17 Передвиж. Воды по раст. Концевые двигатели водного тока.. Верхний концевой двигатель растений
17 Передвиж. Воды по раст. Концевые двигатели водного тока..
Т. к. вода поглощается в осн. корневой системой, возникает необходимость ее передвижения по растению. Этот процесс составляет т. н. восходящий ток. Следует отметить, что это название отражает характер передвижения и его локализацию в растении. Он проходит по мертвым тканям стебля или черешка — сосудам у покрытосеменных и трахеидам у голосеменных. Однако эта локализация не абсолютна: вода способна перемещаться и по флоэмной системе. Передвижение по сосудам идет с большей скоростью (20 см/ч), чем по трахеидам (только 5 см/ч).
Если принять во внимание всю протяженность пути восходящего тока, то его можно будет разбить на 2 не равных по длине участка: Мертвые гистологические элементы в середине проводящего пути — сосуды или трахеиды. Протяженность значительна, но вода проходит по нему сравнительно легко, так как двигается пассивно по мертвым элементам, не испытывая с их стороны значительного сопротивления. Живые клетки корня и листа, находящиеся в начале и в конце пути передвижения. Этот путь пространственно короток, но преодолевается он с большим трудом, так как клеточные мембраны препятствуют передвижению воды. Этот ток снабжает водой все органы и ткани, приводя их в состояние тургора. Восходящий ток воды захватывает мин. ионы, поглощенные корнем, переносит их и тем самым облегчает распределение (но не поглощение!) по растению.
Концевые двигатели водного тока
Для того чтобы вода двигалась по растению (и не просто двигалась, а поднималась вверх), необходимы затраты энергии, точки приложения которой располагаются по концам тока, вследствие чего они получили название концевых двигателей.
Нижний концевой двигатель, или корневое давление. Его роль проявляется в основном при активном поглощении — нагнетании воды. При участии актомиозиноподобных сократительных белков он не только подает воду в корневую систему, но и проталкивает ее дальше в сосуды корня и вверх по стеблю. Нагнетание воды — активный энергозависимый процесс, который сильнее всего выражен в коре корня. Сила, развиваемая концевым двигателем, невелика, она может обеспечить подъем воды на высоту не более 1 м., т. е. достаточна для травянистых растений и небольших кустарников. Верхний концевой двигатель, или присасывающая сила транспирации. При постоянном испарении воды в листьях растений разбивается сосущая сила, отсасывающая воду от ближайших клеток и передающаяся др. кл., по к-ым передвигается вода, вплоть до сосудов. В сосудах нет цитоплазмы, поэтому нет и осмотического давления, и всасывание жидкости происходит при участии всей величины сосущей силы. Она позволяет поднять воду на несколько метров. Этой силы дост. для обеспечения водой кустарников и небольших деревьев.
Поднятие воды по стволу
Многие древесные раст. имеют большую протяженность ствола. У таких растений на помощь приходят силы сцепления между молекулами воды, которые очень велики и могут достигать 30 — 35 МПа. Этой силы достаточно для того, чтобы поднять воду на 1 — 2 км, что значительно превышает высоту любого дерева. Силы сцепления молекул воды действуют только при определенных условиях: водные струи в сосудах должны идти непрерывно, без пузырьков воздуха. Если же в них попадает в-х, передвижение воды прерывается. Непрерывность водных струй в сосудах достигается их сильным натяжением и плотным прилеганием к стенкам сосудов. Водные струи оказываются как бы висящими, они удерживаются в подвешенном состоянии сосущей силой трансп. и снизу поддерживаются силой корневого Р. При сильном натяжении водных струй пр-ит натяжение и самих сосудов, при этом у последних наблюдается увелич. длины и уменьшение поперечного сечения (подобно эластичной резине). Это создает предпосылки к смыканию стенок сосудов, что привело бы к их закрыванию и прекращению водного тока. Однако этого не пр-ит, т. к. сосуды имеют внутренние утолщения стенок — кольчатые, спиральные, лестничные, к-ые наподобие каркаса растягивают их и предохр. от смыкания.
Величина его незначительна — всего 20 см воды на 1 см2 площади сосудов. Однако и этого достаточно при условии не очень сильного напряжения атмосферных факторов.
studfiles.net
| Основной силой, вызывающей поступление и передвижение воды в растении, является процесс транспирации, в результате которого возникает градиент водного потенциала. Как уже упоминалось, водный потенциал — это мера энергии, используемой водой для передвижения. Чем меньше насыщенность водой данной системы, тем меньше (более отрицателен) ее водный потенциал. При потере воды растением в процессе транспирации создается ненасыщенность клеток листа водой, как следствие, водный потенциал становится более отрицательным. Поступление воды идет в сторону более отрицательного водного потенциала. Так называемый верхний концевой двигатель водного тока в растении — это транспирация листьев. Однако опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется. Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлений, как гуттация и плач. Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, препятствующей испарению, выделяют капельно-жидкую воду с небольшим количеством растворенных веществ — гуттируют. Выделение жидкости идет через специальные водные устьица — гидатоды. Выделяющаяся жидкость — гутта. Таким образом, процесс гуттации является результатом одностороннего тока воды, происходящего в отсутствие транспирации, и, следовательно, вызывается какой-то иной причиной. К такому же выводу можно прийти и при рассмотрении явления плача растений. Если срезать побеги растения и к срезанному концу присоединить стеклянную трубку, то по ней будет подниматься жидкость. Анализ показывает, что это — вода с растворенными веществами, получившая название пасоки. В некоторых случаях, особенно в весенний период, плач наблюдается и при надрезе веток растений. Именно на этом основано вытекание сока при надрезе ствола березы весной. Определения показали, что объем выделяющейся жидкости (пасоки) во много раз превышает объем корневой системы. Таким образом, плач — это не просто вытекание жидкости в результате пореза. Это приводит к выводу, что плач, как и гуттация, связан с наличием одностороннего тока воды через корневые системы, не зависящего от транспирации. Силу, вызывающую в растении односторонний ток воды с растворенными веществами, не зависящую от процесса транспирации, называют корневым давлением. Наличие корневого давления позволяет говорить о нижнем концевом двигателе водного тока. Корневое давление можно измерить, присоединив манометр к концу, оставшемуся после срезания надземных органов растения, или поместив корневую систему в серию растворов различной концентрации и подобрав такую, при которой плач прекращается. Оказалось, что величина корневого давления равна примерно 1—1,5 бара (ДА. Сабинин). Было показано также, что плач осуществляется только в тех условиях, в которых нормально протекают все процессы жизнедеятельности клеток корня. Не только умерщвление клеток корня, но и снижение интенсивности их жизнедеятельности, в первую очередь интенсивности дыхания, прекращает плач. В отсутствие кислорода, под влиянием дыхательных ядов, при пониженной температуре плач приостанавливается. Все сказанное позволило Д. А. Сабинину дать следующее определение: плач растений — это прижизненный односторонний ток воды и питательных веществ, связанный с затратой энергии. Однако механизм этого явления до настоящего времени не получил достаточного объяснения. Существуют две основные точки зрения. Ряд исследователей (А. Крафтс, Т. Бройер и др.) полагают, что вода передвигается пассивно, осмотическим путем по градиенту водного потенциала. Согласно этой гипотезе низкий (более отрицательный) водный потенциал создается в сосудах ксилемы благодаря поступлению туда из почвы растворимых солей, а также из-за слабого противодавления малоэластичных стенок сосудов. Соли поступают активно, для чего и требуется затрата энергии. Таким образом, поступление воды связано в этом случае с энергетическими затратами и с активной жизнедеятельностью клеток корня не прямо, а косвенно. В этой связи корневое давление можно определить как давление, развивающееся в проводящих элементах ксилемы благодаря метаболической активности корней. Поскольку активное поступление зависит от энергии, выделяемой при дыхании, работа нижнего концевого двигателя — корневого давления зависит от присутствия кислорода, ингибиторов и активаторов дыхательного обмена. Вместе с тем существует и другая точка зрения (В.Н. Жолкевич, Л.В. Можаева, Н.В. Пильщиков), согласно которой корневое давление складывается из двух составляющих: осмотической и метаболической. Работа последней требует непосредственной затраты энергии АТФ. Высказывается предположение, что при этом большая роль принадлежит сократительным актиноподобным белкам, энергозависимое сокращение и расслабление которых вызывает изменения гидростатического давления в клетках. В результате на пути водного тока в направлении сосудов ксилемы создаются локальные градиенты водного потенциала, что и способствует проталкиванию воды в сосуды. Вопрос этот требует дальнейшей экспериментальной разработки. Сила, развиваемая нижним концевым двигателем водного тока, во много раз меньше той, которая обусловливается транспирацией. Это проявляется и в том, что скорость вытекания воды из ксилемы значительно уступает скорости транспирации, поэтому в обычных условиях вода в сосудах находится не под положительным, а под отрицательным давлением. Однако все же нижний концевой двигатель способствует подаче воды. Большинство исследователей считает, что прекращение работы нижнего концевого двигателя является показателем начала страдания растений от недостатка воды (засухи). Особенное значение этот двигатель водного тока приобретает в отсутствие транспирации. Так, это может иметь место ранней весной, когда воздух насыщен водой и транспирация ослаблена. У некоторых многолетних растений в зимний период сосуды ксилемы заполнены воздухом и весной поступление воды идет за счет работы только нижнего концевого двигателя. |
fizrast.ru
Основные двигатели водного тока
Поглощение воды корневой системой идет благодаря работе двух концевых двигателей водного тока: верхнего концевого двигателя, или присасывающей силы испарения (транспирации), и нижнего концевого двигателя, или корневого давления. Основной силой, вызывающей поступление и передвижение воды в растении, является присасывающая сила транспирации в результате которой возникает градиент водного потенциала. Водный потенциал — это мера энергии, используемой водой для передвижения. Водный потенциал и сосущая сила одинаковы по абсолютному значению, но противоположны по знаку. Чем меньше насыщенность водой данной системы, тем меньше (более отрицателен) ее водный потенциал. При потере воды растением в процессе транспирации создается ненасыщенность клеток листа водой, как следствие, возникает сосущая сила (водный потенциал падает). Поступление воды идет в сторону большей сосущей силы, или меньшего водного потенциала.
Таким образом, верхний концевой двигатель водного тока в растении — это присасывающая сила транспирации листьев, и его работа мало связана с жизнедеятельностью корневой системы. Действительно, опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется. Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлений, как гуттация.
Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, препятствующей испарению, выделяют капельно-жидкую воду с небольшим количеством растворенных веществ — гуттация. Выделение жидкости идет через специальные водные устьица — гидатоды. Выделяющаяся жидкость — гутта. Таким образом, процесс гуттации является результатом одностороннего тока воды, происходящего в отсутствие транспирации, и, следовательно, вызывается какой-то иной причиной. К такому же выводу можно прийти и при рассмотрении явления плач растений. Если срезать побеги растения и к срезанному концу присоединить стеклянную трубку, то по ней будет подниматься жидкость. Анализ показывает, что это вода с растворенными веществами — пасока. В некоторых случаях, особенно в весенний период, плач наблюдается и при надрезе веток растений. Определения показали, что объем выделяющейся жидкости (пасоки) во много раз превышает объем корневой системы. Таким образом, плач — это не просто вытекание жидкости в результате пореза. Все сказанное приводит к выводу, что плач, как и гуттация, связан с наличием одностороннего тока воды через корневые системы, не зависящего от транспирации. Силу, вызывающую односторонний ток воды по сосудам с растворенными веществами, не зависящую от процесса транспирации, называют корневым давлением. Наличие корневого давления позволяет говорить о нижнем концевом двигателе водного тока. Корневое давление можно измерить, присоединив манометр к концу, оставшемуся после срезания надземных органов растения, или поместив корневую систему в серию растворов различной концентрации и подобрав такую, при которой плач прекращается. Оказалось, что величина корневого давления равна примерно 0,1—0,15 МПа. Концентрация наружного раствора, останавливающего плач, значительно выше концентрации пасоки. Это позволило высказать мнение, что плач может идти против градиента концентрации. Было показано также, что плач осуществляется только в тех условиях, в которых нормально протекают все процессы жизнедеятельности клеток. Не только умерщвление клеток корня, но и снижение интенсивности их жизнедеятельности, в первую очередь интенсивности дыхания, прекращает плач. В отсутствие кислорода, под влиянием дыхательных ядов, при пониженной температуре плач приостанавливается. Таким образом - плач растений — это прижизненный односторонний ток воды и питательных веществ, зависящий от аэробной переработки ассимилятов. Односторонний ток воды в корне. Объясняется тем, что клетки корня поляризованы в определенном направлении. Это проявляется в том, что в разных отсеках одной и той же клетки процессы обмена веществ различны. В одной части клетки идут усиленные процессы распада, в частности, крахмала на сахара, вследствие чего концентрация клеточного сока возрастает. Следовательно, если со стороны, где клетка обращена к внешней среде, она насыщена водой, то, с другой стороны, наблюдается секреция воды, тургорное давление, которое превышает осмотическое, и клетка будет выделять воду. Надо учитывать, что весь этот механизм будет работать только при достаточном количестве воды в среде и при ненарушенном обмене веществ.
Согласно другой гипотезе, зависимость плача растений от интенсивности дыхания является косвенной. Энергия дыхания используется для поступления ионов в клетки коры, откуда они десорбируются в сосуды ксилемы. В результате концентрация солей в сосудах ксилемы повышается, что и вызывает поступление воды
studfiles.net
1.2 Основные двигатели водного тока. Водный режим у растений
Похожие главы из других работ:
Биохимия растений. Процессы брожения. Фотосинтез
3. Поступление воды в растение. Верхние и нижние «двигатели» водного потока
теневыносливый растение орошение микрофлора Основным источником влаги является вода, находящаяся в почве, и основным органом поглощения воды в растение-- корневая система. Роль этого органа, прежде всего, заключается в том...
Водний обмін у рослин
Розділ 1. Загальна характеристика водного обміну рослинного організму
Вода відіграє важливу роль у життєдіяльності організму завдяки своїм унікальним фізичним і хімічним властивостям. Молекула води складається з двох атомів водню, приєднаних до одного атому кисню. Атом кисню відтягує електрони від водню...
Водный обмен растений
1.2 Основные двигатели водного тока
Поглощение воды корневой системой идет благодаря работе двух концевых двигателей водного тока: верхнего концевого двигателя, или присасывающей силы испарения (транспирации), и нижнего концевого двигателя, или корневого двигателя...
Водный режим растений
1. Общая характеристика водного обмена растительного организма
Водный режим растений, водообмен, поступление воды в растение и отдача её растением, необходимые для его жизнедеятельности (обмена веществ, роста, развития, размножения)...
Закономерности линейно-весового роста сиговых рыб Обь-Иртышского бассейна
1. Основные понятия
Абсолютная длина тела (общая длина или длина всей рыбы) - от вершины рыла до вертикали конца наиболее длинной лопасти хвостового плавника при горизонтальном положении рыбы. Принимается при указании предельных размеров рыбы...
Кровоносна система
2.4 Вікові та видові особливості стійкості та чутливості риб до токсикантів водного середовища
Дослідження ролі вікового чинника у визначенні стійкості риб до отрут важливе для розуміння шляхів впливу токсикантів у природній водоймі на популяцію виду в цілому та на окремі стадії життєвого циклу...
Микоплазмы: представители, особенности строения и метаболизма, систематика
2. Основные представители
Представители группы микоплазм (роды Mycoplasma, Acholeplasma и Spiroplasma) - паразитические бактерии. Возбудитель относится к разделу Tenericutes. Согласно современной классификации, они относятся к семейству Мycoplasmataceae. Это семейство разделяют на 2 рода - род Мycoplasma...
Морфология внутреннего строения рыб
3. Основные представители
Основная масса промысловых рыб относится к высшим костным рыбам. Подкласс лучеперых включает более 95 % всех видов современных рыб (треска, окунь, сельдь и др.)- Скелет костных рыб имеет костную ткань...
Особенности строения клетки
3. Транспирация и ее биологическое значение. Особенности верхнего "двигателя" водного потока
В основе расходования воды растительным организмом лежит процесс испарения - переход воды из жидкого в парообразное состояние, происходящий при соприкосновении органов растения с ненасыщенной водой атмосферой...
Пристосування тварин до життя у водному середовищі
РОЗДІЛ 1. Загальна характеристика водного середовища
Водне середовище має свої певні властивості і тому організми-гідробіонти мають різноманітні специфічні пристосування до життя у воді. Водне середовище менш мінливе, має відносно постійні показники...
Процесс транспирации у разных сортов рода Cucurbita
Глава III. ОБЩАЯ ХАРАКТЕРИСТИКА ВОДНОГО БАЛАНСА РАСТЕНИЙ
Вода играет огромную роль в жизни растения. Прямо или косвенно она участвует во всех жизненных процессах, протекающих в клетке. Вода является элементом структуры мембран и цитоплазмы...
Свободные аминокислоты нервной системы
10. Основные аминокислоты
Лизин пока мало исследован в аспекте его значения для нервной системы. Пути деградации лизина в мозге точно не установлены, но они отличаются от локализованных в печени. Лизин в мозге может катаболировать через образование пипеколовой кислоты...
Ткани и их функции в растительных организмах
5. Основные ткани
...
Токсикологія риб
4. Вікові та видові особливості стійкості та чутливості риб до токсикантів водного середовища
Дослідження ролі вікового чинника у визначенні стійкості риб до отрут важливе для розуміння шляхів впливу токсикантів у природній водоймі на популяцію виду в цілому та на окремі стадії життєвого циклу...
Фитобентос континентальных водоемов
1.3 Основные представители
Водяная сеточка (Hydrodictyon reticulatum) Род пресноводных зелёных водорослей порядка протококковых. В СССР -- один вид, Н. reticulatum. Слоевище до 1 м длиной, имеет вид свободноплавающей мешковидной сетки. Рис...
bio.bobrodobro.ru
Механизм передвижеия воды из корня в надземные органы. Верхний и нижний концевые двигатели.
Т. к. вода поглощается в осн. корневой системой, возникает необходимость ее передвижения по растению. Этот процесс составляет т. н. восходящий ток. Следует отметить, что это название отражает характер передвижения и его локализацию в растении. Он проходит по мертвым тканям стебля или черешка — сосудам у покрытосеменных и трахеидам у голосеменных. Однако эта локализация не абсолютна: вода способна перемещаться и по флоэмной системе. Передвижение по сосудам идет с большей скоростью (20 см/ч), чем по трахеидам (только 5 см/ч).Если принять во внимание всю протяженность пути восходящего тока, то его можно будет разбить на 2 не равных по длине участка: Мертвые гистологические элементы в середине проводящего пути — сосуды или трахеиды. Протяженность значительна, но вода проходит по нему сравнительно легко, так как двигается пассивно по мертвым элементам, не испытывая с их стороны значительного сопротивления. Живые клетки корня и листа, находящиеся в начале и в конце пути передвижения. Этот путь пространственно короток, но преодолевается он с большим трудом, так как клеточные мембраны препятствуют передвижению воды. Этот ток снабжает водой все органы и ткани, приводя их в состояние тургора. Восходящий ток воды захватывает мин. ионы, поглощенные корнем, переносит их и тем самым облегчает распределение (но не поглощение!) по растению.Концевые двигатели водного тока. Для того чтобы вода двигалась по растению (и не просто двигалась, а поднималась вверх), необходимы затраты энергии, точки приложения которой располагаются по концам тока, вследствие чего они получили название концевых двигателей.Нижний концевой двигатель, или корневое давление. Его роль проявляется в основном при активном поглощении — нагнетании воды. При участии актомиозиноподобных сократительных белков он не только подает воду в корневую систему, но и проталкивает ее дальше в сосуды корня и вверх по стеблю. Нагнетание воды — активный энергозависимый процесс, который сильнее всего выражен в коре корня. Сила, развиваемая концевым двигателем, невелика, она может обеспечить подъем воды на высоту не более 1 м., т. е. достаточна для травянистых растений и небольших кустарников. Верхний концевой двигатель, или присасывающая сила транспирации. При постоянном испарении воды в листьях растений разбивается сосущая сила, отсасывающая воду от ближайших клеток и передающаяся др. кл., по к-ым передвигается вода, вплоть до сосудов. В сосудах нет цитоплазмы, поэтому нет и осмотического давления, и всасывание жидкости происходит при участии всей величины сосущей силы. Она позволяет поднять воду на несколько метров. Этой силы дост. для обеспечения водой кустарников и небольших деревьев.Поднятие воды по стволу Многие древесные раст. имеют большую протяженность ствола. У таких растений на помощь приходят силы сцепления между молекулами воды, которые очень велики и могут достигать 30 — 35 МПа. Этой силы достаточно для того, чтобы поднять воду на 1 — 2 км, что значительно превышает высоту любого дерева. Силы сцепления молекул воды действуют только при определенных условиях: водные струи в сосудах должны идти непрерывно, без пузырьков воздуха. Если же в них попадает в-х, передвижение воды прерывается. Непрерывность водных струй в сосудах достигается их сильным натяжением и плотным прилеганием к стенкам сосудов. Водные струи оказываются как бы висящими, они удерживаются в подвешенном состоянии сосущей силой трансп. и снизу поддерживаются силой корневого Р. При сильном натяжении водных струй пр-ит натяжение и самих сосудов, при этом у последних наблюдается увелич. длины и уменьшение поперечного сечения (подобно эластичной резине). Это создает предпосылки к смыканию стенок сосудов, что привело бы к их закрыванию и прекращению водного тока. Однако этого не пр-ит, т. к. сосуды имеют внутренние утолщения стенок — кольчатые, спиральные, лестничные, к-ые наподобие каркаса растягивают их и предохр. от смыкания. Величина его незначительна — всего 20 см воды на 1 см2 площади сосудов. Однако и этого достаточно при условии не очень сильного напряжения атмосферных факторов.
Физиологическая значимость процессов минерального питания, коэффициент накопления, органогены, определение макро- и микроэлементов, их представители. Первичные и вторичные макроэлементы. Тройное правило Арнона.
Растения могут поглощать и перемещать неорганические и органические вещества. Раньше считали, что для роста растений (фотосинтезирующих автотрофов) необходимы только минеральные элементы. Однако в последние годы показано, что многие водоросли требуют витамины В12, а также веществ, содержащих кобальт и т. д.Для наземных растений основным источником воды и минеральных веществ служит почва, не менее важна и атмосфера, которая обеспечивает фотосинтезирующие растения СО2, и, в свою очередь, сама атмосфера поглощает О2 и воду, выделяемую при транспирации. В системе растение-почва-атмосфера происходит значительный круговорот веществ, особенно воды и СО2.Концентрация минеральных веществ в самих растениях в большинстве случаев мало связана с их концентрацией в среде. Например, соотношение K+/Na+ в растениях обычно составляет 5–20, тогда как в среде, как правило, больше Na+. С другой стороны, элементы, содержащиеся в окружающей среде в низких концентрациях, могут накапливаться в растениях в значительном количестве.Эта способность растений характеризуется так называемым коэффициентом накопления:
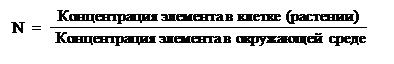 |
Еще раз отметим, что ни присутствие, ни концентрация минеральных элементов в самих растениях не является критерием необходимости. Хорошо зарекомендовало себя в этом отношении (относительно необходимости) так называемое тройное правило Арнона, которое было предложено еще в 1939 году. Элемент признается необходимым в случае, когда: 1. Растение без него не может закончить свой жизненный цикл; 2. Другой элемент не может заменить функцию изучаемого элемента; 3. Элемент непосредственно включен в метаболизм растения.Питательные элементы, которые используются растениями из субстрата в больших количествах (содержание их в золе 10–1–10–2 %), называют макроэлементами, а необходимые в значительно меньшем количестве (содержание 10–3 % и ниже) – микроэлементами.
Вода и СО2 рассматриваются отдельно, но в этом контексте необходимо считать их макроэлементами. К макроэлементам относятся K, Ca, Mg, N, P, S (Н2О и СО2), для галофитов следует добавить Na и Cl. Микроэлементы: Mn, Zn, Cu, Cl, B, Mo, сюда же относится и Fe, который располагается на границе макро- и микроэлементов.Водорослям необходим один или несколько из следующих элементов: Со, Si, I, V. Некоторым высшим растениям необходимы селен, кремний.Кроме необходимых элементов для роста, в оптимальных пределах должны быть и другие факторы: рН, температура, обеспечение О2, градиент водного потенциала, а для зеленых растений – освещенность соответствующей интенсивности и протяженности.Пассивный транспорт – перемещение веществ путем диффузии по градиенту электрохимического потенциала без затраты энергии (простая и, в какой-то мере, облегченная диффузия).Активный транспорт – перемещение веществ против градиента электрохимического потенциала с затратой метаболической энергии, как правило в форме АТФ или редокс-цепей.Для того чтобы понять механизмы трансмембранного переноса элементов минерального питания, остановимся на рассмотрении некоторых физико-химических закономерностях, определяющих движение ионов в растворе и мембране. Начнем с процессов пассивного транспорта (в частности вспомним законы диффузии). Согласно первому закону Фика, поток (Ф) прямо пропорционален коэффициенту диффузии D и градиенту концентрации dC/dх в точке х в данный момент времени.
Ф = – D
cyberpedia.su
Транспирация и верхний концевой двигатель
Химия и химическая технология
Статьи Рисунки Таблицы О сайте Реклама Передвижение воды по стеблю — ксилемный, или дальний, транспорт — большей частью представляют себе как пассивное движение по непрерывному акропетальному градиенту водного потенциала при участии двух концевых двигателей — нижнего (корневое давление) и верхнего (присасывающее действие транспирации), полагая, что никаких промежуточных двигателей в стебле нет. Правда, движению воды способствует непрерывность водной фазы в растении — от эпидермальных клеток корня до мезофилла листа — и колоссальное натяжение водных нитей в сосудах за счет свойственных воде огромных сил сцепления (см. главу I). Именно благодаря натяжению водных нитей в сосудах и непрерывности водной фазы всякое воздействие на лист, изменяющее скорость транспирации, или на корневую систему, изменяющее скорость поступления воды, влекут за собой мгновенную гидродинамическую реакцию, улавливаемую с помощью чувствительного датчика и аналогичную пульсовой волне в системе кровообращения. Скорость волны во много раз превышает скорость передвижения самой жидкости. Гидродинамические реакции возникают раньше биоэлектрических и, возможно, выполняют в растении даже какую-то информационную роль [337]. Но гидродинамические реакции к промежуточным двигателям непосредственного отношения не имеют. Теория промежуточных двигателей, как отмечает Н. А. Максимов [234], была опровергнута рядом опытов. Так, Е. Ф. Вотчал в своем обширном труде, опубликованном в 1897 г., установил, что вода движется по положенному горизонтально живому отрезку древесного ствола в несколько метров длиной с одинаковой скоростью как от нижнего конца к верхнему, так и наоборот, от верхнего к нижнему и что, следовательно, в древесине отсутствуют какие-либо клапаны, не пропускающие воду вниз а без таких клапанов не могли бы работать предполагаемые двигатели. Подобные же наблюдения были сделаны и другими учеными. Страс-бургеру (1893) и другим исследователям удалось показать, что введенные в перерезанные сосуды ядовитые растворы, например растворы пикриновой кислоты, беспрепятственно поднимаются по древесному стволу на много метров вверх, вплоть до самых листьев, хотя и отравляют на своем пути живые клетки. Точно так же удалось наблюдать беспрепятственное поднятие воды через участки травянистого стебля, убитые нагреванием, замораживанием или действием ядовитых веществ. Через некоторое время, однако, такие убитые участки стеблей пере- [c.147] Смотреть страницы где упоминается термин Транспирация и верхний концевой двигатель: [c.8] Смотреть главы в:Физиология растений -> Транспирация и верхний концевой двигатель
© 2018 chem21.info Реклама на сайте
chem21.info
Обратная связь ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Как цель узнает о ваших желаниях прежде, чем вы начнете действовать. Как компании прогнозируют привычки и манипулируют ими Целительная привычка Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Тренинг уверенности в себе Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Как слышать голос Бога Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| Поглощение воды корневой системой идет благодаря работе двух концевых двигателей водного тока: верхнего концевого двигателя, или присасывающей силы испарения (транспирации) , и нижнего концевого двигателя, или корневого двигателя. Основной силой, вызывающей поступление и передвижение воды в растении, является присасывающая сила транспирации, в результате которого возникает градиент водного потенциала. Водный потенциал – это мера энергии, используемой водой для передвижения. Водный потенциал и сосущая сила одинаковы по абсолютному значению, но противоположны по знаку. Чем меньше насыщенность водой данной системы, тем меньше (более отрицателен) ее водный потенциал. При потере воды растением в процессе транспирации создается ненасыщенность клеток листа водой, как следствие, возникает сосущая сила (водный потенциал падает), поступление воды идет в сторону большей сосущей силы, или меньшего водного потенциала.[1] Таким образом, верхний концевой двигатель водного тока в растении – это присасывающая сила транспирации листьев, и его работа мало связана с жизнедеятельностью корневой системы. Действительно, опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется. Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация. Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, препятствующей испарению, выделяют капельно-жидкую воду с небольшим количеством растворенных веществ – гуттация. Выделение жидкости идет через специальные водные устьица – гидаторы. Выделяющаяся жидкость – гутта. Таким образом, процесс гуттации является результатом одностороннего тока воды, происходящего в отсутствие транспирации, и, следовательно, вызывается какой-то иной причиной. К такому же выводу можно прийти и при рассмотрении явления плач растений. Если срезать побеги растения и к срезанному концу присоединить стеклянную трубку, то по ней будет подниматься жидкость. Анализ показывает, что это вода с растворенными веществами – пасока. В некоторых случаях, особенно в весенний период, плач наблюдается и при надрезе веток растений. Определения показали, что объем выделяющейся жидкости (пасоки) во много раз превышает объем корневой системы. Таким образом, плач – эт6о не просто вытекание жидкости в результате пореза. Все сказанное приводит к выводу, что плач, как и гуттация, связана с наличием одностороннего тока воды через корневые системы, не зависящего от транспирации. Силу, вызывающую односторонний ток воды по сосудам с растворенными веществами, не зависящую от процесса транспирации, называют корневым давлением. Наличие корневого давления позволяет говорить о нижнем концевом двигателе водного тока. Корневое давление можно измерить, присоединив манометр к концу, оставшемуся после срезания надземных органов растения, или поместив корневую систему в серию растворов различной концентрации и подобрав такую, при которой плач прекращается. Оказалось, что корневое давление равняется примерно 0,1 – 0,15 МПа (Д.А.Сабинин). Определения, проведенные советскими исследователями Л.В.Можаевой, В.Н.Жолкевичем, показали, что концентрация наружного раствора, останавливающего плач, значительно выше концентрации пасоки. Это позволило высказать мнение, что плач может идти против градиента концентрации. Было показано также, что плач осуществляется только в тех условиях, в которых нормально протекают все процессы жизнедеятельности клеток. Не только умерщвление клеток корня, но и снижение интенсивности их жизнедеятельности, в первую очередь интенсивность дыхания, прекращает плач. В отсутствии кислорода, под влиянием дыхательных ядов, при понижении температуры плач приостанавливается. Все сказанное позволило Д.А.Сабинину дать следующее определение: плач растений – это прижизненный односторонний ток воды и питательных веществ, зависящий от аэробной переработки ассимелятов. Д.А.Сабинин предложил схему, объясняющую механизм одностороннего тока воды в корне. Согласно этой гипотезе, клетки корня поляризованы в определенном направлении. Это проявляется в том, что в разных отсеках одной и той же клетки процессы обмена веществ различны. В одной части клетки идут усиленные процессы распада, в частности, крахмала на сахара, вследствие чего концентрация клеточного сока возрастает. На противоположном конце клетки преобладают процессы синтеза, благодаря чему концентрация растворенных веществ в этой части клетки уменьшается. Надо учитывать, что все эти механизмы будут работать только при достаточном количестве воды в среде и не нарушенном обмене веществ.[1] Согласно другой гипотезе, зависимость плача растений от интенсивности дыхания является косвенной. Энергия дыхания используется для поступления ионов в клетки коры, откуда они десорбируются в сосуды ксилемы. В результате концентрация солей в сосудах ксилемы повышается, что и вызывает поступление воды. |

 Отёска стен и прирубка косяков - Когда на доме не достаёт окон и дверей, красивое высокое крыльцо ещё только в воображении, приходится подниматься с улицы в дом по трапу.
Отёска стен и прирубка косяков - Когда на доме не достаёт окон и дверей, красивое высокое крыльцо ещё только в воображении, приходится подниматься с улицы в дом по трапу. Дифференциальные уравнения второго порядка (модель рынка с прогнозируемыми ценами) - В простых моделях рынка спрос и предложение обычно полагают зависящими только от текущей цены на товар.
Дифференциальные уравнения второго порядка (модель рынка с прогнозируемыми ценами) - В простых моделях рынка спрос и предложение обычно полагают зависящими только от текущей цены на товар.megapredmet.ru