
Контрольная - Размножение растений - файл n1.doc. Размножение высших растений
Био: Вегетативное размножение высших растений
Вегетативное размножение - один из способов бесполого размножения; образование дочерних особей из многоклеточных вегетативных органов материнского организма (или частей этих органов). Оно очень распространено в природе. Благодаря вегетативному размножению растения быстро расселяются, захватывая новые территории. В некоторых случаях семенное размножение бывает затруднено, и вегетативное размножение оказывается единственным возможным путем. Кроме того, вегетативное размножение активно использует человек при размножении сельскохозяйственных и декоративных растений, т.к. оно позволяет сохранить ценные сортовые признаки.ВИДЕО 1. Размножение корнем. 1.1. Корневыми отпрысками - малина, яблоня, рябина, тополь, сирень, вишня и т.д. Из придаточных почек на корнях вырастают побеги - корневые отпрыски. со временем старые участки корней разрушаются, и отпрыски становятся самостоятельными растениями.
1. Размножение корнем. 1.1. Корневыми отпрысками - малина, яблоня, рябина, тополь, сирень, вишня и т.д. Из придаточных почек на корнях вырастают побеги - корневые отпрыски. со временем старые участки корней разрушаются, и отпрыски становятся самостоятельными растениями.  1.2. Корневыми черенками размножаются только те растения. у которых на корнях могут формироваться придаточные почки. Корневой черенок - это отрезок корня длиной 15-25 см, на котором могут образовываться придаточные почки. На посаженном в почву черенке образуются придаточные корни , а из почек развиваются надземные побеги. Одуванчик, шиповник, садовая малина.
1.2. Корневыми черенками размножаются только те растения. у которых на корнях могут формироваться придаточные почки. Корневой черенок - это отрезок корня длиной 15-25 см, на котором могут образовываться придаточные почки. На посаженном в почву черенке образуются придаточные корни , а из почек развиваются надземные побеги. Одуванчик, шиповник, садовая малина. 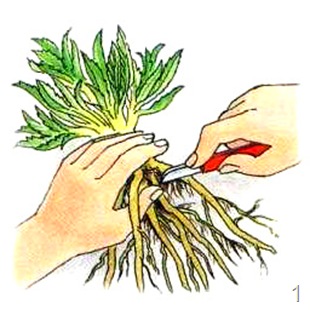
1.3. Корневыми клубнями. Корневые клубни - это подземные видоизмененные корни с запасом питательных веществ. На корнеклубнях могут развиваться придаточные почки. георгина.

2. Размножение листом.
так размножают некоторые виды комнатных растений - бегонию, сенполию (фиалку), сансевьеру. Листья сажают во влажный песок, и на них развиваются придаточные почки и придаточные листья. Иногда достаточно даже части листа.


3. Размножение надземным побегом.
3.1. Стеблевыми черенками (это отрезок побега с несколькими почками) размножают смородину, розу, тополь, иву и многие другие деревья и кустарники. Для этого черенки длиной 25-30 см сажают в хорошо подготовленную почву. К осени на черенках вырастут придаточные корни, и тогда черенки выкапывают и пересаживают на постоянное место.

3.2. Отводками размножают смородину, крыжовник, яблони, Для этого нижние ветви пригибают к земле и в месте пригиба надрезают кору. К осени в месте надреза развиваются придаточные корни, побег можно отрезать от материнского растения и пересадить на постоянное место.

3.3. Размножение ползучими побегами - "усами" характерно для живучки, лугового чая, земляники, хлорофитума; в узлах ползучих побегов образуются придаточные корни.

 3.4. Размножение прививкой в естественных условиях встречаются крайне редко. Но очень распространено в садоводстве. Почку-глазок или черенок культурного растения сращивают со стеблем-дичком (который обладает большей мощностью, неприхотливостью и морозостойкостью). Глазок или черенок культурного растения называют привоем, а дичок - подвоем. так размножают плодовые деревья - яблоню, грушу, вишню, сливу...
3.4. Размножение прививкой в естественных условиях встречаются крайне редко. Но очень распространено в садоводстве. Почку-глазок или черенок культурного растения сращивают со стеблем-дичком (который обладает большей мощностью, неприхотливостью и морозостойкостью). Глазок или черенок культурного растения называют привоем, а дичок - подвоем. так размножают плодовые деревья - яблоню, грушу, вишню, сливу... Прививка черенком - копулировка
 Прививка глазком - окулировка
Прививка глазком - окулировка 
4. Размножение подземными видоизмененными побегами.
4.1. Размножение корневищем характерно для огромного числа растений. многие из них являются злостными сорняками. Крапива, пырей, сныть, тысячелистник, ландыш, ветренница, орхидеи и др.

4.2. Луковицы характены для однодольных растений - тюльпанов, гиацинтов, амариллисов, нарциссов и др. Одна луковица может образовать несколько маленьких луковиц-деток, которые можно рассадить.

4.3. Клубнями размножается картофель и топинамбур. Если клубней недостаточно, можно использовать часть клубня с глазком (почкой).

5. Размножение культурой ткани. Мы живем в 21м веке, и на помощь сельскому хозяйству приходит клеточная инженерия. В специальных камерах на питательной среде выращиваются клетки растительных образовательных тканей, из которых формируются целые растения с признаками. идентичными признакам родительской особи. такой способ размножения позволяет получить большое количество посадочного материала, что немаловажно при введении нового сорта в культуру. Хорошо размножается культурой ткани картофель, гвоздика, герберы, орхидеи

myblog-bio.blogspot.com
Размножение растений | Агро Журнал
Каждый организм, в том числе и растение, имеет способность к воспроизведению себе подобных. Именно этот процесс и обеспечивает длительное, как правило, существование вида. Воспроизведение организмов возможно благодаря их способности производить потомство.В том случае, если образование потомства приводит к увеличению численности данного вида, т.е. дочерних особей становится больше чем материнских, этот процесс можно назвать размножением. Выделяют два основных способа размножения растений: бесполое и половое.
При бесполом размножении (в широком смысле) особь либо отделяет от себя неспециализированные вегетативные зачатки (почки, клубни, луковицы и пр.), и здесь мы имеем дело с вегетативным размножением. Нередко в качестве зачатков растение производит специализированные клетки, называемые спорами (греч. спора — посев). Спорами размножаются все низшие и многие высшие растения, за исключением голосеменных и цветковых, хотя и у последних образуются споры. Формирование спор на растении называют спороношением.
У многих низших растений, например водорослей, споры возникают в результате обычного деления клетки — митоза, поэтому и называют их митоспорами. Кроме того, как правило, споры низших растений (зооспоры) имеют жгутики, с помощью которых передвигаются в воде.У всех высших растений, в отличие от низших, споры развиваются в специализированных многоклеточных органах — спорангиях и образуются они в результате редукционного деления клетки — мейоза. Другими словами, все без исключения высшие растения образуют мейоспоры. Как правило, споры высших растений не имеют жгутиков и окружены твердой оболочкой.
В случае же полового процесса, при слиянии гаплоидных гамет образуется диплоидная зигота. Выделяют несколько основных типов полового процесса у растений.
1. Хологамия (греч. холос — целый, гамео — вступать в брак) — сливаются одноклеточные организмы. Такой способ характерен только для наиболее примитивных низших растений.
У большинства же, как низших, так и высших растений гаметы образуются в специализированных органах — гаметангиях.
Изогамия (греч. изо — равный) — в этом случае сливаются совершенно одинаковые по размеру и внешнему виду гаметы, одну из которых условно можно считать женской, другую — мужской.
Гетерогамия (греч. гетерос — различный) — постепенно в процессе эволюции гаметы становятся разных размеров: женские более крупные, хотя они и не утрачивают подвижность; подвижные мужские гаметы — сперматозоиды имеют более мелкие размеры.Оогамия (греч. оон — яйцо) — на данной стадии крупные женские гаметы (яйцеклетки) утрачивают подвижность, однако более мелкие мужские гаметы — сперматозоиды все еще ее сохраняют.
Необходимо отметить, что все без исключения высшие растения являются оогамными.
2. И, наконец, у семенных растений даже более мелкие мужские гаметы утрачивают подвижность. Такие неподвижные мужские гаметы называют спермиями.
Отдел Моховидные (Bryophyta)
Моховидные — это обширная группа растений (около 30000 видов), объединяемых нередко под общим названием — мхи. Однако в более точном, научном смысле мхами называют лишь представителей одного класса — настоящих или листостебельных мхов, поскольку кроме них отделе представлены еще 2 класса: антоцеротовые и печеночники.
Среди высших растений моховидные образуют наиболее обособленную группу. Своеобразие этой группы настолько велико, что в ботанике выделилась особая наука — бриология, в рамках которой занимаются исключительно изучением мхов. Для моховидных создаются совершенно особые определители и флоры, поскольку в стандартные общие флоры — они не вписываются.
Моховидная наиболее примитивная и древняя группа высших растений. Их древность вполне убедительно подтверждается ископаемыми находками. Во всяком случае, в карбоне они уже определенно существовали. Можно предположить, что именно в палеозое наметились основные линии развития моховидных, такие как печеночники, сфагновые и бриевые мхи.
Моховидные представляют собой не только древнюю группу. Это еще и слепая ветвь в эволюции растений.
Однако мхи заняли особое место в природе, особую нишу, и в настоящее время принимают заметное участие в сложении растительного покрова Земли. При этом они нередко оказывают существенное воздействие на среду обитания других растений и животных.
Так, в условиях повышенного увлажнения в умеренных широтах северного и южного полушарий идет болотообразовательный процесс, при этом накапливаются значительные толщи торфяных отложений с преобладающим участием моховидных. Особенно характерно это для областей, где осадки преобладают над испарением.
Например, заболоченность Западной Сибири по самым скромным оценкам составляет около 50% и эта территория по праву может считаться уникальным болотным регионом.
Общеизвестно ландшафтное значение моховидных в тундровой зоне, там даже выделяется особая подзона моховых тундр.
В достаточно больших количествах эти растения встречаются и в областях тропических. Так, в высокогорьях тропиков на высоте более 3000 м выделяется пояс мшистых лесов. Моховидные облепляют древесные растения настолько густо, что не видно даже стволов и ветвей. Все скрыто в сплошном зеленом футляре.
Если говорить об условиях обитания моховидных, то следует отметить, что их обычно рассматривают как растения, связанные в своей жизни с избыточным увлажнением. Однако значение этой зависимости не следует преувеличивать. Да, бриофиты действительно в изобилии растут на болотах, в сырых лесах и тундрах. Но есть среди них и ксерофиты. Так, в некоторых пустынях Tortula покрывает до 90% поверхности. Стало быть, существуют не только моховые тундры, но и моховые пустыни! Моховидные практически вездесущи.
Почти все моховидные обнаруживают, подобно лишайникам, поразительную устойчивость к длительному пересыханию. При увлажнении они снова оживают. Пожалуй, после лишайников моховидные являются самыми выносливыми растениями.
В одних случаях моховидные играют большую роль в сложении растительного покрова, в других незначительную, но всегда они непременный компонент растительного мира любого региона. Никакое ботаническое изучение территории, даже пустынной, не будет полным без учета бриофлоры.
Необыкновенно многообразны и субстраты, на которых селятся моховидные. Сравнительно немногие виды живут на поверхности почвы. Другие освоили скалы, камни, стволы, ветви, пни, валежины.
И, наконец, еще одна уникальная особенность моховидных. В сравнении с другими высшими растениями — это растения космополиты, они имеют чрезвычайно широкие ареалы (область распространения вида).
Причины: маленькие размеры тела, угнетенность полового процесса, повышенная толерантность (устойчивость), плюс дерновинная форма роста.
Таким растениям легче отыскать подходящую экологическую нишу, которую можно назвать микронишей.
Своеобразие моховидных, как самостоятельного отдела высших растений, особенно заметно проявляется в цикле их развития.
Жизненный цикл большинства растений складывается из 2-х фаз развития, или ядерных фаз, последовательно сменяющих друг друга: растение с диплоидным набором хромосом — спорофит, сменяется растением с гаплоидным набором хромосом — гаметофитом.
У очень небольшого числа низших растений спорофит и гаметофит неразличимы. Но у большинства низших и у всех высших растений (в том числе и моховидных) гаметофит и спорофит совсем непохожи друг на друга. Если не знать жизненного цикла растения, то спорофазу и гаметофазу можно принять за совершенно разные виды и это не раз случалось в истории ботаники.
Соотношение размеров и продолжительность жизни спорофита и гаметофита неодинаковы в разных группах растений. Но у большинства высших растений в жизненном цикле преобладает спорофаза. Под словом «преобладает» понимается такое положение, когда спорофит крупнее гаметофита и дольше живет. Собственно, все, что мы называем высшим растением: кочка осоки, цветущий куст, огромный ствол дерева с листьями, корнями, плодами и семенами — все это и есть спорофит.
Таким образом, можно смело утверждать, что подавляющее большинство высших растений пошло в эволюции по пути преимущественного развития спорофазы — спорофита (осуществили диплоидную линию эволюции). Гаметофит занимает весьма незначительную часть жизненного цикла. Более того, наблюдается тенденция к неуклонному сокращению роли гаметофазы в жизненном цикле. Если у папоротников гаметофит — это крохотное, самостоятельно питающееся растение — заросток, то у покрытосеменных гаметофит имеет микроскопические размеры, состоит из нескольких клеток и живет внутри спорофита. Чем выше по эволюционной лестнице продвинулось растение, тем более редуцированный гаметофит оно имеет.
Моховидные же представляют исключение. Они пошли в эволюции своей дорогой. У них все наоборот по сравнению с остальными высшими растениями, т.е. в жизненном цикле этих растений преобладает гаметофит. По сравнению с другими высшими растениями гаметофит значительно расширяет сферу своей физиологической деятельности: он не только обеспечивает половое размножение, но и выполняет основные вегетативные функции — фотосинтез, водоснабжение и минеральное питание. Однако стебли и листья, развивающиеся на гаметофите моховидных не являются настоящими, поэтому стебли моховидных в бриологии принято называть каулидиями, а листья — филлидиями.
Спорофит же у моховидных имеет меньшие размеры, лишен листьев и фактически низведен до роли спороносящего органа.
Однако рассмотрим все по порядку.
Из одноклеточной гаплоидной споры у моховидных развивается многоклеточная ветвистая зеленая нить — протонема. Она очень похожа на нитчатые зеленые водоросли. Однако в отличие от зеленых водорослей протонема имеет не прямые, а косые перегородки между клетками и хлоропласты вместо хроматофоров.
На протонеме появляются многочисленные выводковые почки, из которых вырастают облиственные побеги гаметофоры. Их так называют потому, что на гаметофорах образуются половые органы — гаметангии: мужские — антеридии и женские — архегонии.
Поскольку на протонеме мхов образуются многочисленные почки, большинству из них присущи групповые формы роста — плотные, компактные дерновинки. Это дает мхам возможность переносить неблагоприятные условия, быстрее поглощать и сохранять влагу. Компактное произрастание растений также обеспечивает половое размножение.
Виды моховидных можно разделить на однодомные и двудомные растения. У однодомных — половые органы — антеридии и архегонии находятся на одном растении, у двудомных — на разных. Так, кукушкин лен является двудомным растением.
Гаметангии располагаются обычно не поодиночке, а группами на верхушках облиственных гаметофоров, причем перемежаются с многочисленными стерильными нитями — парафизами. Парафизы защищают гаметангии и предохраняют их от высыхания. Обычно скопления гаметангиев окружены специальными, сильно видоизмененными листовыми выростами.
Антеридии имеют вид дисковых мешочков на ножке с однослойной оболочкой. Все их внутреннее содержимое заполнено сперматогенной тканью. Из каждой клетки этой ткани образуются подвижные двухжгутиковые сперматозоиды.
Архегоний, в типичном случае, представляет собой колбовидное тельце, с расширенной нижней частью — брюшком, где помещается крупная яйцеклетка. В брюшке также размещается брюшная канальцевая клетка. Горловина архегония называется шейкой, в ней расположены шейковые канальцевые клетки.
При созревании архегония брюшная и шейковые канальцевые клетки расслизняются и слизь немного выступает наружу. При этом внутри архегония образуется своеобразный слизистый канал, по которому сперматозоиды легко достигают яйцеклетки: слизь облегчает их продвижение.
Оплодотворение происходит при наличии капельножидкой воды. Сперматозоиды подплывают к архегонию, проникают внутрь и, двигаясь по слизистому каналу, подходят к яйцеклетке. При слиянии гамет (сперматозоида и яйцеклетки) появляется диплоидная зигота и ее дальнейшее развитие происходит внутри архегония.
Из зиготы формируется спорофит, который у мхов называется — спорогоном. Это очень яркое и меткое название. Действительно спорогон моховидных растений не имеет никакой иной функции кроме производства спор (гонит споры).
Строение спорогона удобно рассмотреть на примере кукушкина льна обыкновенного (Politrichum commune). Он представляет собой коробочку на ножке, прикрытую волосистым колпачком.
Интересно отметить, что спорогон в целом — диплоидное образование, в то время как сидящий на его вершине колпачок — гаплоидный. Как такое могло получиться? Когда зигота начинает делиться внутри архегония — она его разрывает и растущий спорогон поднимает верхнюю часть архегония вверх. Эта верхняя часть архегония впоследствии и преобразуется в колпачок.
Сама коробочка состоит из нескольких основных структурных частей. В ее составе можно выделить урночку — своеобразную емкость для спор. Сверху урна прикрыта крышечкой, наподобии туристского котелка. Нижняя расширенная часть урны называется апофизой. В апофизе имеются хлоропласты, за счет которых спорогон может фотосинтезировать и, хотя бы частично, снабжает себя пластическими веществами. Отверстие урны, помимо крышечки, прикрыто тонкой белой пергаментообразной пленочкой — эпифрагмой. После отделения крышечки эпифрагма надежно прикрывает устье урны и препятствует преждевременному высыпанию спор.
В центре разрезанной повдоль коробочки обнаруживается колонка, которая по периметру окружена полостью. В этой полости размещается так называемый споровый мешок. Он подвешен к внутренним стенкам урны на особых флорофиллоносных нитях, которые играют роль своеобразных аммортизаторов.
Споры образуются в результате редукционного деления спорогенной ткани, заполняющей споровый мешок. Кукушкин лен обладает весьма тонким механизмом высеивания спор.
Дело в том, что пленочка — эпифрагма крепится к краям зубцов так называемого перистома, окаймляющих устье урны. Клетки зубцов перистома имеют неравномерно-утолщенные оболочки. Кроме того, перистом отличается удивительной гигроскопичностью, способностью впитывать влагу. Поэтому во влажную погоду зубцы перистома набухают, изгибаются и плотно прижимают эпифрагму к отверстию урны, препятствуя прорастанию спор внутри урны. В сухую погоду зубцы перистома расправляются (выпрямляются), теряя влагу, и эпифрагма слегка приподнимается над отверстием урны. В образовавшиеся между зубцами прорехи небольшими порциями высыпаются споры, когда коробочка раскачивается на ветру. При этом в сухую погоду споры разносятся на большое расстояние.
Таким образом, развитие спорофита как самостоятельного организма у моховидных не происходит. Он прикреплен к гаметофиту и живет в значительной мере за его счет. Еще совсем недавно считалось, что спорофит мохообразных паразитирует на гаметофите. Однако это не вполне верно, поскольку спорофит, по крайней мере, в молодом возрасте — фотосинтезирует, и, значит, о полном паразитизме говорить нельзя.
Другие исследователи считают, что спорофит произрастает на гаметофите эпифитно.
Правильнее будет сказать, что обе фазы образуют сложный двуединый организм, с главенствующей ролью гаметофита.
Следует отметить, что размножение с помощью спор в большинстве групп моховидных не является главным.
Для того, чтобы возник спорофит, необходимо оплодотворение. Для многих видов это почти непреодолимая трудность. Дело в том, что для этого необходима вода, в которой могут перемещаться сперматозоиды. И если антеридии и архегонии разделены пространством хотя бы в несколько сантиметров, то даже дождевой воды может не хватать, для реализации оплодотворения. А ведь нередко мужские и женские особи разделены сотнями и даже тысячами метров. Есть, например, очень интересный вид Tortula, у которого мужские особи отмечены только в Европе, а женские — исключительно в Северной Америке. В таких условиях оплодотворение просто нереально.
И моховидные нашли выход в исключительно активном вегетативном размножении. Практически любая часть гаметофита — кусочки листьев, стеблей при благоприятных условиях может дать протонему, а через нее и новые растения. Большой кусочек слоевища или веточка могут развиваться в целое растение, минуя стадию протонемы.
Моховидные в подавляющем большинстве многолетние растения. Но независимо от продолжительности жизни все они низкорослы. Размеры моховидных колеблются от миллиметров до нескольких сантиметров у большинства видов. Значительно реже встречаются более крупные моховидные. Так, живущие в воде мхи рода Fontinalis могут иметь стебли длиной до 60 см. и более. Такими же длинными являются и стебли некоторых мхов — эпифитов из рода Dawsonia, поселяющихся на стволах и ветвях деревьев. Это — гиганты среди мохообразных.
В силу многих причин гаплоидная линия эволюции, избранная моховидными, оказалась невыгодной. Остались отрезанными многие возможные пути эволюционного развития. Именно поэтому моховидные обречены оставаться мелкими растениями, кроме того, у них слабо развита водопроводящая и механическая система.
В целом, структурные возможности гаплоидного гаметофита оказались значительно ниже.
Моховидные — единственный отдел растительного царства в жизненном цикле которых безраздельно и полно господствует гаметофаза.
www.agrojour.ru
Споровые растения - высшие, размножение, цикл
Споровые растения
К споровым относятся растения. размножение и распространение которых осуществляется посредством спор, причем споры образуются двумя путями – бесполым и половым. Споровыми растениями являются водоросли. грибы. лишайники и высшие споровые растения (папоротники. плауны. хвощи. мхи. некоторые ископаемые растения).
В процессе эволюции около 400 млн. лет назад от многоклеточных зеленых водорослей возникли риниофиты – первые высшие растения, размножающиеся спорами, которые дали начало всем современным высшим споровым и семенным растениям. Это вымершая группа растений. В жизненном цикле высших споровых растений, как и у некоторых водорослей, чередуются особи бесполового и полового поколений, которые размножаются, соответственно, бесполым и половым путем. В полном жизненном цикле, который обеспечивает непрерывность жизни организмов, происходит чередование гаметофита (полового) и спорофита (бесполого поколения). На спорофите образуются органы бесполого размножения, на гаметофите – полового.
В жизненном цикле высших споровых растений, как и у некоторых водорослей, чередуются особи бесполового и полового поколений, которые размножаются, соответственно, бесполым и половым путем. В полном жизненном цикле, который обеспечивает непрерывность жизни организмов, происходит чередование гаметофита (полового) и спорофита (бесполого поколения). На спорофите образуются органы бесполого размножения, на гаметофите – полового.
Высшие споровые растения после выхода на сушу в ходе эволюции претерпевали метаморфозы в двух направлениях. Так сформировались две крупные эволюционные группы – гаплоидная и диплоидная. К первой ветви относятся мхи, у которых лучше развит гаметофит, а спорофит занимает подчиненное положение. К диплоидной ветви принадлежат папоротники, хвощи и плауны. У них гаметофит редуцирован, и выглядит как заросток.
Размножение
Из спор, которые формируют особи бесполого поколения, вырастают особи полового поколения. Они имеют специальные мужские и женские половые органы, в которых развиваются мужские и женские половые клетки (гаметы) – подвижные сперматозоиды и неподвижные яйцеклетки. Для оплодотворения сперматозоид должен попасть во внешнюю среду и оплодотворить яйцеклетку, которая находится внутри женского полового органа. Для перемещения сперматозоида необходима вода. Из оплодотворенной яйцеклетки образуется зародыш. Он прорастает и превращается в особь бесполого поколения, которая размножается спорами.
Грибы, лишайники и водоросли размножаются подвижными зооспорами, спорангиеспорами, неподвижными апланоспорами. Для этих растений характерно половое размножение ооспорами, зигоспорами, вегетативное размножение также посредством спор. Из спор появляется такое же растение, как материнское. Тогда как у высших споровых растений такое растение, как то, что сформировало споры, появляется после сложного процесса оплодотворения.
Спора представляет собой клетку с двойной оболочкой. Споры у определенного вида растений могут быть одинаковыми по размерам или разными. Микроспоры – мелкие, макроспоры – крупные. Микроспоры обычно дают начало мужским заросткам, а макроспоры – женским.
Высшие споровые растения распространены в разных климатических условиях, но большинство из них обитает на влажных участках суши, так как для полового размножения им нужна вода. Однако некоторые виды этих растений встречаются и в пустынях.
Рекомендуем ознакомится: http://beaplanet.ru
worldunique.ru
Контрольная - Размножение растений - n1.doc
Контрольная - Размножение растенийскачать (269.5 kb.)Доступные файлы (1):
n1.doc
Реклама MarketGid: План 1. Размножение как одно из основных свойств живого организма. Типы размножения. Способность к регенерации как основа вегетативного размножения. Способы вегетативного размножения 32. Бесполое размножение спорами. Спорангии, их строение и расположение у разных растений. Изоспория и ее биологическое значение 6
3. Половое размножение высших растений. Гаметы и зигота. Гаметофит и спорофит. Чередование поколений у высших растений и его значение. Закономерности эволюции размножения высших растений. Закономерности эволюции размножения высших растений. 8
4. Размножение хвойных растений. Анатомо-морфологические особенности спорофита. Строение шишек. Мега- и микроспорогенез. Оплодотворение. Развитие семян 10
Список использованной литературы 13
1. Размножение как одно из основных свойств живого организма. Типы размножения. Способность к регенерации как основа вегетативного размножения. Способы вегетативного размножения
Размножение или репродукция ? одно из основных свойств живого. Размножение ? способность производить себе подобных особей. Целостныйорганизм состоит из дискретных единиц ? клеток. Существование вида поддерживается размножением.
Известны две формы размножения: бесполое и половое.
При половом размножении необходимо, как правило, наличие двух особей, и новый организм возникает из специализированных половых клеток или особей, выполняющих эти функции.
В основе полового размножения лежит половой процесс ? объединение в наследственном материале генетической информации из разных источников ? родителей ? для развития потомка. Организмам свойственна двойственная наследственность.
Преимущество полового размножения состоит в перекомбинации лучших наследственных признаков обоих родителей, что является источником изменчивости. Потомство более жизнеспособно и приспособлено к условиям существования. Быстрее происходит эволюция.
Бесполое размножение широко распространено во всех группах растений. В простейшем виде при таком типе размножения родительская особь разделяется на две части, каждая из которых развивается в самостоятельный организм. Этот способ размножения, называемый делением, встречается, как правило, лишь у одноклеточных организмов. Клетка при этом делится путем митоза.
Многие многоклеточные организмы также способны успешно размножаться путем отделения жизнеспособных участков вегетативного тела, из которых формируются полноценные дочерние особи. Этот тип бесполого размножения в мире растений нередко называют вегетативным. Способность к вегетативному размножению очень характерна для растений и грибов на всех уровнях их организации.
В основе вегетативного размножения лежит его способность к регенерации, т.е. к восстановлению утраченных или поврежденных органов, а также развитию целого растения из отдельных частей. Регенерация происходит благодаря интенсивному делению клеток. Степень регенерации зависит от видов растений, а также от тех факторов, которые могут стимулировать или тормозить деление клеток – это условия внешней среды и питания, стимуляторы или ингибиторы роста и др. Предполагается, что механизм воздействия разных факторов на степень регенерации заключается в изменении физиологического состояния, в первую очередью тех клеток и тканей, которые непосредственно участвуют в регенерации, а затем уже смежных тканей, органов и всего организма в целом.
Нередко при этом растения размножаются обрывками или частями таллома или частями вегетативных органов. Многие нитчатые и пластинчатые водоросли, талломы лишайников свободно распадаются на части, каждая из которых легко становится самостоятельным организмом. Так могут размножаться и некоторые цветковые растения, обитающие в воде. Примером растения, размножающегося исключительно вегетативно, на территории Европы является двудомная элодея канадская (Elodea canadensis), попавшая сюда из Северной Америки. При этом в Европу были занесены лишь женские экземпляры, не способные образовывать семена в отсутствие мужских растений. Несмотря на отсутствие семенного возобновления растение это исключительно быстро размножается и стремительно осваивает новые местообитания.
Вегетативное размножение может быть естественным и искусственным (рис. 1).
Для естественного вегетативного размножения могут служить специальные образования, имеющие запас питательных веществ: клубни (картофель), луковицы (лук, тюльпан), клубнелуковицы (шафран), корневища (осот, ландыш), усы (земляника).
Для искусственного вегетативного размножения используют части самого растения: черенки (часть стебля), листья (бегония), отводки (ветви пригибают к земле и на них образуются придаточные корни (например, у малины), корневые отпрыски, производят деление кустов, прививки. Прививка состоит в пересадке одного растения (побега или почки) на нижнюю часть другого побега. Пересаженную часть растения ? донора называют привоем, а реципиента ? подвоем. Получившееся растение имеет корневую систему подвоя, а побег ? привоя.

Рисунок 1 ? Вегетативное размножение растений:
1 ? вегетативное размножение земляники надземными ползучими побегами,
2 ? подземные клубни картофеля (темный клубень ? старый, из которого выросло все растение), 3 ? молодило размножается откидышами,
4 ? луковички в соцветии дикого лука, 5 ? луковицы ? « детки» в луковице,
6 ? клубнелуковица шафрана, 7 ? то же в продольном разрезе, 8 ? корневище осоки,
9 ? корневище ириса, 10 ? корневище купены, 11 ? элодея, размножающаяся частями побегов.
2. Бесполое размножение спорами. Спорангии, их строение и расположение у разных растений. Изоспория и ее биологическое значение
Размножение спорами свойственно водорослям, мхам и папоротникообразным. Споры развиваются в специальных органах-спорангиях.Спорангий ? одноклеточный (у многих низших растений) или многоклеточный (у высших) орган, в котором образуются споры. Некоторые одноклеточные зелёные водоросли целиком превращаются в спорангий. У многоклеточных водорослей, например у улотрикса, ульвы, Спорангием могут стать клетки, не отличающиеся от других клеток. У эктокарпуса, ламинарии спорангием становятся клетки, морфологически отличающиеся от других и занимающие на талломе определённое положение. У некоторых неклеточных растений спорангии образуются на талломах, отделяясь перегородкой. У некоторых оомицетов спорангии становятся конидиями, опадают и прорастают. Названия спорангиев. низших растений отражают особенности строения возникающих в них спор (зооспорангий), их число (моноспорангий, тетраспорангий), внешний вид спорангия (цистокарпий), способ образования спор (митоспорангии, мейоспорангии) и т.п. С мейоспорангиями связана смена ядерных фаз в циклах развития растений. Высшие растения образуют только мейоспорангии. У мохообразных спорангий представлен коробочкой спорогония. Спорангий папоротникообразных развиваются на спорофиллах или в их пазухах. Спорангии могут быть одиночными или развиваться группами (сорусы), свободными или сросшимися (синангии). Равноспоровые папоротникообразные образуют спорангии одного типа, продуцирующие споры, прорастающие в обоеполые заростки, разноспоровые ? спорангии двух типов (микро- и мегаспорангии), образующие микро- и мегаспоры, из которых развиваются мужские и женские заростки. Все семенные растения ? разноспоровые. Мегаспорангию у них гомологичен нуцеллус семезачатка. Микроспорангию у покрытосеменных гомологично гнездо пыльника.
Изоспория ? равноспоровость, образование у растений спор равного размера. Характерна для папоротников (исключая водные папоротники ? сальвинии, марсилии, азоллы), хвощей, плаунов (исключая селагинеллы). У некоторых хвощей из внешне одинаковых спор развиваются в зависимости от условий питания и освещения более мелкие ? мужские заростки с антеридиями или более крупные женские с архегониями. Таким образом, регулируя условия прорастания спор, можно изменить пол заростка.
3. Половое размножение высших растений. Гаметы и зигота. Гаметофит и спорофит. Чередование поколений у высших растений и его значение. Закономерности эволюции размножения высших растений. Закономерности эволюции размножения высших растений.
Половое же размножение осуществляется путем слияния (копуляции) мужской и женской гамет с образованием зиготы. Половое семенное размножение происходит только у семенных растений.Спорофит ? бесполое поколение, образует споры. В результате прорастания спор образуется гаметофит.
Гаметофит ? половое поколение, образует гаметы. При слиянии гамет (оплодотворении) образуется зигота, из которой вырастает спорофит.
Чередование поколений у высших растений ? закономерная смена в жизненном цикле организмов поколений, размножающихся половым и бесполовым путем.
В жизненном цикле всех высших растений, за исключением мохообразных, спорофит резко преобладает над гаметофитом, особенно у древесных форм. Эволюция высших растений шла по пути преобразования и усложнения органов размножения с сокращением и последующей редукцией гаметофита как самостоятельного растения. Параллельно происходило усложнение процесса полового размножения. Микроспоры постепенно преобразовались в пыльцу, а спорангии со спорами – в семязачатки. После оплодотворения семязачатка пыльцой возникает семя. В соответствии со способом размножения все высшие растения делят на споровые и семенные.
Жизненный цикл схематически можно представить в следующем виде: спорофит ? споры ? прорастание ? гаметофит ? гаметы (n) ? оплодотворение ? зигота ? (2 n) ? спорофит.
Гаметы: женская ? одна неподвижная яйцеклетка, мужские ? много. У споровых ? сперматозоиды (имеются жгутики, подвижны), у семенных ? спермии (жгутиков нет, неподвижны).
Споровые растения. Гаметофит вырастает из споры как самостоятельный организм, на нем образуются мужские и женские половые органы, а в них ? гаметы. Сперматозоиды выходят наружу и активно передвигаются в воде к женским половым органам, где происходит оплодотворение. Образуется зигота, из которой на гаметофите вырастает спорофит. Он производит споры, которые служат для расселения.
Мхи. Спорофит и гаметофит не разделены. Само растение (гаметофит) ? преобладает. Спорофит (коробочка со спорами) образуется на гаметофите.
Папоротники, хвощи, плауны. Спорофит и гаметофит разделены. Само растение (спорофит) ? преобладает. Гаметофит (заросток) ? отдельно живущее маленькое недолговечное растение. На нем вырастает спорофит. Вначале гаметофит обеспечивает его питанием.
Семенные. Спорофит и гаметофит не разделены. Само растение (спорофит) ? преобладает. В половых органах спорофита (шишки или цветки) образуются и прорастают споры, формируя очень маленькие гаметофиты (несколько клеток). Мужские гаметофиты (пыльцевые зерна) переносятся к женским (опыление) и прорастают: формируется пыльцевая трубка, доставляющая спермии к яйцеклетке. В результате оплодотворения возникает зародыш ? новое поколение спорофита. Зародыши находятся внутри семян, которые служат для расселения.

Рисунок 2 ? Редукция гаметофита в эволюции
4. Размножение хвойных растений. Анатомо-морфологические особенности спорофита. Строение шишек. Мега- и микроспорогенез. Оплодотворение. Развитие семян
Особенности размножения хвойных рассмотрим на примере сосны обыкновенной (Pinus sylvestris).Сосна ? крупное дерево, размножающееся семенами. Семена сосны и других хвойных образуются в шишках. Эти шишки называются женскими стробилами.
Менее известны так называемые мужские шишки (стробилы), в которых созревает пыльца. Они появляются ранним летом и существуют весьма непродолжительное время.
Строение мужского стробила. Микроспорогенез и микрогаметогенез. На продольном срезе через мужскую шишку хорошо выделяется ось, к которой крепятся микроспорофиллы. В основании каждого из них размещаются гнезда двух крупных спорангиев (пыльцевых мешков). Гнезда микроспорангиев заполнены археспориальной тканью. Процесс микроспорогенеза состоит в том, что клетки археспориальной ткани делятся путем мейоза, в результате образуются тетрады гаплоидных микроспор.
Каждая микроспора (пыльцевое зерно) имеет две оболочки: внутреннюю тонкую ? интину и наружную, грубую и прочную ? экзину. Характерной особенностью микроспор хвойных являются так называемые воздушные мешки, повышающие аэродинамические качества пыльцы, которая распространяется ветром. Гаплоидное ядро микроспоры начинает делиться обычным митотическим путем. В результате двух, следующих друг за другом митотических делений, образуется четыре клетки: две проталлиальные, антеридиальная и сифоногенная.
Проталлиальные клетки являются рудиментом вегетативного тела заростка. Эти клетки существуют очень непродолжительное время и вскоре разрушаются.
Из сифоногеннойклетки развивается пыльцевая трубка. Все дальнейшие изменения происходят с антеридиальной клеткой. Она делится еще один раз и, в результате этого деления, появляются клетка-ножка и спермагенная клетка. Назначение клетки ножки не вполне понятно, а вот при делении спермагенной клетки образуется две мужские гаметы ? спермии. На этом процесс микрогаметогенеза заканчивается.
Строение женского стробила. Мегаспорогенез и мегагаметогенез. Женская шишка (стробил) также имеет ось, к которой крепятся чешуи двух типов: пленчатые кроющие чешуи, в пазухах которых развиваются крупные плодущие или семенные чешуи. В основании семенных чешуй размещается по два семязачатка.
Оба процесса: мегаспорогенез и мегагаметогенез происходят в семязачатке. Процесс мегаспорогенеза организован очень просто: внутри нуцеллуса обособляется клетка археспория, которая и становится материнской клеткой мегаспор. В результате редукционного деления этой клетки образуется тетрада мегаспор, три из которых быстро погибают и идут на питание единственной оставшейся мегаспоре.
Из гаплоидной мегаспоры вырастает женский гаметофит, который со временем занимает практически весь объем семени. По своей функциональной роли женский гаметофит является питательной тканью семени ? эндоспермом. Необходимо подчеркнуть, что эндосперм голосеменных растений является гаплоидной тканью, в отличие от триплоидного эндосперма цветковых растений.
На верхнем микропилярном полюсе гаметофита образуются два типичных архегония. В брюшке каждого из них находится крупная яйцеклетка. После формирования яйцеклетки женский гаметофит готов к оплодотворению.
Оплодотворение совершается внутри семязачатка, при этом спермии по пыльцевой трубке проникают к архегониям и один из них сливается с яйцеклеткой. Вторая пара гамет обычно не дает зиготы. Такой тип оплодотворения называют сифоногамией.
Из зиготы со временем развивается зародыш семени. При делении зиготы образуется две клетки: терминальная и базальная. Из терминальной клетки развивается так называемый подвесок, а из базальной ? зародыш.
Список использованной литературы
- Андреева И.И., Родман Л.С. Ботаника. ? М: Колос, 2005. ? 528с.
- Бавтуто Г.А., Еремин В.М. Ботаника. Морфология и анатомия растений. Учеб. пособие. ?Мн., Высшая школа, 1997. ? 375с.
- Барабанов Е.И. Ботаника: учебик для студ.высш.учеб.заведений. ? М: Академия, 2006. ? 448с.
- Ботаника. Систематика высших, или наземных, растений / А. Г. Еленевский, М. П. Соловьева, В. Н. Тихомиров. ? М.: Академия, 2004. ? 432с.
- Чебышев Н. В. Биология (Учебник) / Н.В.Чебышев, Г.Г. Гринева, М.В. Козарь, С.И. Гуленков. ? М.: ВУНМЦ, 2000. ? С.81?84.
gendocs.ru









