
Дополнительный материал по биологии на тему "Вакуоль". Содержимое вакуолей клеток растений
17. Вакуоли. Строение, химический состав и функции. Лизосомы и периксомы.
Лизосомы представляют собой неотъемлемую часть состава клетки. Они являются разновидностью везикул. Эти клеточные помощники, являясь частью вакуома, покрыты оболочкой из мембраны и наполнены гидролитическими ферментами. Важность существования лизосом внутри клетки обеспечена секреторной функцией, которая необходима в процессе фагоцитоза и аутофагоцитоза.
Лизосомы существуют как в животных и растительных клетках, так и в клетках грибов. Строение и функции лизосом могут отличаться. Они могут иметь разные размеры, форму, особенности структуры и химического состава. В обобщенном варианте лизосомы – это мембранные пузырьки, образованные из везикул и эндосом, с кислотной средой (PH от 1,5 до 4-4,5).
Отличительной чертой строения и функций лизосом, помимо кислой реакции внутренней среды, от других внутриклеточных органелл является наличие белковых ферментов. Ферменты участвуют в процессе распада полимеров, имеющихся в организме, а именно белков, углеводов, жиров и нуклеиновых кислот. Размеры лизосом колеблются от 0.3 до 0.5 мкм. в зависимости от обитания.
Функции лизосом
Такой большой набор ферментов внутри микроскопического пузырька обусловлен рядом важных функций лизосом, выполняемых в организме. У живых организмов, имеющих внутриклеточное пищеварение, лизосомы принимают участие в переваривании питательных веществ, поступивших в клетку в процессе эндоцитоза. При эндоцитоце клетка захватывает не только питательные вещества, но и бактерии и другие вещества, попадающие в организм. Ферменты лизосом переваривают захваченные частицы, сливаясь с вакуолями.
Ферменты лизосом могут оказывать благотворное воздействие на клетку и организм в целом. Лизосомы переваривают не только необходимые вещества, но и уничтожают вредные, ненужные клетке элементы (участвуют в обновлении клеток). Ферменты лизосомы расщепляют и произведенные самой клеткой белковые соединения. Этот процесс назван биологами аутофагией. При автолизе лизосомы самоуничтожают клетки, которые могут оказатьпагубно влияние на организм. Этот процесс происходит при развитии организма и дифференцировании клеток, отвечающих за специализированные процессы.
Также, к функциям лизосом можно отнести их участвуют в обмене веществ. Переваренные лизосомами вещества поступают в межклеточную жидкость или плазму крови и вовлекаются в обмен веществ. В растительных организмах лизосомы способны накапливать ионы, пигменты, белки и вторичные метаболиты. Нарушение работы лизосом может оказать влияние на работу всего организма. При сбоях могут быть вызваны процессы накопления и развитие сложных заболеваний. При разрыве лизосом в гиалоплазму попадают расщепляющие ферменты. Подобное происходит при действии излучения или при некрозе, сопровождаясь повышением активности ферментов лопнувших лизосом.Вакуоль — ограниченный мембраной органоид, содержащийся в некоторых эукариотических клетках и выполняющий различные функции (секреция, экскреция и хранение запасных веществ, аутофагия, автолиз и др.) . Вакуоли и их содержимое рассматриваются как обособленный от цитоплазмы компартмент. Вакуоли особенно хорошо заметны в клетках растений: во многих зрелых клетках растений они составляют более половины объёма клетки. Одна из важных функций растительных вакуолей — накопление ионов и поддержание тургора (тургорного давления) . Вакуоль — это место запаса воды.
Мембрана, в которую заключена вакуоль, называется тонопласт. Вакуоль в растительной клетке одна и занимает довольно большой объем, в то время как вакуолей в животной клетке несколько, и каждая имеет свои функции: пищеварительные, фагоцитозные, сократительные. Вакуоли образуются из пузырьков аппарата Гольджи и могут быть сократительными (которые служат для осмотической регуляции) и центральными.
Вакуоль состоит из полости в цитоплазме, которая может достигать до 90% от общего объема всей клетки, из вакуолярной мембраны, которая ограничена от этой полости тонопластом и заполненна клеточным соком (концентрированный водный раствор минеральных солей, сахаров, органических кислот, пигментов и отходов жизнедеятельности клетки).
Вакуоль выполняет такие функции, как обеспечение роста растительной клетки за счет растяжения, происходящего в результате поступления в нее воды. Вакуоль также регулирует напряженное состояние клеточной стенки, которое происходит из-за давления внутриклеточной жидкости.
Содержащиеся в вакуоли гидролитические ферменты способствуют повторное использование таких компонентов, как хлоропласт. Также в вакуоли скапливаются отходы жизнедеятельности клетки: млечный сок, танины и алкалоиды. Эти вещества делают растение несъедобным.
Помимо отходов в вакуоли также хранятся питательные вещества, которые используются цитоплазмой клетки при необходимости: это инсулин, сахароза и минеральные соли.
studfiles.net
Дополнительный материал по биологии на тему "Вакуоль"

Выполнила учитель биологии
Тушева Вера Ивановна
Содержание стр
Цель работы……………………………………………………………………..3
Образование вакуолей………………………………………………………….4
Строение вакуолей и химический состав клеточного сока……….................5
Функции вакуолей……………………………………………………………...7
Вывод………………………….……………………………………………………8
Список используемой литературы…….………………………………………….9
Вакуоль – это пространство клетки, заполненное клеточным соком.
Целями работы является:
знакомство с процессом образования вакуоли и с основными ее функциями;
изучение строения вакуоли и химическим составом ее клеточного сока;
определение важности вакуоли в растительной и животной клетке;
создание теоретической базы для дальнейшего изучения предмета «Ботаника», необходимой на семинарских и лабораторных занятиях.
Образование вакуолей
Центральная вакуоль, характерна для большинства зрелых растительных клеток (рис 1), возникает в процессе роста и дифференциации клеток путем слияния многочисленных мелких вакуолей, которые обычно имеются в меристематических (эмбриональных) клетках. Происхождение этих цитоплазматических вакуолей до конца не выяснено. Одним из наиболее вероятных способов заложения вакуолей является образование цистернами гранулярного эндоплазматического ретикулума локальных расширений, мембраны которых теряют рибосомы. Эти расширения затем изолируются, округляются и превращаются в вакуоли, а ретикулярная мембрана становиться их тонопластом. Другой важный путь – отчленения ретикулярными цистернами агранулярных пузырьков., которые, сливаясь друг с другом, дают начало мелким вакуолям. Образование вакуолей может происходить и при участии элементов аппарата Гольджи (пузырьков и диктиосомных цистерн).
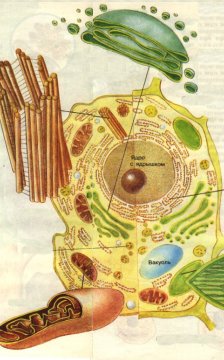
Рис. 1. Строение клетки:
1 – ядро с ядрышком; 2 – вакуоль; 3 – хлоропласт; 4 – эндоплазматическая сеть с рибосомами; 5 – аппарат Гольджи; 6 – лизосома; 7 - митохондрия
Строение вакуолей и химический состав клеточного сока.
Вакуоли содержаться почти во всех растительных клетках. Они представляют собой полости в клетке, заполненные обычно водянистым содержанием – клеточным соком. От цитоплазмы клеточный сок изолирован избирательно проницаемой вакуолярной мембраной – тонопластом (лат. tonus – напряжение; пластос - оформленный). Для большинства зрелых клеток высших растений характерна центральная вакуоль (рис. 2). Она настолько крупна (занимает до 70-90% объема клетки), что протопласт со всеми органеллами располагается в виде очень тонкого постенного слоя, выстилающего клеточную оболочку.
Рис. 2. Клетка палисадного мезофилла листа тополя под световым микроскопом (схемат.):
Г – гиалоплазма; Гр – граны хлоропласта; КЗ – крахмальное зерно; КО – клеточная оболочка; ЛК – липидная капля; Хл – хлоропласт; ЦВ – центральная вакуоль; Я – ядро; Яд – ядрышко.
В этом слое тонопласт выступает как внутренняя пограничная мембрана протопласта. В местах протопласта часто располагается очень близко к плазмалемме, а на участках, где они есть, тонопласт отдаляется от плазмалеммы, но при этом общая толщина слоя гиалоплазмы, одевающего органеллы, не увеличиваются. В постенном протопласте обычно встречаются мелкие цитоплазматические вакуоли (рис. 3).
Иногда в центре клетки, в ядерном кармашке, располагается ядро. А кармашек связан с постенным слоем цитоплазмы тончайшими цитоплазматическими тяжами, пересекающими центральную вакуоль.
Вакуолярное содержимое – клеточный сок – представляет собой водный раствор различных веществ, являющихся продуктами жизнедеятельности протопласта (в основном запасными веществами и отбросами). Таким образом, основной компонент клеточного сока – вода. В ней накапливаются многочисленные соединения – минеральные и органические. – которые находятся в состоянии истинного или коллоидного раствора и реже в виде оформленных включений.
Рис. 3. клетка мезофилла листа липы при небольшом (5000) увеличении электронного микроскопа (схемат.):
В – цитоплазматическая вакуоль; Г – гиалоплазма; Гр – грана; ГЭР – цистерна гранулярного эндоплазматического ретикулума; Д – диктиосома; КЗ – крахмальное зерно; КО – клеточная оболочка; ЛК – липидная капля; М – митохондрия; Мж – межклетник; Мк – микротельце; Пд – плазмодесма; Пг – пластоглобула; Пл – плазмолемма; По – полисома; ПЯ – пора в ядерной оболочке; СП – срединная пластинка; Т – тонопласт; Ти – межгранный тилакоид; Хл – хлоропласт; Хм – хроматин; ЦВ – центральная вакуоль; Я – ядро; Яд – ядрышко; ЯО – ядерная оболочка.
Реакция клеточного сока обычно слабокислая или нейтральная, реже щелочная. По химическому составу и консистенции веществ клеточный сок отличается от протопласта. Это различие обусловлено активностью вакуолярной мембраной, обладающей свойствами избирательной проницаемости и, следовательно, не пропускающей одни вещества и транспортирующие другие вещества в вакуоль против градиента концентрации. Поэтому основные функции тонопласта, как и плазмалеммы, не синтетические, а барьерные и транспортные.
В живой клетке клеточный сок обычно не имеет никакой внутренней структуры, т.е. является оптически пустым, чем и обусловлено название вакуоли. Однако ряд веществ клеточного сока реагирует с фиксаторами и красителями, поэтому на фиксированных препаратах в нем может выявляться определенная структура.
Вещества, входящие в состав клеточного сока, чрезвычайно разнообразные – это углеводы (сахара и полисахариды), белки, органические кислоты и их соли, аминокислоты, минеральные ионы, алкалоиды, гликозиды, пигменты, танины и другие растворимые и воде соединения. Большинство из них относятся к группе эргастических веществ – продуктов метаболизма протопласта, которые могут появляться и исчезать в различные периоды жизни клетки. Многие вещества клеточного сока образуются только в растительных клетках.
Химический состав и концентрация клеточного ока изменчивы и зависят от вида растения, органа, типа и состояния клетки. Некоторые из перечисленных выше классов соединений могут в определенных клетках вообще отсутствовать. Другие накапливаются в больших количествах.
Наиболее обычными веществами клеточного сока являются сахара, прежде всего сахароза, также глюкоза и фруктоза. Они играют роль запасных энергетических веществ и служат важнейшим питательным материалом клетки. Сахароза, накапливающаяся в большом количестве в клеточном соке корнеплодов сахарной свеклы и сердцевины стеблей сахарного тростника. Имеет большое народнохозяйственное значение. Так как служит источником получения сахара. Глюкоза (или виноградный сахар) и фруктоза. Как показывают их названия. Накапливаются в больших количествах главным образом в сочных плодах и также широко используются человеком. Для ряда групп растений (кактусовые, толстянковые, орхидные) характерно накопление в клеточном соке полисахаридов в виде слизей.
Клетки созревающих семян накапливают в вакуолях в виде коллоидного раствора большое количество белков, поэтому их называют белковыми вакуолями. Синтез этих белков происходит на прикрепленных рибосомах гранулярного эндоплазматического ретикулума, с которым тесно связано развитие белковых вакуолей. При обезвоживании семян на поздних этапах их развития из вакуолей удаляется вода. Концентрация белка в клеточном соке повышается и он переходит в состояние твердого геля. Дегидратированные вакуоли зрелых семян называют белковыми тельцами или алейроновыми зернами. Белковые вакуоли встречаются не только в клетках семян, но и в ряде других клеток растений.
Из органических кислот в клеточном соке наиболее часто встречаются лимонная, яблочная, янтарная и щавелевая. Эти кислоты находятся в большом количестве, например, в клеточном соке незрелых плодов, придавая им кислый вкус. При созревании плодов органические кислоты могут использоваться как субстраты дыхания, поэтому кислый вкус плодов обычно исчезает. Соли органических кислот вместе с минеральными ионами играют наибольшую роль в осмотических процессах в клетке.
В состав клеточного сока часто входят дубильные вещества-танины. Это безазотистые циклические соединение (производный фенолов) вяжущего вкуса. Для некоторых клеток высших растений накопление танинов становиться одной из основных функций. Содержимое центральной вакуоли таких клеток в прижизненном состоянии имеет зеленовато-желтый цвет, а после фиксации с применением осмия или окраски хлоридом железа оно выглядит темным. Таниноносные клетки могут быть рассеяны в беспорядке поодиночке среди других клеток данной ткани или собраны в группы. Особенно богаты танинами клетки коры стеблей и корней (дуб, ива, ель, бадан), незрелых плодов (грецкий орех), листьев (чай), и некоторых патологических выростов (например, клетки так называемых чернильных орешков на листьях дуба). При отмирании клетки танины окисляются, пропитывают клеточную оболочку и придают ей темно-коричневый цвет. Эти вещества обладают антисептическими свойствами и поэтому защищают растение от инфекции. Техническое значение танинов состоит в том, что с их помощью дубят кожу. При этом составляющие кожу белки переводятся в нерастворимое состояние и перестают набухать; кожа становится мягкой, не ослизняющейся в воде.
Алкалоиды – разнообразные в химическом отношении азотсодержащие вещества гетероциклической природы, имеющие горький вкус. Они обладают щелочными свойствами и содержатся в клеточном соке, как правило. В виде солей. Они обычно бесцветны, редко окрашены (оранжевый цвет млечного сока чистотела). Алкалоиды характерны для клеток высших растений и у других организмов встречаются редко. В настоящее время описано свыше 2000 алкалоидов. Состав их часто характерен для определенных групп растений (вида, рода). К алкалоидам принадлежат многие растительные яды. Некоторые из них, такие, как кофеин (в семенах кофе), атропин (во всех органах белладонны), хинин (в коре хинного дерева), морфин, кодеин (в плодах мака) и др., широко используются в медицине как лекарственные средства. Млечный сок, вытекающий при ранении ряда растений, представляет собой содержимое вакуолей, часто богатое алкалоидами (мак, чистотел) или каучуком (гевея, одуванчик).
Гликозиды – обширная группа природных веществ, соединения сахаров со спиртами, альдегидами, фенолами и другими веществами. Ряд гликозидов растений используется в медицине (например, сердечные гликозиды, добываемые из наперстки и ландыша). К гликозидам принадлежат также пигменты клеточного сока – флавоноиды. Одни из них – антоцианы (греч. антос – цветок; кианос - синий) – придают клеточному соку красный, синий или фиолетовый цвет; другие – флавоны – желтый. Цветовая гамма (от фиолетового до красного) окрашенных частей цветков многих растений обусловлена присутствием в их клеточном соке антоцианов. Различие в оттенках цвета связано с различной реакцией клеточного сока: если она кислая, то господствуют красные тона, нейтральная – фиолетовые, а при слабощелочной реакции – синие. На возникновение оттенков оказывает влияние также образование антоцианами комплексов с различными металлами. Флавоны обусловливают желтый цвет лепестков ряда растений (льнянка, коровяка, примулы, многих бобовых и сложноцветных). Яркая окраска цветков, вызываемая флавоноидами клеточного сока или каротиноидами пластоглобул хромопластов, выполняет функцию привлечения насекомых-опылителей.
Значение органических кислот, танинов, алкалоидов и гликозидов клеточного сока в обмене веществ клетки выяснено еще недостаточно. Раньше их обычно рассматривали как конечные продукты обмена. В настоящее время показано, что многие из них могут вновь вовлекаться в процессы метаболизма и поэтому их можно рассматривать и как запасающие вещества.
Функции вакуолей.
Вакуоли в растительных клетках выполняют две основные функции – накопление запасных веществ и отбросов и поддержание тургора. Первая функция ясна из приведенного выше описания химического состава клеточного сока. Вторая функция вакуолей требует пояснения. Концентрация ионов и сахаров в клеточном соке центральной вакуоли. Как правило. Выше, чем в оболочке клетки; тонопласт значительно замедляет диффузию из вакуоли этих веществ и в то же время легко проницаем для воды. Поэтому при достаточном насыщении оболочки водой последняя будет поступать в вакуоль путем диффузии. Такой однонаправленный процесс диффузии воды через избирательно проницаемую для растворенных веществ мембрану носит название осмоса. Поступающая в клеточный сок вода оказывает давление на постенный протопласт, а через него и на оболочку, вызывая напряженное, упругое ее состояние или тургор (от латинского - набухать) клетки.
Тургор обеспечивает сохранение сочными органами (например, листьями, неодревесневшими стеблями) формы и положения в пространстве, а также сопротивление их к действию механических факторов. Если клетку погрузить в гипертонический раствор какой-нибудь соли или сахара (т.е. в раствор большей концентрации, чем концентрация клеточного сока). То начинается осмотический выход воды из вакуоли. В результате этого объем ее сокращается, эластичный постенный протопласт отходит от оболочки по направлению к центру клетки. Тургор исчезает, наступает плазмолиз клетки.
Плазмолиз обычно обратим, и при доступе воды или переносе клеток в гипотонический раствор вода снова энергично поглощается центральной вакуолью, протопласт опять прижимается к оболочке. Тургор восстанавливается.
Плазмолиз может служить показателем живого состояния вакуолизированной клетки, ибо отмершая клетка не плазмолизируется, так как не содержит избирательно проницаемых мембран, а в живой, как уже говорилось, слой постенной цитоплазмы может быть так тонок, что его невозможно увидеть в световой микроскоп; при отхождении от оболочки он становиться различимым.
Потеря тургора при плазмолизе вызывает завядание растения. При завядании на воздухе в условиях недостаточного водоснабжения тонкие оболочки клеток сморщиваются одновременно с протопластом и делаются складчатыми.
Тургорное давление не только поддерживает форму неодревесневших частей растений, оно является также одним из факторов роста клетки, обеспечивая рост клеток растяжением. У животных клеток центральная вакуоль отсутствует, из-за чего они растяжением не растут, увеличение их размера происходит главным образом за счет увеличения количества цитоплазмы, поэтому размер животных клеток обычно меньше, чем растительных.
Выводы
Вакуоль характерна для большинства растительных клеток.
Центральная вакуоль возникает в процессе роста и дифференциации клеток путем слияния многочисленных мелких вакуолей.
Образование вакуолей может происходить при участии аппарата Гольджи.
Клеточный сок - водный раствор различных веществ, являющихся продуктами жизнедеятельности протопласта (в основном запасными веществами и отбросами).
Тургорное давление не только поддерживает форму неодревесневших частей растений, оно является также одним из факторов роста клетки, обеспечивая рост клеток растяжением.
Список используемой литературы.
: Васильев А.Е, Воронин Н.С, Еленевский и др. Ботаника: морфология и анатомия растений – М.: Просвещение, 1988. 480 с.
Биология. Живой организм (учебник для общеобразовательных школ) – Алматы: Кiтап, 2006. – 208 с.
infourok.ru
|
ТОП 10: |
Клетки как низших, так и высших растительных организмов содержат в цитоплазме вакуоли, несущие ряд важных физиологических нагрузок (рис. 195). У молодых клеток может быть несколько мелких вакуолей, которые по мере роста и дифференцировки клетки сливаются друг с другом и образуют одну или несколько крупных вакуолей, занимающих до 90% объема всей клетки. Центральные вакуоли отделены от цитоплазмы одинарной мембраной, сходной по толщине с плазмалеммой. Мембрана, ограничивающая центральные вакуоли, носит название тонопласта. Возникают центральные вакуоли из мелких пузырьков, отщепившихся от аппарата Гольджи. Такие первичные вакуоли растут в объеме, сливаются друг с другом и в конце концов образуют одну или несколько крупных вакуолей, оттесняющих цитоплазму с ядром и органоидами к периферии клетки. Полость вакуоли заполнена так называемым клеточным соком, представляющим собой водный раствор, в который входят различные неорганические соли, сахара, органические кислоты и их соли и другие низкомолекулярные соединения, а также некоторые высокомолекулярные вещества (например, белки). Центральные вакуоли растений выполняют многообразные и важные функции. Одной из главных ее функций является поддержание тургорного давления клеток. Растворенные в соке вакуолей молекулы определяют его осмотическую концентрацию. Соответствующая молекулярная концентрация сока вакуолей и полупроницаемые свойства как ее мембраны, тонопласта, так и плазмалеммы способствуют тому, что вакуоль функционирует в качестве осмометра и придает клетке необходимую прочность и тургисцентность (напряженность). Другая функция определяется тем, что вакуоль представляет собой большую полость, отделенную от метаболирующей гиалоплазмы мембраной, тонопластом, обладающим свойствами полупроницаемости и через котрый может происходить, как и через плазматическую мембрану, активный транспорт различных молекул. В тонопласте обнаружен АТФ-зависимый Н+-насос, направленный внутрь вакуолей, участвующий в транспорте сахаров. Поэтому вакуоли могут использоваться клетками как накопительные резервуары не только для отложения запасных веществ, но и для выброса метаболитов, для экскреции. Так выводятся, секретируются из клетки все водорастворимые метаболиты. Нерастворимые в воде органические компоненты могут превращаться в растворимые глюкозиды, соединяясь с молекулами сахаров. Перечень экскретируемых в вакуоли метаболитов очень обширен. Это различные алкалоиды (например, никотин, кофеин) и полифенолы. В вакуолях происходит отложение многих глюкозидов, к которым относятся различные пигменты, например антоцианы. Из неорганических веществ в вакуолярном соке накапливаются фосфаты калия, натрия, кальция, могут накапливаться соли органических кислот (оксалаты, цитраты и др.). Это придает вакуолярному соку отчетливую кислую реакцию (рН от 2 до 5). Таким образом, можно считать, что тонопласт участвует в процессах экскреции. Другой обширный ряд функций вакуолей связан с накоплением запасных веществ, таких, как сахара и белки. Сахара в вакуолях содержатся в виде растворов, встречаются и резервные полисахариды типа инулина. В вакуолях происходит запасание белков, что характерно для семян. Поступление белков в вакуоли, вероятнее всего, связано со способностью вакуолей ЭР и АГ сливаться с тонопластом. Запасание белков семян злаковых происходит в так называемых алейроновых вакуолях, которые заполняются альбуминами и глобулинами, после чего вакуоли обезвоживаются, превращаясь в твердые алейроновые зерна. При прорастании семян эти зерна обводняются и снова превращаются в вакуоли. В таких новообразованных вакуолях выявляется активность некоторых ферментов, кислой фосфатазы, a-амилазы, глюкозидазы, протеиназы и РНКазы. Следовательно, алейроновые вакуоли отчасти напоминают лизосомы, где происходит переваривание запасных белков при прорастании семян. Гидролитические ферменты были обнаружены не только в алейроновых вакуолях, но и в мелких и крупных центральных вакуолях. Наблюдалась неоднократно инвагинация, впячивание тонопласта внутрь вакуолей, при этом часть “втянутого” материала оказывается в полости вакуоли и там деградирует. Возможно, так выполняется аутофагическая функция вакуолей, участвующих в гидролизе дефектных клеточных компонентов. Лизосомными свойствами обладают вакуоли дрожжей. Было обнаружено, что стенки вакуолей дрожжей тоже могут образовывать впячивания внутрь, затем они отщепляются от тонопласта и растворяются внутри вакуоли. Сферосомы Это мембранные пузырьки, встречающиеся в клетках растений, они окрашиваются липофильными красителями, имеют высокий коэффициент преломления и поэтому хорошо видны в световой микроскоп. Сферосомы образуются из элементов эндоплазматического ретикулума. На конце цистерны ЭР начинает накапливаться осмиофильный материал, затем от этого участка отшнуровывается и начинает расти мелкий пузырек, достигающий диаметра 0,1-0,5 мкм. Это “просферосома”, окруженная одинарной мембраной. Рост сферосом и перестройка их содержимого связаны с накоплением в них масла, так что сферосома постепенно превращается в масляную каплю. Отложение липидов начинается между осмиофильными слоями мембраны. Кроме жиров в составе сферосом обнаруживают белки и среди них фермент липазу, расщепляющую липиды. Пероксисомы (микротельца) Это небольшие вакуоли (0,3-1,5 мкм), одетые одинарной мембраной, отграничивающей гранулярный матрикс, в центре которого располагается сердцевина, или нуклеоид (ничего не имеющий общего с нуклеоидом бактерий и вобще к ядерным структурам не относящийся). В зоне сердцевины часто, особенно в пероксисомах печеночных клеток, видны кристаллоподобные структуры, состоящие из регулярно упакованных фибрилл или трубочек. Изолированные сердцевины пероксисом содержат фермент уратоксидазу (рис. 196, 207б). Пероксисомы обнаружены у простейших (амебы, тетрахимена), у низших грибов (дрожжи), у высших растений в некоторых эмбриональных тканях (эндосперм) и в зеленых частях, способных к фотореспирации, у высших позвоночных животных они обнаруживаются главным образом в печени и почках. В печени крыс на клетку число пероксисом колеблется от 70 до 100. Пероксисомы часто локализуются вблизи мембран ЭР. У зеленых растений пероксисомы часто находятся в тесном контакте с митохондриями и пластидами. Впервые пероксисомы были выделены из печени и почек. Во фракциях пероксисом обнаруживается ферменты, связанные с метаболизмом перекиси водорода. Это ферменты (оксидазы, уратоксидаза, оксидаза d-аминокислот) окислительного дезаминирования аминокислот, при работе которых образуется перекись водорода (Н2О2 ) и каталаза, разрушающая ее. В пероксисомах печени каталаза составляет до 40 % всех белков и локализована в матриксе. Так как Н2О2 является токсическим веществом для клеток, то каталаза пероксисом может играть важную защитную роль. Пероксисомы цыплят и лягушек кроме уратоксидазы содержат ряд ферментов катаболизма пуринов. У животных и некоторых растений (проростки клещевины) пероксисомы играют важную роль при превращении жиров в углеводы. Так, в клетках эндосперма клевещины в пероксисомах (глиоксисомах) содержатся ферменты глиоксалатного цикла. Пероксисомы не содержат никаких нуклеиновых кислот и все белки, из которых они состоят, кодируются ядерными генами, но их относят к саморепродуцирующимся органеллам. В пероксидах происходит накопление специфических белков, которые синтезируются в цитозоле, и имеют свои сигнальные участки. В мембране пероксисом есть рецепторный белок, который узнает транспортируемые белки. Белки мембран пероксисом, также как и липиды приходят из цитозоля. Такое накопление содержимого и рост мембраны приводят к общему росту пероксисомы, которая затем с помощью неизвестного пока механизма делится на две – самореплицируется. Секреция белков и образование мембран у бактерий В принципе рост плазматической мембраны и её производных у бактерий происходит тем же образом, что и образование мембран у эукариотических клеток. Как известно, синтез белков у бактерий осуществляется на 70s рибосомах, которые также, как и у клеток высших организмов, имеют двоякую локализацию. Большая часть рибосом бактериальных клеток образует полисомы в цитоплазме, около 25% рибосом связано с плазматической мембраной. Такие рибосомы участвуют как в синтезе белков мембраны, так и в синтезе экскретируемых белков. Многие бактериальные клетки получают питательные вещества за счет деградации полимеров около бактериальной поверхности. Для этого бактерии должны выделять гидролизирующие ферменты в окружающую среду. Это они делают намного проще, чем эукариотические клетки: часть их рибосом, локализованных на внутренней (цитоплазматической) поверхности плазматической мембраны, синтезирует белки, которые, подобно секреторным белкам, проходят через мембрану и оказываются вне клетки. Выделенные гидролазы застревают в компонентах муреиновой бактериальной стенки и там функционируют. На других рибосомах, связанных с мембранами, идет синтез белков для построения самой мембраны, подобно тому, что происходит в гранулярном ЭР эукариотических клеток. Так что в этом отношении бактерию можно уподобить вакуоли гранулярного ЭР, вывернутой наизнанку. На примере бактерий хорошо изучен путь синтеза липидных компонентов мембран. Так, было найдено, что синтез фосфоэтидилэтаноламина происходит с помощью ферментов, являющихся интегральными белками плазматической мембраны, активные участки которых находятся на цитоплазматической стороне мембраны. Синтезированные здесь липиды встраиваются во внутренний липидный слой. Оказалось, что новосинтезированные липиды довольно быстро обнаруживаются и во внешнем слое мембраны за счет работы переносчиков – флиппаз. |
infopedia.su
Вакуоли в растительных клетках
Вакуоли - это полости, заполненные клеточным соком - сложным раствором различных водорастворимых соединений. Среди этих соединений имеются:
- Минеральные соли;
- Органические кислоты;
- Сахариды;
- Аминокислоты;
- Дубильные вещества;
- Водорастворимые пигменты: антоцианы, флавоны, беталанины;
- Некоторые белки и ферменты.
В вакуолях сосредоточено основное количество клеточной воды. Объем клетки возрастает исключительно за счет жидкости в вакуолях, а не цитоплазмы и ее структурных элементов. Объем центральной вакуоли поддерживается постоянным притоком молекул воды под воздействием осмотических явлений.
Клеточный сок часто бывает окрашен в различные оттенки красного или синеватого цвета. Распространенными красителями вакуолярной жидкости является антоцианы, которые меняют свой цвет в зависимости от соотношения с другими флавоноидных соединений, наличие ионов металла и величины рН сока. Если вакуолярного раствор имеет щелочную реакцию, антоциановые пигменты приобретают разные оттенки синего окраску (лепестки васильков, цикория, льна). Если вакуолярного раствор имеет кислую реакцию, то клеточный сок имеет красный цвет (лепестки роз, плоды вишни, калины, клубники).
Форма и размеры вакуолей изменяются от возраста клетки, уровня ее метаболической активности и выполняемых функций. В меристематических клетках они называются провакуолямы и их не видно в световой микроскоп. Постепенно с ростом и развитием клеток размер вакуолей резко увеличивается за счет их слияния и увеличения объема. В дифференцированных клетках находятся одна или несколько вакуолей в центральной части протопласта. Цитоплазма и ее структурные компоненты занимают пристенные положения или изредка пронизывают вакуолярного полость тонкими тяжами. Вакуоли отделены от цитоплазмы одной мембраной - тонопластом.
Центральная вакуоль взрослой клетки образуется в результате слияния мелких вакуолей меристематичнгих клеток. Формирование вакуолей изучено с помощью электронного микроскопа. Первыми появляются мельчайшие провакуоли, образующиеся из мелких пузырьков на тяжах эндоплазматической сети, что свидетельствует о родстве ее мембранных структур с мембраной вакуолей.
Функции вакуолей в растительной клетке.
- С их участием осуществляются осмотические процессы клеток, которые лежат в основе поглощения и движения воды, питательных веществ и поддерживается напряжен (тургорного) состояние клеточных оболочек.
- Благодаря отдельным вакуолях или их совокупности в клетке - ВАК достигается минимизация объема цитоплазмы и существенное увеличение поверхности раздела между цитоплазмой и тонопластом.
- Центральное положение и значительный объем вакуоли обеспечивает эффективное пространственное размещение хлоропластов с приближением их к поверхности клетки, условия оптимального освещения.
- Вакуоли принимают участие в детоксикации цитоплазмы, выполняя функцию буферной системы ее защиты от метаболических или ионных стрессов.
- Вакуоли свойственна лизосомных функция-расщепление некоторых макромолекул и отработанных органелл.
- Как внутренняя среда, вакуоли поддерживают гомеостаз растительной клетки, а процесс вакуолизации является необходимым условием роста клетки.
- Вакуоли часто является вместилищем различных вторичных метаболитов: алкалоидов, фловоноидив, вредных ионов и веществ. - Они также являются резервуаром, где откладываются про запас некоторые питательные вещества: сахариды, органические кислоты, белки, которые периодически включаются в обмен веществ и используются для нужд клетки как строительный и энергетический материал.
- Вакуоли также участвуют в явлении плазмолиза, так как процесс отставание цитоплазматического содержимого от клеточной оболочки, обусловленный уменьшением количества воды в вакуолях путем осмотических процессов.
Таким образом, вакуоли растительной клетки - полифункциональные образования.
worldofscience.ru
Обратная связь ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Как цель узнает о ваших желаниях прежде, чем вы начнете действовать. Как компании прогнозируют привычки и манипулируют ими Целительная привычка Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Тренинг уверенности в себе Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Как слышать голос Бога Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
| Комплекс Гольджи Открыт в 1898 году итальянским ученым Гольджи. Присутствует во всех эукариошках, за исключением эритроцитов. Обычно элементы комплекса Гольджи располагаются около ядра рядом с центросомой в животных клетках, а в растительных – по периферии. Участки комплекса имеют вид сложных сетей. Ячейки этой сети могут быть связаны друг с другом или же располагаться отдельно в виде вогнутых кого-то там, вакуолярных образований. При этом морфология комплекса очень зависит от фазы клеточного цикла. Это динамически сложная организованная и поляризованная система вакуолей. Комплекс Гольджи в типичном случае – это собранные в небольшой зоне отдельные скопления, участки вакуолей, которые называются цистернами. Стопка уплощенных цистерн – диктиосома. В диктиосоме плотно друг к другу на расстоянии 20 – 25 нм располагаются плоские мембранные цистерны, между которыми находятся прослойки гиалоплазмы. Каждая цистерна имеет диаметр около одного мкм и переменную толщину. В центре цистерны мембраны более сближены (25 нм), а по краям цистерны имеются ампулярные расширения. Ширина их не постоянна. Количество таких цистерн в стопке варьирует от пяти до десяти штук. У одноклеточных встречается до двадцати штук. Кроме этих структурных единиц в состав комплекса относится много мелких вакуолей, располагающихся главным образом в периферических участках. Эти мелкие вакуоли отшнуровываются от ампулярных окончаний и обрамляют всю диктиосому. В диктиосоме принято различать два полюса: 1) Проксимальный или формирующийся. Цис-полюс. Располагается ближе к ядру клетки. 2) Дистальный или зрелый транс полюс. 3) Медиальная часть – серединочка. На транс полюсе к комплексу примыкает участок, состоящий из трубчатых элементов и массой мелких вакуолей. Это опушенные пузырьки с каким-то белком. Принимают активное участие в экзоцитозе. Область – транссеть комплекса Гольджи. Именно в этой области происходит разделение и сортировка секретируемых продуктов. Ближе к плазмолемме располагается область более крупных вакуолей, которые являются продуктом слияния более мелких вакуолей с образованием секреторных вакуолей. В 1924 году Насоновым было выдвинуто предположение, что комплекс Г. является органоидом, обеспечивающим разделение (сепарация) и накопление веществ в клетке. Одна и та же клетка может участвовать в синтезе многих белков. Эта клетка изолирует их друг от друга и направляет к клеточной поверхности. Кроме того, в комплексе Гольджи происходит не только перекачка этих продуктов, но также и их постепенное созревание, модификация, которая заканчивается сортировкой продуктов. Модификация белков связана с вторичным гликозелированием, с реакцией фосфолирирования, с образованием полисахаридов матрикса клеточной стенки, с образованием сложных гликопротеинов, реакцией сульфатирование и реакцией отщепления олигосахаридных последовательностей и присоединением других белков. Буратино дрочил-дрочил, да и сгорел… Трансцистерна с накопленными продуктами распадается и мы имеем дело с более крупными секреторными пузырьками, но весь этот огромный массив секреторных пузырьков можно разбить на три потока: 1) Гидролитические ферменты (гидролазы), которые направляются в компартмент лизосом. Известно, что только белки предшественники гидролаз имеют особую маннозную группу в своем составе. В цис-цистернах эти группы фосфолирируются. И далее, вместе с другими белками, переносятся от цистерны к цистерне в транс участок. Мембраны транс полюса содержат особый рецептор. Манноза-6-фосфат, который специфически узнает фосфолиророванные маннозные группировки лизосом элементов и взаимодействует с ними. Это связывание осуществляется при нейтральных значениях рН внутри цистерн трансполюса. На мембранах пузырьков манноза-6-фосфатные рецепторы образуют группы, которые концентрируются в окаймленных клатрином пузырьках. Оторвавшись от трансполюса, эти пузырьки теряют клатрин, сливаются с эндосомами, перенося свои лизосомные ферменты (гидролазы), связанные с мембранными рецепторами в эту вакуоль. Внутри эндосом, благодаря работе протонного переносчика, происходит закисление среды и начиная где-то с рН = 6 лизосомные ферменты гидролазы отсоединяются от маноза-6-фосфата активируются и начинают работать – переваривать органику. Участки же мембран, вместе с рецепторами, возвращаются путем рециклинга обратно в транссеть комплекса Гольджи. 2) Белки, которые накапливаются в секреторных вакуолях и выделяются из клетки только при получении специальных сигналов. Это путь стимулируемой секреции. Считают, что та часть белков, которая накапливается в секреторных вакуолях и выводится из клетки после поступления сигнала либо нервного, либо гормонального, проходят такую же процедуру отбора сортировки на рецепторах цистерн комплекса Гольджи. Эти секреторные белки попадают сначала в мелкие вакуоли, которые тоже одеты клатрином. В секреторных вакуолях происходит агрегация накопленных белков в виде плотных секреторных гранул. Концентрация белка при этом в вакуолях повышается в 2000 раз по сравнению с концентрацией белка в цистернах комплекса. Секреторные вакуоли выбрасываются из клетки путем экзоцитоза после получения клеткой соответствующего стимула. 3) Путь конститутивной (постоянной) секреции. Клетки могут постоянно выделять белки, которые связывают их с субстратами. Кроме того, непрерывно идет поток мембранных пузырьков к плазмолемме, в которых находятся элементы гликокаликса и мембранных протеинов. Этот поток не подлежит сортировке в рецепторной транссистеме комплекса Гольджи. Внешне они не отличаются от других пузырьков. Также являются окаймленными секреторными пузырьками.
Функции комплекса Гольджи: 1) Участие в сегрегации и накопление продуктов синтезированных в ЭПР; 2) участие в химических перестройках и созревании органики. Главным образом, это перестройка олигосахаридных компонентов гликопротеинов в составе водорастворимых секретов или в составе мембран; 3) Это синтез полисахаридов, их взаимосвязь с белками, приводящая к образованию мукопротеинов, гликопротеинов; 4) Выведение готовых секретов за пределы клетки; 5) Формирование лизосом.
Характеристика лизосом
Лизосомы не являются самостоятельными структурами клетки, т.к. они образуются за счет активности ЭПР и комплекса Гольджи. И в этом отношении они очень похожи на секреторные вакуоли. Основная их функция заключается в участии в процессах внутриклеточного расщепления, как экзогенных, так и эндогенных макромолекул. Открыты были французским ученым Де Дювом в 1955 году. Окружены одной липопротеидной мембраной. Содержат кислые гидролитические ферменты (гидролазы), расщепляющие белки, липиды и т.д. (всю органику). По специфичности делятся примерно на 40 видов, например, нуклеазы, гликозидазы, сульфидазы и т.д. Оптимум действия гидролаз рН – 5,5 (кислая среда). Самая характерная – кислая фосфотаза. Протонный насос создает и поддерживает кислое значение среды. Этот насос является АТФ-зависимым. Было высказано предположение, что, вероятнее всего, мембраны лизосом защищены от действия кислых гидролаз олигосахаридными участками, которые присоединяются к гидролазам в ЭПР в результате процесса первичного гликозелирования. Эти участки либо не узнаются гидролазами вообще, либо просто мешают гидролазами взаимодействовать с ними.
Гидролазы обладают способностью быстро активироваться: 1) Развитый рецепторный аппарат; 2) За счет микротрубочек лизосомы очень активно перемещаются в клетке; 3) Они способны к локальному разрушению мембраны при контакте с эндосомами.
Лизосомы представлены несколькими фракциями. Подразделяются на первичные, вторичные, остаточные тельца или тело лизосомы и аутофагосомы. Первичные лизосомы – мелкие мембранные пузырьки, около 100 нм, с бесструктурным содержимым, содержащим набор ферментов. Их практически невозможно отличить от других секреторных пузырьков. Часть из них имеет клатриновую оболочку. Первичные лизосомы в дальнейшем сливаются с эндосомами, поступившими в клетку снаружи и образуют вторичную лизосому или внутриклеточную пищеварительную вакуоль. При слиянии первичной лизосомы с эндосомой происходит диссоциация комплексом маноза-6-фосфатного рецептора гидролаза из-за кислой среды в лизосоме. Свободный фермент после потери фосфатной группы вступает в работу. Расщепляет полимеры до мономеров, затем происходит транспорт в состав гиалоплазмы клетки, где они включаются в процессы. Первичная лизосома может вновь слиться с эндосомой. Процесс слияния и переваривания - гетерофагицескай цикл. Однако переваривание и расщепление молекул может идти и не до конца. В этом случае, в полостях лизосом происходит накопление непереваренных продуктов и вторичная лизосома превращается в остаточное тельце или в тело лизосомы. Остаточные тельца уже содержат меньше гидролаз. В них происходит уплотнение содержимого, его перестройка. Часто в остаточных тельцах наблюдается вторичная структуризация непереваренных липидов, которые образуют сложные слоистые структуры. Во вторичных лизосомах происходит и отложение специфических продуктов обмена веществ и пигментов. Например, липофусцин. Кроме гетерофагического цикла, в клетках происходит и аутофагический цикл, связанный с перевариванием собственных структур клетки. Аутофагосомы присутствуют и в клетках простейших, и в клетках растений, и у животных. По своей морфологии их относят к вторичным лизосомам, но с тем отличием, что в составе этих вакуолей встречаются фрагменты или даже целыецитоплазматические структуры, такие как митохондрии, пластиды, элементы ЭПР, рибосомы, гранулы гликогена и другие вещества. Полагают, что процесс образование аутофагосом связан с выстраиванием вокруг клеточного органоида первичных лизосом, затем их слияние друг с другом. Таким образом они отделяют структуру, подлежащую перевариванию, от основной цитоплазмы. Полагают, что аутофагосомы задействованы в процессе абоптоза (запрограммированная гибель). Такой функции подвергаются митохондрии печени, которые живут 9 – 10 суток. Установлено, что число лизосом увеличивается в клетке при патологии. Даже при разрыве клетки, лизосомные гидролазы теряют свою активность и переваривания клетки не происходит. Ферменты несомненно участвуют в автолизе, но, скорее всего, это вторичное явление, а не причина гибели. Лизосомные накопления – первичная генная мутация, приводящая к потере активности отдельных ферментов, участвующих в функционировании лизосом.
Вакуоли растительных клеток.
Молодые растительные клетки имеют несколько мелких вакуолей. Далее вакуоли сливаются и образуют одну крупную вакуоль, которая занимает примерно 90% объема клетки, оттесняя органоиды к периферии. Отделены вакуоли одинарной мембранной, сходной по толщине с плазмолеммы.Тонопласт. Полость вакуоли заполнена клеточным соком, представляющим собой водный раствор, в который входят различные неорганические соли, сахара, органические кислоты и их соли, некоторые высоко молекулярные вещества и т.д. Центральные вакуоли растений выполняют множество функций: 1) Поддержание тур горного внутреннего давления клеток. Тургор определяют растворенные в клеточном соке вакуолей молекулы. 2) Вакуоли используются клетками для складирования метаболитов иди для осуществления процесса экскреции, т.е. выделения. Через тонопласт осуществляется активный транспорт различных молекул. Здесь находится протонная помпа, через которую, путем механизма с импорта, происходит транспорт сахаров. Таким образом выводятся из клетки все растворимые метаболиты, нерастворимые в воде неорганические вещества превращаются в центральной вакуоли в растворимые глюкозиды, соединяясь с молекулами сахаров. Также происходит отложение различных пигментов, неорганических кристаллов, солей. За счет складирования этих соединений вакуолярной сок имеет выраженную кислую реакцию от 2 рН до 5. 3) Накопление запасных продуктов. В первую очередь это сахара и белки. Отложение белков в живых молодых клетках редко. В основном они откладываются в семенах. Образование алейроновых зерен связано со способностью мембран ЭПР и комплекса Гольджи сливаться с тонопластом. 4) Гидролиз. В центральной вакуоли обнаружены гидролазы. Тонопласт может впячиваться внутрь вакуоли, получаются складки, которые отщепляются и плывут в вакуоль. Аутофагическая функция вакуоли это. Лизосомными свойствами обладают вакуоли дрожжей. Отдельной структурой вакуолярной системы являются сферосомы. Они встречаются в растительных клетках и окрашиваются липофильными красителями, имея очень высокий коэффициент преломления. Образуются они их элементов ЭПР. Пузырек отшнуровывается. Это Просферосома. Имеет одинарную мембрану. В ней происходит накопление масла. Она постепенно растет, в ней происходит перестройка и в конце-концов она превращается в масляную каплю окруженную мембраной. Но сферосомы выполняют также функцию расщепления белков. В них находится фермент, который обеспечивается особый тип метаболизма.
Пероксисомы
Пероксисомы. Обнаружены у простейших, низших грибов, высших растений, эмбриональных тканях, у высших позвоночных в печено и почках. Локализация – рядом с мембранами ЭПР. В растительных клетках пероксисомы имеют теснейший контакт с митохондриями. Это органоиды, обладающие ферментативной системой образования и утилизацией перекиси водорода, глиоксилата и т.д. Пероксисомы регулируют окислительно-восстановительной равновесие внутри клетки и концентрацию активных форм кислорода. Были открыты тем же Де Дювом в 1960 году. Это небольшие вакуоли, диаметром от 0,3 до 1,5 мкм, одетые одинарной мембраной, отграничивающей гранулярный матрикс, в центре которого располагается сердцевина. У млекопитающих представлены пероксисомы двумя формами – универсальными мелкими и более крупными, характерными в основном для клеток печени и почек. Мембраны пероксисом либо из мембран гладкого ЭПР, либо из предшествующих пероксимом в процессе роста и деления, подобно митохондриям и хлоропластам. Пероксисомы относятся к саморедуцирующимся органеллам, хотя в них нет ни нуклеиновых кислот, ни аппарата для репликаций. В пероксисоме происходит накопление специфических белков и липидов, которые синтезируются в цитозоле, затем поступают в пероксисому. Происходит ее рост, а затем деление на две. В пероксисомах обнаруживаются: оксидазы, оксидаза-d-ам.к. Эти ферменты являются маркерами аминокислот. Это ферменты окислительного дезаминирования. При работе этих ферментов образуется перекись водорода и каталаза, которая разрушает перекись. Каталаза составляет примерно 40%. Де Дю, открывший пероксисомы, пришел к заключению, что эти органоиды представляют собой реликты того времени, когда митохондрии еще не образовались. По сути, это органоид, выполняющий функцию биологического окисления. Они находятся на перекрестке биохимических путей, которые замыкают реакции, осуществляющиеся в разных компартментах клетки. Осуществляя реакции, сопровождающиеся высвобождением энергии, пероксисомы связывают окислительный метаболизм в клетке с конструктивным и генерируют пути синтеза разнообразных соединений, например, превращение жиров в углеводы. И регуляция окислительного-восстановительного равновесия и регуляция концентрации активных форм кислорода, которые вызывают повреждение клетки. |

 Отёска стен и прирубка косяков - Когда на доме не достаёт окон и дверей, красивое высокое крыльцо ещё только в воображении, приходится подниматься с улицы в дом по трапу.
Отёска стен и прирубка косяков - Когда на доме не достаёт окон и дверей, красивое высокое крыльцо ещё только в воображении, приходится подниматься с улицы в дом по трапу. Дифференциальные уравнения второго порядка (модель рынка с прогнозируемыми ценами) - В простых моделях рынка спрос и предложение обычно полагают зависящими только от текущей цены на товар.
Дифференциальные уравнения второго порядка (модель рынка с прогнозируемыми ценами) - В простых моделях рынка спрос и предложение обычно полагают зависящими только от текущей цены на товар.megapredmet.ru
Вакуоль. Функции вакуоли | Биология
Вакуолями называются крупные мембранные пузырьки или полости в цитоплазме, заполненные преимущественно водным содержимым. Они образуются из пузыревидных расширений эндоплазматического ретикулума (ЭР) или из пузырьков комплекса Гольджи (КГ). В меристематических клетках растений возникает много небольших вакуолей из пузырьковидных расширений ЭР. Увеличиваясь, они сливаются в центральную вакуоль, которая занимает большую часть объема клетки (до 70-90 %) и может быть пронизана тяжами цитоплазмы. Окружающая ее мембрана - тонопласт - имеет толщину мембраны ЭР (около 6 нм) в отличие от более толстой, более плотной и менее проницаемой плазмалеммы.
Содержимое вакуоли составляет клеточный сок. Он представляет собой водный раствор различных неорганических и органических веществ. Большинство из них относится к группе продуктов метаболизма протопласта, которые могут появляться и исчезать в различные периоды жизни клетки. Химический состав и концентрация клеточного сока очень изменчивы и зависят от вида растения, органа, ткани и состояния клетки. В клеточном соке содержатся соли, сахара (прежде всего сахароза, глюкоза, фруктоза), органические кислоты (яблочная, лимонная, щавелевая, уксусная и др.), аминокислоты, белки. Эти вещества являются промежуточными продуктами метаболизма, временно выведенными из обмена и изолированными тонопластом. Они являются запасными веществами клетки. Помимо запасных веществ, которые могут вторично использоваться в обмене веществ, клеточный сок содержит фенолы, танины (дубильные вещества), алкалоиды, которые выводятся из обмена в вакуоль и таким путем изолируются от цитоплазмы.
Танины особенно часто встречаются в клеточном соке (а также в цитоплазме и оболочках) клеток листьев, коры, древесины, незрелых плодов и семенных оболочек. Алкалоиды присутствуют, например, в семенах кофе (кофеин), плодах мака (морфин) и белены (атропин), стеблях и листьях люпина и др. Считается, что танины с их вяжущим вкусом, алкалоиды и токсичные полифенолы выполняют защитную функцию, так как отпугивают травоядных животных и предотвращают поедание этих растений.
В вакуолях также часто накапливаются конечные продукты жизнедеятельности клеток.
В клеточном соке многих растений содержатся пигменты которые придают клеточному соку пурпурный, красный, желтый, синий или фиолетовый цвет. Эти пигменты главным образом и определяют окраску лепестков цветков (например, роз, георгинов, фиалок, примулы и др.), плодов, почек и листьев, а также окрашивают корнеплоды некоторых растений (например, свеклы).
Клеточный сок некоторых растений содержит физиологически активные вещества - фитогормоны (регуляторы роста), фитонциды, ферменты. В последнем случае вакуоли действуют, как лизосомы. После гибели клетки ферменты, высвобождаясь из вакуолей, вызывают автолиз клетки.
Вакуоли играют главную роль в поглощении воды растительными клетками. Вода путем осмоса через тонопласт поступает в вакуоль, клеточный сок которой является более концентрированным, чем цитоплазма, и оказывает давление на цитоплазму, а тем самым и на оболочку клетки. В результате в клетке развивается тургорное давление, которое поддерживает относительную жесткость растительных клеток, а также обусловливает растяжение клеток во время их роста. В запасающих тканях растений вместо одной центральной вакуоли часто бывает несколько вакуолей, в которых скапливаются запасные питательные вещества, как, например, жировые вакуоли (содержащие растительные масла) или белковые (алейроновые) вакуоли.
Читать далее
ed-lib.ru
Вакуоли растительных клеток.
Количество просмотров публикации Вакуоли растительных клеток. - 387
Клетки как низших, так и высших растительных организмов содержат в цитоплазме вакуоли, выполняющие важные физиологические функции (рис. 195).
По мере роста и дифференцировки клетки мелкие вакуоли могут сливаться друг с другом и образовывать одну или несколько крупных вакуолей, занимающих до 90% объёма всей клетки. Вакуоли отделены от цитоплазмы одинарной мембраной, сходной по толщине с плазмалеммой. Мембрана, ограничивающая центральные вакуоли, носит название тонопласта. Возникают центральные вакуоли из мелких пузырьков, отщепившихся от аппарата Гольджи. Такие первичные вакуоли растут в объёме, сливаются друг с другом и образуют одну или несколько крупных вакуолей, оттесняющих цитоплазму с ядром и органоидами к периферии клетки. Полости вакуолей заполнены клеточным соком, представляющим собой водный раствор, в котором находятся неорганические соли, сахара, органические кислоты и их соли и другие низкомолекулярные соединения, а также некоторые высокомолекулярные вещества (к примеру, белки).
Главной функцией центральных вакуолей является поддержание тургорного давления клеток. Растворенные в соке вакуолей молекулы определяют его осмотическую концентрацию. Соответствующая молекулярная концентрация сока вакуолей и полупроницаемые свойства как ее мембраны, тонопласта͵ так и плазмалеммы способствуют тому, что вакуоль функционирует в качестве осмометра и придает клетке необходимую прочность и тургисцентность (напряженность).
Другая функция центральных вакуолей состоит по сути в том, что тонопласт обладает свойствами полупроницаемости и через него происходит, как и через плазматическую мембрану, активный транспорт различных молекул. В тонопласте обнаружен АТФ-зависимый Н+-насос, направленный внутрь вакуолей, участвующий в транспорте сахаров. По этой причине вакуоли используются клетками как накопительные резервуары не только для отложения запасных веществ, но и для выброса метаболитов, для экскреции. Так выводятся, секретируются из клетки все водорастворимые метаболиты. Нерастворимые в воде органические вещества превращаются в растворимые глюкозиды, соединяясь с молекулами сахаров. Перечень экскретируемых в вакуоли метаболитов очень обширен. Это различные алкалоиды (к примеру, никотин, кофеин) и полифенолы. В вакуолях происходит отложение многих глюкозидов, к которым относятся различные пигменты, к примеру антоцианы.
Из неорганических веществ в вакуолярном соке накапливаются фосфаты калия, натрия, кальция, могут накапливаться соли органических кислот (оксалаты, цитраты и др.). Это придает вакуолярному соку отчетливую кислую реакцию (рН от 2 до 5).
Таким образом тонопласт участвует в процессах экскреции.
Другой ряд функций вакуолей связан с накоплением запасных веществ, таких, как сахара и белки. Сахара в вакуолях содержатся в виде растворов, встречаются и резервные полисахариды типа инулина. В вакуолях происходит запасание белков, что характерно для семян. Поступление белков в вакуоли, связано со способностью вакуолей ЭР и АГ сливаться с тонопластом. Запасание белков семян злаковых происходит в так называемых алейроновых вакуолях, которые заполняются альбуминами и глобулинами, после чего вакуоли обезвоживаются, превращаясь в твердые алейроновые зерна. При прорастании семян эти зерна обводняются и снова превращаются в вакуоли. В таких новообразованных вакуолях выявляется активность некоторых ферментов, кислой фосфатазы, a-амилазы, глюкозидазы, протеиназы и РНКазы. Следовательно, алейроновые вакуоли отчасти напоминают лизосомы, где происходит переваривание запасных белков при прорастании семян.
Сферосомы. Это мембранные пузырьки, встречающиеся в клетках растений. Сферосомы образуются из элементов эндоплазматического ретикулума. На конце цистерны ЭР начинает накапливаться осмиофильный материал, затем от этого участка отшнуровывается и начинает расти мелкий пузырек, достигающий диаметра 0,1-0,5 мкм. Это “просферосома”, окруженная одинарной мембраной. Рост сферосом и перестройка их содержимого связаны с накоплением в них масла, так что сферосома постепенно превращается в масляную каплю. Отложение липидов начинается между осмиофильными слоями мембраны. Кроме жиров в составе сферосом обнаруживают белки и среди них фермент липазу, расщепляющую липиды.
4.3.5.Пероксисомы (микротельца)
Это небольшие вакуоли (0,3-1,5 мкм), одетые одинарной мембраной, отграничивающей гранулярный матрикс, в центре которого располагается сердцевина, или нуклеоид (ничего не имеющий общего с нуклеоидом бактерий и вобще к ядерным структурам не относящийся).
В зоне сердцевины часто, особенно в пероксисомах печеночных клеток, видны кристаллоподобные структуры, состоящие из регулярно упакованных фибрилл или трубочек. Изолированные сердцевины пероксисом содержат фермент уратоксидазу (рис. 196, 207б).
Пероксисомы обнаружены у простейших (амебы, тетрахимена), у низших грибов (дрожжи), у высших растений в некоторых эмбриональных тканях (эндосперм) и в зеленых частях, способных к фотореспирации, у высших позвоночных животных они обнаруживаются главным образом в печени и почках. В печени крыс на клетку число пероксисом колеблется от 70 до 100.
Пероксисомы часто локализуются вблизи мембран ЭР. У зеленых растений пероксисомы часто находятся в тесном контакте с митохондриями и пластидами.
Впервые пероксисомы были выделены из печени и почек. Во фракциях пероксисом обнаруживается ферменты, связанные с метаболизмом перекиси водорода. Это ферменты (оксидазы, уратоксидаза, оксидаза d-аминокислот) окислительного дезаминирования аминокислот, при работе которых образуется перекись водорода (Н2О2 ) и каталаза, разрушающая ее. В пероксисомах печени каталаза составляет до 40 % всех белков и локализована в матриксе.
У животных и некоторых растений (проростки клещевины) пероксисомы играют важную роль при превращении жиров в углеводы.
Пероксисомы относят к саморепродуцирующимся органеллам. В пероксидах происходит накопление специфических белков, которые синтезируются в цитозоле, и имеют свои сигнальные участки. В мембране пероксисом есть рецепторный белок, который узнает транспортируемые белки. Белки мембран пероксисом, также как и липиды приходят из цитозоля. Такое накопление содержимого и рост мембраны приводят к общему росту пероксисомы, которая затем с помощью неизвестного пока механизма делится на две – самореплицируется.
referatwork.ru









