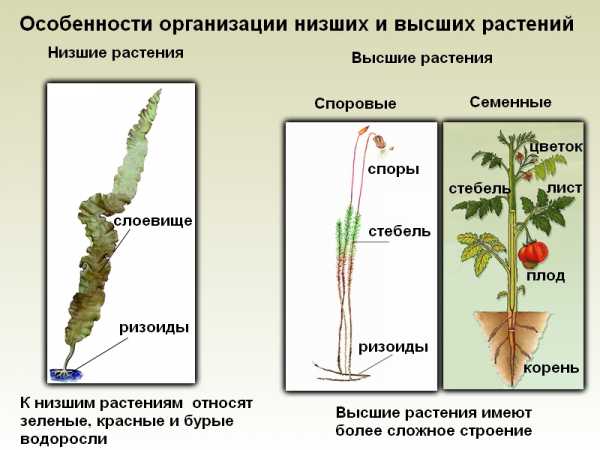
Симбиоз корней с бактериями. Симбиоз бобовых растений с клубеньковыми бактериями
Симбиоз корней с бактериями.
Помимо микоризы (микосимбиотрофии), которая часто встречается в природе, существует другой симбиоз, который встречается не так широко, как первый. Это симбиоз корней растения с бактериями (бактериосимбиотрофия).
Наиболее часто у бобовых растений, но иногда и у некоторых других растений на корнях образуются паренхимные наросты, которые еще называют клубеньками. Внутри этих клубеньков находится множество клубеньковых бактерий. Особенность этих бактерий в том, что они могут фиксировать атмосферный азот в виде соединений, которые усваиваются растениями. Например, бобовые клевер и люцерна могут накапливать в своих клубеньках 150-300 кг/га азота. Поэтому в сельском хозяйстве бобовые часто высаживают для того, чтобы обогатить почву азотом.
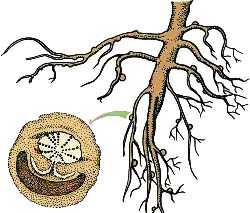 Паренхимные наросты или клубеньки
Паренхимные наросты или клубеньки
Запасающие корни.
В корнях любых растений, как правило, в некоторых количествах откладываются запасные питательные вещества такие, как сахар, крахмал, инулин и т.д. Но встречаются случаи, когда эта запасающая функция гипертрофирована и выходит на первый план. Корни при этом утолщаются и становятся мясистыми.
Такие видоизмененные стержневые корни, которые выполняют функцию запасания назвали «корнеплодами». Наиболее часто такая структура встречается у двулетников. К примеру, это морковь, свекла, репа, редис и т.д. В формировании этих корнеплодов принимает участие также и часть стебля – гипокотиль (или подсемядольное колено).
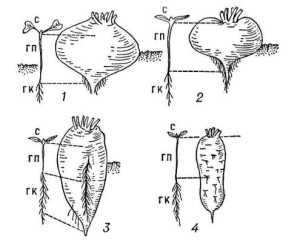
Корнеплоды на рисунке: 1 – брюква; 2 – свёкла египетская;
3 – свёкла сорта Маммут; 4 – морковь;
с – семядоли; гп – гипокотиль; гк – главный корень.
У некоторых видов растений встречаются так называемые корневые шишки, которые являются сильно утолщенными придаточными корнями. Это, например, георгина, любка, чистяк и т.д. Между корневыми шишками и «корнеплодами» встречаются многочисленные переходы.
Корневые шишки
Похожие статьи:
poznayka.org
Симбиоз с клубеньковыми бактериями - PDF
физиологии и биохимии растений
МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ ФЕДЕРАЦИИ Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования «КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ УНИВЕРСИТЕТ»
Подробнее ТЕМА «Энергетический обмен»
1. К автотрофным организмам относят 1) мукор 2) дрожжи 3) пеницилл 4) хлореллу ТЕМА «Энергетический обмен» 2. В процессе пиноцитоза происходит поглощение 1) жидкости 2) газов 3) твердых веществ 4) комочков
Подробнее БИОЛОГИЯ НАУКА О ЖИВой ПРИРОДе
БИОЛОГИЯ НАУКА О ЖИВой ПРИРОДе Биология изучает: ЧТО ИЗУЧАЕТ БИОЛОГИЯ z строение и жизнедеятельность живых организмов; z законы индивидуального и исторического развития организмов. 3 СИСТЕМА ОРГАНИЧЕСКОГО
Подробнее Энергетический обмен
Энергетический обмен Клетка открытая система. Гомеостаз Клетка открытая система, обмен веществ осуществляется только в том случае, если клетка получает все необходимые для нее вещества из окружающей среды
Подробнее СИСтЕМа ОРГаНИчЕСКОГО МИРа
СИСтЕМа ОРГаНИчЕСКОГО МИРа Все существующие на Земле организмы разделены на четыре царства: Дробянки, Грибы, Растения, Животные. Дробянки относятся к прокариотам (доядерным организмам), грибы, растения,
Подробнее Группа Ф.И.О. Билет 1
Группа Ф.И.О. Билет 1 1. Какие из перечисленных макромолекул обладают какими-либо общими характеристиками: ДНК, РНК, белки, углеводы, липиды? Укажите, какие именно общие свойства Вы выделяете для каждого
Подробнее Лекция 5. Дыхание растений
Лекция 5. Дыхание растений Общая характеристика и этапы дыхания Дыхание представляет собой окислительный распад органических веществ, синтезированных в процессе фотосинтеза, протекающий с потреблением
Подробнее БЛОК 2 Клетка как биологическая система.
1. К макроэлементам относятся: БЛОК 2 Клетка как биологическая система. 1) кислород, углерод, водород, азот 2) кислород, железо, золото 3) углерод, водород, бор 4) селен, азот, кислород 1) 2. Органоид,
Подробнее Вестник 3. Снисаренко Т.А., Медведева И. В.
УДК 582.288.45 Снисаренко Т.А., Медведева И. В. СТАРЕНИЕ КЛУБЕНЬКОВ TRIFOLIUM PRATENSE ПОД ВОЗДЕЙСТВИЕМ СТРЕССОГЕННЫХ ФАКТОРОВ Аннотация. В ходе эксперимента изучалось стрессогенное воздействие на морфо-функциональное
Подробнее ВВЕДЕНИЕ В ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ
ВВЕДЕНИЕ В ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ Жизнедеятельность организмов включает: а) обмен веществ и энергии; б) передача генетической информации; в) механизмы регуляции. Нарушение любого звена приводит к патологии.
Подробнее Пленарное заседание (пятница)
Предварительная программа Международной конференции "Микробиология в решении современных проблем сельскохозяйственного производства" (19-21 мая 2011 г.) 19.05. (четверг) 8.00-16.00. Заезд и регистрация
Подробнее ФГБОУ ВПО ДальГАУ. Классификация корней
Морфология и анатомия корня 1. Морфология корня 2. Метаморфозы корня 3. Клубеньки и микориза 4. Анатомия корня однодольных растений 5. Анатомия корня двудольных растений 1. Морфология корня Корень подземный
Подробнее Инструкция по выполнению работы
1 Инструкция по выполнению работы На выполнение работы по биологии отводится 45 минут. Работа включает в себя 21 задание. Ответы к заданиям 1 17 записываются в виде одной цифры, которая соответствует номеру
Подробнее Вариант 1 Вариант 2 Вариант 3 Вариант 4
Вариант 1 1 Биохимический состав растительной клетки. Белки, нуклеиновые кислоты, углеводы, липиды. 2 Водный баланс растений. Уравнение водного баланса. Водный дефицит. Завядание растений. 3 Интенсивность
Подробнее 3. Объем дисциплины и виды учебной работы:
1. Цели и задачи дисциплины: Цель дисциплины сформировать у студентов знания об использовании в промышленности физико-химических принципов работы живой клетки, ознакомить с методами и технологиями производства
Подробнее ОТЗЫВ НАУЧНОГО РУКОВОДИТЕЛЯ
ОТЗЫВ НАУЧНОГО РУКОВОДИТЕЛЯ о работе Плюта Владимира Александровича по кандидатской диссертации «Особенности образования биопленок и Quorum Sensing регуляция при действии антибактериальных агентов», представленной
Подробнее Задания С3 по биологии
Задания С по биологии. Какие механизмы обеспечивают работу иммунной системы человека? Элементы ответа ) Узнавание организм узнает чужеродные антигены и их продукты и выделяет антитела ) Специфичность антитела
Подробнее Тема Строение и функции пластид
Тема 2. 3. Строение и функции пластид Хромопласт Пропластида ТемноС та в Этиопласт е Амилопласт С т в е т Хлоропласт Взаимопревращения разных видов пластид Общий вид растительной клетки 5 мкм Хлоропласт
Подробнее Задания С5 по биологии
Задания С5 по биологии. Почему процесс оплодотворения у цветковых растений назван двойным? Что получается в результате этого оплодотворения? (допускаются иные формулировки ответа, не искажающие его смысла)
Подробнее СОДЕРЖАНИЕ ДИСЦИПЛИНЫ
СОДЕРЖАНИЕ ДИСЦИПЛИНЫ Содержание разделов дисциплины: 1. История развития агрохимии и задачи науки на современном этапе гумусная (Валериус, Тэер) и минеральная теории питания растений (А.Т.Болотов, Ж.Б.Буссенго,
Подробнее ID_2853 1/6 neznaika.pro
1 Клетка, её жизненный цикл (установление соответствия) Ответами к заданиям являются слово, словосочетание, число или последовательность слов, чисел. Запишите ответ без пробелов, запятых и других дополнительных
Подробнее Планируемые результаты
Планируемые результаты Ученик научится: - характеризовать особенности строения и процессов жизнедеятельности биологических объектов (клеток, организмов), их практическую значимость; - применять методы
Подробнее docplayer.ru
Живая природа - Клубеньковые бактерии бобовых 4
| | В клетках дегенерирующей ткани клубенька появляются многочисленные вакуоли, ядра теряют способность окрашиваться, часть клеток клубеньковых бактерий лизируется, часть мигрирует в окружающую среду в виде мелких кокковидных клеток-артроспор. Прекращают функционировать в этот период и инфекционные нити. Клетки хозяина утрачивают тургор и сжимаются теми соседними клетками, которым он еще свойствен. Старые клубеньки темные, дряблые, мягкие. При надрезе из них выступает водянистая слизь. Процессу разрушения клубенька, начинающегося с опробковения клеток сосудистой системы, способствуют понижение фотосинтетической активности растения, сухость или чрезмерная влажность среды. В разрушенном, ослизненном клубеньке обнаруживаются часто простейшие, грибы, бациллы и мелкие палочковидные клубеньковые бактерии. Состояние растения-хозяина оказывает влияние на длительность функционирования клубенька. Так, по данным Ф. Ф. Юхимчука (1957), кастрируя или удаляя цветы люпина, можно продлить период его вегетации и вместе с тем время активной деятельности клубеньковых бактерий. Клубеньки многолетних растений, в отличие от клубеньков однолетних, могут функционировать в течение многих лет. Так, например, карагана имеет многолетние клубеньки, в которых процесс старения клеток идет одновременно с образованием новых. У вистерии (глицинии китайской) также функционируют многолетние клубеньки, образуя на корнях хозяина шаровидные вздутия. К концу вегетационного периода бактероидная ткань многолетних клубеньков деградирует, но весь клубенек не отмирает. На следующий год он вновь начинает функционировать. Факторы, определяющие симбиотические взаимоотношения клубеньковых бактерий с бобовыми растениями. Для симбиоза, обеспечивающего хорошее развитие растений, необходим определенный комплекс условий среды. Если условия окружающей среды будут неблагоприятными, то, даже несмотря на высокую вирулентность, конкурентную способность и активность микросимбионта, эффективность симбиоза будет низкой. Для развития клубеньков оптимальная влажность 60—70% от полной влагоемкости почвы. Минимальная влажность почвы, при которой еще возможно развитие клубеньковых бактерий в почве, приблизительно равна 16% от полной влагоемкости. При влажности ниже этого предела клубеньковые бактерии обычно уже не размножаются, но тем не менее они не погибают и могут длительное время сохраняться в неактивном состоянии. Недостаток влаги приводит и к отмиранию уже сформировавшихся клубеньков. Нередко в районах с недостаточным увлажнением многие бобовые растения развиваются, не образуя клубеньков. Поскольку размножение клубеньковых бактерий в отсутствие влаги не происходит, в случае засушливой весны инокулированные (искусственно зараженные) семена необходимо вносить глубже в почву. Например, в Австралии семена с нанесенными на них клубеньковыми бактериями глубоко заделывают в почву. Интересно, что клубеньковые бактерии почв засушливого климата более стойко переносят засуху, чем бактерии почв влажного климата. В этом проявляется их экологическая приспособленность. Избыточная влажность, как и ее недостаток, также неблагоприятна для симбиоза — из-за снижения степени аэрации в зоне корней ухудшается снабжение корневой системы растения кислородом. Недостаточная аэрация отрицательно влияет и на живущие в почве клубеньковые бактерии, которые, как известно, лучше размножаются при доступе кислорода. Тем не менее высокая аэрация в зоне корней приводит к тому, что кислород начинают связывать восстановители молекулярного азота, снижая степень азотфиксации клубеньков. Важную роль во взаимоотношениях клубеньковых бактерий и бобовых растений играет температурный фактор. Температурные характеристики разных видов бобовых растений различны. Также и разные штаммы клубеньковых бактерий имеют свои определенные температурные оптимумы развития и активной фиксации азота. Следует отметить, что оптимальные температуры развития бобовых растений, образования клубеньков и азотфиксации не совпадают. Так, в природных условиях образование клубеньков может наблюдаться при температурах несколько выше 0 °С, азотфиксация при таких условиях практически не происходит. Возможно, лишь арктические симбиозирующие бобовые растения связывают азот при очень низких температурах. Обычно же этот процесс происходит лишь при 10 °С и выше. Максимальная азотфиксации ряда бобовых растений наблюдается при 20—25 °С. Температура выше 30 °С отрицательно влияет на процесс азотонакопления. Экологическая адаптация к температурному фактору у клубеньковых бактерий значительно меньше, чем у многих типичных сапрофитных форм. По мнению Е. Н. Мишустина (1970), это объясняется тем, что естественной средой обитания клубеньковых бактерий являются ткани растений, где температурные условия регулируются растением-хозяином. Большое влияние на жизнедеятельность клубеньковых бактерий и образование клубеньков оказывает реакция почвы. Для разных видов и даже штаммов клубеньковых бактерий значение рН среды обитания несколько различно. Так, например, клубеньковые бактерии клевера более устойчивы к низким значениям рН, чем клубеньковые бактерии люцерны. Очевидно, здесь также сказывается адаптация микроорганизмов к среде обитания. Клевер растет на более кислых почвах, чем люцерна. Реакция почвы как экологический фактор оказывает влияние на активность и вирулентность клубеньковых бактерий. Наиболее активные штаммы, как правило, легче выделить из почв с нейтральными значениями рН. В кислых почвах чаще встречаются неактивные и слабовирулентные штаммы. Кислая среда (рН 4,0— 4,5) оказывает непосредственное влияние и на растения, в частности нарушая синтетические процессы обмена веществ растений и нормальное развитие корневых волосков. В кислой среде у инокулированных растений резко сокращается срок функционирования бактероидной ткани, что ведет к снижению степени азотфиксации. В кислых почвах, как отмечает А. В. Петербургский, в почвенный раствор переходят соли алюминия и марганца, неблагоприятно действующие на развитие корневой системы растений и процесс азотоусвоения, а также снижается содержание усвояемых форм фосфора, кальция, молибдена и углекислоты. Неблагоприятную реакцию почвы лучше всего устраняет известкование. Размеры симбиотической азотфиксации определяются в значительной степени условиями питания растения-хозяина, а не клубеньковых бактерий. Клубеньковые бактерии как эндотрофные симбионты растений зависят в основном от растения при получении углеродсодержащих веществ и минеральных элементов питания. Для клубеньковых бактерий ткань хозяина представляет такую питательную среду, которая может удовлетворить даже самый требовательный штамм вследствие содержания в ткани всех типов питательных веществ. Тем не менее после внедрения клубеньковых бактерий в ткань растения-хозяина их развитие определяется не только внутренними процессами, но и в значительной степени зависит от действия внешних факторов, оказывающих влияние на весь ход инфекционного процесса. Содержание или отсутствие того или иного питательного вещества в окружающей среде может быть определяющим моментом для проявления симбиотической азотфиксации. Степень обеспеченности бобовых растений доступными формами минеральных соединений азота определяет эффективность симбиоза. На основании многочисленных лабораторных и вегетационных опытов известно, что чем больше азотсодержащих соединений в окружающей среде, тем с большим трудом внедряются бактерии в корень. Сельскохозяйственная практика требует однозначно решить задачу — целесообразнее удобрять бобовые культуры азотом или же правы те исследователи, которые утверждают, что минеральный азот подавляет симбиотическую азотфиксацию бобовых культур и поэтому экономически выгоднее такие растения азотом не удобрять. На кафедре агрономической и биологической химии Московской сельскохозяйственной академии им. К. А. Тимирязева были проведены опыты, результаты которых дали возможность получить картину поведения симбионтов в условиях вегетационных и полевых опытов при обеспеченности растений разными дозами азота в среде. Установлено, что повышение содержания растворимых азотсодержащих соединений в среде в полевых условиях при оптимальных условиях произрастания растений не препятствует их симбиозу с клубеньковыми бактериями. Снижение доли атмосферного азота, усваиваемого растениями при повышенной обеспеченности минеральным азотом, имеет только относительный характер. Абсолютное количество азота, усвоенного бактериями из атмосферы, практически не снижается, даже нередко увеличивается по сравнению с растениями, выращивающимися в присутствии клубеньковых бактерий, но без внесения в почву азота. Большое значение в активации усвоения азота бобовыми растениями имеет фосфорное питание. При низком содержании фосфора в среде проникновение бактерий в корень происходит, но клубеньки при этом не образуются. Бобовым растениям присущи некоторые особенности в обмене фосфорсодержащих соединений. Семена бобовых отличаются повышенным содержанием фосфора. Запасной фосфор при прорастании семян используется не так, как У Других культур,— сравнительно равномерно для формирования всех органов, а в большей степени сосредоточиваясь в корнях. Поэтому в ранние сроки развития бобовые растения, в отличие от злаковых, в большей степени удовлетворяют свои потребности в фосфоре за счет семядолей, а не запасов почвы. Чем крупнее семена, тем меньше бобовые растения зависят от фосфора почвы. Однако при симбиотическом способе существования потребность бобовых растений в фосфоре выше, чем при автотрофном. Поэтому при недостатке фосфора в среде у инокулированных растений ухудшается снабжение растений азотом. Бобовые растения, как известно, выносят с урожаем значительно больше калия, чем другие сельскохозяйственные культуры. Поэтому калийные и особенно фосфорно-калийные удобрения существенно повышают продуктивность азотфиксации бобовыми растениями. Положительное действие калия на образование клубеньков и интенсивность азотфиксации связано в значительной степени с физиологической ролью калия в углеводном обмене растения. Кальций нужен не только для устранения излишней кислотности почвы. Он играет специфическую роль в развитии клубеньковых бактерий и обеспечении нормального симбиоза бактерий с растением-хозяином. Потребность клубеньковых бактерий в кальции частично может быть компенсирована стронцием. Интересно, что клубеньковые бактерии тропических культур, растущих на кислых латеритных почвах, не нуждаются в кальции. В этом опять проявляется экологическая адаптация клубеньковых бактерий, поскольку тропические почвы содержат очень небольшие количества кальция. Для симбиотической азотфиксации необходимы также магний, сера и железо. При недостатке магния тормозится размножение клубеньковых бактерий, снижается их жизнедеятельность, подавляется симбиотическая азотфиксация. Сера и железо оказывают также благоприятное влияние на образование клубеньков и процесс азотфиксации, в частности играя несомненную роль в синтезе леггемоглобина. Из микроэлементов особо отметим роль молибдена и бора. При недостатке молибдена клубеньки плохо образуются, в них нарушается синтез свободных аминокислот и подавляется синтез леггемоглобина. Молибден вместе с другими элементами с переменной валентностью (Fe, Сo, Сu) служит посредником при переносе электронов в окислительно-восстановительных ферментных реакциях. При дефиците бора в клубеньках не формируются сосудистые пучки, и вследствие этого нарушается развитие бактероидной ткани. На формирование клубеньков у бобовых растений большое влияние оказывает углеводный обмен растений, определяемый рядом факторов: фотосинтезом, наличием в среде углекислого газа, физиологическими особенностями растений. Улучшение углеводного питания благоприятно сказывается на инокуляционном процессе и азотонакоплении. С . практической точки зрения большой интерес представляет использование соломы и соломистого свежего навоза для удобрения бобовых растений как источника углеводов. Но в первый год после внесения соломы в почву при ее разложении накапливаются токсические вещества. Следует отметить, что не все виды бобовых растений чувствительны к токсическим продуктам распада соломы; горох, например, не реагирует на них. Определенное значение в симбиозе клубеньковых бактерий и бобовых растений имеют биологические факторы. Большое внимание уделяется влиянию ризосферной микрофлоры на клубеньковые бактерии, которое может иметь как стимуляционный, так и антагонистический характер в зависимости от состава микроорганизмов ризосферы. Много работ посвящено изучению фагов клубеньковых бактерий. Большинство фагов спо собны лизировать различные виды бактерий, некоторые специализированы лишь в отношении отдельных видов или даже штаммов клубеньковых бактерий. Фаги могут препятствовать внедрению бактерий в корень, вызывать лизис клеток в ткани клубенька. Фаги наносят большой ущерб, лизируя препараты клубеньковых бактерий на заводах, вырабатывающих нитрагин. Среди различных видов насекомых, наносящих вред клубеньковым бактериям, особенно выделяется полосатый клубеньковый долгоносик, личинки которого разрушают клубеньки па корнях многих видов бобовых растений (главным образом однолетних). Широко распространен и щетинистый клубеньковый долгоносик. Ранней весной самки клубеньковых долгоносиков откладывают от 10 до 100 яиц. Через 10—15 дней из яиц развиваются небольшие (до 5,5 мм), червеобразные, согнутые, белые, со светло-бурой головкой личинки, питающиеся преимущественно клубеньками и корневыми волосками. Только что вылупившиеся личинки проникают в клубенек и питаются его содержимым. Более взрослые личинки разрушают клубеньки снаружи. Одна личинка за 30—40 дней уничтожает 2—6 клубеньков. Особенно большой вред они наносят в сухую и жаркую погоду, когда развитие растений замедляется. Клубеньки люцерны и некоторых других видов бобовых растений повреждает также большой люцерновый долгоносик. Самки жука откладывают до 400 яиц, из которых развиваются безногие, дугообразные, желтовато-белые, с бурой головкой, покрытые бурыми щетинками личинки. Их длина 10— 14 мм. Цикл развития большого люцернового долгоносика протекает в течение двух лет. Наличие нематод в корневой зоне различных видов бобовых растений отмечают многие исследователи. В прикорневой зоне гороха, например, обнаружено 47 видов нематод, среди них 25 паразитических. На корнях молодых растений фасоли, люпина, клевера может паразитировать широко распространенная ростковая нематода. Самки этого вида, питающиеся корнями растений, откладывают яйца в ткани растения. Весь жизненный цикл развивающейся из яиц нематоды протекает обычно внутри тканей. В степных районах на корнях люцерны, клевера и сои обнаружена степная нематода. Самки перед откладкой яиц проникают в корень, куда откладывают от 12 до 20 яиц. В корнях личинки проходят три личиночные стадии развития, нарушая функции корня и клубеньков. Распространение клубеньковых бактерий в природе. Являясь симбиотическими организмами, клубеньковые бактерии распространяются в почвах, сопутствуя определенным видам бобовых растений. После разрушения клубеньков клетки клубеньковых бактерий попадают в почву и переходят к существованию за счет различных органических веществ подобно другим почвенным микроорганизмам. Почти повсеместное распространение клубеньковых бактерий является доказательством высокой степени их адаптируемости к различным почвенноклиматическим условиям, способности вести симбиотический и сапрофитный способ жизни. Схематизируя имеющиеся к настоящему времени данные по распространению клубеньковых бактерий в природе, можно сделать следующие обобщения. В целинных и окультуренных почвах присутствуют обычно в больших количествах клубеньковые бактерии тех видов бобовых растений, которые имеются в составе дикой флоры или культивируются длительное время в данной местности. Численность клубеньковых бактерий всегда наивысшая в ризосфере -бобовых растений, несколько меньше их в ризосфере других видов и мало в почве вдали от корней. В почвах встречаются как эффективные, так и неэффективные клубеньковые бактерии. Имеется много данных о том, что длительное сапрофитное существование клубеньковых бактерий, особенно в почвах с неблагоприятными свойствами (кислых, засоленных), ведет к снижению и даже утрате активности бактерий. Перекрестная заражаемость разных видов бобовых растений нередко приводит в природе и сельскохозяйственной практике к появлению на корнях клубеньков, недостаточно активно фиксирующих молекулярный азот. Это, как правило, зависит от отсутствия в почве соответствующих видов клубеньковых бактерий. Особенно часто такое явление наблюдается при использовании новых видов бобовых растений, которые либо заражаются неэффективными видами бактерий перекрестных групп, либо развиваются без клубеньков. |
| |
naturalia.ucoz.ru