
Биология и медицина. Стела у растений это
Стела и ее типы - это... Что такое Стела и ее типы?
Если сделать несколько последовательных поперечных срезов через осевые органы различных представителей плаунов, хвощей или папоротников, а также через молодые стебли (или корни) сосны или дуба, то можно легко убедиться в том, что система проводящих тканей у этих столь разных растений построена по разному архитектурному принципу. В одних случаях она представляет собой сплошной цилиндр, в других случаях внутренняя часть этого цилиндра полая или заполнена паренхимной тканью (сердцевиной). Наконец, во многих случаях проводящая ткань вокруг сеюдцевины не сплошная, но рассечена, причем рассечена по-разному у разных групп высших растений. Первичную проводящую систему осевых органов высших растений вместе со связанными с ней паренхимными тканями называют стелой (от греч. stela, stele — столб). Снаружи стела окружена корой, но граница между ними не всегда ясно выражена и в некоторых случаях анатомы затрудняются в проведении наружной границы стелы.
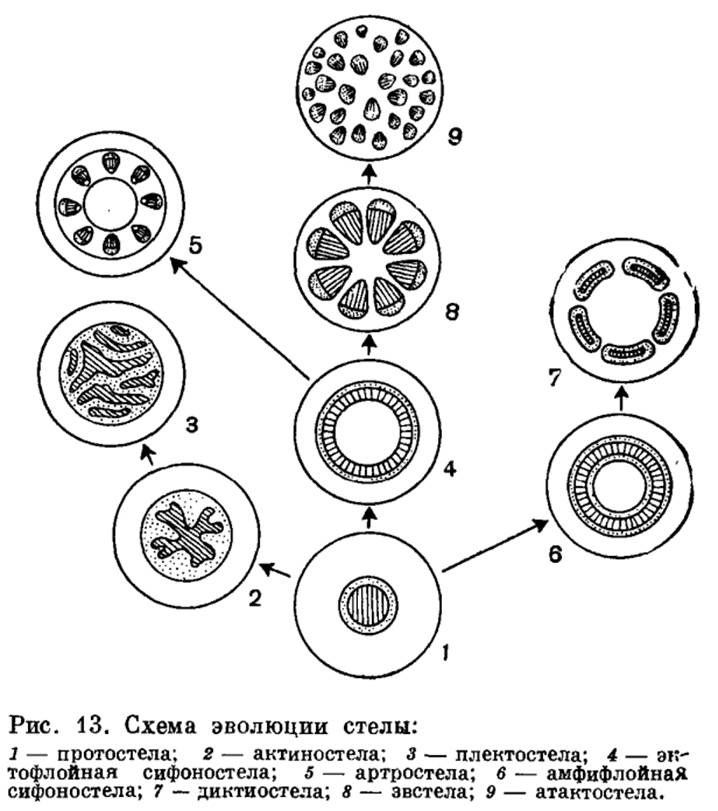
Изучение разных типов стелярной организации представляет большой интерес для понимания путей эволюции высших растений (рис. 13). Наиболее примитивным типом стелы является та, в которой проводящая ткань образует сплошную массу и центральное ядро, состоящее из ксилемы, полностью окружено слоем флоэмы. Это протостела. Протостела не только очень проста по структуре, но и представляет собой исходный тип, от которого в процессе эволюции произошли все другие типы стелы. Она была характерна для риниофитов и многих других вымерших форм. В современной флоре протостела характерна для плаунов и некоторых папоротников и встречается в корнях почти всех растений. На поперечном сечении протостела может иметь разную форму. Наиболее примитивным типом является протостела, состоящая из центрального пучка первичной ксилемы, окруженной цилиндром флоэмы. Подобную протостелу называют гаплостелой (от греч. haplos — простой). Более подвинутым типом является протостела с радиальными выступами ксилемы, называемая актиностелой (от греч. aktinos — луч). Она была характерна для девонского рода астероксилон (Asteroxylon) в встречается у некоторых примитивных видов плаунов. Результатом дальнейшей специализации актиностелы является так называемая плектостела (от греч. plectos — плетеный), характерная для стеблей большинства видов плауна. В плектостеле ксилема разбита на отдельные пластинки, расположенные параллельно или радиально друг к другу.
В процессе эволюции протостела дала начало сифоностеле (от греч. siphon — труба). В отличие от протостелы сифоностела имеет трубчатое строение и обладает центральной паренхимной частью, или сердцевиной. Эволюция сифоностелы сопровождается появлением паренхимных участков (лакун) в самом проводящем цилиндре. Благодаря лакунам создается непрерывная связь коры и сердцевины. Различные типы сифоностелы очень характерны для многих папоротников.
Когда сифоностела разбита на сеть или ряды продольных тяжей, получается рассеченная стела, или диктиостела (от греч. diktyon — сеть), которая, как и сифоностела, характерна для многих папоротников.
Особым типом стелы, производным от сифоностелы, является полициклическая сифоностела. Полициклическая стела— наиболее сложный тип стелярной организации— характерна для некоторых папоротников. Полициклические стелы являются всегда сифоностелическими. Они имеют внутреннюю проводящую систему, связанную с внешней сифоностелой. Эта внутренняя связь бывает всегда у узла. Типичные полициклические стелы имеют два или больше концентрических цилиндров проводящей ткани. В некоторых случаях внешний цилиндр является сифоностелой, а в других — диктиостелой.
У хвощей и родственных форм из сифоностелы возникает особый тип стелы, названный артростелой (от греч. arthron — член, сустав). Артростела имеет членистое строение и характеризуется наличием центральной полости и протоксилемных лакун (каринальных каналов). Она характерна для хвощей.
В результате рассечения сифоностелы возникает так называемая эвстела (от греч. eu — хорошо), характерная для большинства семенных растений.
В некоторых случаях, как у ряда травянистых двудольных и большинства однодольных, проводящие пучки эвстелы разбросаны по сердцевине и коре так, что сходство с единой сетью или кольцом теряется совершенно. Ввиду своеобразия подобной стелярной организации она выделяется в особый тип стелы, названный атактостелой (от греч. ataktos — беспорядочный). Атактостела развилась в результате крайнего рассечения эвстелы и представляет собой в эволюционном отношении крайне специализированную форму центрального цилиндра.
Сифоностелы, эвстелы и атактостелы почти всегда имеют больший диаметр, чем гаплостелы. Особенно хорошо видно это в тех случаях, когда стебель (или корень) бывает вначале гаплостелическим, а затем становится актиностелическим, плектостелическим, сифоностелическим или полициклическим. Эти факты приводят к выводу, что с изменением диаметра стелы изменяется также ее организация. Между величиной и формой стелы существует определенное соотношение, которое имеет большое физиологическое значение. Для нормального функционирования растения требуется достаточно большая поверхность соприкосновения между элементами ксилемы и живыми клетками. В ганлостеле, имеющей небольшой диаметр, отношение поверхности ксилемы к ее объему является относительно высоким и поэтому достаточным для ее жизнедеятельности. Но с увеличением размеров гаплостелы поверхность должна будет увеличиваться как квадрат ее линейных размеров, а объем как куб. Таким образом, с увеличением гаплостелы должно резко измениться отношение поверхности к объему. Это должно влиять как на поверхностные взаимоотношения ксилемы и флоэмы, так и на взаимоотношения между проводящей системой и другими частями оси. С другой стороны, если увеличение диаметра стебля или корня сопровождается изменением сложности очертания проводящей системы, то первоначальные пропорции между поверхностью и объемом проводящей ткани приблизительно сохраняются. Таким образом, возрастание сложности и размеров нужно рассматривать как приспособление для сохранения нормальных физиологических отношений внутри стебля или корня. Так как при данном объеме трубчатая или звездчатая масса ксилемы более эффективна для механической поддержки, чем сплошная цилиндрическая масса, то увеличение сложности стелярной организации дает также некоторые чисто физические преимущества.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
dic.academic.ru
Стела стебля растения
Стела стебля растения
Стела, занимающая центральную часть стебля, состоит из проводящих тканей, сердцевины (иногда она разрушается), перицикла (если он имеется) и тех постоянных тканей, которые из него возникают. Кнаружи от перицикла располагается первичная кора . В состав сформировавшейся первичной коры, которая образуется из части меристемы , располагающейся между образовательным кольцом и протодермой , входит паренхима , нередко колленхима и иногда секреторные элементы. Последовательность расположения этих тканей бывает различной. Паренхима (чаще хлорофиллоносная) обычно находится непосредственно под эпидермой , которая в отличие от эпиблемы корня имеет устьица . Колленхима может располагаться или глубже паренхимы, или также непосредственно под эпидермой . Самый внутренний слой первичной коры - эндодерма . Однако в стебле она никогда не бывает столь отчетливо развитой, как в корне . Нередко в эндодерме откладываются крахмальные зерна , поэтому ее часто называют крахмалоносным влагалищем. Иногда в ней заметны кристаллы оксалата кальция . Типичная эндодерма , клетки которой имеют лигнифицированные стенки, называемые поясками Каспари , обычно развивается лишь в корневищах . Изредка, как, например, в корневище ландыша , она бывает даже двуслойной.
Под проводящей тканью располагается сердцевина, обычно она состоит из относительно тонкостенных паренхимных клеток. В сердцевине часто откладываются запасные питательные вещества. Здесь же нередко встречаются идиобласты , заполненные таннидами , кристаллами, слизью и др. Иногда часть сердцевины разрушается и образуется полость. Периферическая часть сердцевины называется перимедуллярной зоной.
Типы стелы у высших растений разных таксонов неодинаковы. Представления об эволюции типов стелы обобщены в стелярной теории, основы которой в 1886 г. заложил французский ботаник ван Тигем. На рисунке 56 изображены различные типы стел и их вероятные генетические отношения.
Рассмотрим более подробно формирование и особенности стелы двудольных и однодольных . Прежде всего отметим, что у однодольных весь прокамбий полностью расходуется на формирование первичных проводящих тканей и первичная структура стебля сохраняется в течение всей жизни растений. У двудольных часть клеток прокамбия сохраняется и преобразуется в клетки камбия , за счет которых обеспечивается затем вторичный рост стебля в толщину.
В тех случаях, когда тяжи прокамбия у двудольных сближены и между ними из клеток образовательного кольца дифференцируется паренхима сердцевинных лучей, формируются изолированные проводящие пучки, располагающиеся кольцом. Флоэма в них ориентирована к периферии стебля, а ксилема - к центру. Такой тип строения стебля называется пучковым.
На границе стелы и первичной коры нередко располагаются волокна механической ткани . Иногда они образуют сплошное кольцо, подобно тому, что наблюдается в стебле льна , либо собраны около пучков (стебель клевера ). Их происхождение может быть перициклическим, но во многих случаях волокна возникают в первичной флоэме в результате дифференцировки клеток прокамбия .
Стела, главным компонентом которой являются расположенные кольцом проводящие пучки, получила название эустелы ( рис. 56 ). Эустела характерна для большинства двудольных .
В стеблях однодольных инициальное кольцо не возникает, а прокамбий распадается на множество тонких тяжей. В силу этого стела однодольных устроена иначе, чем стела двудольных . Особенности формирования проводящей системы у однодольных приводят к тому, что в большинстве случаев многочисленные закрытые (лишенные камбия) пучки равномерно распределены по всей толще стебля, занятой клетками основной паренхимы ( рис. 56 ). Хорошо выраженной сердцевины здесь нет, а граница центрального цилиндра в связи со слаборазвитым перициклом также не ясна. Такой тип стелы называют атактостелой .
medbiol.ru
Типы стели. Эволюция стели
Отличия высших растений от низших.
Высшие растения отличаются от низших по многим признакам, однако главное их отличие в том, что они по преимуществу - обитатели суши, устроены несравненно более сложно и более разнообразны, чем низшие растения. В отличие от низших растений, высшие растения представляют собой сложно дифференцированные многоклеточные организмы, приспособленные к жизни в наземной среде (за исключением немногочисленных и явно вторичных водных форм), с правильным чередованием двух поколений: полового (гаметофит) и бесполого (спорофит). На спорофитах высшисх растений развиваются многоклеточные спорангии, в которых образуются неподвижные споры, на гаметофитах — многоклеточные половые органы (гаметангии), однако у некоторых голосеменных и у всех цветковых растений гаметангии в процессе эволюции исчезли.
Спорангии одинаковые или чаще дифференцированы на 2 типа (микроспорангии и мегаспорангии), гаметангии — всегда двух типов — мужские (антеридии) и женские (архегонии). Как антеридии, так и архегонии защищены многоклеточными стенками. Женская гамета (яйцеклетка) всегда одна, неподвижна. Зигота у высших растений развивается в многоклеточном зародыше, который проходит первые стадии развития внутри женского гаметофита. Спорофит у высших растений, как правило, расчленён на 3 основных органа — лист, стебель и корень. В большинстве случаев у высших растений развивается специальная проводящая ткань — ксилема и флоэма, у некоторых групп редуцировавшаяся. Имеется эпидерма с кутикулой и типичными устьицами. Хлорофилл у высших растений не содержит дополнительных пигментов, окраска фотосинтезирующих частей зелёная. Еще более показательны различия вклада высших и низших растений в продуктивность и биомассу биосферы. Низшие растения производят примерно 1/3 первичной биологической продукции, но ввиду того, что длительность жизни большинства из них невелика, биомасса низших растений составляет доли процента от общей массы продуцентов биосферы.Впрочем, разделение на высшие и низшие только по среде обитания будет не совсем верным. Часть высших растений ведет водный образ жизни, они называются гидрофитами. Среди гидрофитов много обитателей пресноводных водоемов (Nymphaeaceae, Lemnaceae, Potamogetonaceae и др.). В то же время, многие водоросли ведут наземный образ жизни и заселяют поверхность почвы и даже скальных пород. Однако в обоих случаях водная и наземная среда вторичны, то есть гигрофиты - это высшие растения, которые перешли от сухопутного к водному образу жизни, а наземные водоросли - водные растения, которые поселились на суше. Все водоросли могут нормально функционировать только в периоды, когда субстрат их обитания обильно увлажнен. В другое время они находятся в диапаузе.
Типы стели. Эволюция стели.
Стела занимает центральную часть стебля, состоит из проводящих тканей, сердцевины (иногда она разрушается), перицикла (если он имеется) и тех постоянных тканей, которые из него возникают. Кнаружи от перицикла (наружной границы стелы) располагается первичная кора.
Протостела - наиболее примитивный тип стелы, в которой проводящая ткань образует сплошную массу и центральное ядро, состоящее из ксилемы, полностью окружено слоем флоэмы. Протостела не только очень проста по структуре, но и представляет собой исходный тип, от которого в процессе эволюции произошли все другие типы стелы. Она была характерна для риниофитов и многих других вымерших форм. В современной флоре протостела характерна для плаунов и некоторых папоротников и встречается в корнях почти всех растений. На поперечном сечении протостела может иметь разную форму. Наиболее примитивным типом является протостела, состоящая из центрального пучка первичной ксилемы, окруженной цилиндром флоэмы. Подобную протостелу называют гаплостелой. Более подвинутым типом является протостела с радиальными выступами ксилемы, называемая актиностелой. Она встречается у некоторых примитивных видов плаунов. Результатом дальнейшей специализации актиностелы является так называемая плектостела, характерная для стеблей большинства видов плауна. В плектостеле ксилема разбита на отдельные пластинки, расположенные параллельно или радиально друг к другу.
В процессе эволюции протостела дала начало сифоностеле. В отличие от протостелы сифоностела имеет трубчатое строение и обладает центральной паренхимной частью, или сердцевиной. Эволюция сифоностелы сопровождается появлением паренхимных участков (лакун) в самом проводящем цилиндре. Благодаря лакунам создается непрерывная связь коры и сердцевины. Различные типы сифоностелы очень характерны для многих папоротников.
Когда сифоностела разбита на сеть или ряды продольных тяжей, получается рассеченная стела, или диктиостела, которая, как и сифоностела, характерна для многих папоротников.
У хвощей и родственных форм из сифоностелы возникает особый тип стелы, названный артростелой. Артростела имеет членистое строение и характеризуется наличием центральной полости и протоксилемных лакун (каринальных каналов). Она характерна для хвощей.
В результате рассечения сифоностелы возникает так называемая эвстела, характерная для большинства семенных растений.
В некоторых случаях, как у ряда травянистых двудольных и большинства однодольных, проводящие пучки эвстелы разбросаны по сердцевине и коре так, что сходство с единой сетью или кольцом теряется совершенно. Ввиду своеобразия подобной стелярной организации она выделяется в особый тип стелы, названный атактостелой. Атактостела развилась в результате крайнего рассечения эвстелы и представляет собой в эволюционном отношении крайне специализированную форму центрального цилиндра.
Жизненная форма растений, биологическая форма, биоморфа — внешний облик растений, отражающий их приспособленность к условиям среды. Форма, в которой вегетативное тело растения находится в гармонии с внешней средой в течение всей жизни, от семени до отмирания.
Жизненная форма растения не остается постоянной, а может меняться по мере взросления и старения. Важнейшую роль в становлении жизненной формы играет внешняя среда. Но это, конечно, не означает, что жизненная форма любого растения бесконечно пластична и зависит только от непосредственно действующих в данный момент условий.
Каждый вид растений реагирует на внешние воздействия в рамках своих наследственно закрепленных возможностей. Земляника, например, не станет развесистым деревом даже в самой благоприятной для роста и ветвления обстановке. Подразумевается, что в сложившейся жизненной форме каждого вида проявляются черты наследственной, выработанной в процессе естественного отбора приспособленности к определенному комплексу внешних факторов.
Использовав и обобщив предложенные в разное время классификации, отечественный ботаник И.Г. Серебряков предложил называть жизненной формой своеобразный габитус определенных групп растений, возникающий в результате роста и развития в определенных условиях – как выражение приспособленности к этим условиям.
В основу своей классификации И.Г. Серебряков положил признак продолжительности жизни всего растения и его скелетных осей. Он выделил следующие жизненные формы растений:
А. Древесные растения
Деревья
Кустарники
Кустарнички
studopedya.ru
Биология для студентов - 24. Стелярная теория
Стела – совокупность первичных проводящих пучков вместе с заключенными между ними прочими тканями и примыкающим к коре перициклом.

Эволюция типов стелы (по Бавтуто, Еремин, 1996): 1 – протостела; 2 – актиностела; 3 – плектостела; 4 – эктофлойная сифоностела; 5 – артростела; 6 – амфифлойная сифо- ностела; 7 – диктиостела; 8- эустела; 9 – атактостела.
В 1886 году Ф. Тигем и А.Дулио впервые ввели понятие «стела», заложив основы стелярной теории – учения о типах строения и закономерностях эволюции центрального цилиндра всех высших растений.
Наиболее древний и примитивный тип стелы – протостела – присущ первым наземным растениям (риниофитам). Последующая эволюция стелы была связана с увеличением поверхности проводящих тканей. Из протостелы развилась актиностела, ксилема которой на поперечном срезе имеет вид звезды. Данный тип стелы также характерен для примитивных растений (плауновидных, вымерших хвощевидных). Переход от протостелы к актиностеле связан с возникновением пучков, идущих в боковые органы; кроме того, в актиностеле проводящие ткани имеют большую поверхность соприкосновения с окружающими живыми тканями, что способствует лучшему проведению веществ.
Результат дальнейшей специализации актиностелы – появление плектостелы, характерной для осевых органов плауновидных. В плектостеле ксилема представлена отдельными участками, расположенными параллельно или радиально друг к другу. В процессе эволюции протостела дала также начало сифоностеле. Сифоностела имеет трубчатое строение и обладает сердцевиной. Возникновение сифоностелы обеспечило существование более крупных растений, т. к. расположение ксилемы на периферии сделало стебли более прочными. Возникновение сердцевины увеличило поверхность соприкосновения проводящих тканей с живыми клетками, а также создало возможность запасания питательных веществ. Известны два типа сифоностелы:
- эктофлойная (флоэма охватывает ксилему снаружи),
- амфифлойная (флоэма окружает ксилему снаружи и изнутри).
Дальнейшее эволюционное усложнение стелы связано с развитием крупных боковых органов. В местах их отхождения от стебля в стеле, в области узлов, образуются большие листовые лакуны (прорывы), заполненные паренхимой. Вследствие этого сифоностела паренхимными полосами (радиальными лучами), идущими от сердцевины к коре в радиальном направлении, рассекается на отдельные пучки, сохраняющие общее положение по цилиндрической поверхности (на поперечном срезе – кольцом). Возникают диктиостела, артростела и эустела. Диктиостела (характерна для папоротников, например, для орляка, щитовника, кочедыжника) имеет вид сетчатой трубки. Она возникает из амфифлойной сифоностелы вследствие образования большого количества листовых прорывов. Эустела встречается у семенных растений и составлена из коллатеральных открытых пучков, расположенных по кругу. Артростела (характерна для хвощей) является разновидностью эустелы, представлена закрытыми пучками, расположенными вокруг центральной полости стелы и соединяющимися в узлах.
Последнее звено в эволюции стелы стебля представляет собой атактостела однодольных растений. Ей присущи закрытые коллатеральные пучки, сложная картина прохождения пучков по стеблю (дугообразные индивидуальные листовые следы) с их разбросанным расположением по всему поперечному сечению стебля. Эволюция типов стелы показана.
vseobiology.ru
Стела и ее типы
Если сделать несколько последовательных поперечных срезов через осевые органы различных представителей плаунов, хвощей или папоротников, а также через молодые стебли (или корни) сосны или дуба, то можно легко убедиться в том, что система проводящих тканей у этих столь разных растений построена по разному архитектурному принципу. В одних случаях она представляет собой сплошной цилиндр, в других случаях внутренняя часть этого цилиндра полая или заполнена паренхимной тканью (сердцевиной). Наконец, во многих случаях проводящая ткань вокруг сеюдцевины не сплошная, но рассечена, причем рассечена по-разному у разных групп высших растений. Первичную проводящую систему осевых органов высших растений вместе со связанными с ней паренхимными тканями называют стелой (от греч. stela, stele — столб). Снаружи стела окружена корой, но граница между ними не всегда ясно выражена и в некоторых случаях анатомы затрудняются в проведении наружной границы стелы.
Изучение разных типов стелярной организации представляет большой интерес для понимания путей эволюции высших растений (рис. 13). Наиболее примитивным типом стелы является та, в которой проводящая ткань образует сплошную массу и центральное ядро, состоящее из ксилемы, полностью окружено слоем флоэмы. Это протостела. Протостела не только очень проста по структуре, но и представляет собой исходный тип, от которого в процессе эволюции произошли все другие типы стелы. Она была характерна для риниофитов и многих других вымерших форм. В современной флоре протостела характерна для плаунов и некоторых папоротников и встречается в корнях почти всех растений. На поперечном сечении протостела может иметь разную форму. Наиболее примитивным типом является протостела, состоящая из центрального пучка первичной ксилемы, окруженной цилиндром флоэмы. Подобную протостелу называют гаплостелой (от греч. haplos — простой). Более подвинутым типом является протостела с радиальными выступами ксилемы, называемая актиностелой (от греч. aktinos — луч). Она была характерна для девонского рода астероксилон (Asteroxylon) в встречается у некоторых примитивных видов плаунов. Результатом дальнейшей специализации актиностелы является так называемая плектостела (от греч. plectos — плетеный), характерная для стеблей большинства видов плауна. В плектостеле ксилема разбита на отдельные пластинки, расположенные параллельно или радиально друг к другу.
В процессе эволюции протостела дала начало сифоностеле (от греч. siphon — труба). В отличие от протостелы сифоностела имеет трубчатое строение и обладает центральной паренхимной частью, или сердцевиной. Эволюция сифоностелы сопровождается появлением паренхимных участков (лакун) в самом проводящем цилиндре. Благодаря лакунам создается непрерывная связь коры и сердцевины. Различные типы сифоностелы очень характерны для многих папоротников.
Когда сифоностела разбита на сеть или ряды продольных тяжей, получается рассеченная стела, или диктиостела (от греч. diktyon — сеть), которая, как и сифоностела, характерна для многих папоротников.
Особым типом стелы, производным от сифоностелы, является полициклическая сифоностела. Полициклическая стела— наиболее сложный тип стелярной организации— характерна для некоторых папоротников. Полициклические стелы являются всегда сифоностелическими. Они имеют внутреннюю проводящую систему, связанную с внешней сифоностелой. Эта внутренняя связь бывает всегда у узла. Типичные полициклические стелы имеют два или больше концентрических цилиндров проводящей ткани. В некоторых случаях внешний цилиндр является сифоностелой, а в других — диктиостелой.
У хвощей и родственных форм из сифоностелы возникает особый тип стелы, названный артростелой (от греч. arthron — член, сустав). Артростела имеет членистое строение и характеризуется наличием центральной полости и протоксилемных лакун (каринальных каналов). Она характерна для хвощей.
В результате рассечения сифоностелы возникает так называемая эвстела (от греч. eu — хорошо), характерная для большинства семенных растений.
В некоторых случаях, как у ряда травянистых двудольных и большинства однодольных, проводящие пучки эвстелы разбросаны по сердцевине и коре так, что сходство с единой сетью или кольцом теряется совершенно. Ввиду своеобразия подобной стелярной организации она выделяется в особый тип стелы, названный атактостелой (от греч. ataktos — беспорядочный). Атактостела развилась в результате крайнего рассечения эвстелы и представляет собой в эволюционном отношении крайне специализированную форму центрального цилиндра.
Сифоностелы, эвстелы и атактостелы почти всегда имеют больший диаметр, чем гаплостелы. Особенно хорошо видно это в тех случаях, когда стебель (или корень) бывает вначале гаплостелическим, а затем становится актиностелическим, плектостелическим, сифоностелическим или полициклическим. Эти факты приводят к выводу, что с изменением диаметра стелы изменяется также ее организация. Между величиной и формой стелы существует определенное соотношение, которое имеет большое физиологическое значение. Для нормального функционирования растения требуется достаточно большая поверхность соприкосновения между элементами ксилемы и живыми клетками. В ганлостеле, имеющей небольшой диаметр, отношение поверхности ксилемы к ее объему является относительно высоким и поэтому достаточным для ее жизнедеятельности. Но с увеличением размеров гаплостелы поверхность должна будет увеличиваться как квадрат ее линейных размеров, а объем как куб. Таким образом, с увеличением гаплостелы должно резко измениться отношение поверхности к объему. Это должно влиять как на поверхностные взаимоотношения ксилемы и флоэмы, так и на взаимоотношения между проводящей системой и другими частями оси. С другой стороны, если увеличение диаметра стебля или корня сопровождается изменением сложности очертания проводящей системы, то первоначальные пропорции между поверхностью и объемом проводящей ткани приблизительно сохраняются. Таким образом, возрастание сложности и размеров нужно рассматривать как приспособление для сохранения нормальных физиологических отношений внутри стебля или корня. Так как при данном объеме трубчатая или звездчатая масса ксилемы более эффективна для механической поддержки, чем сплошная цилиндрическая масса, то увеличение сложности стелярной организации дает также некоторые чисто физические преимущества.
Поделитесь на страничкеslovar.wikireading.ru
(жр) Стела и её типы — Zbio
Материал из Zbio
А. Л. ТахтаджянЕсли сделать несколько последовательных поперечных срезов через осевые органы различных представителей плаунов, хвощей или папоротников, а также через молодые стебли (или корни) сосны или дуба, то можно легко убедиться в том, что система проводящих тканей у этих столь разных растений построена по разному архитектурному принципу. В одних случаях она представляет собой сплошной цилиндр, в других случаях внутренняя часть этого цилиндра полая или заполнена паренхимной тканью (сердцевиной). Наконец, во многих случаях проводящая ткань вокруг сердцевины не сплошная, но рассечена, причем рассечена по-разному у разных групп высших растений. Первичную проводящую систему осевых органов высших растений вместе со связанными с ней паренхимными тканями называют стелой (от греч. stela, stele — столб). Снаружи стела окружена корой, но граница между ними не всегда ясно выражена и в некоторых случаях анатомы затрудняются в проведении наружной границы стелы.
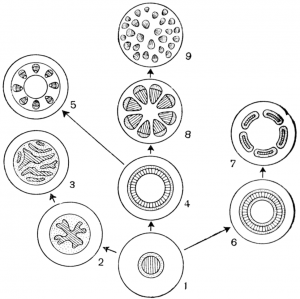
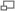 Рис. 13. Схема эволюции стелы:1 - протостела; 2 - актиностела; 3 - плектостела; 4 - эктофлойная сифоностела; 5 - артростела; 6 - амфифлойная сифоностела; 7 - диктиостела; 8 - эвстела; 9 - атактостела.
Рис. 13. Схема эволюции стелы:1 - протостела; 2 - актиностела; 3 - плектостела; 4 - эктофлойная сифоностела; 5 - артростела; 6 - амфифлойная сифоностела; 7 - диктиостела; 8 - эвстела; 9 - атактостела. Изучение разных типов стелярной организации представляет большой интерес для понимания путей эволюции высших растений (рис. 13). Наиболее примитивным типом стелы является та, в которой проводящая ткань образует сплошную массу и, центральное ядро, состоящее из ксилемы, полностью окружено слоем флоэмы. Это протостела. Протостела не только очень проста по структуре, по и представляет собой исходный тип, от которого в процессе эволюции произошли все другие типы стелы. Она была характерна для риниофитов и многих других вымерших форм. В современной флоре протостела характерна для плаунов и некоторых папоротников и встречается в корнях почти всех растений. На поперечном сечении протостела может иметь разную форму. Наиболее примитивным типом является протостела, состоящая из центрального пучка первичной ксилемы, окруженной цилиндром флоэмы. Подобную протостелу называют гаплостелой (от греч. haplos — простой). Более подвинутым типом является протостела с радиальными выступами ксилемы, называемая актиностелой (от греч. aktinos — луч). Она была характерна для девонского рода астероксилон (Asteroxylon) и встречается у некоторых примитивных видов плаунов. Результатом дальнейшей специализации актиностелы является так называемая плектостела (от греч. plectos — плетеный), характерная для стеблей большинства видов плауна. В плектостеле ксилема разбита на отдельные пластинки, расположенные параллельно или радиально друг к другу.
В процессе эволюции протостела дала начало сифоностеле (от греч. siphon — труба). В отличие от протостелы сифоностела имеет трубчатое строение и обладает центральной паренхимной частью, или сердцевиной. Эволюция сифоностелы сопровождается появлением паренхимных участков (лакун) в самом проводящем: цилиндре. Благодаря лакунам создается непрерывная связь коры и сердцевины. Различные типы сифоностелы очень характерны для многих папоротников.
Когда сифоностела разбита на сеть или ряды продольных тяжей, получается рассеченная стела, или диктиостела (от греч. diktyon — сеть), которая, как и сифоностела, характерна для многих папоротников.
Особым типом стелы, производным от сифоностелы, является полициклическая сифоностела. Полициклическая стела- наиболее сложный тип стелярной организации- характерна для некоторых папоротников. Полициклические стелы являются всегда сифоностелическими. Они имеют внутреннюю проводящую систему, связанную с внешней сифоностелой. Эта внутренняя связь бывает всегда у узла. Типичные полициклические стелы имеют два или больше концентрических цилиндров проводящей ткани. В некоторых случаях внешний цилиндр является сифоностелой, а в других — диктиостелой.
У хвощей и родственных форм из сифоностелы возникает особый тип стелы, названный артростелой (от греч. arthron — член, сустав). Артростела имеет членистое строение и характеризуется наличием центральной полости и протоксилемных лакун (каринальных каналов). Она характерна для хвощей.
В результате рассечения сифоностелы возникает так называемая эвстела (от греч. eu — хорошо), характерная для большинства семенных растений.
В некоторых случаях, как у ряда травянистых двудольных и большинства однодольных, проводящие пучки эвстелы разбросаны по сердцевине и коре так, что сходство с единой сетью или кольцом теряется совершенно. Ввиду своеобразия подобной стеляриой организации она выделяется в особый тип стелы, названный атактостелой (от греч. ataktos — беспорядочный). Атактостела развилась в результате крайнего рассечения эвстелы и представляет собой в эволюционном отношении крайне специализированную форму центрального цилиндра.
Сифоностелы, эвстелы и атактостелы почти всегда имеют больший диаметр, чем гаплостелы. Особенно хорошо видно это в тех случаях, когда стебель (или корень) бывает вначале гаплостелическим, а затем становится актиностелическим, плектостелическим, сифоностелическим или полициклическим. Эти факты приводят к выводу, что с изменением диаметра стелы изменяется также ее организация. Между величиной и формой стелы существует определенное соотношение, которое имеет большое физиологическое значение. Для нормального функционирования растения требуется достаточно большая поверхность соприкосновения между элементами ксилемы и живыми клетками. В гаплостеле, имеющей небольшой диаметр, отношение поверхности ксилемы к ее объему является относительно высоким и поэтому достаточным для ее жизнедеятельности. Но с увеличением размеров гаплостелы поверхность должна будет увеличиваться как квадрат ее линейных размеров, а объем как куб. Таким образом, с увеличением гаплостелы должно резко измениться отношение поверхности к объему. Это должно влиять как на поверхностные взаимоотношения ксилемы и флоэмы, так и на взаимоотношения между проводящей системой и другими частями оси. С другой стороны, если увеличение диаметра стебля или корня сопровождается изменением сложности очертания проводящей системы, то первоначальные пропорции между поверхностью и объемом проводящей ткани приблизительно сохраняются. Таким образом, возрастание сложности и размеров нужно рассматривать как приспособление для сохранения нормальных физиологических отношений внутри стебля или корня. Так как при данном объеме трубчатая или звездчатая масса ксилемы более эффективна для механической поддержки, чем сплошная цилиндрическая масса, то увеличение сложности стелярной организации дает также некоторые чисто физические преимущества.
molbiol.ru
Стела (ботаника) — WiKi
Сте́ла, или Стель, или Сте́ле, или Центра́льный цили́ндр, или Осево́й цилиндр — комплекс первичных тканей, лежащих внутри от первичной коры сосудистых растений.
Основы стелярной теории заложил французский ботаник Филипп ван Тигем.
Протостела
Первые три типа стел (протостелы) характерны для наиболее примитивных сосудистых растений.
Для них характерно отсутствие сердцевины и листовых лакун (для таких растений характерны энационные листья), центрархная или экзархная дифференцировка ксилемы, которая состоит только из трахеид, и отсутствие камбия.
- Гаплостела — наиболее примитивный тип стелы. Она состоит из сплошного тяжа ксилемы, покрытого слоем флоэмы. Ксилема — центрархного типа и содержит только трахеиды. Характерна для риниофитов и некоторых современных споровых растений.
- Актиностела («звёздчатая» стела) развилась из гаплостелы, ксилема в которой располагается в форме звезды, между лучами которой располагается флоэма. Ксилема экзархного типа, дифференцируется центростремительно. С развитием актиностелы поверхность контакта с живыми непроводящими тканями растения увеличилась, от стелы начали отходить листовые проводящие пучки. Ксилема сложена трахеидами и расположенными в её центральной части волокнистыми трахеидами, флоэма — ситовидными клетками и лубяной паренхимой. Актиностела характерна для плаунов.
- Плектостела («губчатая» стела) — следующий этап развития, в котором лучи ксилемы рассекаются флоэмой и паренхимой. Ксилема здесь также экзархного типа. Ксилема сложена трахеидами, флоэма — ситовидными клетками и лубяной паренхимой. Она характерна также для плаунов.
Сифоностела
Для более продвинутых растений с теломными листьями характерны следующие стелы (сифоностелы):
- Соленостела — (греч. solen — трубка) — у папоротников — ксилема с двух сторон окружена лубом флоэмой, причем внутренняя флоэма ограничивает снаружи паренхимную сердцевину.
- Диктиостела — у папоротниковидных. Кольцо проводящих тканей, окружающее сердцевину, разорвано паренхиматизированными листовыми лакунами на небольшие участки — меристелы. Меристела состоит из мезархной ксилемы, включающей в себя трахеиды и клетки паренхимы, флоэмы и перицикла и окружена эндодермой. Сердцевина дифференцируется из избытка клеток, образовавшихся в процессе деления апикальной клетки.
- Эустела характерна для голосеменных и двудольных. Состоит из пучков, расположенных кольцом, и отделенных друг от друга паренхимными сердцевинными лучами. У травянистых растений лучи более широкие, у древесных — более узкие, иногда даже из 1 ряда клеток.
Литература
- Серебрякова Т. И., Воронин Н. С., А. Г. Еленевский и др. «Ботаника с основами фитоценологии. Анатомия и морфология растений» — М.: «Академкнига», 2006.
Ссылки
ru-wiki.org










