
Мир науки. Развитие споровых растений
Общая характеристика высших споровых растений
Предпосылки выхода растений на сушу. Изучая древние ископаемые остатки растений и животных, ученые пришли к выводу, что климат Земли, ее атмосфера, очертания материков и океанов многократно менялись. Появление
атмосферного кислорода и озонового слоя позволило живым организмам выйти на сушу. Выход растений на сушу привел к появлению группы высших растений и стал новым этапом в развитии растительного мира нашей планеты.
Считают, что первые наземные растения появились около 450 млн лет назад на мели водоемов. По окаменевшими остатками ученые восстановили их вид. Они имели тело, которое еще не было четко расчлененное на корневую и стебельную части. Роль листьев выполняли чешуи, ветви вилчасто галузилися. Такое ветвление характеризуется постоянным раздвоением оси стебля при образовании новых ветвей, и, как считают ученые, является примитивным признаком. На верхушках некоторых ветвей размещались спорангии, в которых созревали споры. Как и современные, первые наземные растения имели покровную, ведущую и основную ткани, но еще не имели механической, так высотой были не более полуметра (вспомните строение и функции тканей растений).
Подобные по строению вымершие организмы принадлежат к отдела риниофиты и считаются переходным звеном к высших растений.
Особенности строения споровых растений. К современным споровых растений относятся представители отделов Мохообразные, Плауновидные, Хвощевидные и Папоротникообразные. Это наземные организмы, и только некоторые из них приспособились к жизни в водной среде. При огромном разнообразии внешнего вида эти растения имеют общие черты строения. Тело споровых (кроме мхов) делится на корень и побег. В связи с переходом к наземному способу существования у споровых растений возникают покровные, механические и проводящие ткани. Покровные ткани защищают растения от высыхания, они устьица для газообмена и испарения. Во покровной тканью расположена механическая, которая обеспечивает растению сопротивления. Движение веществ от корня к листьям и наоборот осуществляется с помощью проводящих тканей. Пространство между покровной, механической и ведущей тканями в споровых заполнен основной тканью, выполняющей фотосинтезирующую и запасающую функции. Размножаются эти растения спорами, и определило их название.
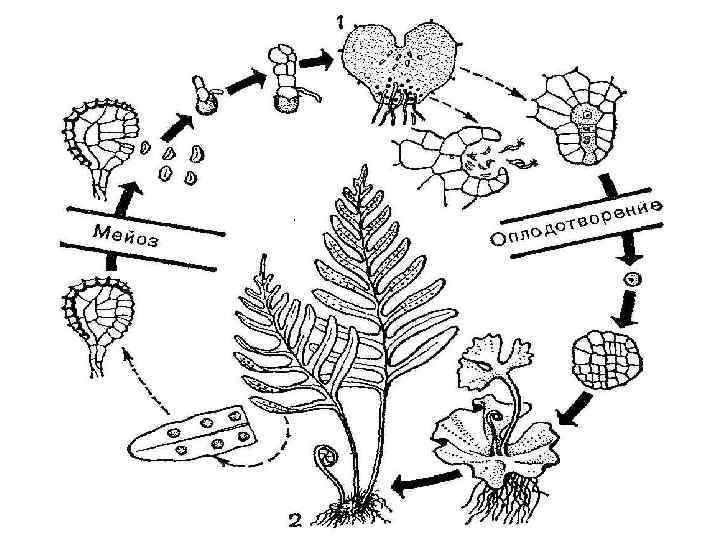
Размножение и развитие споровых растений. В споровых растений развитие происходит путем ряда преобразований со сменой поколений - полового и бесполого.
Изменение полового поколения бесполым в цикле развития растений называется чередованием поколений. Оно возникло еще в некоторых водорослей (бурых, зеленых), а для высших растений является обязательным.
Бесполое поколение, или спорофит, для размножения образует неполовые клетки - споры. Из споры вырастает заросток, на котором формируются женские и мужские половые органы. В них созревают гаметы. Поколение, образует гаметы, будет половым поколением в жизненном цикле развития споровых растений, или гаметофитом. Гаметы участвуют в оплодотворении. Оплодотворение происходит только в водно-капельном среде, поэтому споровые растения распространены на влажных участках суши, хотя некоторые существуют даже в пустынях (подумайте, какие условия для этого должны быть). Из зиготы, которая образуется после оплодотворения вырастает спорофит. Цикл развития начинается снова. У растений разных систематических групп, с которыми вы познакомитесь дальше, гаметофит и спорофит имеют различный вид. Выход растений на сушу обусловил изменение их строения и процессов размножения. Наземные растения, в отличие от водорослей, имеют тело, разделенное на подземные и надземные органы (корень и побег), а также разнообразные ткани (покровную, ведущую, механическую и др.), позволяет им приспособиться к жизни на суше.
worldofscience.ru
ДИПЛОИДНАЯ ЛИНИЯ ЭВОЛЮЦИИ ВЫСШИХ СПОРОВЫХ РАСТЕНИЙ
 ДИПЛОИДНАЯ ЛИНИЯ ЭВОЛЮЦИИ ВЫСШИХ СПОРОВЫХ РАСТЕНИЙ
ДИПЛОИДНАЯ ЛИНИЯ ЭВОЛЮЦИИ ВЫСШИХ СПОРОВЫХ РАСТЕНИЙ
 • Все высшие растения кроме мхов относят к диплоидной эволюционной ветви развития. В их жизненном цикле господствующим поколением является спорофит бесполое поколение.
• Все высшие растения кроме мхов относят к диплоидной эволюционной ветви развития. В их жизненном цикле господствующим поколением является спорофит бесполое поколение.
 Эволюция жизненного цикла • У всех высших растений спорофит зеленое автотрофное растение, в то время как гаметофит в той или иной степени редуцирован и даже переходит у разноспоровых и семенных растений в полную зависимость от спорофита, развиваясь внутри оболочки мегаспоры и за счет питательных веществ спорофита.
Эволюция жизненного цикла • У всех высших растений спорофит зеленое автотрофное растение, в то время как гаметофит в той или иной степени редуцирован и даже переходит у разноспоровых и семенных растений в полную зависимость от спорофита, развиваясь внутри оболочки мегаспоры и за счет питательных веществ спорофита.
 Споровые в наземных условия • Приспособления наземных растений выразились в возрастающем расхождении спорофита и гаметофита, выполняющих различные функции: • появление защищенных оболочкой многоклеточных половых органов, глубокая морфологическая дифференциация спорофита и возникновение многоклеточных спорангиев, развитие проводящей и механической систем тканей, появление эпидермы, кутикулы и устьиц.
Споровые в наземных условия • Приспособления наземных растений выразились в возрастающем расхождении спорофита и гаметофита, выполняющих различные функции: • появление защищенных оболочкой многоклеточных половых органов, глубокая морфологическая дифференциация спорофита и возникновение многоклеточных спорангиев, развитие проводящей и механической систем тканей, появление эпидермы, кутикулы и устьиц.
 Массовое производство спор в наземных условиях • В наземных условиях большая часть спор попадает в неблагоприятные условия и погибает, поэтому возникает необходимость их массового производства. • Производство большого количества спор требует накопления достаточного количества органического вещества, что неизменно приводит к увеличению размеров самого спорофита и расширению его фотосинтезирующей поверхности.
Массовое производство спор в наземных условиях • В наземных условиях большая часть спор попадает в неблагоприятные условия и погибает, поэтому возникает необходимость их массового производства. • Производство большого количества спор требует накопления достаточного количества органического вещества, что неизменно приводит к увеличению размеров самого спорофита и расширению его фотосинтезирующей поверхности.
 Расчленение спорофита • Увеличение размеров спорофита неизбежно сопровождается его внешним и внутренним расчленением, так как для нормального функционирования организма необходимо определенное соотношение между его поверхностью и объемом
Расчленение спорофита • Увеличение размеров спорофита неизбежно сопровождается его внешним и внутренним расчленением, так как для нормального функционирования организма необходимо определенное соотношение между его поверхностью и объемом
 Появление проводящей системы • Увеличение размеров и возрастающее расчленение спорофита сопровождается появлением сложной проводящей системы
Появление проводящей системы • Увеличение размеров и возрастающее расчленение спорофита сопровождается появлением сложной проводящей системы
 Водное питание в наземных условиях • Решение проблем водного режима привело к образованию ксилемы, или древесины, обеспечивающей водопроведение. Первоначально ксилема состояла лишь из вытянутых клеток с множеством пор трахеид • У семенных растений в процессе эволюции она пополнилась более крупными сосудами трахеями разного типа. Органические вещества передвигались по особым ситовидным трубкам элементам флоэмы.
Водное питание в наземных условиях • Решение проблем водного режима привело к образованию ксилемы, или древесины, обеспечивающей водопроведение. Первоначально ксилема состояла лишь из вытянутых клеток с множеством пор трахеид • У семенных растений в процессе эволюции она пополнилась более крупными сосудами трахеями разного типа. Органические вещества передвигались по особым ситовидным трубкам элементам флоэмы.
 Стелярное строение • Флоэма и ксилема в центральном цилиндре различных групп высших растений располагается поразному. В связи с эти выделяют так называемые типы стелярного строения
Стелярное строение • Флоэма и ксилема в центральном цилиндре различных групп высших растений располагается поразному. В связи с эти выделяют так называемые типы стелярного строения
 Стелы разных групп споровых • Наиболее примитивным типом стелы является протостела риниофитов и некоторых других споровых. Позже появились плектостела (плауны), сифоностела (папоротники), артростела (хвощи) и диктиостела (папоротники). Для большинства семенных растений характерны эвстела, а для однодольных особая атактостела.
Стелы разных групп споровых • Наиболее примитивным типом стелы является протостела риниофитов и некоторых других споровых. Позже появились плектостела (плауны), сифоностела (папоротники), артростела (хвощи) и диктиостела (папоротники). Для большинства семенных растений характерны эвстела, а для однодольных особая атактостела.
 Эволюция стелы
Эволюция стелы
 Возникновение листьев • В результате • У всех дальнейшего плауновидных они морфологического возникли в качестве расчленения выростов (энациев) спорофита возникли на осевых органах. специальные органы • Это так называемые фотосинтеза энационные листья
Возникновение листьев • В результате • У всех дальнейшего плауновидных они морфологического возникли в качестве расчленения выростов (энациев) спорофита возникли на осевых органах. специальные органы • Это так называемые фотосинтеза энационные листья
 Теломные листья • Листья большинства споровых и семенных растений возникли в результате уплощения и срастания конечных веточек теломов или их систем. Это теломные листья, или плосковетки.
Теломные листья • Листья большинства споровых и семенных растений возникли в результате уплощения и срастания конечных веточек теломов или их систем. Это теломные листья, или плосковетки.
 Эволюция теломных листьев • Теломные листья с самого начала были спороносными, выполняя как функции фотосинтеза так и размещения органов бесполого размножения. • В процессе дальнейшей эволюции происходило постепенное пространственное разделение этих функций.
Эволюция теломных листьев • Теломные листья с самого начала были спороносными, выполняя как функции фотосинтеза так и размещения органов бесполого размножения. • В процессе дальнейшей эволюции происходило постепенное пространственное разделение этих функций.
 Происхождение листьев
Происхождение листьев

 Теломная теория
Теломная теория
, после") Эволюция ветвления • Исходной формой ветвления спорофитов было равнодихотомическое ( риниофиты, плауны, псилот), после чего растения выработали неравную дихотомию (анизотомию), • Дальнейшим шагом в эволюции ветвления стало дихоподиальное ветвление, которое постепенно перешло в моноподиальное (хвойные) и, наконец, в процессе эволюции возникло симподиальное ветвление, широко распространенное у цветковых растений.
Эволюция ветвления • Исходной формой ветвления спорофитов было равнодихотомическое ( риниофиты, плауны, псилот), после чего растения выработали неравную дихотомию (анизотомию), • Дальнейшим шагом в эволюции ветвления стало дихоподиальное ветвление, которое постепенно перешло в моноподиальное (хвойные) и, наконец, в процессе эволюции возникло симподиальное ветвление, широко распространенное у цветковых растений.
 Эволюция ветвления
Эволюция ветвления
 Функции листьев у споровых • В отличие от безлистных риниофитов на листьях высших споровых диплоидной линии эволюции развиваются спорангии. • Теломные листья в отличие от энационных с самого начала были спороносными (фертильными). Такие спороносные листья, или спорофиллы, исполняли сначала функцию фотосинтеза и бесполого размножения
Функции листьев у споровых • В отличие от безлистных риниофитов на листьях высших споровых диплоидной линии эволюции развиваются спорангии. • Теломные листья в отличие от энационных с самого начала были спороносными (фертильными). Такие спороносные листья, или спорофиллы, исполняли сначала функцию фотосинтеза и бесполого размножения
 Разделение функций листьев В процессе эволюции происходило разделение этих функций. Так у ряда папоротников (ужовниковые, осмунда, анемия) разделение происходит в пределах листа на фертильную и стерильную части. У других (страусопер) разделение происходит между стерильными (трофофиллы) и фертильными (спорофиллы) листьями, обычно отличающимися по форме.
Разделение функций листьев В процессе эволюции происходило разделение этих функций. Так у ряда папоротников (ужовниковые, осмунда, анемия) разделение происходит в пределах листа на фертильную и стерильную части. У других (страусопер) разделение происходит между стерильными (трофофиллы) и фертильными (спорофиллы) листьями, обычно отличающимися по форме.
 Два типа листьев страусопера Трофофиллы Спорофиллы
Два типа листьев страусопера Трофофиллы Спорофиллы
 Стробилы споровых побеги со спорофиллами называются стробилами
Стробилы споровых побеги со спорофиллами называются стробилами
 Среди представителей диплоидной линии эволюции имеются равноспоровые и разноспоровые растения • У равноспоровых морфологически одинаковые споры прорастают в обоеполые, самостоятельно существующие заростки. • У разноспоровых на спорофите образуются спорангии двух типов: мегаспорангии с мегаспороами и микроспорангии с микроспорами.
Среди представителей диплоидной линии эволюции имеются равноспоровые и разноспоровые растения • У равноспоровых морфологически одинаковые споры прорастают в обоеполые, самостоятельно существующие заростки. • У разноспоровых на спорофите образуются спорангии двух типов: мегаспорангии с мегаспороами и микроспорангии с микроспорами.
 Разноспоровые растения • Споры разноспоровых растений прорастают в однополые, сильно редуцированные микроскопические гаметофиты, которые способны существовать лишь за счет питательных веществ мегаспоры, под защитой ее оболочки и даже самого мегаспорангия. • Особенно сильно редуцирован мужской гаметофит, нередко представляющий всего одну две ризоидальные клетки
Разноспоровые растения • Споры разноспоровых растений прорастают в однополые, сильно редуцированные микроскопические гаметофиты, которые способны существовать лишь за счет питательных веществ мегаспоры, под защитой ее оболочки и даже самого мегаспорангия. • Особенно сильно редуцирован мужской гаметофит, нередко представляющий всего одну две ризоидальные клетки
 Генеральная линия эволюции • Таким образом, разноспоровость всегда сопровождается сильнейшей редукцией гаметофита. • Редукция гаметофитов, т. е. полового поколения генеральная линия эволюции всего растительного мира!
Генеральная линия эволюции • Таким образом, разноспоровость всегда сопровождается сильнейшей редукцией гаметофита. • Редукция гаметофитов, т. е. полового поколения генеральная линия эволюции всего растительного мира!
 Отделы высших споровых диплоидной линии эволюции
Отделы высших споровых диплоидной линии эволюции
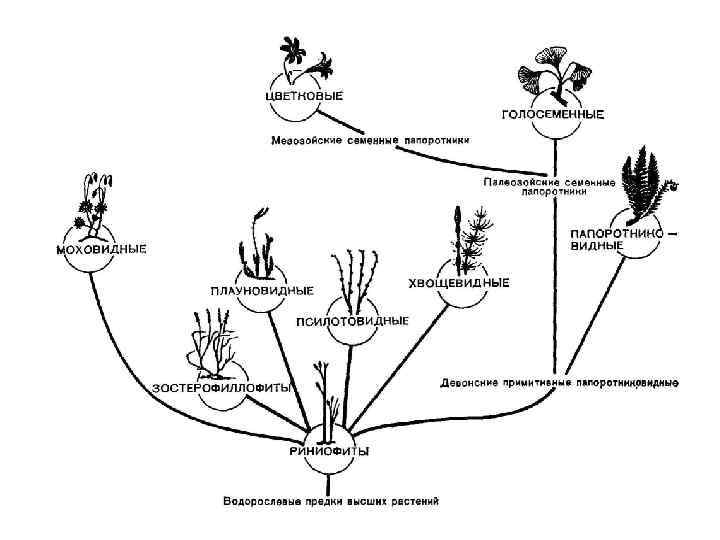
 или псилофитовидные • Впервые они были найдены в 1859 г. канадским геологом") Риниофиты (RHYNIOPHYTA) или псилофитовидные • Впервые они были найдены в 1859 г. канадским геологом Дж. Досоном в девонских отложениях. Это были не похожие ни на одно из известных науке в то время растения с дихотомически разветвленными стеблями без листьев и верхушечными спорангиями.
Риниофиты (RHYNIOPHYTA) или псилофитовидные • Впервые они были найдены в 1859 г. канадским геологом Дж. Досоном в девонских отложениях. Это были не похожие ни на одно из известных науке в то время растения с дихотомически разветвленными стеблями без листьев и верхушечными спорангиями.
") Псилофит – голорос первичный (Psilophyton princeps)
Псилофит – голорос первичный (Psilophyton princeps)
 Находка риниофитов • Растение было названо Psilophyton princeps голорос первичный. Необычное растение не укладывалось в существующие тогда системы классификации. • Повторно риниофиты были обнаружены в 1912 г. в местечке Райни в Шотландии в пластах раннедевонского (415 млн. лет назад) кремнистого черта.
Находка риниофитов • Растение было названо Psilophyton princeps голорос первичный. Необычное растение не укладывалось в существующие тогда системы классификации. • Повторно риниофиты были обнаружены в 1912 г. в местечке Райни в Шотландии в пластах раннедевонского (415 млн. лет назад) кремнистого черта.
 и хорнеофит (Horneophyton).") Представители риниофитов • Новые находки получили названия риния (Rhynia) и хорнеофит (Horneophyton).
Представители риниофитов • Новые находки получили названия риния (Rhynia) и хорнеофит (Horneophyton).
 Представители риниофитов • Позже были найдены другие представители риниофитов: Cooksonia, Yarravia, Trimerophyton и др. ).
Представители риниофитов • Позже были найдены другие представители риниофитов: Cooksonia, Yarravia, Trimerophyton и др. ).
 Появление синангиев • Ярравия
Появление синангиев • Ярравия
 Строение риниофитов • Спорофит риниофитов имел протостелическое строение, а механические ткани и камбий отсутствовали. Имелись устьица примитивного типа. Спорангии были крупные, терминальные и в них формировались многочисленные споры, по характеру оболочки типичные для высших растений. Споры развивались в тетрадах и имели трехлучевой рубец.
Строение риниофитов • Спорофит риниофитов имел протостелическое строение, а механические ткани и камбий отсутствовали. Имелись устьица примитивного типа. Спорангии были крупные, терминальные и в них формировались многочисленные споры, по характеру оболочки типичные для высших растений. Споры развивались в тетрадах и имели трехлучевой рубец.
present5.com
| Споры у споровых растений образуются в специальных
органах – многоклеточных спорангиях на диплоидном спорофите. Споры лишены
ундулиподиев и неподвижны, рассеваясь с помощью ветра. У равноспоровых
высших растений все споры одинаковые. У эволюционно более продвинутых таксонов
образуются споры разной величины: мелкие – микроспоры и крупные – мегаспоры.
При прорастании спор формируется гаплоидный гаметофит, называемый заростком,
с половыми органами – архегонии и антеридии. Обоеполый гаметофит
формируется равноспоровых растений. У разноспоровых растений микроспоры
при прорастании образуют мужской гаметофит с антеридиями, а мегоспоры -
женский с архегониями. В антеридиях развиваются подвижные с ундулиподиями
мужские гаметы – сперматозоиды или неподвижные спермии. В архегониях созревают
женские гаметы – яйцеклетки. Процесс оплодотворения у этой группы растений,
так же как и у низших, может осуществляться только в присутствии капельно-жидкой
воды с образованием зиготы, дающей начало многоклеточному зародышу, из
которого развивается диплоидный спорофит. Редукция числа хромосом происходит
при образовании спор, почему гаметофиты всегда гаплоидны.
Моховидные - наиболее обособленные из всех групп наземных растений, занимающие преимущественно влажные местообитания, с регрессивным развитием спорофита. Моховидные делят на три класса – печеночные, антоцеротовые и листостебельные мхи. Это единственная в истории растительного мира линия эволюции, характеризующаяся преобладанием в жизненном цикле полового поколения - гаплоидного гаметофита, в то время как у всех остальных наземных растений доминирует диплоидный спорофит. Как и для всех высших растений, для них характерно правильное чередование полового и бесполого поколений. При этом половое и бесполое поколения - одно растение. Спорофит моховидных, или диплоидное бесполое поколение, целиком и полностью в своей жизнедеятельности зависит от гаметофита и представляет собой небольшую коробочку со спорами – спорогон, в нижней части прикрепленную гаусториями к телу (талому) гаметофита. Половое размножение. Развитие полового поколения моховидных начинается с момента прорастания споры и формирования нитчатого (у большинства представителей) или пластинчатого (у сфагновых мхов) образования – протонемы с почками. Из почек вырастает пластинчатое слоевище или облиственные стебли гаметофоры с половыми органами (гаметангиями). Органы полового размножения – многоклеточные архегонии и антеридии, чаще всего собранны группами на верхушках побегов и ветвей или же расположены на верхней поверхности талломов. Моховидные могут быть однодомными или двудомными. У листостебельных мхов архегонии и антеридии окружены специализированными окрашенными листьями и наблюдается половой диморфизм – гаметофиты двудомных видов, несущие антеридии, часто сильно редуцированы до так называемых карликовых мужских растений, которые после созревания антеридиев отмирают. У некоторых видов карликовые мужские экземпляры поселяются и образуют антеридии прямо на листьях женских растений. Для протекания полового процесса необходимо наличие капельно-жидкой среды.. Антеридии продуцируют сперматозоиды с ундулиподиями (жгутиками), а яйцеклетка расположена в архегонии, расположенные на верхней поверхности таллома. Антеридии имеют вид продолговатых или округлых мешочков на ножке, а архегонии обычно имеют бутылкообразный вид - суженную шейку и брюшко, где находится крупная яйцеклетка. Половое размножение гаметофитов происходит весной или в начале лета. Талломы печеночных мхов – раздельнополые, органы размножения размещаются на особых подставках и возвышаются над слоевищем. Гаметангии моховидных располагаются обычно группами, тем самым облегчая протекание процесса оплодотворения. Сперматозоидов образуется очень много, так что жидкость, покрывающая таллом обычно имеет молочный оттенок. В слизи архегония содержатся атрактанты, химические вещества, указывающие направление движения мужских гамет. Движение сперматозоида возможно только в воде – во время дождя или тумана мужская половая клетка достигает архегония и по образовавшемуся к этому времени узкому каналу проникает внутрь, где и сливается с яйцеклеткой. Образовавшаяся диплоидная зигота через определенный промежуток времени (от нескольких месяцев до двух лет) дает начало спорофиту (спорогону), заканчивающемуся коробочкой, в которой созревают споры. Сначала формируется подставка, которая врастает в гаметофит и действует как всасывающий орган, через который спорофит получает питательные вещества. У бриевых мхов, наиболее обширного подкласса листостебельных мхов, спорогоны бывают как верхушечные, так и боковыми. Бесполое размножение. Образованию спор в спорангиях предшествует мейоз, число хромосом уменьшается вдвое, т.е. споры имеют гаплоидный набор хромосом. В коробочке спорофита находятся материнские клетки спор, которые мейотически делятся с образованием гаплоидных спор. Оставшиеся клетки образуют пружинки, или элатеры. После зимнего покоя весной развитие спорофита возобновляется и сформировавшаяся коробочка с колпачком выносится на ножке над талломом. При этом споры становятся многоклеточными, в них появляется хлоропласт и спора превращается в гаметофит. Созревшая коробочка при подсыхании раскрывается четырьмя створками. Элатеры способствуют рассеиванию спор, разрыхляя их массу. Споры разносятся ветром и, попав в благоприятные условия, прорастают, формируя таллом гаметофитного поколения. Вегетативное размножение. Оно характерно для многих видов моховидных и осуществляется фрагментами слоевища или выводковыми почками, образующимися по краю или на нижней стороне талломов. Выводковые почки развиваются в особых органах – выводковых корзиночках, называемые выводковыми тельцами. Плауновидные (Lycopodiophyta). Сюда входят более высокоорганизованная группа растений, чем моховидные. В настоящее время широко распространенны плауновые (Lycopodiales), являющиеся равноспоровыми, и также шильниковые, или полушниковые (Isoetopsida) – разноспоровыми растениями. Спорофиллы по форме, размерам и цвету похожи наобычные вегетативные листья. Чередуясь с ассимиляционными листьями, они образуют на стебле спороносные зоны или собраны в расположенные на верхушках ветвей стробилы, часто называемые спороносными колосками. Верхушечные побеги плауновых, являющихся равноспоровыми растениями, у большинства представителей рода плаун заканчиваются спороносными колосками – стробилами, состоящими из спорофилов, находящихся на стерженьках. Спорофиллы несут одиночные спорангии с большим количеством мелких тетраэдрической формы спор. При прорастании споры формируется маленький зародыш, представляющий собой обоеполый гаметофит без хлорофилла. В присутствии гиф гриба гаметофит медленно в течении 10-12 лет растет и развивается. На его верхней стороне образуются многочисленные антеридии и архегонии. Из образовавшейся после слияния половых гамет диплоидной зиготе вырастает взрослое растение. У представителей рода баранец спорофиллы не отличаются от обычных ассимиляционных листьев и отчетливо выраженных спороносных колосков виды этого рода не образуют. На верхушечных листьях баранца обыкновенного нередко образуются выводковые почки, при созревании специальный механизм отбрасывает их на расстояние до 0,5 м. У шильниковых, класс разноспоровых растений, спорангии располагаются на стебле, в пазухе листа или на его верхней поверхности. Гаметофиты однополые, сильно редуцированные, обычно не покидающие оболочки споры. У большинства силагинелл (Selaginella) колоски обоеполые, они расположены в одном стробиле. У них имеются спорангии двух типов – микроспорангии с большим числом мелких микроспор, и мегаспорангии – с четырьмя крупными мегаспорами. Сильно редуцированный мужской и женский гаметофиты обычно не покидают оболочку споры и после оплодотворения зародыш спорофита долгое время связан с гаметофитом, используя для роста запасные питательные вещества микроспоры. Содержимое микроспоры превращается в мужской заросток с одиночным антеридием, внутри которого после митоза образуются сперматозоиды со жгутиками. Заросток – редуцированное гаметофитное поколение, не способное фотосинтезу и питающееся за счет питательных веществ, содержащихся в споре. Из мегаспоры развивается заростки - женские гаметофиты, образующие ризоиды и способные к фотосинтезу. На поверхности женского заростка формируется архегоний, внутри которых находится по одной яйцеклетке, образовавшей в результате митоза. При этом зрелые гаметофиты плаунов не отделяются от спорофита. У полушниковых очень крупные мега- и микроспорангии образуются в ямках, лежащих в основании листьев на их верхней поверхности ниже основания язычка, и прикрыты пленчатой складкой – индузием. Редукция гаметофита у полушниковых усиливается, по сравнению с селагинеллами, и он представлен нередко всего лишь несколькими клетками. В мегаспорангиях образуется от 50 до 2350 мегоспор, а число микроспор в микроспорангии достигает миллиона. Хвощевидные (Equisetophyta). В современном растительном покрове хвощи представлены одним родом хвощ (Eqisetum). Стебли у них бывают зеленые или двух типов – зеленые (фотосинтезирующие) и бесхлорофильные (спороносные). У некоторых хвощевидных формируются спорангиофоры, спорофиллы особого строения, которые образуют на стебле спороносные зоны, чередующиеся с обычными вегетативными листьями, или сидят на концах осей, образуя стробилы. Спорангии у хвощей располагаются на спорангиофорах, расположенных чаще всего в верхушечных стробилах. Спорангиофоры в виде шестиугольных щитков на ножках размещены на оси стробила мутовками. На внутренней стороне щитка располагаются 4-16 вытянутых вдоль ножки спорангиев. При созревании спор щитки подсыхают и раздвигаются, наружная стенка спорангия при подсыхании легко разрушается, и споры, имеющие элатеры, рассеваются ветром. Хвощи равноспоровые растения, споры у них мелкие и многочисленные, часто снабжены элатерами. Одно- или обоеполый гаметофит (заросток) представлен очень маленьким, величиной в несколько миллиметров, зеленым растением. У хвоща полевого (Equisetum arvense) ранней весной на корневищах вырастают бледно-розовые спороносные побеги. На узлах этих побегов сидят мутовки листьев, а заканчиваются побеги спороносными колосками. У большинства других видов хвощей спороносные колоски образуются на верхушке ассимилирующих побегов. Развивающиеся из рассеивавшихся спор гаметофиты подушковидные и зеленые. Архегонии и антеридии погружены в ткань заростка. Оплодотворение осуществляется только при наличии воды на поверхности гаметофита. После оплодотворения образуется зигота и без периода покоя из нее вырастает новое бесполое поколение – спорофит. Папортниковидные, или папортники (Polypodiophyta). Большинство папортников относятся к равноспоровым растениям. Спорангии развиваются на обыкновенных зеленых листьях, на специальных спороносных частях листа или на специализированных листьях. Располагаются они одиночно или группами – сорусами. Сорусы расположены на нижней стороне листьев (спорофиллов). У многих папортников они состоят из выпуклого ложа (рецептакула), к которому с помощью ножек прикрепляются спорангии. Защиту развивающихся спорангиев обеспечивают покрывальца, или индузии, а также завернутый край пластинки листа, в их числе щитовник мужской, орляк обыкновенный, кочедыжник женский и т.д. Споры у них образуются в середине-конце летнего периода в спорангиях, которые сидят группами по 5-8 на нижней поверхности листа в сорусах. Внутри каждого спорангия в результате мейотического деления диплоидных материнских клеток спор образуются гаплоидные споры. При созревании спорангия с помощью специального приспособления в виде кольца споры рассеиваются с помощью тока воздуха. Из споры в благоприятных условиях развивается заросток (гаметофит), который у равноспоровых папортников гаметофит обитает на поверхности почвы и прикреплен к ней многочисленными ризоидами. Он обоеполый, зеленый, маленький, часто сердцевидной формы, быстро развивается и рано созревает. На нижней стороне заростка развиваются архегонии и антеридии. Оплодотворение протекает при наличии капельно-жидкой воды в дождливую погоду или при росе. Сперматозоиды имеют большое число жгутиков или имеет штопоровидную форму, активно двигаясь мужская гамета достигает яйцеклетки и сливается с ней. Движение сперматозоидов обусловлено реакцией хемотаксиса на яблочную кислоту, выделяемую клетками шейки архегония. Оплодотворение обычно перекрестное, поскольку антеридии созревают раньше архегониев. Зигота дает начало диплоидному зародышу, первое время который поглащает питательные вещества гаметофита, пока эту функцию не начнут выполнять собственные корни и листья спорофита. Гаметофит вскоре увядает и отмирает, а зародыш развивается в диплоидный спорофит. У разноспоровых папортников (марсилиевых и сальвиниевых), водных по своей экологии растений, гаметофит сильно редуцирован и имеет микроскопические размеры. У основания подводных листьев сальвинии плавающей (Salvinia natana), типичного водного папортника, гроздями висят шаровидные замкнутые сорусы, в которых располагаются спорангии различного размера – в одних в небольшом числе крупные мегаспорангии, в других – многочисленные мелкие по размерам микроспорангии. Полного развития в них получают лишь одна мегаспора и обычно 64 микроспоры соответственно. Мега- и микросорусы со спорангиями зимуют на дне водоема. Весной в результате разрушения индузия соруса спорангии всплывают. Многочисленные антеридии прорастают, из них выходят сперматозоиды, а из мегаспоры развивается зеленый веерообразный заросток – женский гаметофит с архегониями. Обилие питательных веществ в мегаспоре создает благоприятные условия для развивающегося половогопоколения – гаметофита. Из образовавшейся в результате оплодотворения зиготы формируется зародыш, а из него – взрослое растение. Первое время зародыш сальвинии питается за счет заростка – женского гаметофита |
neisri.narod.ru







