
Ростовые движения растений. Способы движения растений
Движения растений адаптация и устойчивость растений к неблагоприятным факторам
9
I Движения растений
II Адаптация и устойчивость растений к неблагоприятным факторам
1. Стресс, адаптация, устойчивость
2. Неспецифическая реакция растений на стресс.
3. Засухоустойчивость и устойчивость к перегреву.
4. Устойчивость растений к низким температурам.
5. Солеустойчивость.
6. Устойчивость к недостатку кислорода.
7. Газоустойчивость.
8. Радиоустойчивость.
9. Устойчивость растений к тяжелым металлам.
10. Устойчивость растений к патогенам.
I Движения растений
Выделяют следующие способы движения у растений:
1.Движения цитоплазмы и органоидов.
2. Локомоторные движения с помощью жгутиков (таксисы).
3. Верхушечный рост (корневые волоски, пыльцевые трубки).
4. Ростовые движения (нутации, тропизмы, ростовые настии).
5. Тургорные движения (движения устьиц, настии, сейсмонастии).
Из всех перечисленных форм движения 3-5 типы свойственны только растениям.
Движение цитоплазмы осуществляется за счет взаимодействия актиновых микрофилламентов эндоплазмой, находящейся в состоянии золя и микротрубочками гелеобразной эктоплазмы. Этот процесс является энергозависимым и нуждается в присутствии ионов Са2+ в концентрациях менее 0,1 мкмоль/л. Различают колебательное, циркуляционное, ротационное и фонтанирующее движения цитоплазмы. Средняя скорость движения цитоплазмы у разных клеток варьирует от 2-5 мкм/с до 1350.
В основе локомоторных способов движения клеток лежит функционирование систем сократительных белков, обеспечивающих превращение химической энергии АТФ в механическую энергию, например, движения жгутиков. Перемещение всего организма в пространстве под действием односторонне действующих факторов (раздражителей) называется таксисом. Этот тип движения характерен для одноклеточных жгутиковых водорослей, для гамет и зооспор водорослей, для сперматозоидов мхов, плаунов, хвощей и папоротников. В основе движения жгутика лежит функционирование белковой системы тубулин-динеин, которая за счет энергии АТФ обеспечивает механическое движение этого органа.
Перемещение в пространстве корневых волосков, пыльцевых трубок, гиф грибов, протонемы мхов осуществляется за счет верхушечного роста. Это высокополяризованный тип роста, при котором в клетках соблюдается строгая пространственная локализация интенсивности синтеза вдоль продольной оси клетки. Секреторные везикулы транспортируются к кончику клетки, куда доставляют материал для синтеза стенки, а также синтазы и литические ферменты, которые разрыхляют структуру стенки, делая ее неспособной противостоять высокому тургорному давлению. Микрофибриллы стенки раздвигаются и новые компоненты встраиваются в разрыхленную стенку.
Тропизмы – это ростовые движения растений, обусловленные изгибанием или искривлением органов в ответ на факторы среды, действующие односторонне. Они осуществляются в растущих частях растений и, как правило, являются следствием более быстрого роста клеток растяжением на одной стороне побега, корня или листа. Различают геотропизм (ориентация в пространстве и изгибание под действием гравитационного поля), фототропизм (ростовые изгибы под действием одностороннего освещения), хемотропизм (ростовая реакция на градиент химических соединений), тигмотропизм (ростовые изгибы в ответ на прикосновение, например, у усиков некоторых лазящих растений), травмотропизм (изгиб в сторону от нанесенного повреждения).
Фототропизм. Согласно гипотезе Н.Г.Холодного- Ф.Вента фототропический стимул вызывает поперечную электрическую поляризацию клеток в верхушке колеоптиля, причем неосвещенная сторона приобретает положительный заряд, что приводит к перераспределению ауксина. ИУК передвигается на затененную сторону, в результате чего рост этой стороны усиливается. С помощью подобного механизма затененные участки листовых пластинок выходят из тени, создавая эффект «мозаики листьев».
Настии – обратимые движения (изгибы) органов с дорсивентральным строением в ответ на изменение диффузно действующих факторов внешней среды (например, открывает и закрывает цветков при смене дня и ночи). Направленность настических движений зависит от структуры самого органа. При более быстром росте верхней стороны орган (лист, лепесток) изгибается книзу (эпинастия), при обратном соотношении скоростей роста– кверху (гипонастия). Различают фото-, термо-, гидро-, хемо-, тигмо-, сейсмо-, травмо- и электронастии. Настические движения обеспечивают защиту органов (закрывание цветков, опускание листьев) или захват предметов (движение усиков, железистых волосков росянки).
Нутации – круговые или колебательные движения органов растений. Так, растущий побег совершает качания, в результате чего его верхушка колеблется относительно продольной оси. Особенно хорошо круговые нутации выражены у стеблей вьющихся растений (лиан). Длительность одного оборота у этих растений – от 2 до12 ч. Молодой побег вьющегося растения сначала растет вертикально, затем его верхушка изгибается и занимает горизонтальное положение. Вслед за этим наблюдаются круговые нутации и побег обвивается вокруг опоры. У многих лазающих растений круговые нутации совершает вся верхняя часть побега. Круговые нутации лиан и лазящих растений служат для поиска опоры, необходимой для движения к свету.
Обратимые тургорные движения также обусловлены дорсивентральным строением органов и вызываются теми же диффузно действующими факторами внешней среды. Однако они не зависят от процессов роста и обуславливают двигательную активность тех органов, которые закончили свой рост. Такие движения осуществляются благодаря увеличению и уменьшению в вакуолях специализированных клеток концентрации осмотически активных веществ (К+, малата), в результате чего соответственно увеличиваются или уменьшаются поглощение воды и тургорное давление. Например, движения устьиц или опускание листьев на ночь (никтинастии). К медленным тургорным движениям относят также выделение железистыми волосками росянки муравьиной кислоты в ответ на прикосновение шершавых предметов.
К быстрым тургорным движениям относятся сейсмонастические (индуцируемые сотрясением) движения сложных листьев некоторых бобовых (мимозы, клевера), кислицы и др. В ответ на механический удар или встряхивание нижние клетки главного сочленение мимозы способны быстро ( в течение секунд) терять тургор, что приводит к опусканию листа. Поднимается лист медленнее (30-60 мин). Раздражение вызывает возникновение в листе потенциала действия с амплитудой 50-100 мВ, который распространяется со скоростью 0,5 – 4 см/с. Физиологическое значение опускания листьев у мимозы при сотрясении, по-видимому, связано с предотвращением повреждений от сильных ливней и ураганных ветров.
studfiles.net
Ростовые движения растений.
Количество просмотров публикации Ростовые движения растений. - 606
Фотопериодизм и термопериодизм и способы его регулирования у растений.
Период покоя у семян и приемы ускорения их прорастания.
Вынужденный покой семян – такое физиологическое состояние семян, при котором жизнеспособные семена не прорастают из-за неблагоприятных внешних условий (температура, влажность).
Глубокий (органический) покой – не зависит от внешних условий, а связан с особенностями внутреннего ритма развития зародыша и строением семенных покровов. Он подразделяется на:
1.Эндогенный – связан с накоплением в семенах ингибиторов роста͵ которые блокируют ДНК.
2.Экзогенный – связан с непроницаемость семенной кожуры для воды и газов.
3.Комбинированный – связан с непроницаемостью семенных покровов и неполным формированием зародыша.
Существуют разнообразные пути преодоления покоя у семян растений:
1.Стратификация – переслаивание семян древесных растений с влажным песком и выдерживание их при низких положительных температурах.
2.Скарификация – нарушение целостности оболочки семян с твердым покрытием механическим путем, кислотами, кипятком.
3.Промывание семян проточной водой – удаляются с поверхности семян ингибиторов роста.
4.Воздействие светом – под действием света разрушаются ингибиторов роста растений.
Внешние факторы, влияющие на переход растения от вегетативной фазы к генеративной:
1. Наличие углеводов в растении.
2. Высокий уровень фосфорного питания.
3. Хорошее освещение.
4. Благоприятный водный режим.
5. Благоприятный температурный режим.
6. Длина светового дня.
Внутренние причины:
1. Изменения в обмене веществ, вызванные гормональными регуляторами роста и развития растений.
Фотопериодизм – реакция растений на продолжительность дня и ночи (не связан с фотосинтезом и не зависит от интенсивности света).
Термопериодизм (яровизация) – процесс, протекающий в озимых формах однолетних и двулетних растений под действием низких положительных температур, вызывающий ускорение развития этих растений.
Тропизмы – движения органов растений и изменения их положения в пространстве. Тропизмы бывают:
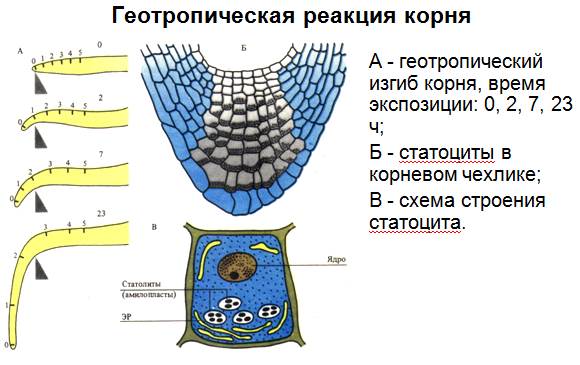
1. Геотропизм – движения растений в результате силы земного притяжения (корень).
2. Фототропизм – движение листьев и стеблей растений к источнику света.
3. Гелиотропизм – движение растений за ходом солнца (подсолнечник).
4. Хемотропизм – рост корней в направлении наибольшей концентрации питательных веществ.
5. Аэротропизм – Движение корней в сторону кислорода воздуха.
6. Гидротропизм – движение корней в сторону воды.
7. Тигмотропизм – ростовые движения на прикосновение.
8. Травмотропизм – изгибы растений, позволяющие изгибать повреждения растений об острые предметы.
Настии – обратимые движения органов (листьев, лепестков, чашелистиков) в ответ на диффузное действие температуры, влажности, света. Οʜᴎ подразделяются:
1. Фотонастии – раскрывание лепестков и листьев на солнце.
2. Термонастии – движения, связанные со сменой температуры.
3. Гидронастии – движения, связанные с изменением влажности.
4. Никтинастии – движение цветков и листьев при смене дня и ночи при комплексном воздействии температуры и света.
5. Сейсмонастии – движения листьев от толчка или прикосновения (листья мимозы).
6. Автонастии – ритмические движения листьев, не связанные с воздействием факторов среды.
Нутации – круговые или колебательные движения органов растений:
1. Круговые нутации у стеблей вьющихся растений (лиан), а также лазающих в поисках опоры
2. Медленные настические тургорные движения устьиц и фотонастии листьев.
Таксисы – перемещение всего организма в пространстве под влиянием односторонне действующих раздражителей (хемотаксис, фототаксис) – для одноклеточных организмов в воде.
referatwork.ru
|
Поиск Лекций
Сравнивая растительный и животный организмы, обычно говорят, что в отличие от животных растения не передвигаются, прикреплены в течение жизни к определенному месту. Однако движение как одно из свойств живой материи характерно и для растений. Движениями растений называют изменения расположения их органов в пространстве, обусловленные разными внешними факторами. Органы прикрепленных растений изменяют свое положение в пространстве благодаря росту, точнее, растяжению клеток и колебаниям тургорного давления. Ткани, непосредственно участвующие в движениях, имеют растяжимые и относительно тонкие клеточные стенки. К таким клеткам относятся, например, тонкостенные клетки основной паренхимы; клетки, находящиеся в основании черешков (подушечках) листьев бобовых растений; моторные клетки в эпидерме некоторых злаков, замыкающие клетки устьиц. Изменения тургорного давления происходят в результате транспирации, а также выхода воды из вакуоли в свободные пространства клеточных стенок и в межклетники. Однако нередко вместе с водой выходят и растворенные вещества. Следовательно, существует тесная связь между движениями и водным обменом растения и содержанием в его клетках минеральных веществ. В основе движений лежит универсальное свойство живой материи – раздражимость. В зависимости от типа раздражителя движения высших растений делят на две группы: тропизмы и настии. Многие раздражители действуют на растение направленно, с одной стороны. Растение способно различать направление действия света или вещества, силы тяжести. В ответ на одностороннее действие внешних факторов побеги или корни изгибаются. В результате изменяется расположение органов в пространстве. Эти изменения положения органов, вызываемые односторонне действующим внешним раздражителем, получили название тропизмов. В зависимости от природы раздражителя (свет, сила тяжести, химические вещества, прикосновение, электрический ток, поранение) тропизмы называют фото-, гео-, хемо-, тигмо-, электро-, травмотропизмами. Характер ответной реакции может быть разным. Те органы, которые поворачиваются к раздражителю, называют положительно тропными, а те, которые отворачиваются от раздражителя, – отрицательно тропными. Тропизмы чаще связаны с более быстрым ростом клеток на одной стороне стебля, корня или черешка, реже – с изменением тургорного давления. Геотропизмом называют движения растений в результате воздействия силы земного притяжения. Корень растения обладает положительным геотропизмом: – он растет вниз, а стебель – отрицательным, ибо растет в противоположном направлении. Весьма характерны в этом отношении проростки, помещенные в горизонтальном положении во влажную среду, или деревья, растущие на склонах. У первых корень вскоре будет изгибаться вниз, а стебель – вверх. Вторые будут ориентированы в направлении радиуса Земли, а не перпендикулярно поверхности склона. Ориентация к горизонтальной плоскости сразу же проявляется у прорастающего семени и у укореняющегося черенка. Корень углубляется в почву, где добывают воду и минеральные вещества, стебель же поднимается вверх, вынося листья навстречу падающим солнечным лучам.
Объяснения тому факту, что при геотропизме определяющим воздействием является сила земного притяжения, были получены еще в начале XIX в. с помощью простого, но остроумного приема. Проростки растений помещались на вращающееся колесо и увлажнялись водой. Растения росли таким образом, что стебли и корни их располагались по радиусам в плоскости вращения колеса: корни по направлению к периферии, а стебли – к центру, Т.е. первые росли по направлению центробежной силы, а стебли – в обратном направлении. Растения не знали ни верха, ни низа. Рост их определялся только направлением центробежной силы, заменившей силу земного притяжения. Наиболее чувствительными к силе тяжести являются кончик корня и зона роста стебля, а у злаков – верхушка колеоптиля. Первым, кто открыл место восприятия силы земного притяжения, был Ч. Дарвин. В его опытах корень с удаленной точкой роста утрачивал способность реагировать на эту силу и продолжал расти в том же положении, в котором он находился. Голландский ученый Ф. Вент и советский академик Н.Г. Холодный установили, что если к поверхности среза прикрепить отрезанный кончик корня или верхушку колеоптиля злаков, то тот же корень снова приобретает способность к проявлению геотропической чувствительности. Было высказано предположение, что дело здесь заключается в наличии в кончике корня или верхушке колеоптиля злаков фитогормонов. При вертикальном положении растений ауксины из верхушек перетекают в зону растяжения равномерно по всем сторонам стебля и корня. При горизонтальном положении растения ауксин накапливается на нижней стороне стебля и корня. Объяснить это можно тем, что нижняя сторона органа приобретает положительный заряд, а верхняя сторона - отрицательный. Ауксин обычно перемещается в виде аниона и поэтому концентрируется на нижней стороне. Стебель имеет гораздо более высокий оптимум концентрации ауксина (примерно 10-5моль/ л), чем корень (10-10-10-11 моль/л). Обладая различной чувствительностью к концентрации ауксина, корень и стебель, поэтому совершенно по-разному относятся к повышенной концентрации ауксина на нижней стороне горизонтально ориентированного органа. При накоплении ростового вещества на нижней стороне стебля рост ее ускоряется и вызывает изгиб кверху, а та же концентрация в нижней стороне корня приводит к противоположному эффекту: рост клеток ингибируется, и корень изгибается вниз. Изложенная теория Вента - Холодного объяснила вопрос, над решением которого трудились ученые в течение более ста лет. Очевидно, что в процессе длительной эволюции сохранялись лишь те особи, у которых под действием силы тяжести наблюдалась указанная реакция на неравномерное распределение ауксина. Ростовые движения растений, вызванные односторонним действием света, получили название фототропизма.Если изгиб направлен в сторону света, то говорят о положительном, а в обратную сторону – отрицательном фототропизме. Положительный фототропизм стеблей и колеоптилей объясняется тем, что сторона органа, обращенная к свету, растет медленнее, чем противоположная. Механизм фототропической реакции также связан с ауксинами: на затененной стороне ауксина больше, чем на освещенной. Поэтому растяжение клеток на затененной стороне происходит быстрее, чем на освещенной, и растение поворачивается в сторону большей освещенности.
Воспринимают одностороннее действие света верхушки надземных частей растений. Корни и корневища редко реагируют на этот фактор. Но у некоторых растений, например из семейства крестоцветных, корни проявляют иногда и отрицательный фототропизм. Если в специальную фототропическую камеру с небольшимотверстием в боковой стенке поместить проростки, например злаков, то уже через несколько часов верхушка колеоптиля повернется в сторону луча света, а изгиб произойдет в зоне растяжения. Если же, как показал впервые Ч. Дарвин, надеть на верхушку колеоптиля непроницаемый для света колпачок, то фототропическая реакция не проявляется. Фототропизм имеет колоссальное значение в жизни растений. Именно благодаря нему листья принимают положение, наиболее благоприятное для осуществления процесса фотосинтеза. Листья многих растений занимают более освещенные места, располагаясь так, чтобы не затенять друг друга. Это явление, которое называется листовой мозаикой, отчетливо наблюдается у клена, вяза, орешника, плюща и многих других растений. Фототропические движения листьев особенно важны в затененных условиях. В значительной мере нормальный рост растений в комнатах, цехах заводов и фабрик и других закрытых помещениях обязан фототропизму. Некоторые растения с такой силой реагируют на изменение направления света, что следуют за движением солнца на небосводе (гелиотропизм). К таким растениям относятся хорошо известные подсолнечник, череда и некоторые другие. Не все лучи спектра одинаковы в проявлении фотопериодических изгибов. Наиболее активными являются сине-фиолетовые лучи. Желтая, красная, ультрафиолетовая части спектра в этом отношении неактивны. Хемотропическиеростовые изгибы связаны с односторонним действием тех или иных химических веществ. Наиболее ярким примером хемотропизма является рост корней в сторону больших концентраций питательных веществ в почве. Большое значение хемотпропизм имеет и для нормальной жизнедеятельности грибов и высших растений, относящихся к сапрофитам и паразитам. Их гифы и присоски растут в сторону источника органических веществ. Явление хемотропизма наблюдается и при прорастании пыльцы и росте пыльцевой трубки. В данном случае, очевидно, основное влияние оказывает наличие выделяемых созревающими яйцеклетками веществ, которые управляют движением пыльцевых трубок. В числе этих веществ находятся некоторые специфические органические соединения, а также бор и кальций. Отрицательный хемотропизм корней наблюдается в случаях воздействия на них различных ядов. Движения корней растений в сторону кислорода воздуха получило название аэротропизма,а в сторону более оптимальной влажности почвы – гидротропизма.Первый можно наблюдать в избыточно увлажненных или сильно уплотненных почвах, второй при недостатке воды в почве. Эти виды тропизмов имеют важное экологическое значение, обеспечивая существование растений в стрессовых условиях, вызываемых недостатком кислорода или воды. Под тигмотропизмом подразумевают ростовую реакцию растений на прикосновение. Особенно сильно выражен тигмотропизм стеблевых и листовых усиков цепляющихся растений. Сторона усика при соприкосновении с опорой растет медленнее противоположной, что и приводит к обвиванию усика вокруг опоры. Эти движения происходят с затратой энергии. Предполагают участие в этих процессах сократительных белков и фитогормонов. Различают также травмотропизмы,позволяющие избегать соприкосновения с острыми предметами или химическими веществами, вызывающими травмы, радиотропизмы – ростовые изгибы и движения относительно источника радиоактивного излучения и некоторые другие. Настия – это движение органов растения, вызываемое раздражителем, не имеющим строгого направления, а действующим равномерно на все растение. Факторами, вызывающими настические движения, являются изменения температуры, освещенности, влажности воздуха в течение суток, перед дождем или после него. Если тропизмы – это движения органов в ответ на изменения напряженности внешнего фактора в пространстве, то настии – это движения органов, возникающие под действием смены условий во времени. Названия настии, как и тропизмов, зависят от природы раздражителей, которые их вызывают. Различают термо-, фото-, хемо-, гидро-, тигмо-, сейсмо-, электро- и травмонастии. К настическим движениям способны лишь двусторонне-симметричные органы (листья, лепестки). Примером термонастииможет служить раскрывание цветков тюльпана при высокой температуре и закрывание при низкой. К фотонастическим движениям относятся раскрывание цветков кувшинки и соцветий одуванчика при хорошем освещении и закрывание при уменьшении количества света, поднимание листовых пластинок кислицы на свету и опускание в темноте. Наиболее распространенными являются никтинастические движения, т.е. вызываемые сменой дня и ночи, когда одновременно меняются условия температуры и освещения. Очень многие цветки открываются утром и закрываются на ночь или наоборот. Раздражителем, вызывающим никтинастическую реакцию, может быть изменение температуры или только интенсивности света, или одновременно температуры и интенсивности света. Никтинастические движения облегчают опыление цветков в благоприятную погоду и защищают внутренние органы цветка при неблагоприятных условиях. Разновидностью настических изгибов являются сеисмонастии, вызываемые прикосновением или сотрясением, например листьев стыдливой мимозы: клетки одной стороны сочленения теряют клеточный сок с последующим мгновенным падением тургора, сжиманием клеток и опусканием черешков и листочков сложных листьев. Передается возбуждение при помощи электрического импульса или особых гормонов.
Механизм настий до конца не изучен. Предполагают, что настии происходят благодаря неравномерному росту разных сторон лепестков или листочков околоцветника. Если быстрее растет верхняя сторона, то происходит эпинастия, в результате которой раскрываются цветки, почки, опускаются листья. Если быстрее растет нижняя сторона, то происходит гипонастия, в результате которой органы закрываются. Открывания и закрывания цветков могут повторяться в определенном ритме. При каждом новом открывании и закрывании происходит рост клеток соответственно верхней или нижней стороны. Если процессы повторяются много раз, то лепестки или листочки околоцветника значительно удлиняются. Неравномерный рост, возможно, обусловлен гормонами. Это доказывается с помощью опыта. Если срезать верхушку молодого растения томата и на поверхность среза нанести пасту с ИУК, то через некоторое время все оставшиеся листья опустятся (эпинастия). В основе сейсмонастий, а также изменения положения листьев при переходе от дня к ночи лежит быстрое изменение тургорного давления в клетках на различных сторонах органа. Изменения тургора в моторных клетках, расположенных на разных сторонах подушечек, вызываются увеличением или уменьшением концентрации в них ионов калия. Калий перемещается из сокращающихся клеток в растягивающиеся благодаря работе калиевых насосов. За ним поступает вода, и клетки набухают. Центральная вакуоль при этом распадается на множество мелких вакуолей. Тургорные движения обратимы. Лучше изучен механизм опускания листьев у стыдливой мимозы, у которой различают два механизма проведения возбуждения: химический и электрический. Химический механизм состоит в том, что раздраженные клетки выделяют вещество, передвигающееся по флоэме и паренхиме. Когда это вещество дойдет до подушечки, происходит ответная реакция (листочек складывается). Реакцию вызывает очень низкая концентрация этого вещества 10-8 г/мл. Предполагают, что этими веществами являются аминокислоты, например, глутамат и аланин или малат. При электрическом механизме проведения возбуждения сразу после прикосновения в ее клетках возникает потенциал действия, равный -140 мВ (потенциал покоя равен -160 мВ). Потенциал действия распространяется от клетки к клетке по проводящим пучкам со скоростью 2-5 см/с. Он передается в подушечки у основания листочков и всего листа, в результате в моторных клетках уменьшается тургорное давление (вода выходит в межклетники), и лист складывается. Изменение тургорного давления зависит от ионов калия и хлора, концентрация которых в клетке регулируется ионными насосами. Возбуждение моторных клеток приводит к увеличению проницаемости плазмалеммы, ионы хлора и калия выходят, одновременно резко падает тургорное давление. Возможно, что в выделении воды участвуют и сократительные белки. На это указывает высокое содержание АТФ в моторных клетках, сильно уменьшающееся во время движения. |
|


poisk-ru.ru
I Движения растений
Количество просмотров публикации I Движения растений - 161
АДАПТАЦИЯ И УСТОЙЧИВОСТЬ РАСТЕНИЙ К НЕБЛАГОПРИЯТНЫМ ФАКТОРАМ
I Движения растений
II Адаптация и устойчивость растений к неблагоприятным факторам
1. Стресс, адаптация, устойчивость
2. Неспецифическая реакция растений на стресс.
3. Засухоустойчивость и устойчивость к перегреву.
4. Устойчивость растений к низким температурам.
5. Солеустойчивость.
6. Устойчивость к недостатку кислорода.
7. Газоустойчивость.
8. Радиоустойчивость.
9. Устойчивость растений к тяжелым металлам.
10. Устойчивость растений к патогенам.
Выделяют следующие способы движения у растений:
1.Движения цитоплазмы и органоидов.
2. Локомоторные движения с помощью жгутиков (таксисы).
3. Верхушечный рост (корневые волоски, пыльцевые трубки).
4. Ростовые движения (нутации, тропизмы, ростовые настии).
5. Тургорные движения (движения устьиц, настии, сейсмонастии).
Из всех перечисленных форм движения 3-5 типы свойственны только растениям.
Движение цитоплазмы осуществляется за счёт взаимодействия актиновых микрофилламентов эндоплазмой, находящейся в состоянии золя и микротрубочками гелеобразной эктоплазмы. Этот процесс является энергозависимым и нуждается в присутствии ионов Са2+ в концентрациях менее 0,1 мкмоль/л. Различают колебательное, циркуляционное, ротационное и фонтанирующее движения цитоплазмы. Средняя скорость движения цитоплазмы у разных клеток варьирует от 2-5 мкм/с до 1350.
В корне локомоторных способов движения клеток лежит функционирование систем сократительных белков, обеспечивающих превращение химической энергии АТФ в механическую энергию, к примеру, движения жгутиков. Перемещение всего организма в пространстве под действием односторонне действующих факторов (раздражителей) принято называть таксисом. Этот тип движения характерен для одноклеточных жгутиковых водорослей, для гамет и зооспор водорослей, для сперматозоидов мхов, плаунов, хвощей и папоротников. В корне движения жгутика лежит функционирование белковой системы тубулин-динеин, которая за счёт энергии АТФ обеспечивает механическое движение этого органа.
Перемещение в пространстве корневых волосков, пыльцевых трубок, гиф грибов, протонемы мхов осуществляется за счёт верхушечного роста. Это высокополяризованный тип роста͵ при котором в клетках соблюдается строгая пространственная локализация интенсивности синтеза вдоль продольной оси клетки. Секреторные везикулы транспортируются к кончику клетки, куда доставляют материал для синтеза стенки, а также синтазы и литические ферменты, которые разрыхляют структуру стенки, делая ее неспособной противостоять высокому тургорному давлению. Микрофибриллы стенки раздвигаются и новые компоненты встраиваются в разрыхленную стенку.
Тропизмы - ϶ᴛᴏ ростовые движения растений, обусловленные изгибанием или искривлением органов в ответ на факторы среды, действующие односторонне. Οʜᴎ реализуются в растущих частях растений и, как правило, являются следствием более быстрого роста клеток растяжением на одной стороне побега, корня или листа. Различают геотропизм (ориентация в пространстве и изгибание под действием гравитационного поля), фототропизм (ростовые изгибы под действием одностороннего освещения), хемотропизм (ростовая реакция на градиент химических соединений), тигмотропизм (ростовые изгибы в ответ на прикосновение, к примеру, у усиков некоторых лазящих растений), травмотропизм (изгиб в сторону от нанесенного повреждения).
Фототропизм. Согласно гипотезе Н.Г.Холодного- Ф.Вента фототропический стимул вызывает поперечную электрическую поляризацию клеток в верхушке колеоптиля, причем неосвещенная сторона приобретает положительный заряд, что приводит к перераспределению ауксина. ИУК передвигается на затененную сторону, благодаря чему рост этой стороны усиливается. С помощью подобного механизма затененные участки листовых пластинок выходят из тени, создавая эффект ʼʼмозаики листьевʼʼ.
Настии – обратимые движения (изгибы) органов с дорсивентральным строением в ответ на изменение диффузно действующих факторов внешней среды (к примеру, открывает и закрывает цветков при смене дня и ночи). Направленность настических движений зависит от структуры самого органа. При более быстром росте верхней стороны орган (лист, лепесток) изгибается книзу (эпинастия), при обратном соотношении скоростей роста– кверху (гипонастия). Различают фото-, термо-, гидро-, хемо-, тигмо-, сейсмо-, травмо- и электронастии. Настические движения обеспечивают защиту органов (закрывание цветков, опускание листьев) или захват предметов (движение усиков, железистых волосков росянки).
Нутации – круговые или колебательные движения органов растений. Так, растущий побег совершает качания, благодаря чему его верхушка колеблется относительно продольной оси. Особенно хорошо круговые нутации выражены у стеблей вьющихся растений (лиан). Длительность одного оборота у этих растений – от 2 до12 ч. Молодой побег вьющегося растения сначала растет вертикально, затем его верхушка изгибается и занимает горизонтальное положение. Вслед за этим наблюдаются круговые нутации и побег обвивается вокруг опоры. У многих лазающих растений круговые нутации совершает вся верхняя часть побега. Круговые нутации лиан и лазящих растений служат для поиска опоры, крайне важно й для движения к свету.
Обратимые тургорные движения также обусловлены дорсивентральным строением органов и вызываются теми же диффузно действующими факторами внешней среды. При этом они не зависят от процессов роста и обуславливают двигательную активность тех органов, которые закончили свой рост. Такие движения реализуются благодаря увеличению и уменьшению в вакуолях специализированных клеток концентрации осмотически активных веществ (К+, малата), благодаря чему соответственно увеличиваются или уменьшаются поглощение воды и тургорное давление. К примеру, движения устьиц или опускание листьев на ночь (никтинастии). К медленным тургорным движениям относят также выделение железистыми волосками росянки муравьиной кислоты в ответ на прикосновение шершавых предметов.
К быстрым тургорным движениям относятся сейсмонастические (индуцируемые сотрясением) движения сложных листьев некоторых бобовых (мимозы, клевера), кислицы и др. Размещено на реф.рфВ ответ на механический удар или встряхивание нижние клетки главного сочленение мимозы способны быстро ( в течение секунд) терять тургор, что приводит к опусканию листа. Поднимается лист медленнее (30-60 мин). Раздражение вызывает возникновение в листе потенциала действия с амплитудой 50-100 мВ, который распространяется со скоростью 0,5 – 4 см/с. Физиологическое значение опускания листьев у мимозы при сотрясении, по-видимому, связано с предотвращением повреждений от сильных ливней и ураганных ветров.
referatwork.ru









