
13.2. Репродуктивные процессы в цветке. Зрелое пыльцевое зерно у цветковых растений содержит
Пыльцевое зерно - Справочник химика 21
Оплодотворение у цветковых растений происходит следующим образом пыльцевые зерна, которые поп али на рыльце пестика, прорастают, и из них развиваются пыльцевые трубки, растущие в ткани пестика к зародышевому мешку (фиг. 12). Эти пыльцевые зерна несут два ядра спермиев одно из них оплодотворяет яйцеклетку, а другое — сливается с центральным ядром (фиг. 11). Итак, у цветковых растений происходит (как правило) двойное оплодотворение, после оплодотворения из яйцеклетки путем последовательных митозов возникает [c.37] Зрелые пыльцевые зерна [c.36]В цветочных тычинках, совокупность которых называется андроцеем, образуется пыльца (микроспоры), служащие для опыления и в дальнейшем для оплодотворения. Андроцей является мужским половым аппаратом растений — пыльцевое зерно, пыльцевая клетка, каждая из разъединенных клеток, находящихся в тычинках при прорастании пылинок образуют спермин, оплодотворяющие яйцеклетку. Применение пыльцы для-косметики получает все более широкое распространение. [c.123]
Люминесцентная микроскопия. Рассматривают сухой порошок или отдельные части соцветия, цветка наблюдается собственная (первичная) флюоресценция сырья в ультрафиолетовом свете. Наиболее характерное свечение имеют кутикула, различные трихомы (волоски, железки), механические элементы, пыльцевые зерна, включения клеток в зависимости от их химического состава. [c.258]
Сходная дифференциация, при которой наследственная конституция пыльцевых зерен определяет их особенности, обнаружена и у растений, принадлежащих к другим родам. На фиг. 18 показаны различные пыльцевые зерна одного гибридного растения из рода энотера. Пыльца содержит зерна обоих типов в равном количестве. Половина пыльцевых зерен имеют относительно крупные размеры и содержат очень мелкие [c.45]
Цветки. Чаще готовят препараты из порошка цветков или отдельных частей цветка (соцветия), которые рассматривают обычно без включающей жидкости. В цветках часто содержатся флавоноиды, каротиноиды и другие вещества, обладающие флюоресценцией. Отчетливо видны пыльцевые зерна, имеющие желтое, зеленовато-желтое или голубоватое свечение. [c.284]
| Фиг. 152. Гаплоидное (Л), диплоидное (Б) и тетра-плоидное (В) пыльцевые зерна одного сорта гиацинта с 12, 24 и 48 хромосомами соответственно. |
У цветковых растений гаплофаза редуцирована еще больше и представлена лишь пыльцевыми зернами и зародышевым мешком. Пыльцевые зерна еще можно считать до известной степени свободными организмами, тогда как зародышевый мешок неотделим от материнского растения. Известно, что пыльцевые зерна образуются в пыльниках, которые на ранней стадии развития содержат обычно больщое число материнских клеток пыльцы (фиг. 10, вверху слева). Мейоз происходит в материнских клетках пыльцы, которые в результате превращаются в тетрады, т. е. в группы из четырех гаплоидных клеток. Эти клетки затем отделяются друг от друга и образуют зрелые пыльцевые зерна. Сначала эти пыльцевые зерна имеют лишь по одному ядру, но затем путем митоза они превращаются в пыльцевые зерна, состоящие из двух клеток— вегетативной и генеративной. Ядро вегетативной клетки располагается в центре пыльцевого зерна и больше не делится генеративное же ядро делится еще раз и дает начало двум ядрам спермиев каждое из них локализуется в отдельной клетке (фиг. 10, нижний ряд слева). Такое пыльцевое зерно созрело и способно к оплодотворению. [c.35]
Первый митоз в пыльцевом зерне [c.36]
У кукурузы имеется пара аллелей W-w, действие которых проявляется уже в пыльцевых зернах. Доминантный аллель [c.44]
Подсчет, проведенный в одном из исследований, показал, что на 3437 синих зерен приходилось 3482 неокрашенных пыльцевых зерна, что очень близко к теоретически ожидаемому отношению 1 1. На фиг. 17 показана пыльца одного гибрида сорго, у которого наблюдается аналогичное явление. [c.44]
Ультрафиолетовые лучи имеют значительно большую длину волны, а потому и меньшую энергию они не вызывают ионизации и обладают меньшей проникающей способностью, чем обсуждавшиеся выше излучения. Однако ультрафиолетовые лучи могут проникать в мельчайшие объекты (например, пыльцевые зерна, бактерии, споры грибов), вызывая у [c.214]
Пыльцевые зерна, содержащие один из аллелей, темные, а зерна, содержащие другой аллель, светлые. [c.45]
Известно, что у цветковых растений каждая материнская клетка пыльцы дает четыре пыльцевых зерна, которые вначале соединены друг с другом и образуют тетраду. У мхов также образуются тетрады, состоящие из четырех гаплоидных спор, развившихся из одной материнской клетки. У мхов во многих случаях удалось провести тетрадный анализ были выделены отдельные тетрады и изучались индивидуально четыре растения, развившиеся из спор такой тетрады. При этом оказалось, что если коробочка мха была гетерозиготна по паре аллелей Аа), то в тетраде всегда были споры двух типов. Две из них давали начало растениям мха типа А, тогда как две другие споры давали растения типа а. [c.46]
И аЬ. Это одинаково относится как к пыльцевым зернам, так и к яйцеклеткам. Ввиду того что соединение гамет различных типов при образовании гибридами потомства происходит совершенно случайно, это случайное соединение можно выразить формулой АВ АЬ + аВ + аЬ) х АВ + АЬ + аВ - - аЬ). Результаты такого соединения станут более ясными, если 16 образующихся при этом комбинаций представить в виде следующей таблицы [c.52]
Генетический анализ самостерильности у разных видов растений показал, что она регулируется различными генетическими механизмами. Самый распространенный и вместе с тем наиболее тонкий способ связан с наличием множественных аллелей, проявляющих свое действие на стадии гаплофазы в пыльцевом зерне, а также в ткани пестика. Этот механизм впервые был выяснен на одном виде растений табака. Оказалось, что эти растения обычно гетерозиготны по аллелям самостерильности. Если эти аллели обозначить 51, 82, 5з, 54,,..5 , то одно растение может иметь конституцию 152, другое — 525,ч [c.154]
Как это будет подробнее объяснено в гл. XXV, подобные пыльцевые зерна и зародышевые мешки, как правило, функционируют хуже, чем те, у которых число хромосом соответствует таковому одного из исходных видов, т. е. равно 7 или 14. Особенно чувствительна к подобному несбалансированному числу хромосом пыльца поэтому среди пыльцевых зерен гибрида нормально функционируют лишь те, которые содержат 7 или 14 хромосом. А поскольку подавляющее большинство пыльцевых зерен гибрида имеет промежуточное число хромосом (8, 9, 10, 11, 12 или 13), то в результате получается очень высокий процент стерильной пыльцы, что можно обнаружить непосредственно под микроскопом. Только незначительная часть пыльцевых зерен имеет нормальный внешний вид, а в большинстве случаев они сморщены и более или менее пустые. Поэтому в таких случаях нетрудно определить процент нормальных по внешнему виду пыльцевых зерен. Опыты по прорастанию пыльцы позволяют получить еще более достоверные данные о ее способности к оплодотворению. [c.305]
Хромосомы видны в ядрах, находящихся в профазе пер-ВОГО митоза пыльцевого зерна. Видна зависимость между числом хромосом и размером пыльцевых зерен. [c.325]
В отличие от нормально образующихся четырех клеток — тетрад, содержащих редуцированное, т. е. половинное, число хромосом. Пыльцевые зерна, развивающиеся из клеток диады, вдвое крупнее обычных (фиг. 152, Л и Б). В других случаях тормозится второе деление мейоза, но при этом также в результате образуются нередуцированные пыльцевые зерна и яйцеклетки. В редких случаях возникают как пыльцевые зерна, так и зародышевые мешки, у которых число хромосом вдвое больше соматического (фиг. 152, В). [c.326]
Более того, иногда наблюдается элиминация хромосом, т. е. отдельные хромосомы остаются в цитоплазме, не включаясь ни в пыльцевое зерно, ни в зародышевый мешок. Таким образом, в большинстве случаев пыльцевые зерна и зародышевые мешки получают неполный, несбалансированный набор хромосом, дающий летальный эффект. Итак, в большинстве случаев гаплоид будет полностью или почти полностью стерильным. [c.331]
Вид АА образует пыльцевые зерна и яйцеклетки с хромосомным набором А. Однако у гибридов АВ мейоз часто бывает настолько сильно нарушен, что гаметы с редуцированным набором хромосом образоваться не могут. Мы уже раньше видели, что это объясняется отсутствием гомологии между оказавшимися вместе хромосомами разных видов, что обусловливает нарушение конъюгации хромосом и приводит к образованию нежизнеспособных гамет с несбалансированным хромосомным набором. [c.334]
Узнавание клеток Rhizobium корнями бобовых-всего липп. один пример специфического взаимодействия растительной клетки с другими клетками. Это могут быть клетки других видов (например, патогенных грибов) или того же вида (например, пыльцевого зерна, попавшего на рыльце пестика). В настоя- [c.179]
В тех случаях, когда скрещиваемые виды имеют одинаковое число хромосом, но конъюгация несколько нарушена вследствие структурных различий, во время мейоза возникают нарушения, сходные с теми нарушениями, которые наблюдаются у гибридов, полученных от скрещивания между видами с разным числом хромосом. В подобных случаях также возникает более или менее выраженная стерильность. Эта стерильность будет иной, чем та, в основе которой лежит ненормальное развитие самих половых органов. Мы различаем гаплонтную и диплонтную стерильность. К стерильности первого типа относятся те случаи, когда гаплонты, т. е. пыльцевые зерна и зародышевые мешки цветковых растений, гибнут вследствие их собственной конституции. Подобная ненормальная конституция является следствием случайного распределения хромосом при мейозе и заключается либо в чисто количественном нарушении хромосомного баланса (например, наличие лишних хромосом), либо в нарушениях баланса, [c.305]
Специфическое взаимодействие пыльцевых зерен с рыльцем пестика-хорошо изученный пример функционирования лектинов. Это взаимодействие побуждает клетки рыльца выделять воду, а увлажнение пыльцевого зерна в свою очередь индуцирует образование длинной пыльцевой трубки, необходимой для оплодотворения (рис. 19-26). Около половины всех известных цветковых растений имеют генетически детерминированные механизмы, препятствующие самоопылению и таким образом обеспечивающие аутбридинг. У крестоцветных, например, молекулярные компоненты системы узнавания-крупный гликопротеин, присутствующий в клейком секрете рыльца, и опознающий его лектин, находящийся на поверхности пыльцевых зерен,-кодируются генным комплексом 5. Пыльца прорастает на рыльце и оплодотворяет яйцеклетку только в том случае, если у скрещиваемых растений экспрессируются разные аллели генов этого комплекса. Если пыльцу предварительно обработать очищенным гликопротеином, выделенным из рыльца растения, у которого экспрессируется тот же самый набор аллелей, она не прорастет даже на рыльце совместимого партнера. По-видимому, взаимодействие пыльцевого лектина со своим гликопротеином служит сигналом, эффективно блокирующим прорастание пыльцевого зерна и, следовательно, самооплодотворение. Специфические лектины и их эндогенные рецепторы в настоящее время выделены и охарактеризованы для многих систем узнавания растительных клеток. [c.180]
В жизненном цикле цветковых растений преобладает диплофаза, а гаплофаза обычно ограничена пыльцевыми зернами и зародышевым мешком. Однако интересно отметить, что в исключительных случаях могут возникать целиком гаплоидные растения. Хотя эти гаплоилы имеют такое же число хромосом, как и гаметы диплоидного растения, они развиваются [c.329]
| Рис. 19-67. Из пыльцевого зерна можно вырастить растение с гаплоидным набором хромосом. Слева показано нормальное диплоидное растение табака, а справа-гаплоидное, выросшее из одного шлпьцевого зерна. Гаплоидные растения, иногда возникающие в природе, обычно отличаются от диплоидных меньшими размерами. (С любезного разрешения 1. Оип е11.) |
А 1 роиесс созревания мужских клеток. Б. Процесс созревания женских клеток. В обоих сл чаях мейоа приводит к образованию комплексов из четырех клеток (тетрад) с поло-винпым числом хромосом, из которых образуются пыльцевые зерна и зародышевый [c.36]
А. Продольный срез через цветок во время оплодотворения. Некоторые пыльцевые зерна проросли на рыльце, одна нз пыльцевых трубок достигла зародышевого мешка. Б. Продольный срез через верхнюю часть столбика. Видны прорастаюш,ие пыльцевые зерна с пыльцевыми трубками разной длины. У —пыльцевое зерно 2 —пыльцевая трубка 3 —рыльце — столбик 5 —семяпочка —покровы семяпочки 7 —зародышевый мешок 5 —ядро пыльцевой трубки 9 —спермин / — лепестки // — чашелистики /2 —пыльник с пыльцевыми зернами —завязь / — тычиночная нить /5 —антиподы /6 —полярные ядра /7 —синергиды, /5 —яйцеклетка /9 —микропиле —нектарные железы. [c.38]
Влияние различных генов на жизнеспособность может иногда проявляться в гаплофазе. Как показал Г. Нильссон, у растения Oenothera Lamar kiana, имеющего очень крупные цветки и пестики, пыльцевые зерна, несущие ген R (красные прожилки листьев), образуют пыльцевые трубки, которые растут быстрее, чем пыльцевые трубки из зерен, несущих ген г (белые прожилки листьев). Если пыльцу обоих типов нанести на рыльце одного и того же пестика, то между двумя типами пыльцы начинается настоящее соревнование в прорастании, из которого пыльца с геном R выходит победителем. Поэтому все это явление было названо сертацией. Подобный дифференциальный рост пыльцевых трубок, представляющий собой частный случай плейотропии, в дальнейшем был найден и у других видов цветковых растений. [c.68]
Если же растение SiSi опыляется пыльцой растения S3S4, то пыльцевые зерна прорастают быстро и развиваются нормальные трубки. В том случае, если пыльцевое зерно несет s-аллели, отличные от тех, которые присутствуют в тканях пестика (и во всем растении), рост пыльцевой трубки протекает нормально если же пыльцевое зерно несет тот же s-аллель, что и пестик, то рост тормозится. [c.155]
Эта зависимость особенно четко проявляется при такой комбинации перекрестного опыления, как S1S2 X S2S3. Оказалось, что в таком случае рост половины пыльцевых зерен задерживается, половина же растет нормально. Ясно также, что задерживается рост пыльцевых зерен S2, так как пестик содержит тот же самый аллель. В этом случае функционируют только пыльцевые зерна S3. [c.155]
Подобные случаи хорошо известны, например, для дурмана Datura stramonium). Однако чаще некоторая часть материнских клеток пыльцы (и материнских клеток зародышевого мешка) содержит прямые транслокационные кольца, в которых лежащие рядом хромосомы направляются к одному и тому же полюсу. В результате образуются абортивные пыльцевые зерна и зародышевые мешки. Хорошая пыльца часто составляет лишь 50%. Это показывает, что частота зигзагообразных и прямых колец в первой метафазе примерно одинакова. [c.173]
chem21.info
13.2. Репродуктивные процессы в цветке
В цветке последовательно протекают важнейшие процессы, обеспечивающие вначале бесполое размножение и образование очередного поколения гаметофитов, а затем и половое размножение, завершающееся образованием нового поколения спорофитов в виде зародышей, расположенных в семенах. Цветковые являются разноспоровыми растениями. Они образуют два типа спор: микро- и мегаспоры.
Клетки бесполого размножения – микроспоры, образуются в ходе процесса, называемого микроспорогенезом в микроспорангиях, которыми являются гнезда пыльника. В результате ряда последовательных митотических делений клеток археспория, то есть образовательной ткани пыльника, возникают материнские клетки микроспор (микроспороциты), содержащие, как и все растение, диплоидный набор хромосом. Далее каждая материнская клетка в результате мейотического деления образует тетраду гаплоидных микроспор. Сформировавшаяся микроспора имеет оболочку и единственное ядро.
В ходе превращения микроспоры в пыльцевое зерно происходит два митотических деления и под защитой оболочки микроспоры возникают сначала две, далее три клетки – редуцированный мужской гаметофит. После формирования такого редуцированного гаметофита трехклеточная структура называется пыльцевым зерном (мужской особью). Пыльцевое зерно в начале развития содержит две клетки: маленькую генеративную и более крупную, часто называемую вегетативной. Микрогаметогенез крайне упрощен. Генеративная клетка делится однократно и из нее образуются две безжгутиковые мужские гаплоидные гаметы – спермии. В таком состоянии пыльцевое зерно готово к половому размножению. При попадании на рыльце пестика цветка вегетативная клетка пыльцевого зерна удлиняется и становится так называемой «пыльцевой трубкой», с мощью которой уже в семязачатке завершается оплодотворение ( рис. 13.13).
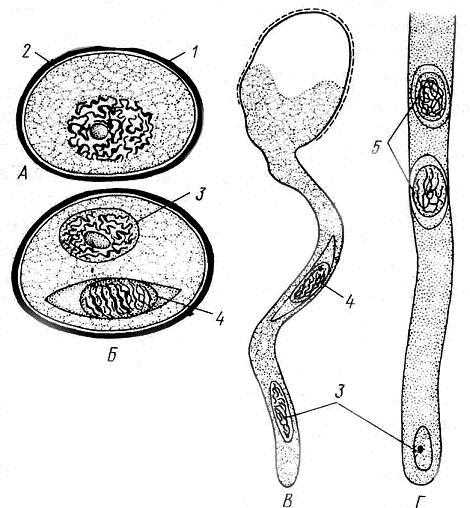
Рис. 13.13. Микроспора, пыльцевое зерно и его прорастание . А – микроспора; Б – пыльцевое зерно; В – формирование пыльцевой трубки; Г– часть пыльцевой трубки: 1 – экзина, 2 – интина, 3 – вегетативная клетка, дающая начало пыльцевой трубке, 4 – генеративная клетка, 5 – спермий.
Зрелое пыльцевое зерно окружено сложно устроенной оболочкой. Оболочка, часто называемая спородермой, состоит из двух главных слоев: внешнего, более толстого – экзины, и относительно тонкого внутреннего – интины. Экзина, содержащая особое высокомолекулярное вещество спорополленин, характеризуется необычайной стойкостью: она не растворяется в кислотах и щелочах, выдерживает температуру до 300 °С и сохраняется в фоссилизованном (окаменевшем) состоянии миллионы лет в геологических отложениях. Это позволяет вести спорово-пыльцевой анализ почв, донных отложений и осадочных толщ, определять, какие виды росли в разные геологические периоды. Экзина микроспор имеет на поверхности разнообразные скульптурные утолщения.
Особенности скульптуры важны при морфологической характеристике пыльцевых зерен отдельных таксонов. Наиболее развиты такие утолщения у энтомофильных растений. Интина менее стойка, чем экзина. Она состоит из целлюлозы и пектина и как бы облегает содержимое пыльцевого зерна.
Перенос пыльцевых зерен из тычинок на рыльца пестиков называется опылением. Для того чтобы образовался зародыш семени, должны произойти опыление и оплодотворение. Опыление впервые появляется у голосеменных, однако наибольшее разнообразие механизмов опыления и его совершенство достигаются у цветковых. Различают два типа опыления – самоопыление, или автогамию (от греческого «аутос» – сам), и перекрестное опыление, или ксеногамию (от греческого «ксенос» – чужой, «гамос» – брак). При самоопылении рыльце опыляется пыльцой того же цветка или пыльцой других цветков этой особи растения – гейтоногамия (девственное опыление). Обычно самоопыление осуществляется в распустившихся цветках, но иногда происходит в цветках закрытых, нераспустившихся (клейстогамных). В генетическом отношении все эти способы вполне равноценны.
Если перенос пыльцы осуществляется между цветками разных особей, то происходит перекрестное опыление. Оно свойственно не менее 90% видов цветковых растений. Перекрестное опыление обеспечивает обмен генами, поддерживает высокий уровень гетерозиготности популяций, определяет единство и целостность вида. Это создает широкое поле для деятельности естественного отбора. Строгое самоопыление встречается относительно редко (например, у гороха) и может вести к расщеплению вида на ряд чистых линий, то есть делает популяции гомозиготными. Иногда это приводит к затуханию микроэволюции. Но самоопыление способствует изоляции вновь возникших в результате мутаций форм, обособляя и фиксируя их в чистых линиях. По-видимому, для эволюционного процесса оптимально сочетание самоопыления и перекрестного опыления, что чаще всего и имеет место в природе. Однако преобладает перекрестное опыление и поэтому у цветковых обычно есть специальные приспособления морфологического и физиологического характера, предотвращающие или ограничивающие самоопыление. К ним относятся двудомность, дихогамия, гетеростилия, самонесовместимость. Наиболее надежное средство предотвращения самоопыления – двудомность, но при этом часть особей популяций (мужские особи) не дает семян. Однодомность устраняет автогамию, но не предохраняет от генетически равноценной ей гейтоногамии. Функциональная раздельнополость получила название дихогамии (от греческого «дихе» – на две части). Она проявляется в разновременном созревании пыльцы и рылец в цветках одного и того же растения. Дихогамия чаще всего встречается в форме протандрии. В этом случае раньше вызревает пыльца. При протогинии (другой форме дихогамии) происходит более раннее созревание рылец и завязей, в которых заключены семязачатки.
Иногда бывает полная физиологическая самонесовместимость. Она выражается в подавлении при самоопылении прорастания пыльцы на рыльце пестика той же особи. Самонесовместимость встречается у покрытосеменных более широко, чем двудомность. Она зарегистрирована более чем у 10000 видов цветковых.
Явление гетеростилии состоит в том, что у некоторых видов имеются две или даже три формы цветков (находящиеся на разных особях), различающиеся по длине столбиков и тычиночных нитей. В силу этого самоопыление у этих растений в значительной мере затруднено и дает обычно ничтожное количество семян. Классическими примерами растений, характеризующихся гетеростилией, служат виды первоцветов (Primula) и болотное растение дербенник иволистный (Lythrum salicaria).
Механизмы перекрестного опыления весьма разнообразны и являются главным объектом изучения особого раздела ботаники, называемого антэкологией. Их подразделяют на два основных типа – биотическое и абиотическое опыление. Биотическое опыление осуществляется животными, абиотическое – с помощью неживых факторов внешней среды. Наибольшее значение среди механизмов биотического опыления имеет энтомофилия (от греческого «энтомон» – насекомое, «филео» – любить). Насекомые сыграли выдающуюся роль в эволюции цветка. Цветки привлекают насекомых запасом пищи: пыльцой, нектаром. Для привлечения опылителей служат яркая окраска и характерный запах цветков; некоторое значение имеет их форма. Конкретные механизмы перекрестного опыления, осуществляемого насекомыми, очень разнообразны и связаны как со строением цветка, так и с особенностями тела насекомого. Видам некоторых семейств (бобовые, губоцветные, орхидные) свойственны особые способы опыления.
Помимо насекомых существенную роль в биотическом опылении, особенно в тропиках, играют птицы (орнитофилия), летучие мыши и некоторые нелетающие млекопитающие (например, крысы). Абиотическое опыление связано с переносом пыльцы ветром (анемофилия, от греческого «анемос» – ветер), реже (у болотных и водных растений) – водой. Анемофилы – это преимущественно растения открытых пространств. У анемофильных цветковых растений, как правило, невзрачный, сильно редуцированный околоцветник и крупные, часто мохнатые рыльца с огромной воспринимающей пыльцу поверхностью. Очень часто мелкие цветки анемофилов собраны в плотные или многоцветковые соцветия, пыльца их обильна, легка, а экзина почти лишена скульптурных утолщений, обычных для пыльцы энтомофильных растений.
Второй тип спор разноспоровых цветковых растений – мегаспоры образуются в процессе мегаспорогенеза, протекающего в семязачатке. Центральная часть семязачатка – нуцеллус, представляет собой мегаспорангий цветковых растений. Семязачаток состоит из диплоидных клеток, поскольку он является частью растения-спорофита. Одна из клеток нуцеллуса является материнской клеткой мегаспор. Она редукционно делится и формирует тетраду линейно расположенных гаплоидных клеток. (рис. 13. 14).
Рис. 13.14. Стадии развития мегаспор в семязачатке лобелии (Lobelia cardinalis)
Это и есть мегаспоры цветковых растений. Обычно три из четырех клеток-мегаспор в семязачатке отмирают, единственная жизнеспособная мегаспора развивается в женский гаметофит, или зародышевый мешок.
Формирование зародышевого мешка, получившее название мегагаметогенез, происходит путем трех последовательных делений ядра мегаспоры. В результате первого деления образуется двухъядерная клетка, которая сильно увеличивается в длину. Далее каждое из двух ядер делится еще дважды и возникает восьмиядерная клетка, имеющая по четыре ядра у каждого полюса. На этом деления завершаются. В центр клетки от каждого их полюсов перетекает по одному гаплоидному ядру и они сливаются в диплоидное вторичное ядро. В результате цитокинеза остальные ядра (по три у каждого полюса) формируют дочерние гаплоидные клетки. В итоге возникает семиклеточный зародышевый мешок. На микропилярном его конце обычно имеется гаплоидная яйцеклетка в окружении двух гаплоидных клеток-синергид. На халазальном конце расположены три гаплоидные клетки-антиподы. Основной объем зародышевого мешка составляет центральная диплоидная клетка (см. рис. 13.10).
Для осуществления оплодотворения необходимы два условия: зрелая жизнеспособная пыльца, попавшая на рыльце пестика, и сформировавшийся зародышевый мешок в семязачатке. Оплодотворению предшествует прорастание пыльцевого зерна. Оно начинается с разбухания зерна и выхода из апертуры пыльцевой трубки. В густой цитоплазме кончика пыльцевой трубки идут интенсивные физиологические процессы, вследствие которых несколько размягчаются ткани рыльца и столбика, в которые внедряется пыльцевая трубка. По мере роста пыльцевой трубки в нее переходят ядро вегетативной клетки и оба спермия. В огромном большинстве случаев пыльцевая трубка проникает в мегаспорангий (нуцеллус) через микропиле семязачатка, реже – иным образом. Проникнув в зародышевый мешок, пыльцевая трубка разрывается (под действием разницы осмотического давления в ней и давления в нуцеллусе) и ее содержимое изливается в зародышевый мешок. Один из спермиев сливается с яйцеклеткой и образуется диплоидная зигота, дающая затем начало зародышу. Второй спермий сливается со вторичным ядром, располагающимся в центре зародышевого мешка, что приводит к образованию триплоидного ядра, развивающегося затем в специальную питательную ткань – эндосперм (от греческого «эндон» – внутри, «сперма» – семя). Весь этот процесс получил название двойного оплодотворения. Он впервые описан в 1898 г. выдающимся русским цитологом и эмбриологом С.Г.Навашиным. Прочие клетки зародышевого мешка (антиподы и синергиды) разрушаются при проникновении пыльцевой трубки. Однако при гибели яйцеклетки они могут выполнить ее функцию.
Биологический смысл двойного оплодотворения, в отличие от голосеменных, у которых гаплоидный эндосперм развивается независимо от процесса оплодотворения, состоит в том, что у покрытосеменных триплоидный эндосперм образуется лишь в случае оплодотворения. С учетом гигантского числа семян этим достигается существенная экономия энергетических и пластических ресурсов.
У многих цветковых (около 10% видов) в процессе эволюции половое размножение замещается различными формами бесполого. Из них наиболее известен апомиксис. Семена у апомиктичных растений образуются без оплодотворения. Апомиксис исключает генетическое расщепление, поэтому апомиктические формы образуют клоны, в пределах которых все особи имеют одинаковую генетическую и соматическую конституцию. Хорошим примером растения с апомиктическим образованием семян служит одуванчик с его поразительно высокой жизнеспособностью. Нередко у многолетних корневищных растений преобладает вегетативное размножение, а семенное ограничено.
studfiles.net
Репродуктивные процессы в цветке
Философия Репродуктивные процессы в цветке
просмотров - 104
В цветке последовательно протекают важнейшие процессы, обеспечивающие вначале бесполое размножение и образование очередного поколения гаметофитов, а затем и половое размножение, завершающееся образованием нового поколения спорофитов в виде зародышей, расположенных в семенах. Цветковые являются разноспоровыми растениями. Οʜᴎ образуют два типа спор: микро- и мегаспоры.
Клетки бесполого размножения – микроспоры, образуются в ходе процесса, называемого микроспорогенезом в микроспорангиях, которыми являются гнезда пыльника. В результате ряда последовательных митотических делений клеток археспория, то есть образовательной ткани пыльника, возникают материнские клетки микроспор (микроспороциты), содержащие, как и все растение, диплоидный набор хромосом. Далее каждая материнская клетка в результате мейотического деления образует тетраду гаплоидных микроспор. Сформировавшаяся микроспора имеет оболочку и единственное ядро.
В ходе превращения микроспоры в пыльцевое зерно происходит два митотических деления и под защитой оболочки микроспоры возникают сначала две, далее три клетки – редуцированный мужской гаметофит. После формирования такого редуцированного гаметофита трехклеточная структура принято называть пыльцевым зерном (мужской особью). Пыльцевое зерно в начале развития содержит две клетки: маленькую генеративную и более крупную, часто называемую вегетативной. Микрогаметогенез крайне упрощен. Генеративная клетка делится однократно и из нее образуются две безжгутиковые мужские гаплоидные гаметы – спермии. В таком состоянии пыльцевое зерно готово к половому размножению. При попадании на рыльце пестика цветка вегетативная клетка пыльцевого зерна удлиняется и становится так называемой «пыльцевой трубкой», с мощью которой уже в семязачатке завершается оплодотворение (рис. 13.13).
Рис. 13.13. Микроспора, пыльцевое зерно и его прорастание . А – микроспора; Б – пыльцевое зерно; В – формирование пыльцевой трубки; Г– часть пыльцевой трубки: 1 – экзина, 2 – интина, 3 – вегетативная клетка, дающая начало пыльцевой трубке, 4 – генеративная клетка, 5 – спермий.
Зрелое пыльцевое зерно окружено сложно устроенной оболочкой. Оболочка, часто называемая спородермой, состоит из двух главных слоев: внешнего, более толстого – экзины, и относительно тонкого внутреннего – интины. Экзина, содержащая особое высокомолекулярное вещество спорополленин, характеризуется необычайной стойкостью: она не растворяется в кислотах и щелочах, выдерживает температуру до 300 °С и сохраняется в фоссилизованном (окаменевшем) состоянии миллионы лет в геологических отложениях. Это позволяет вести спорово-пыльцевой анализ почв, донных отложений и осадочных толщ, определять, какие виды росли в разные геологические периоды. Экзина микроспор имеет на поверхности разнообразные скульптурные утолщения.
Особенности скульптуры важны при морфологической характеристике пыльцевых зерен отдельных таксонов. Наиболее развиты такие утолщения у энтомофильных растений. Интина менее стойка, чем экзина. Она состоит из целлюлозы и пектина и как бы облегает содержимое пыльцевого зерна.
Перенос пыльцевых зерен из тычинок на рыльца пестиков принято называть опылением. Для того чтобы образовался зародыш семени, должны произойти опыление и оплодотворение. Опыление впервые появляется у голосеменных, однако наибольшее разнообразие механизмов опыления и его совершенство достигаются у цветковых. Различают два типа опыления – самоопыление, или автогамию (от греческого «аутос» – сам), и перекрестное опыление, или ксеногамию (от греческого «ксенос» – чужой, «гамос» – брак). При самоопылении рыльце опыляется пыльцой того же цветка или пыльцой других цветков этой особи растения – гейтоногамия (девственное опыление). Обычно самоопыление осуществляется в распустившихся цветках, но иногда происходит в цветках закрытых, нераспустившихся (клейстогамных). В генетическом отношении все эти способы вполне равноценны.
В случае если перенос пыльцы осуществляется между цветками разных особей, то происходит перекрестное опыление. Оно свойственно не менее 90% видов цветковых растений. Перекрестное опыление обеспечивает обмен генами, поддерживает высокий уровень гетерозиготности популяций, определяет единство и целостность вида. Это создает широкое поле для деятельности естественного отбора. Строгое самоопыление встречается относительно редко (к примеру, у гороха) и может вести к расщеплению вида на ряд чистых линий, то есть делает популяции гомозиготными. Иногда это приводит к затуханию микроэволюции. Но самоопыление способствует изоляции вновь возникших в результате мутаций форм, обособляя и фиксируя их в чистых линиях. По-видимому, для эволюционного процесса оптимально сочетание самоопыления и перекрестного опыления, что чаще всего и имеет место в природе. При этом преобладает перекрестное опыление и в связи с этим у цветковых обычно есть специальные приспособления морфологического и физиологического характера, предотвращающие или ограничивающие самоопыление. К ним относятся двудомность, дихогамия, гетеростилия, самонесовместимость. Наиболее надежное средство предотвращения самоопыления – двудомность, но при этом часть особей популяций (мужские особи) не дает семян. Однодомность устраняет автогамию, но не предохраняет от генетически равноценной ей гейтоногамии. Функциональная раздельнополость получила название дихогамии (от греческого «дихе» – на две части). Она проявляется в разновременном созревании пыльцы и рылец в цветках одного и того же растения. Дихогамия чаще всего встречается в форме протандрии. В этом случае раньше вызревает пыльца. При протогинии (другой форме дихогамии) происходит более раннее созревание рылец и завязей, в которых заключены семязачатки.
Иногда бывает полная физиологическая самонесовместимость. Она выражается в подавлении при самоопылении прорастания пыльцы на рыльце пестика той же особи. Самонесовместимость встречается у покрытосеменных более широко, чем двудомность. Она зарегистрирована более чем у 10000 видов цветковых.
Явление гетеростилии состоит в том, что у некоторых видов имеются две или даже три формы цветков (находящиеся на разных особях), различающиеся по длине столбиков и тычиночных нитей. В силу этого самоопыление у этих растений в значительной мере затруднено и дает обычно ничтожное количество семян. Классическими примерами растений, характеризующихся гетеростилией, служат виды первоцветов (Primula) и болотное растение дербенник иволистный (Lythrum salicaria).
Механизмы перекрестного опыления весьма разнообразны и являются главным объектом изучения особого раздела ботаники, называемого антэкологией. Их подразделяют на два базовых типа – биотическое и абиотическое опыление. Биотическое опыление осуществляется животными, абиотическое – с помощью неживых факторов внешней среды. Наибольшее значение среди механизмов биотического опыления имеет энтомофилия (от греческого «энтомон» – насекомое, «филео» – любить). Насекомые сыграли выдающуюся роль в эволюции цветка. Цветки привлекают насекомых запасом пищи: пыльцой, нектаром. Для привлечения опылителей служат яркая окраска и характерный запах цветков; неĸᴏᴛᴏᴩᴏᴇ значение имеет их форма. Конкретные механизмы перекрестного опыления, осуществляемого насекомыми, очень разнообразны и связаны как со строением цветка, так и с особенностями тела насекомого. Видам некоторых семейств (бобовые, губоцветные, орхидные) свойственны особые способы опыления.
Помимо насекомых существенную роль в биотическом опылении, особенно в тропиках, играют птицы (орнитофилия), летучие мыши и некоторые нелетающие млекопитающие (к примеру, крысы). Абиотическое опыление связано с переносом пыльцы ветром (анемофилия, от греческого «анемос» – ветер), реже (у болотных и водных растений) – водой. Анемофилы - ϶ᴛᴏ преимущественно растения открытых пространств. У анемофильных цветковых растений, как правило, невзрачный, сильно редуцированный околоцветник и крупные, часто мохнатые рыльца с огромной воспринимающей пыльцу поверхностью. Очень часто мелкие цветки анемофилов собраны в плотные или многоцветковые соцветия, пыльца их обильна, легка, а экзина почти лишена скульптурных утолщений, обычных для пыльцы энтомофильных растений.
Второй тип спор разноспоровых цветковых растений – мегаспоры образуются в процессе мегаспорогенеза, протекающего в семязачатке. Центральная часть семязачатка – нуцеллус, представляет собой мегаспорангий цветковых растений. Семязачаток состоит из диплоидных клеток, поскольку он является частью растения-спорофита. Одна из клеток нуцеллуса является материнской клеткой мегаспор. Она редукционно делится и формирует тетраду линейно расположенных гаплоидных клеток. (рис. 13. 14).
Рис. 13.14. Стадии развития мегаспор в семязачатке лобелии(Lobelia cardinalis)
Это и есть мегаспоры цветковых растений. Обычно три из четырех клеток-мегаспор в семязачатке отмирают, единственная жизнеспособная мегаспора развивается в женский гаметофит, или зародышевый мешок.
Формирование зародышевого мешка, получившее название мегагаметогенез, происходит путем трех последовательных делений ядра мегаспоры. В результате первого деления образуется двухъядерная клетка, которая сильно увеличивается в длину. Далее каждое из двух ядер делится еще дважды и возникает восьмиядерная клетка, имеющая по четыре ядра у каждого полюса. На этом деления завершаются. В центр клетки от каждого их полюсов перетекает по одному гаплоидному ядру и они сливаются в диплоидное вторичное ядро. В результате цитокинеза остальные ядра (по три у каждого полюса) формируют дочерние гаплоидные клетки. В итоге возникает семиклеточный зародышевый мешок. На микропилярном его конце обычно имеется гаплоидная яйцеклетка в окружении двух гаплоидных клеток-синергид. На халазальном конце расположены три гаплоидные клетки-антиподы. Основной объем зародышевого мешка составляет центральная диплоидная клетка (см. рис. 13.10).
Для осуществления оплодотворения необходимы два условия: зрелая жизнеспособная пыльца, попавшая на рыльце пестика, и сформировавшийся зародышевый мешок в семязачатке. Оплодотворению предшествует прорастание пыльцевого зерна. Оно начинается с разбухания зерна и выхода из апертуры пыльцевой трубки. В густой цитоплазме кончика пыльцевой трубки идут интенсивные физиологические процессы, вследствие которых несколько размягчаются ткани рыльца и столбика, в которые внедряется пыльцевая трубка. По мере роста пыльцевой трубки в нее переходят ядро вегетативной клетки и оба спермия. В огромном большинстве случаев пыльцевая трубка проникает в мегаспорангий (нуцеллус) через микропиле семязачатка, реже – иным образом. Проникнув в зародышевый мешок, пыльцевая трубка разрывается (под действием разницы осмотического давления в ней и давления в нуцеллусе) и ее содержимое изливается в зародышевый мешок. Один из спермиев сливается с яйцеклеткой и образуется диплоидная зигота͵ дающая затем начало зародышу. Второй спермий сливается со вторичным ядром, располагающимся в центре зародышевого мешка, что приводит к образованию триплоидного ядра, развивающегося затем в специальную питательную ткань – эндосперм (от греческого «эндон» – внутри, «сперма» – семя). Весь данный процесс получил название двойного оплодотворения. Он впервые описан в 1898 ᴦ. выдающимся русским цитологом и эмбриологом С.Г.Навашиным. Прочие клетки зародышевого мешка (антиподы и синергиды) разрушаются при проникновении пыльцевой трубки. При этом при гибели яйцеклетки они могут выполнить ее функцию.
Биологический смысл двойного оплодотворения, в отличие от голосеменных, у которых гаплоидный эндосперм развивается независимо от процесса оплодотворения, состоит в том, что у покрытосеменных триплоидный эндосперм образуется лишь в случае оплодотворения. С учетом гигантского числа семян этим достигается существенная экономия энергетических и пластических ресурсов.
У многих цветковых (около 10% видов) в процессе эволюции половое размножение замещается различными формами бесполого. Из них наиболее известен апомиксис. Семена у апомиктичных растений образуются без оплодотворения. Апомиксис исключает генетическое расщепление, в связи с этим апомиктические формы образуют клоны, в пределах которых все особи имеют одинаковую генетическую и соматическую конституцию. Хорошим примером растения с апомиктическим образованием семян служит одуванчик с его поразительно высокой жизнеспособностью. Нередко у многолетних корневищных растений преобладает вегетативное размножение, а семенное ограничено.
Читайте также
В цветке последовательно протекают важнейшие процессы, обеспечивающие вначале бесполое размножение и образование очередного поколения гаметофитов, а затем и половое размножение, завершающееся образованием нового поколения спорофитов в виде зародышей, расположенных в... [читать подробенее]
В цветке последовательно протекают важнейшие процессы, обеспечивающие вначале бесполое размножение и образование очередного поколения гаметофитов, а затем и половое размножение, завершающееся образованием нового поколения спорофитов в виде зародышей, расположенных в... [читать подробенее]
oplib.ru









