
Синергизм: влияние чередования культур на улучшение эффективности роста. Синергизм в растениях
антагонизм и синергизм ионов. Уравновешенные растворы.
Р-ры, в которых токсического действия солей нет, называют физиологически уравновенными (морская вода-близка по составу к крови и лимфе животных). Физиологически уравновешенными являются те растворы, количество и соотношение ионов в которых исключают их вредное влияние. Такие растворы обеспечивают нормальный рост, развитие и высокую продуктивность растений. При составлении физиологически уравновешенных растворов необходимо учитывать разный характер взаимодействия ионов. Растворы чистых солей оказывают токсическое действие на организм растения или животного. Еще в конце прошлого века была установлена ядовитость раствора чистой соли NаС1, концентрация которого соответствовала концентрации морской воды. Достаточно было добавить незначительное количество солей кальций и магния, чтобы снять это ядовитое действие NaС1. Взаимодействие ионов, при котором физиологический эффект воздействия смеси солей меньше, чем действие каждой соли в отдельности, называется антагонизмом. Антагонизм ионов проявляется как между разными ионами одной валентности, например Nа+ и К+, Иа+ и Nh5, так и между ионами разной валентности (К+ и Са2+, Nf+ и Мg2+). Явление антагонизма может наблюдаться при развитии корневой системы растений, выращенных в след. р-рах (0,1н): 1)NaCl+KCl+CaCl2; 2)NaCl+CaCl2; 3)CaCl2; 4)NaCl. Прорастание пшеницы или ячменя, появляется в этих р-рах, различно: в 1 и 2 корненая система более нормальная, в 3 и 4 корневая система почти не развита. Явление антагонизма зависит от валентности ионов: чем выше валентность, тем в меньшей концентрации проявляется антагонистическое действие. Так, для того чтобы устранить ядовитое действие на проростки пшеницы односолевого раствора КС1, надо добавить 30 % NaС1 или 5 % СаС12. В основе антагонистического действия ионов лежит их противоположное действие на физико-химические свойства протоплазмы, что отражается на обмене веществ, а также конкуренция при усвоении их растениями. Наблюдаются также синергизм и аддитивность в действии компонентов солей. Синергическое действие ионов заключается в том, что один из них усиливает действие другого. Например, катионы К, Са, Мg оказывают стимулирующее действие на поглощение анионов NОз и РО4 Нитратные и фосфатные анионы благоприятно влияют на усвоение других элементов. Аддитивность — это действие смеси солевых растворов, которое равно сумме действия отдельных компонентов. Например, осмотическое давление питательного раствора равно сумме парциальных осмотических давлений входящих в смесь солей.
25. Пути обезвреживания аммиака в растении.
В процессе дыхания растений образуются органические к-ты:-кетоглутаровая и щавелево-уксусная (ЩУК). Они путем прямого аминирования присоединяют аммиак: 1)HOOC·Ch3·Ch3CO·COOH+Nh4+2H↔HOOC·Ch3Ch3CHNh3·COOH+h3O(глутаминовая к-та) 2)HOOC·COCh3COOH+Nh4+2H↔HOOC·Ch3CHNh3·COOH+h3O(аспаргиновая к-та).
Более простым способом связывания аммиака является образование аммонийных солей органических кислот. У растений с кислым клеточным соком (щавель, бегония, осоки, хвощи) содержание аммонийного азота в десятки раз превосходит содержание амидного азота.
Аммиак в растениях обезвреживается и при образовании мочевины. Исходным продуктом для синтеза мочевины служит аминокислота орнитин, которая, присоединяя аммиак и углекислоту, превращается в цитруллин. Цитруллин присоединяет еще одну молекулу аммиака и образует аргинин. Аргинин под действием фермента аргиназы с участием воды расщепляется на орнитин и мочевину. Мочевина неядовита для растений, хорошо усваивается, использование ее азота для всевозможных синтетических процессов происходит очень легко, так как в растительных тканях имеется фермент уреаза, катализирующий расщепление мочевины.
26 Микроэлементы, их роль в жизни растения.
В кислых почвах мало кобальта и меди, в щелочных почвах их много. Микроэлементы в почве могут быть в составе почвенных минералов, в поглощенном состоянии на поверхности почвенных коллоидов. Водорастворимые формы микроэлементов содержат 1-10% от их общего кол-ва в почве. Из водорастворимых соединений микроэлементы могут легко переходить в обменные формы, поглощаться растениями и м/о. Запас микроэлементов в почве пополняется в процессе разрушения почвенных минералов, а также минерализации орг. в-в, удобрений, микроорганизмов. Кол-во микроэлементов, поступающих в растения, неодинаково, оно зависит от вида растения, почвенно-климатических условий, комплекса агротехнических мероприятий. Микроэлементы и ультрамикроэлменты входят в состав орг.соединений, играющих важную роль в обмене в-в – хелатов - это органические внутрикомплексные соединения циклического строения, содержащие в своей молекуле ион какого-либо Ме, который участвует в образовании кольца. Образование хелатов обеспечивает передвижение Ме по сосудам растения, перенос электронов между кристаллически активными белками. Микроэлементы обуславливают поступление анионов и катионов в растения, влияют на плодоношение растений, способствуют процессу оплодотворения, повышают урожайность семян. Улучшают кач-во продукции с/х растений: увеличивают номерность волокна льна, конопли, повышают содержание сахаров в корнях сахарной свеклы, посевные св-ва семян бобовых и др.культ. В почву вносят микроудобрения, применяют некорневую подкормку растворами солей микроэлементов (плодовых и ягодных культ). Применяют много форм удобрений: медные, марганцевые, борные, комбинированные.
Микроэлементы относятся к группе незаменимых питательных элементов, содержание которых в растительных тканях измеряется тысячными и стотысячными долями процента. Несмотря на то, что микроэлементы требуются в очень малых количествах, в их отсутствие нормальная жизнедеятельность становится невозможной. Недостаток микроэлементов вызывает серьезные физиологические расстройства и нередко приводит к гибели растений уже в раннем возрасте. Объясняется это тем, что микроэлементы главным образом функционируют в регуляторных системах клетки. Они выступают в качестве простетических групп ферментов или кофакторов — активаторов ферментов.
Цинк. Входит в состав фермента поддерживающего уровень СО2 для фотосинтеза. Недостаток цинка приводит к резкому торможению роста и формирования побегов. Растения приобретают розеточную форму.
Молибден. Играет особо важную роль в азотном обмене. При дефиците рост тормозится, особенно чувствительны бобовые и овощные.
Бор. При его недостатке нарушается фотосинтез, превращение и транспорт углеводов, формирование репродуктивных органов.
studfiles.net
влияние чередования культур на улучшение эффективности роста, сайта Зерно
Причина синергизма неизвестна. Но он реально достижим и способен улучшать показатели
Севообороты сейчас включают в себя большее разнообразие культур по сравнению с традиционными системами обработки почвы, где выращивается одна или две культуры. Разнообразие культур нередко приводит к увеличению урожаев за счет эффекта чередования. Мы обратили внимание, что реакция культуры на эффект чередования может быть выражена либо в улучшении эффективности использования ресурсов, либо в увеличении размера растения и повышении его урожайности. Примером эффективной реакции, называемой синергизмом, является то, что ряд культур повышает эффективное использование влаги (ЭИВ) последующими культурами
Культуры производят больше зерна при использовании такого же количества воды в синергетических последовательностях. Для сравнения: культуры, которые увеличивают свою урожайность, потребляют больше воды, производя большее количество зерна. Синергизм разных культур увеличивает устойчивость к сорнякам из-за улучшенной эффективности использования ресурсов. Синергизм еще более ощутим в условиях получения низкой урожайности, связанной со стрессом (например засухой). Причина синергизма культур, вероятно, связана со множеством взаимодействующих факторов, таких как микробиологические изменения, вещества – стимуляторы роста и измененный круговорот питательных веществ. Методом выявления синергизма среди культур может быть количественное определение эффективности использования влаги (ЭИВ) от воздействий предшествующей культуры. No-till-севообороты, включающие синергетическую последовательность культур, повышают продуктивность земли, экономические показатели фермы, здоровье почвы и эффективность использования ресурсов в полузасушливом климате Великих равнин.
1. Некоторые культуры в севообороте улучшают эффективность использования влаги
При долгосрочном исследовании севооборота в регионе Великих равнин мы обнаружили дополнительную выгоду от выращивания разнообразных культур: некоторые культуры улучшают ЭИВ последующими культурами. Например, озимая пшеница (Triticum aestivum L.) после гороха (Pisum sativum L.) при использовании одинакового количества воды производит на 10-25% больше зерна, чем после озимой пшеницы, проса обыкновенного (Panicum miliaceum) или после пара. Такое же увеличение эффективности использования влаги было обнаружено, когда горох выращивался перед яровой пшеницей в Канадских прериях.
По мере дальнейшего исследования чередования культур, проведенного на Великих равнинах, мы обнаружили, что реакция культуры на эффект чередования делилась на две категории: либо повышение эффективности культуры, либо увеличение размера растения, что является хорошим потенциалом для роста урожайности. Эти реакции отличаются, поскольку при росте потенциальной урожайности растению требуется больше воды. Примером повышения потенциальной урожайности является рапс (Brassica napus L.), предшествующий озимой пшенице. Рапс снижает уровень заболеваний корней озимой пшеницы, а здоровые корни извлекают из почвы больше влаги. Дополнительное количество влаги способствует образованию более крупных растений с более высоким потенциалом урожайности. Однако если в почве дополнительной влаги нет, урожайность озимой пшеницы не увеличится.
Подобный контраст в реакции на эффект последовательности происходит с кукурузой (Zea mays L.) и соей (Glycine max Merril). Коуплэнд с соавторами (1993) обнаружил, что соя более эффективно использовала влагу после кукурузы, по сравнению с выращиванием монокультуры сои. В свою очередь кукуруза использует больше влаги для увеличения урожайности после сои, чем в случае выращивания монокультуры кукурузы. При выращивании после сои у кукурузы увеличиваются размер растений и их потенциальная урожайность, а эффективность не увеличивается. Последовательности, которые повышают эффективность культуры, дают больший урожай при использовании одинакового количества влаги.
Такое изменение в эффективности, которое мы называем синергизмом, может помочь производителям улучшать продуктивность культур без необходимости увеличения организационных затрат. Далее приведены результаты наших исследований, способствующие пониманию и интегрированию этого преимущества в системы производства культур.
2. Синергизм при использовании систем No-till и различных севооборотов
Изначально мы наблюдали за проявлением синергизма в ходе исследования систем No-till на северо-востоке Колорадо, где производилось оценивание 15 севооборотов. Севообороты включали в себя такие культуры, как кукуруза, подсолнечник (Helianthus annuus L.), соя, просо, просо итальянское (Setaria italica (L.) Beauv), овес (Avena sativa L.), горох и озимая пшеница. Пар был также включен в некоторые севообороты. Все фазы каждого севооборота были представлены ежегодно. Мы высчитывали ЭИВ для урожайности зерна, основываясь на количестве выпадающих осадков плюс влага, извлеченная из почвы в течение вегетационного периода культуры. Извлечение влаги из почвы с глубины 2 м определялось для каждой культуры с помощью нейтронного влагомера. Ежегодное количество осадков на этом участке в среднем составляло 416 мм.
2.1 Озимая пшеница
Через пять лет после начала исследования эффективность использования влаги озимой пшеницей начала отличаться в разных севооборотах (Anderson, 2005a). Так, ЭИВ посевами озимой пшеницы увеличилась с 9,0 кг/га/мм в севообороте озимая пшеница – пар (ОП-П) до 11,2 кг/га/мм в севообороте озимая пшеница – кукуруза – пар (ОП-К-П; график 1). Сначала мы предполагали, что эффективность использования влаги обусловлена тем, что более длительный интервал между культурами озимой пшеницы в севообороте ОП-К-П подавляет болезни корней озимой пшеницы, как это отмечал Кук (1990).
Однако ЭИВ озимой пшеницы не отличалась между севооборотами озимая пшеница – пар и озимая пшеница – просо – пар (ОП-Пр-П). Более того, ЭИВ озимой пшеницы не увеличивалась в севооборотах с более длительными интервалами между культурами озимой пшеницы; ЭИВ озимой пшеницы была одинаковой при использовании севооборотов озимая пшеница – кукуруза – просо – пар (ОП-К-Пр-П) и озимая пшеница – кукуруза – пар (ОП-К-П). Это изменение в ЭИВ, похоже, связано с наличием кукурузы в севообороте.
Мы также обратили внимание, что горох, выращиваемый в качестве зеленого пара (рост останавливали через 7 недель после посева), улучшал ЭИВ озимой пшеницы в севообороте озимая пшеница – кукуруза – просо – горох (ОП-К-Пр-Г). ЭИВ озимой пшеницей в этом севообороте была 12,3 кг/га/мм, что на 11% выше, чем в севообороте ОП-К-Пр-П и ОП-К-П. В севообороте ОП-К-Пр-Г озимая пшеница производила на 37% зерна больше по сравнению с ОП-П при одинаковом использовании влаги.
Эффективность использования влаги озимой пшеницей не изменялась ни в каких других севооборотах. Например, ЭИВ озимой пшеницы была одинаковой в севооборотах озимая пшеница – просо – подсолнечник – пар (ОП-Пр-Под-П) и ОП-П (график 1). ЭИВ посевами озимой пшеницы увеличивалась только в том случае, если в севообороте были представлены кукуруза и горох.
2.2 Просо обыкновенное
В этом же исследовании мы заметили, что эффективность использования влаги посевами проса отличалась, когда его предшественниками были кукуруза и озимая пшеница. ЭИВ посевами проса составляла 9,3 кг/га/мм после кукурузы, но лишь 7,5 кг/га/мм после озимой пшеницы (Anderson, 2005a). Общее использование влаги просом было одинаковым в обеих последовательностях, тем не менее просо оказывалось на 24% эффективнее в преобразовании влаги в зерно, если выращивалось после кукурузы. Эта разница в ЭИВ была постоянной в течение 4 лет, несмотря на то, что урожайность колебалась в пределах от 1540 до 3510 кг/га.
Когда мы рассматривали эти изменения в ЭИВ у озимой пшеницы и проса, мы не подумали, что отличия могут быть связаны с управлением культурами. Культуры выращивали с применением лучших методов управления для данного региона. Управление плодородием основывалось на проведении анализов почвы и прогнозируемых показателях урожайности и включало в себя стартовые удобрения, вносимые с семенами. Сорняки эффективно контролировались гербицидами, и мы не наблюдали какого-либо нашествия вредных насекомых, которыми можно было бы объяснить эти различия. Мы не знали, как объяснить изменение в ЭИВ, однако кукуруза улучшала использование влаги просом, а ЭИВ озимой пшеницей увеличивалась, когда в севооборот были включены кукуруза и горох. Мы не наблюдали изменений ЭИВ по другим культурам, используемым в данном исследовании.
2.3 Соя
Похожее изменение ЭИВ происходит, когда соя выращивается после кукурузы. Эффективность использования влаги соей увеличивалась на 14%, когда предшественником сои была кукуруза, по сравнению с вариантом, когда соя выращивалась после сои. Для сравнения: ЭИВ кукурузой не изменялась, когда ее выращивали после сои, по сравнению с выращиванием монокультуры кукурузы. Коуплэнд и Крукстон (1992) также подсчитывали использование питательных веществ по сое и кукурузе под влиянием эффекта чередования. Урожайность сои после кукурузы увеличивалась даже без использования большего количества питательных веществ, в то время как кукуруза после сои требовала больше питательных веществ и влаги для увеличения урожайности. Кукуруза и соя по-разному реагировали на эффект чередования культур.
3. Влияние предшествующей культуры на устойчивость к воздействию сорняков
Наши данные показывают, что эффективность использования влаги посевами некоторых культур может быть улучшена предшествующими культурами. Поскольку сорняки конкурируют с культурами за влагу, нам было интересно узнать, будут ли последовательности культур, улучшающие ЭИВ, также увеличивать устойчивость культуры к воздействию сорняков. Поэтому мы провели ряд исследований, в которых проводилась оценка влияния предшествующей культуры на устойчивость озимой пшеницы, кукурузы и сои к сорнякам в районах, где ежегодное количество осадков в среднем составляет 580 мм.
Исследовались виды сорняков, которые являются показательными по равномерному воздействию на эти культуры. Щетинник итальянский (просо итальянское) изучали в посевах кукурузы и сои, поскольку виды Setaria являются преобладающими видами на этих культурах повсеместно в США. Показательным видом для озимой пшеницы была рожь посевная (Secale cereale L.) – распространенный вид в посевах озимой пшеницы (White et al., 2006). Во всех исследованиях каждый участок был разделен на две зоны: без сорняков и засоренный участок для дальнейшего определения точности при оценивании синергизма культур. Сорняки на свободных от сорняков участках контролировали с помощью гербицидов и ручной прополки. Исследования проводились на полях, на которых интенсивное управление сорняками в предыдущие годы привело к низкой густоте стояния местного сорного сообщества.
Управление питательными веществами для кукурузы и озимой пшеницы включало в себя использование стартовых удобрений (N, P, K) с последующим внесением азота по культуре в течение вегетационного периода. Для сои вносились только стартовые удобрения. Количество азотного удобрения было откорректировано с учетом предшествующих культур, таких как бобовые, которые привносят азот в почву. Все культуры были посеяны по No-till-технологиям, а на исследуемых участках No-till применялся в течение 5 лет до начала исследования.
3.1 Озимая пшеница
В нашем первом исследовании мы сравнивали, как влияет соя, яровая пшеница, горох и смесь овса и гороха (фураж) на устойчивость озимой пшеницы к воздействию ржи посевной (Anderson, 2009с). Мы выращивали быстросозревающий сорт сои, которую убирали в конце августа, что давало возможность посеять озимую пшеницу в один и тот же день на всех участках. Травостой ржи посевной был сформирован в озимой пшенице за счет ручного посева – 15 растений на квадратный метр через 3 дня после всходов озимой пшеницы.
При наличии предшествующей культуры урожайность озимой пшеницы отличалась как в условиях без сорняков, так и на засоренных участках (график 2). При наличии ржи посевной урожайность озимой пшеницы после гороха составила 4900 кг/га, а после сои либо яровой пшеницы – менее 3200 кг/га. В условиях выращивания без сорняков потери урожайности из-за ржи посевной составляли лишь 11%, когда озимую пшеницу выращивали после гороха, но уже 32% после сои. Даже в условиях без сорняков урожайность озимой пшеницы была на 12% выше после гороха, чем после сои. Поразительная тенденция: после гороха урожайность озимой пшеницы даже в присутствии ржи посевной была такой же высокой, как и после сои или яровой пшеницы, но без сорняков. Урожайность озимой пшеницы колебалась от 4 до 6,7 т/га в течение 4-х лет исследований, тем не менее влияние гороха было постоянным в течение всех этих четырех лет.
Мы добавили в исследование овес для подавления болезней корней озимой пшеницы. Однако в плане улучшения устойчивости озимой пшеницы к ржи посевной смесь овса и гороха не была такой же благоприятной, как чистый горох (график 2). Когда озимая пшеница выращивалась после смеси овса и гороха, потери урожайности из-за воздействия ржи посевной составили 21%. Овес, очевидно, мешал механизму синергизма между горохом и озимой пшеницей, поскольку устойчивость ко ржи посевной оказалась меньше по сравнению с тем вариантом, когда озимая пшеница шла после одного гороха.
Более низкая урожайность озимой пшеницы, выращиваемой после пшеницы яровой, похоже, была обусловлена болезнями корней, поскольку бобовые культуры и овес снижают заболеваемость корней пшеницы по сравнению с выращиванием монокультуры пшеницы (Krupinsky et al., 2002). Однако мы были удивлены разницей и в урожайности озимой пшеницы, и в устойчивости к сорнякам в тех случаях, когда в качестве предшествующих культур использовали горох и сою (график 2). Разница в уровнях почвенной влаги после уборки предшествующих культур может быть одним из факторов, однако обильные осадки выпадают на этом участке осенью и зимой. Соответственно, когда начинается весенний рост, почвенный профиль обычно находится в нормальной полевой влагоемкости, исключая разницу в уровнях почвенной влаги среди предшествующих культур.
Еще одним фактором мог бы быть круговорот азота, различный после гороха и сои, но высота растения озимой пшеницы в период цветения и биомасса ржи посевной, измеренные при уборке озимой пшеницы, не отличались из-за предшествующей культуры. Очевидно, что горох влияет на рост озимой пшеницы уникальным способом, снижая влияние ржи посевной.
Устойчивость яровой пшеницы к сорнякам также отличается в зависимости от предшествующей культуры. Чечевица (Lens culinaris, Medic) усиливала устойчивость яровой пшеницы к местному сорному сообществу, но горчица (Brassica juncea L.) и рапс не усиливали. Авторы (Moyer et al., 2004) частично связывали этот эффект с изменениями в биологии почвы. Они также обратили внимание, что горчица подавляла рост яровой пшеницы, но не сорняков. Таким образом, некоторые последовательности культур могут благоприятствовать появлению сорняков. Чечевица может оказывать синергетическое воздействие на яровую пшеницу. Как сообщил Кэйси с соавторами, чечевица увеличивала ЭИВ пшеницей на 12% по сравнению с паром или другими культурами.
3.2 Кукуруза
Для дальнейшего изучения синергизма культур и влияния сорняков мы провели сравнение того, как пять культур – соя, рапс, кукуруза, яровая пшеница и горох – влияли на устойчивость кукурузы к воздействию щетинника итальянского. Семена щетинника итальянского были разбросаны на поверхности почвы за день до посева кукурузы; травостой щетинника итальянского составлял примерно 115 сеянцев/м2 в травостое кукурузы. Масса щетинника итальянского в сыром виде была измерена через 7 недель после всхожести и не отличалась при использовании разных предшественников.
Кукуруза была наиболее устойчива к щетиннику зеленому, если выращивалась после сои, при этом урожайность зерна уменьшалась на 50% (график 3). Для сравнения: кукуруза после сои, рапса или яровой пшеницы теряла почти 80% своей урожайности из-за присутствия щетинника итальянского. Кукуруза после кукурузы давала урожайность в 205 кг/га, когда в поле имелся щетинник итальянский, или только 7% от урожайности, наблюдаемой при выращивании после гороха.
Существенное снижение урожайности кукурузы, выращиваемой после кукурузы, включает в себя аллелопатическое воздействие растительных остатков кукурузы на замедление роста сеянцев в течение первых 5-6 недель роста. Производители, которые выращивают монокультуру кукурузу в данном регионе, сводят к минимуму этот аллелопатический эффект за счет механической обработки, чтобы растворить токсические соединения. Они также разгребают растительные остатки из посевного ряда с помощью специальных приспособлений на сеялке (Vetsch and Randail, 2000).
Даже при отсутствии сорняков урожайность кукурузы была существенно больше при ее посеве после гороха, чем после других культур (график 3). Кукуруза после яровой пшеницы, сои или рапса давала урожайность на 13% меньше, чем кукуруза после гороха. А урожайность монокультуры кукурузы составляла лишь 57% от урожайности кукурузы после гороха. Любопытная тенденция прослеживалась в том, что кукуруза после гороха и при наличии щетинника итальянского имела такую же урожайность, как кукуруза после кукурузы на участке без сорняков.
Люцерна (Medicago sativa L.) и подсолнечник также увеличивали урожайность зерна кукурузы по сравнению с урожайностью кукурузы после кукурузы в условиях без сорняков. Однако обнаружилось, что рост урожайности кукурузы благодаря этим культурам-предшественникам не отличался от роста урожайности после сои. Горох, похоже, оказывает уникальное воздействие, увеличивая урожайность зерна и устойчивость кукурузы к сорнякам.
3.3 Соя
Пять культур мы также оценивали на предмет влияния на устойчивость сои к воздействию сорняков, следуя той же самой методологии, когда применялся щетинник итальянский в качестве сорняка для кукурузы (Anderson, 2009а).
Урожайность сои в условиях без сорняков была наивысшей после злаковых культур: овса, яровой пшеницы и кукурузы (график 4), что может быть обусловлено подавлением болезней корней сои. Но при наличии на участке щетинника итальянского урожайность сои была более высокой после кукурузы, чем после овса или яровой пшеницы. Снижение урожайности сои из-за воздействия щетинника итальянского составило лишь 18%, когда сою выращивали после кукурузы, но более 30%, когда предшественниками сои были яровая пшеница или овес. Повышенная устойчивость сои к сорнякам, когда ей предшествовала кукуруза, может быть связана с тем, что кукуруза улучшает ЭИВ соей и помогает сое выдерживать конкуренцию с сорняками за влагу.
Горох улучшал устойчивость сои к щетиннику итальянскому по сравнению с выращиванием монокультуры сои, но не повышал урожайность сои в условиях без сорняков (график 4). Горох, вероятно, взаимодействует с соей по-другому в условиях, когда нет сорняков, чтобы замаскировать синергетическую реакцию, когда в поле присутствует щетинник итальянский.
3.4 Синергизм и устойчивость к сорнякам
Наши результаты относительно устойчивости культур к сорнякам предоставляют еще одно доказательство синергизма; горох улучшал устойчивость озимой пшеницы и кукурузы к сорнякам, в то время как кукуруза благоприятно влияла на сою. Мы были заинтригованы, что и озимая пшеница, и кукуруза реагировали на горох более благоприятно, чем соя (графики 2 и 3).
Эта тенденция может оказаться аномальной, но мы полагаем, что горох улучшает эффективность использования влаги озимой пшеницей и кукурузой, чем сводит к минимуму воздействие сорняков. Как отмечалось ранее, горох улучшает ЭИВ озимой пшеницей по сравнению с другими культурами (Anderson, 2009a), в то время как Коуплэнд с соавторами (1993) обнаружил, что соя не улучшала ЭИВ кукурузой.
4. Горох изменяет взаимосвязь между густотой травостоя кукурузы и урожайностью зерна
Одним из преимуществ генетического усовершенствования кукурузы является повышенная устойчивость к стрессу густоты (Tollenaar et al., 2006). Благодаря этой улучшенной устойчивости урожайность кукурузы выше, если выше густота травостоя. Поскольку горох улучшает устойчивость к воздействию сорняков, мы предположили, что горох может также повысить урожайность кукурузы при более высокой густоте травостоя.
Чтобы проверить эту гипотезу, мы сравнивали урожайность кукурузы при пяти разных показателях густоты травостоя от 45 000 до 73 000 растений на гектар с промежутками в 7000 растений, посеянных в стерню гороха, сои и яровой пшеницы, выращивавшихся в предыдущий год. Обычная густота травостоя, которой придерживаются производители в данном регионе, составляет от 66 000 до 73 000 растений/га. Кукурузу выращивали с применением лучших методов управления в условиях без сорняков.
После сои или яровой пшеницы в качестве предшественников наивысшая урожайность кукурузы достигалась при 73 000 растений на гектар, при этом наблюдалось постепенное увеличение урожайности по мере увеличения густоты травостоя кукурузы (график 5). Однако была отмечена удивительная тенденция: когда кукурузу выращивали после гороха, наибольшая урожайность кукурузы достигалась при 52 000 растений на гектар, что на 21 000 растений меньше, чем когда кукурузу выращивали после сои или яровой пшеницы. Урожайность была в пределах 6500-9800 кг/га в течение трех лет исследований, но тенденции урожайности среди предшествующих культур были стабильными в течение этих лет. Затраты на семена являются основной статьей расходов производителей. Выявленное изменение в оптимальной густоте травостоя кукурузы, выращиваемой после гороха, может сократить затраты на семена более чем на 70$/га.
Мы не совсем уверены в том, почему горох повышал урожайность кукурузы при низкой густоте травостоя по сравнению с соей и яровой пшеницей. Возможно, дело в продуктивности кукурузы в результате влияния расположения растений в пространстве (Lindquist et al., 1994). Густота травостоя 52 000 растений на гектар приводила к тому, что растения кукурузы равномерно располагались внутри рядка и между рядками (ширина междурядья – 50 см).
Баллок с соавторами (1988) обнаружил, что кукуруза, выращиваемая на равноудаленном расстоянии между растениями, давала большую урожайность зерна на единицу площади, чем когда она выращивалась по традиционным нормам касательно расстояний между растениями. Уоткинсон (1980) также отметил, что эффективность удерживания и использования ресурсов наивысшая, когда растения расположены на равном расстоянии друг от друга.
Горох может еще больше повышать эффективность использования ресурсов, уменьшая тем самым необходимость в большом количестве растений.
5. Возможные причины синергизма между культурами
Несколько различных факторов могут принимать участие в синергизме культур, такие как изменения в запасах питательных веществ и влаги (Hobbs, 2007), подавление вредителей (Kirkegaard et al., 2008) или высвобождение ростостимулирующих веществ (Anaya, 1999).
Микробиологическое сообщество и микробиологические заражения культуры тоже изменяются при разнообразии культур. Одним из микробиологических видов, зачастую упоминаемых при влиянии чередования культур, являются микоризы, которые улучшают поглощение питательных веществ и влаги растениями. Чтобы получить представление о возможных причинах синергизма, мы провели дополнительные исследования, которые были направлены на изучение взаимосвязи между горохом и соей. Синергизм гороха заключается в изменении физиологии кукурузы, что повышает эффективность роста.
5.1 Синергизм гороха по отношению к кукурузе сохраняется два года
Нам также было интересно установить, сохранится ли благоприятное влияние гороха на кукурузу на второй год. Райт сообщил о том, что горох повышал урожайность зерна мелкозерновых в течение 2 лет. В первый год горох повышал урожайность ячменя на 21% по сравнению с выращиванием монокультуры ячменя. На следующий год яровая пшеница давала урожайность на 12% больше после последовательности горох – ячмень, чем после последовательности ячмень – ячмень. Крукстон с соавторами (1991) обнаружил подобную тенденцию по сое, когда благоприятное влияние кукурузы на урожайность сои тоже сохранялось в течение 2 лет. Для сравнения, благоприятное влияние сои на урожайность кукурузы длилось только 1 год.
Поэтому мы сравнивали урожайность кукурузы под влиянием последовательности культур и при наличии щетинника итальянского. Горох, соя и кукуруза выращивались в первый год, соя и все обработки во второй год, а на третий год – кукуруза во всех последовательностях. Щетинник итальянский выращивался в кукурузе только в течение третьего года, при этом каждый участок был разбит на две зоны – с сорняками и без сорняков.
Даже на второй год после гороха кукуруза была более устойчивой к воздействию сорняка. Урожайность кукурузы после гороха составила 4500 кг/га в последовательности горох – соя (при наличии щетинника итальянского), но менее 3200 кг/га после таких последовательностей как соя – соя и кукуруза – соя (график 7). Потери урожайности из-за воздействия сорняка после последовательности горох – соя составили 45%, в то время как потери урожайности после других последовательностей составили более 60%.
Синергизм также наблюдался в условиях без сорняков, поскольку урожайность кукурузы была на 8% выше в последовательности горох – соя – кукуруза, чем в последовательности кукуруза – соя – кукуруза.
5.2 Синергизм гороха по отношению к кукурузе больше в условиях низкой урожайности
Кукуруза является более устойчивой к стрессам из-за повышенной эффективности использования ресурсов, но эта реакция зачастую ощутима только в условиях стресса. Чтобы увидеть, был ли синергизм среди культур более ощутим в условиях стресса (низкая урожайность), мы сравнивали единый режим выращивания кукурузы после гороха и сои, который применялся в различных исследованиях в течение семи разных лет.
Наше исследование включало выращивание кукурузы с густотой травостоя 66 000 растений на гектар после гороха и сои; участки сохраняли без сорняков. В среднем за семь лет урожайность кукурузы была на 11% больше после гороха, чем после сои. Но увеличение урожайности благодаря гороху было выше в годы с низкой урожайностью. Урожайность кукурузы была на 4-5% выше после гороха в те годы, когда урожайность была выше 9000 кг/га. Для сравнения, увеличение урожайности составляло от 14 до 22%, когда урожайность кукурузы была менее 6500 кг/га.
Из-за сложности биологии почвы нам, видимо, не удастся определить конкретную причину синергизма культур. Тем не менее мы увидели тенденции, которые могут влиять на управление решениями.
Горох и кукуруза оказывают синергетическое влияние на последующие культуры. Но синергизм среди культур может быть редким; мы обнаружили, что лишь 2 из 11 протестированных культур оказывали такое воздействие.
Синергизм также относится к культурам в последовательности. Например, кукуруза оказывает синергетический эффект на просо и озимую пшеницу, но не на сорго (Sorghum bicolor (L.) Moench). Еще одним примером такой специфичности служит синергетическое влияние кукурузы на озимую пшеницу и просо. Кукуруза улучшала эффективность использования влаги просом в севообороте озимая пшеница – кукуруза – просо (ОП-К-Пр), но не тогда, когда в севооборот добавляли пар (ОП-К-Пр-П). ЭИВ просом не отличалась в севооборотах озимая пшеница – просо (ОП-Пр), озимая пшеница – просо – пар и ОП-К-Пр-П. Тем не менее кукуруза улучшила эффективность использования влаги озимой пшеницей в севооборотах ОП-К-П и ОП-К-Пр-П, включающих в себя пар (график 1). Основываясь на наших тенденциях и наблюдениях Толленаара и Ву, мы полагаем, что синергизм может быть более благоприятным именно в сухих регионах, где стресс растений является обычным явлением.
6. Преимущества синергизма для производства культур
Даже если причина синергизма не известна, измерение эффективности использования влаги культурами под влиянием предшествующей культуры может дать метод для определения потенциальных синергетических последовательностей. Подход ЭИВ является подходящим, потому что культура объединяет в себе различные факторы биологии почвы. После выявления синергетических последовательностей, производители культур будут включать в себя эти благоприятные последовательности, чтобы повысить эффективность производства.
6.1 Увеличение урожайности без увеличения затрат
Синергетические последовательности могут способствовать увеличению урожайности зерна при неизменных затратах ресурсов, как это показано на примере озимой пшеницы и кукурузы (графики 1-3). Польза от синергизма является особенно ощутимой в полузасушливом регионе Великих равнин. До появления No-till-технологий традиционным в регионе был севооборот озимая пшеница – пар. Успешным севооборотом, применяемым в No-till, является севооборот горох – озимая пшеница – кукуруза – просо (Г-ОП-К-Пр). Производители обнаружили, что потенциал урожайности озимой пшеницы в этом севообороте более чем в два раза выше. Урожайность озимой пшеницы редко превышает 2650 кг/га в самые благоприятные годы при выращивании в севообороте озимая пшеница – пар. Для сравнения, урожайность пшеницы может превышать 5400 кг/га в благоприятные годы в севообороте горох – озимая пшеница – кукуруза – просо.
Подобное изменение в потенциале урожайности происходит с просом в данном севообороте. Это увеличение потенциала урожайности озимой пшеницы и проса вдвое. Однако оно не наблюдалось в No-till-севооборотах без синергетических последовательностей, таких как ОП-Пр или ОП-Пр-Под-П. Часть увеличения урожайности зерна в севообороте горох – озимая пшеница – кукуруза – просо может быть обусловлена улучшением использования влаги культурами при No-till-технологиях: урожайность озимой пшеницы и проса на 35-50% больше, чем ожидалось, исходя из запасов влаги и питательных веществ.
Мы полагаем, что благодаря гороху и кукурузе повышается урожайность озимой пшеницы и проса за счет улучшения эффективности использования влаги и питательных веществ.
Благодаря успешному применению севооборота горох – озимая пшеница – кукуруза – просо производители задаются вопросом: нельзя ли создать такие севообороты, в которых все последовательности синергетические.
Вообще говоря, синергетические последовательности довольно редки, поэтому интересно выяснить, сможет ли синергизм быть продлен до уровня севооборота. Покровные культуры могут предоставить такой вариант. Севооборот горох – озимая пшеница – кукуруза – просо содержит две синергетические последовательности (график 6). Озимая пшеница не оказывает синергетическое влияние на кукурузу (Schneekloth et al., 1991), но, как обнаружил Кумар с соавторами, вика мохнатая (Vicia villosa Roth), выращиваемая в качестве покровной культуры, повышает устойчивость помидоров (Solanum lycopersicum L.) к стрессу. Если бы существовала синергетическая покровная культура для кукурузы, производительность севооборота горох – озимая пшеница – кукуруза – просо (Г-ОП-К-Пр) может быть заметно усилена за счет включения этой покровной культуры в севооборот между озимой пшеницей и кукурузой (график 6). Бидербек с соавторами (2005) сообщил, что было бы достаточно коротких интервалов выращивания покровных культур для инициирования благоприятных изменений в микробиологии почвы.
Управление сорняками также может выиграть от синергетических последовательностей. Благодаря разнообразию культур и No-till-технологий производители могут управлять сорняками с помощью экологического подхода, который включает в себя агротехнические методы уменьшения динамики популяции сорняков с использованием гербицидов (Anderson, 2005).
Плотность сорного сообщества уменьшается со временем таким образом, что для некоторых культур уже не нужно использовать гербициды. Производители контролируют сорняки с использованием на 50% меньшего количества гербицидов, чем при традиционном управлении сорняками. Синергизм между культурами будет усиливать современный подход к управлению сорняками: уменьшая влияние сорняков и применение гербицидов, обеспечивая низкую засоренность и высокую урожайность культур.
Рэнди Андерсон,агроном-исследователь,Сельскохозяйственный исследовательский центр,Брукингс, Южная Дакота, США
Опубликовано в №09 2012 г.
www.zerno-ua.com
Синергизм и антагонизм | HarvestDiary
Для хорошего роста и развития растениям, да и не только растениям, но и всему живому, нужно сбалансированное питание. Чтобы обеспечить растениям сбалансированность элементами, необходимо иметь точные знания о физиологической роли каждого питательного элемента, благодаря чему можно понять, каким будет оптимальный срок его внесения. Важным есть и учет взаимодействия различных элементов в организме растения между собой, ведь именно это поможет улучшить условия развития растений. Между различными элементами питания может возникать одно из двух явлений:
· Синергизм – усиление одним элементом действия другого;
· Антагонизм – подавление одним из элементов другого.
То, какое явление будет преобладать, зависит от физико-химических свойств элементов, особенностей почвы и культур, но его несбалансированные реакции могут стать причиной химических стрессов у растений.
Антагонизм
Антагонизм – явление, которое предусматривает торможения поступления одного иона через добавление другого. Оно возникает в том случае, когда совместное действие двух элементов меньше суммы действующих элементов по одиночке. Иными словами, при антагонизме велик объем усвоения растением одного элемента питания, способного привести к дефициту в растительном организме другого питательного элемента. Действие антагонизма проявляется в том, что во время поглощения растением различных элементов, их катионы конкурируют между собой. Антагонизм может проходить двумя путями: так микроэлемент может блокировать поглощение микроэлемента, или микроэлемент ингибирует поглощение макроэлементов.
Наиболее остро антагонизм проявляется тогда, когда ионы являются одноименно заряженными, а также в тех случаях, когда в грунтовом растворе концентрация одних ионов является большей, чем концентрация других.
Ярким примером антагонистического влияния можно назвать реакцию у фосфатов. Таким образом, внесение и усвоения растением в больших количествах фосфора (Р) полностью блокирует усвоение ею кальция (Са) и магния (Мg).
Синергизм и его особенности
Кроме антагонизма, известно и явление синергизма – когда действие одних ионов значительно усиливает влияние других ионов. Наблюдается явление в том случае, когда совместное действие элементов питания выше, чем сумма влияния каждого из них в отдельности. Однако, кроме вышеупомянутого, положительного синергизма, существует и отрицательный – когда токсическое воздействие на растительный организм одной соли значительно усиливает токсичность другой.
Наблюдается синергизм как между катионами и анионами, которые имеют разный заряд, так и между заряженными одноименно. Однако последний вариант чаще всего возможен тогда, когда в почвенном растворе их содержание является достаточно небольшим.
Хорошим примером синергизма является одновременное внесение азота (N) с фосфором (Р), калием (К), кальцием (Са) или магнием (Мg), ведь эти элементы хорошо дополняют действие друг друга и способны улучшить усвоение растением каждого из них.
Кроме элементов антагонистов и синергистов выделяют элементы, которые способны блокировать друг друга. Это можно увидеть из таблицы: при одновременном внесении Zn и Ca, или Ca и Cu, растение будет усваивать только один элемент, например, Zn, или Ca или или Ca, или Cu.
Все особенности антагонистических и синергических явлений, которые возникают при сочетании различных элементов питания, необходимо учитывать во время разработки стратегии питания выращиваемых культур. Ведь неправильное сочетание компонентов питания может привести к негативным последствиям, которые в конечном итоге изменят урожайность культуры в худшую сторону.
Видео по теме :
harvestdiary.com.ua
Уравновешенность питательного раствора, синергизм и антагонизм ионов
Питательный раствор представляет собой водный раствор веществ, необходимых растению для жизни и роста. Находясь в естественных условиях, растения получают их непосредственно из почвы, через корневую систему.
Почвенный раствор можно определить также как жидкую фазу почв, включающую почвенную воду, содержащую растворенные соли, органоминеральные и органические соединения, газы и тончайшие коллоидные золи. В. И. Вернадский считал почвенные растворы одной из важнейших категорий природных вод, «основным субстратом жизни», «основным элементом механизма биосферы».
Почвенным (питательным) раствором называют влагу, находящуюся в почве и содержащую в растворенном состоянии органические и минеральные вещества и газы. Почвенный раствор образуется в результате взаимодействия воды, поступающей в почву, с ее твердой фазой и растворения некоторых органических и минеральных веществ и их производных.
По составу и концентрации почвенного раствора все почвы делят на две группы — незасоленные и засоленные. Незасоленными называют такие почвы, в которых концентрация почвенного раствора невелика и сухой остаток водной вытяжки не превышает 0,25%. К засоленным относят почвы с высокой концентрацией почвенного раствора и сухим остатком водной вытяжки, превышающим 0,25%.
Состав почвенного раствора в незасоленных почвах определяется характером и интенсивностью биологических процессов, особенностями материнских пород и почвообразования, составом обменных катионов почвы. В почвенном растворе этих почв находятся как минеральные, так и органические соединения.
Из минеральных соединений наиболее распространены бикарбонаты кальция и магния — Са(НСОз)2, Mg(HCO3 )2, в меньшей степени встречаются КНСО3 и NaHCO3, сульфаты кальция, магния, калия и натрия, нитраты и фосфаты этих же катионов.
Органические соединения почвенного раствора представлены различными кислотами (щавелевой, винной и др.) и их солями, а также водорастворимыми гумусовыми веществами, ведущее место среди которых принадлежит фульвокислотам и фульватам одно- и двухвалентных катионов. В кислых почвах, кроме того, в состав раствора входят фульваты железа и алюминия.
В засоленных почвах состав и концентрация почвенного раствора зависят от состава и количества легкорастворимых солей, находящихся в самой почве. Преобладающее значение в составе таких почв имеют минеральные соединения, из которых чаще всего присутствуют следующие: хлориды — NaCl, CaCl2, MgCl2, КCl, сульфаты — Na2 SO4, MgSO4, CaSO4, карбонаты — Na2 CO3, и MgCO3; бикарбонаты — NaHCO3, Mg(HCO3 )2, Са(НСОз )2. Из органических соединений в незначительном количестве встречаются гуматы одновалентных катионов.
Состав и концентрация почвенного раствора как в засоленных, так и в незасоленных почвах, непостоянны, и изменяются в течение вегетационного периода. Это объясняется динамикой микробиологических процессов, различной интенсивностью усвоения питательных веществ растениями в течение вегетации, вымыванием растворенных соединений осадками или, наоборот, подъемом их с грунтовыми водами.
Физическое состояние почвенного раствора неоднородно: часть его находится в форме пленочной влаги и недоступна растениям, остальная часть — в виде капиллярной и гравитационной влаги и легко усваивается растениями.
Почвенный раствор обладает определенным осмотическим давлением. У незасоленных почв оно составляет 0,2–0,3 МПа, у засоленных часто превышает 1–2 МПа.
Важное свойство почвенного раствора — его реакция, по характеру которой выделяют кислые, нейтральные и щелочные растворы. Кислая реакция определяется наличием в растворе органических и минеральных кислот и кислых солей, щелочная — карбонатами и бикарбонатами натрия, кальция и магния.
Почвенный раствор служит основным и непосредственным источником элементов питания для растений, так как из него растения усваивают большинство питательных веществ. Он создает определенную среду, в которой развиваются микроорганизмы, и служит, поэтому основным регулятором микробиологической деятельности.
Но в ряде случаев почвенный раствор может оказать и неблагоприятное воздействие на условия жизни организмов и почвообразовательный процесс.
При высокой концентрации почвенного раствора наступает так называемая физиологическая сухость, когда имеющиеся в растворе элементы питания не могут быть усвоены микроорганизмами и растениями. Объясняют это тем, что осмотическое давление концентрированного почвенного раствора значительно превышает осмотическое давление сока в клетках корней или плазме микроорганизмов.
Кислая и щелочная реакции почвенного раствора подавляют развитие и деятельность микроорганизмов. К этому же приводит и присутствие в почвенном растворе ряда соединений, которые даже при малой концентрации губительны для растения. К таким соединениям относятся сода, сероводород и закисные формы железа.
Почвенный раствор играет большую роль в почвообразовании. Так, кислая его реакция способствует подзолообразованию, а высокая концентрация легкорастворимых солей приводит к образованию солончаков или солончаковатых почв.
Концентрацию почвенного раствора уменьшают промыванием почвы пресными водами. Состав его изменяют внесением удобрений, а реакцию — известкованием или гипсованием.
Для выделения почвенного раствора используют методы: выжимание раствора под давлением на специальных прессах, центрифугирования и замещения (вытеснения) другой жидкостью. Количество выделяющегося почвенного раствора зависит от водоудерживающих свойств почвы и степени ее увлажнения. Получение почвенных растворов центрифугованием возможно лишь в почвах с влажностью, близкой к полной влагоемкости. Выделение почвенного раствора замещением его другой жидкостью заключается в том, что через колонку, заполненную исследуемой почвой с естественной влажностью, сверху просачивается вытесняющая жидкость. Наиболее удобен для этой цели этиловый спирт. Почвенный раствор собирается в приемник. Для улучшения фильтрационных свойств тяжелых почв их рекомендуется смешивать с хорошо отмытым кварцевым песком. При использовании указанных методов после выделения раствора в почве остается еще некоторое количество влаги.
Преимущество указанных методов — возможность получения растворов при влажности, характерной для почв в вегетационный период, поэтому практически динамику почвенного раствора можно изучить лишь этими методами.
Состав жидкой фазы почвы в почвоведении также изучают лизиметрическим методом. Этот метод основан на исследовании просачивающихся через определенную толщу почвы дождевых или талых вод, которые собирают в специальный приемник. Недостаток всех лизиметрических установок — возможность получения растворов лишь в периоды сильного увлажнения почв.
Все методы выделения почвенных растворов трудоемки и не получили широкого распространения в практике научных исследований, кроме лизиметрических стационаров.
Некоторое приближение к познанию состава почвенных растворов дает метод извлечения солей из почвы водной вытяжкой в соотношении почва: вода = 1:5. Простота и доступность метода водной вытяжки сделала его массовым при определении засоленности почв и содержания водорастворимых элементов питания растений.
Сравнительное представление о составе почвенного раствора и водной вытяжки из солончака дают следующие данные в м.-экв. на 100г почвы (табл. 1).
Таблица 1
| Вытяжка | Раствор | |
| Cl- | 39,4 | 42,4 |
| SO42- | 14,1 | 7,0 |
| Na+ | 37,0 | 36,4 |
| Mg2+ | 12,0 | 12,6 |
| Ca2+ | 4,6 | 0,6 |
biofile.ru
| ВЗАИМОДЕЙСТВИЕ МАКРО- И МИКРОЭЛЕМЕНТОВ В РАСТЕНИЯХ. АНТАГОНИЗМ И СИНЕРГИЗМ Сбалансированность химического состава живых организмов - основное условие их нормального роста и развития. Взаимодействие химических элементов имеет такое же значение для физиологии растений, как явления дефицита и токсичности. Взаимодействие между химическими элементами может быть антагонистическим или синергическим, и его несбалансированные реакции могут служить причиной химических стрессов у растений. Антагонизм возникает, когда совместное физиологическое действие одного или более элементов меньше суммы действия элементов, взятых по отдельности, а синергизм - когда совместное действие больше. Такие взаимодействия можно связать со способностью одного элемента ингибировать или стимулировать поглощение других элементов растениями. Все эти реакции весьма переменчивы. Они могут происходить внутри клеток, на поверхности мембран, а также в среде, окружающей корни растений. Процессы взаимодействия контролируются многими факторами, и их механизмы еще плохо изучены, хотя некоторые данные все же имеются. Взаимодействия между макро- и микроэлементами, сведенные в табл. 1, ясно показывают, что Са, Р и Mg - главные антагонистические элементы в отношении поглощения и метаболизма многих микроэлементов. Однако и для антагонистических пар элементов наблюдались иногда синергические эффекты, что связано, вероятно, со специфическими реакциями у отдельных генотипов или видов растений. Таблица 1. Взаимодействие между макро- и микроэлементами в растениях
Для практического применения наиболее важно антагонистическое действие Са и Р на такие опасные для здоровья человека тяжелые металлы, как Be, Cd, Pb и Ni. Примечательно, что, хотя антагонистическое действие Р и Са на многие микрокатионы и микроанионы часто рассматривается в литературе, об антагонистическом воздействии Mg на микроэлементы имеются только случайные работы. Взаимодействия между микроэлементами, наблюдающиеся в самих растениях, также показывают, насколько сложны эти процессы, так как они могут быть то антагонистическими, то синергическими. Иногда они проявляются в метаболизме более чем двух элементов. Наибольшее число антагонистических реакций наблюдалось для Fe, Mn, Cu, и Zn, которые, очевидно, являются ключевыми элементами в физиологии растений. Функции этих микроэлементов связаны с процессами поглощения и с энзиматическими реакциями. Среди остальных микроэлементов в антагонистических отношениях к этой четверке часто оказываются Сr, Мо и Se. Синергическое взаимодействие между микроэлементами обычно не наблюдается. Синергизм Cd с такими микроэлементами, как Pb, Fe и Ni, может быть артефактом, возникающим вследствие разрушения физиологических барьеров под действием стресса, вызванного избыточными концентрациями тяжелых металлов. Кроме того, некоторые реакции, происходящие в среде, окружающей корни, и влияющие на потребление микроэлементов корнями, по-видимому, не связаны непосредственно с метаболическими взаимодействиями, однако эти два типа реакций нелегко различить. ИСТОЧНИК: Kabata-Pendias, Alina. Trace elements in soils and plants / Alina Kabata-Pendias. -- 4th ed. – 2010.; Кабата-Пендиас А., Пендиас X. Микроэлементы в почвах и растениях: Пер. с англ. - М.: Мир, 1989. - 439 с, |
stom.tilimen.org
| Для нормальной жизнедеятельности как растительных, так и животных организмов в окружающей их среде должно быть определенное соотношение различных катионов. Чистые растворы солей одного какого-либо катиона оказывают на организм, в том числе и на растительный, губительное влияние, т. е. являются ядовитыми. Причем это влияние проявляется еще до того, как начинают обнаруживаться симптомы голодания растений относительно недостающих элементов питания. При культивировании зародышей рыб в растворе хлористого натрия развиваются уродливые одноглазые особи. У морских ежей, помещенных в растворы хлористого натрия, наблюдается партеногенез. Все это происходит несмотря на то, что NaCl взят в той же концентрации, как и в морской воде. Сходные результаты получены на корнях растений. Так, при помещении проростков пшеницы на чистые растворы КСl или СаСl2 на корнях появлялись вздутия, в затем корни отмирали. Интересно, что в смешанных солевых растворах, содержащих два разных катиона, ядовитое действие не наблюдается. Смягчающее влияние, оказываемое одним катионом на действие другого катиона, называют антагонизмом ионов. Антагонизм ионов проявляется как между разными ионами одной валентности (например, Na+ и К+), так и между ионами разной валентности (К+ и Са2+). При этом в последнем случае антагонистическое влияние сказывается более резко. Так, для того чтобы устранить ядовитое влияние чистой соли КСl, надо прибавить NaCl 30%, а СаСl2 всего 5%. Растворы, которые характеризуются определенным соотношением катионов, благоприятным для роста и развития организмов, называют уравновешенными. К естественным уравновешенным растворам относят, например, морскую воду, плазму крови. Антагонизм — это лишь одно из проявлений взаимного влияния ионов. В целом ряде случаев добавление одного иона угнетает поступление другого. Так, известно торможение поступления железа и связанное с этим торможение синтеза хлорофилла при избытке в среде ионов Мп2+. Конкурентные отношения в процессе поступления обнаружены и для ряда других катионов и анионов. Так, бор усиливает поступление катионов и снижает поступление анионов. Вместе с тем имеются наблюдения, когда действие одного иона усиливает влияние другого. Это явление получило название синергизма. Так, под влиянием фосфора усиливается положительное влияние молибдена. Вопрос о физиологических причинах антагонистического действия ионов до настоящего времени до конца не изучен. Одна из причин этого явления — их влияние на коллоидно-химические свойства цитоплазмы, в частности на гидратацию белков, входящих в ее состав. Известно, что двухвалентные катионы (Са2+ и Mg2+) дегидратируют коллоиды сильнее, чем одновалентные (Na+ и К+). Неодинаково влияние на гидратацию коллоидов и катионов сходной валентности. Так, натрий оказывает более сильное дегидратирующее влияние по сравнению с калием. Сходное влияние в действии отдельных катионов проявляется и на неживых отрицательно заряженных коллоидах. Коагуляция коллоидного раствора под влиянием чистой соли происходит при меньшей ее концентрации по сравнению с концентрацией смешанных растворов, содержащих две соли. Вместе с тем высказываются соображения, что антагонизм ионов в большой степени объясняется их конкуренцией за активные центры ферментов. Так, активность некоторых ферментов дыхания ингибируется Na+ и это снимается добавлением К+. Изменения интенсивности поступления одного иона под влиянием другого могут быть связаны с конкуренцией за места связывания их с переносчиками через плазмалемму во внутреннее пространство клетки. Вопрос о взаимовлиянии ионов заслуживает дальнейшего изучения, тем более что неблагоприятное влияние повышенной концентрации одного катиона может проявляться и в естественных условиях (в почве) и должно учитываться при внесении удобрений. Таким образом, для нормального роста растений необходимо определенное сочетание солей одно- и двухвалентных катионов.
|
fizrast.ru
Синергизм в косметике
Синергизм. Это слово произошло от греческого syneros и означает содействие, сотрудничество. Если мы обратимся к различным толковым словарям, то узнаем, что:
• в медицине термином синергизм обозначают совместное функционирование различных органов;
• в фармакологии – совокупное действие лекарственных веществ, каждое их которых усиливает эффект действия другого;
• в генетике и биологии – взаимное действие разнообразных физиологических и биохимических факторов, которое обуславливает оптимальный конечный эффект;
• в политологии - комбинированное действие на политические, экономические, социальные организации, при котором итоговый эффект превосходит действие, которое совершается каждым компонентом в отдельности.
То есть синергизм – это совместное действие элементов системы в одном направлении, усиление одного элемента другим элементом.
Наше предприятие ООО «КоролёвФарм» занимается производством косметической продукции. И нам интересно знать, как взаимодействуют различные ингредиенты в готовом продукте, сочетание каких видов сырья надо вводить в косметическую продукцию, чтобы получить больший эффект от комбинации используемых компонентов, чем от каждого в отдельности.
Но также надо учитывать и то, что помимо синергизма между ингредиентами возможно возникновение и отрицательного взаимного влияния (антагонизма), которое может привести к ослаблению эффектов от каждого из компонентов.
Рассмотрим эту проблему на косметических кремах.
Все ингредиенты, входящие в состав косметических средств, можно отсортировать по нескольким группам:
1. Вещества, которые создают структуру крема - эмульсию. В первую очередь, это растительные масла, которые вводятся в воду, тщательно перемешиваются (гомогенизируются) до однородного состояния. Если частички масла равномерно распределены в воде, то тогда обеспечивается хорошая впитываемость крема в кожу, в том числе и масляных составляющих, которые при обыкновенном нанесении не очень-то стремятся проникнуть в кожу. В настоящий момент существует оборудование, на котором процесс гомогенизации достиг высочайшего уровня, и на котором можно производить нанокрема с размером частиц меньше 15,7 нм. За счет малого размера частицы преодолевают барьер проницаемость кожи, проникают глубже основной массы клеточных слоев кожи и достигают уровня сосудистой (капиллярной) сети. А именно в частичках эмульсии крема как в растворителе или переносчике и содержатся все другие ингредиенты.
Помимо растительных масел в состав эмульсии вводятся и другие компоненты: эмоленты, стабилизаторы, загустители, эмульгаторы,- которые вводятся в состав продукта по технологической необходимости, обеспечивают легкость процесса эмульгирования, стабильность эмульсии.
2. Эстетические добавки – красители, ароматизаторы.
3. Вещества, необходимые для «долгожительства» косметических продуктов – консерванты.
Все выше упомянутые вещества являются базой для косметических кремов. Но не о них сейчас речь!
Те задачи и функции, которые возлагаются на крем, выполняют так называемые активные компоненты. В основном это витамины, растительные экстракты, растительные и эфирные масла, вытяжки, антиоксиданты. Чтобы крем был более эффективным, необходимо чтобы он содержал всё это разнообразие видов активных ингредиентов, по 2 – 5 наименований по каждому виду. Чем разнообразнее состав активных составляющих крема, тем питательнее он будет для клеток кожи. При выборе компонентов надо учитывать эффект синергизма, который позволит не только сделать косметическое средство более питательным, но и уменьшить процент их ввода в состав крема.
Синергизм витаминов.
Примером синергизма может служить взаимодействие витаминов. В состав косметических кремов обычно входит несколько витаминов. Например, витамины С и Е практически всегда входят в состав кремов парой, они очень хорошо усваиваются, дополняя и усиливая действия друг друга.
Их сподвижником является витамин А. Витамин Е оберегает витамин А от окисления в тканях, поэтому в состав косметической продукции они, как правило, вводятся парой. Комбинация витаминов А и Е дает исключительный защитный и оздоровительный эффект, отсрочивает старение и делает возможным создание мощной антиоксидантной защиты.
Витамин В5, который принимает участие в регуляции липидного обмена, особенно результативен в сочетании с витаминами С, В2, D.
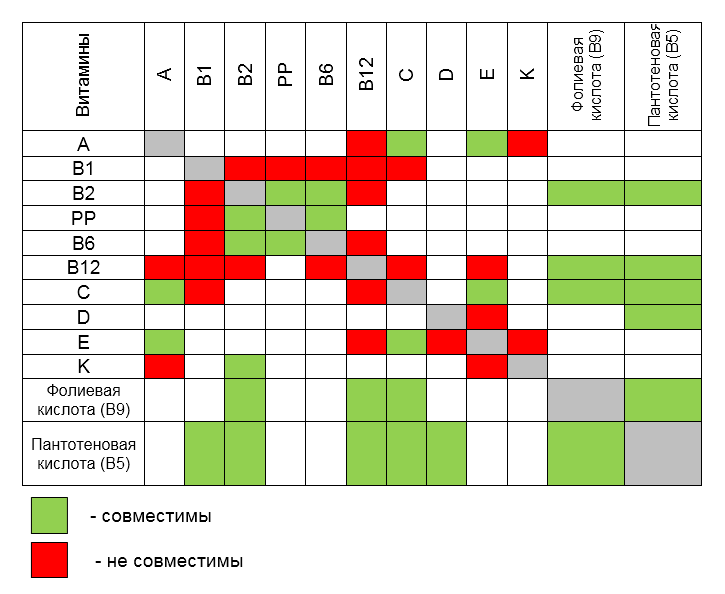
В таблице, которая приведена ниже, отображен не только эффект синергизма между различными витаминами («совместимы»), но и антагонизма («не совместимы»).
Синергизм антиоксидантов
Антиоксиданты — это вещества, которые охраняют организм от свободных радикалов и активных форм кислорода. Это главные стражи молодости и здоровья.
Они вводятся в косметические крема для выполнения сразу нескольких задач:
• для предотвращения окисления масел, которые входят в состав кремов, содержат полиненасыщенные жирные кислоты и которые имеют тенденцию легко окисляться;
• для повышения выработки коллагена и, как следствие, улучшения эластичности кожи;
• для эффективного снижения повреждений от солнечных лучей (устраняют фактор фитостарения).
Самые популярные антиоксиданты: витамины А, С, Е, РР, К, β-каротин (провитамин А), флавоноиды, танины, антоцианы, биофлавоноиды, коэнзим Q10, серосодержащие аминокислоты, селен и другие, а также растительные и ягодные экстракты таких растений как алоэ, зеленый чай, конский каштан, черника, эдельвейс, розмарин, облепиха, красный виноград, гранат, клюква, сосна, рябина, гингко билоба, гамамелис и многих-многих других.
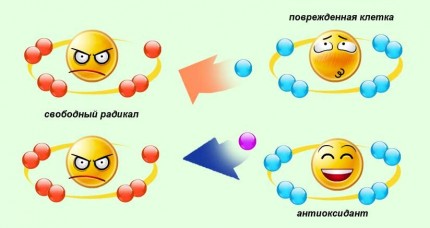
При взаимодействии молекулы антиоксиданта со свободными радикалами первая теряет электрон, разрушается и «выходит из игры».
Поэтому нужны восстанавливающие средства, которые переведут «испорченную» молекулу антиоксиданта в рабочее состояние. Витамин С «возвращает жизнь» витамину Е, но сам при этом окисляется. Соединения, содержащие серу, «реанимируют» витамин С, а биофлавоноиды активно восстанавливают и витамин Е, и витамин С. Подобный синергизм существует и промежду витамином Е и каротиноидами, промежду витамином Е и селеном (витамин Е необходим для сохранения биологических форм селена в активном состоянии). Равным образом селен снижает потребность в витамине Е, поддерживая его уровень в крови.
Коэнзим Q10 (убихинон), представляющий собой витаминоподобный фермент, обладает антиоксидантными свойствами, уменьшает разрушение коллагена, является генератором энергии, снижает утерю кожей гиалуроновой кислоты, а в синергизме с витамином Е работает еще эффективнее, сильнее.
Таким образом синергизм антиоксидантов дает возможность достичь максимальной защиты и высокой стабильности продукта при более низкой концентрации антиоксидантов
Синергизм масел.
 Большой сегмент среди активных ингредиентов косметических кремов занимают масла. Они помогают решить многие проблемы, возникающие с кожей: одни из них обеспечивают питанием сухую кожу и увлажняют ее, другие восстанавливают работу сальных желез, сужают поры, третьи – тонизируют кожу, делают ее упругой, эластичной. Продолжать можно и дальше.
Большой сегмент среди активных ингредиентов косметических кремов занимают масла. Они помогают решить многие проблемы, возникающие с кожей: одни из них обеспечивают питанием сухую кожу и увлажняют ее, другие восстанавливают работу сальных желез, сужают поры, третьи – тонизируют кожу, делают ее упругой, эластичной. Продолжать можно и дальше.
Растительные масла – кладовая микроэлементов, витаминов, ненасыщенный жирных кислот, которые так необходимы для кожи.
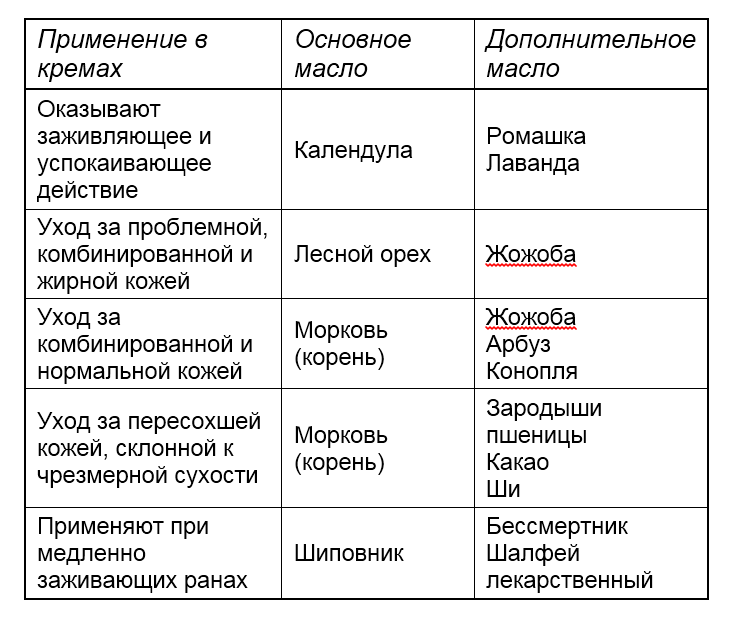
Список масел, которые вводятся в косметические средства, огромен, все они обладают различными свойствами. И, конечно, хочется, чтобы смесь масел, которую мы вводим в крем, сочетала в себе сразу несколько свойств. Существуют так называемые базовые масла, процентное содержание которых должно быть в смеси больше. Они легко впитываются, не забивают поры. А есть дополнительные масла. И так же, как в случае с витаминами, антиоксидантами, необходимо учитывать синергизм масел, когда они помогают друг другу, и антагонизм, когда масла препятствуют положительному взаимному действию.
Приведем несколько примеров синергии растительных масел:
При разработке рецептур кремов и других косметических продуктов технологам всегда надо помнить как о синергизмевводимых компонентов, чтобы получить желаемый эффект при меньших материальных затратах, так и об антагонизме ингредиентов, чтобы не получить обратного результата: отрицательное взаимодействие вводимых компонентов и, как следствие, снижение эффективности косметического средства.
www.korolevpharm.ru










