издательский центр «Академия»
- McClung C. (2006). ‘’Plant Circadian rhythms’’
- Zehring W. A., Wheeler D. A., Reddy P., Konopka R. J., Kyriacou C. P., Rosbash M., and Hall J. C. (1984). P-element transformation with period locus DNA restores rhythmicity to mutant, arrhythmic Drosophila melanogaster. Cell 39, 369—376.
- Bargiello T. A., Jackson F. R., and Young M. W. (1984). Restoration of circadian behavioural rhythms by gene transfer in Drosophila. Nature 312, 752—754.
- Siwicki K. K., Eastman C., Petersen G., Rosbash M., Hall J. C. (1988). Antibodies to the period gene product of Drosophila reveal diverse tissue distribution and rhythmic changes in the visual system. Neuron 1, 141—150.
- Hardin P. E., Hall J. C., and Rosbash M. (1990). Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels. Nature 343, 536—540.
- Liu X., Zwiebel L. J., Hinton D., Benzer S., Hall J. C., and Rosbash M. (1992). The period gene encodes a predominantly nuclear protein in adult Drosophila. J Neurosci 12, 2735—2744.
- Vosshall L. B., Price J. L., Sehgal A., Saez L., and Young M. W. (1994). Block in nuclear localization of period protein by a second clock mutation, timeless. Science 263, 1606—1609.
- Price J. L., Blau J., Rothenfluh A., Abodeely M., Kloss B., and Young M. W. (1998). double-time is a novel Drosophila clock gene that regulates PERIOD protein accumulation. Cell 94, 83-95.
Ссылки
ru-wiki.org










