
Анатомия корня: первичное и вторичное анатомическое строение корня. Растения у которых формируется вторичная анатомическая структура корня
типы, особенности строения, признаки, имеющие значение для описания и диагностики корней.
Зона делящихся клеток, или конус нарастания (апекс) — самая молодая часть длиной в несколько миллиметров. Состоит из клеток-инициалей апикальной меристемы, которые находятся в постоянном делении, и их производных. Кончик корня прикрыт корневым чехликом, который предохраняет молодые клетки от механических и других повреждений. Образуется он наружным (калиптрогенным) слоем меристемы. Имеет клиновидную форму и состоит из живых паренхимных клеток с высоким тургором и тонкими ослизняющимися оболочками. Поверхностный слой чехлика постоянно разрушается, но восполняется изнутри новыми клетками. В корневом чехлике главного корня имеются зерна оберегаемого крахмала, чувствительные к гравитации и обеспечивающие положительный геотропизм — рост корня вертикально вниз.
Зона роста и дифференцировки (2–5 мм) характеризуется растяжением клеток, что обусловливает удлинение корня и появление дерматогена, периблемы и плеромы — предшественников эпиблемы, первичной коры и центрального цилиндра, соответственно.
Зона всасывания, или зона корневых волосков (5–20 мм), расположена над зоной роста, характеризуется наличием корневых волосков — трубчатых выростов клеток эпиблемы. В этой зоне завершается дифференцировка клеток первичных тканей, формируется первичное анатомическое строение, осуществляется поглощение воды и минеральных растворов из почвы.
Зона проведения и укрепления расположена выше зоны всасывания. Она обеспечивает передвижение двух токов веществ, а также укрепление растения благодаря образованию боковых корней. В этой зоне у однодольных растений сохраняется первичное строение с небольшими изменениями. У двудольных и голосеменных в связи с появлением и деятельностью камбия и феллогена корень утолщается и приобретает вторичное анатомическое строение. В некоторых корнеплодах эта зона значительно утолщается за счет разрастания запасающей ткани.
П е р в и ч н о е с т р о е н и е к о р н я характеризуется наличием трех систем тканей — покровно-всасывающей эпидермы с корневыми волосками (эпиблемы), первичной коры и центрального цилиндра.
Эпиблема выполняет всасывающую и защитную функции, состоит из одного слоя тонкостенных, плотно сомкнутых клеток с корневыми волосками, не имеет устьиц, толстой кутикулы и трихом. Поверхность клеток ослизняется, что предохраняет корень от иссушения, излишнего трения, способствует прилипанию и смягчению частичек почвы, развитию полезных бактерий.
Первичная кора, лежащая под эпиблемой, подразделяется на экзодерму, мезодерму и эндодерму. Экзодерму составляет 3–4 слоя крупных, многоугольных, плотно сомкнутых клеток с частично опробковевшими оболочками. Выполняет защитную и опорно-пропускную функции. Мезодерма — многослойная запасающая часть, составляет основную массу первичной коры. Клетки, как правило, живые, крупные, округлые или многоугольные, рыхлые, с тонкими или утолщенными лигнифицированными (у однодольных) оболочками, заполнены обычно крахмальными зернами. Мезодерма выполняет также транспортную и воздухоносную функции. Эндодерма — самый внутренний, обычно однорядный слой первичной коры, граничащий с центральным цилиндром, выполняющий опорно-пропускную функцию. У однодольных утолщаются и опробковевают радиальные и внутренние тангентальные оболочки в виде буквы U. Среди этих мертвых клеток эндодермы напротив лучей ксилемы сохраняются живые пропускные клетки, по которым вода и растворы минеральных веществ проходят к сосудам. У двудольных обычно лишь радиальные (антиклинальные) стенки эндодермы имеют линзовидные опробковевшие утолщения — пятна, или пояски Каспари.
Центральный, или осевой, цилиндр занимает срединную часть корня, окружен кольцом перицикла, из которого формируются боковые корни, а у голосеменных и двудольных также камбий и феллоген, обеспечивающие вторичное утолщение корня. Под перициклом расположен радиальный проводящий пучок. В зависимости от количества участков ксилемы он бывает двух-, трех-, четырех-, пяти-, шести- и многолучевой, или -архный. У двудольных растений лучей ксилемы обычно не более шести, а у однодольных — больше шести (пучок полиархный). В корне нет настоящей сердцевины. В центре могут находиться сосуды, склеренхима, иногда — запасающая паренхима, образующая ложную сердцевину.
В т о р и ч н о е с т р о е н и е к о р н я формируется в зоне укрепления у голосеменных и двудольных покрытосеменных растений благодаря появлению и функционированию вторичных боковых меристем — камбия и феллогена. Камбий образуется из перицикла над лучами первичной ксилемы и из прокамбия или паренхимы под флоэмой. Кнаружи делящийся камбий откладывает вторичную флоэму, а к центру— вторичную ксилему. Первичная ксилема оттесняется к оси корня, а первичная флоэма — к периферии центрального цилиндра. Вторичное строение корня может быть пучкового типа (у некоторых травянистых двудольных растений) или беспучкового (у древесных и некоторых травянистых двудольных). Пучковый тип формируется в том случае, если межпучковый камбий, образовавшийся над лучами первичной ксилемы, продуцирует паренхиму сердцевинных лучей, а пучковый камбий, лежащий под флоэмой,— вторичную флоэму и ксилему коллатерального или биколлатерального проводящего пучка. При беспучковом типе строения кольцо камбия образует сплошные кольца ксилемы и флоэмы. Вторичные преобразования в первичной коре корня связаны с деятельностью феллогена. Если он возникает из перицикла, то пробка, образующаяся кнаружи, изолирует первичную кору, и она отторгается, слущивается. Если феллоген образуется из паренхимы мезодермы, что встречается редко, то часть первичной коры, лежащая под феллогеном, сохраняется. У древесных растений корень обычно беспучкового типа и в древесине формируются кольца годичного прироста. Годичное кольцо — это совокупность тканей, образованных камбием за один вегетационный период. Покровной тканью корней древесных может быть перидерма или корка. Наличие в центре органа радиального проводящего пучка, склеренхимы или сосудов первичной ксилемы отличает корень от стебля и корневища, у которых центральную часть занимает настоящая сердцевина. Отличительной особенностью утолщенных корней — корнеплодов — является значительное разрастание запасающей паренхимы в лубе (сем. зонтичные) или в древесине (сем. крестоцветные). В корнеплодах типа свеклы (сем. маревые) центральный цилиндр сначала имеет беспучковое строение. Затем вокруг него в результате деления клеток первичной флоэмы и перицикла образуются паренхимные кольца. В них возникают кольца камбия. Одни его участки (пучковый камбий) формируют открытые коллатеральные пучки, а другие, лежащие вне пучков (межпучковый камбий), образуют межпучковую запасающую паренхиму. В корнеклубнях, или шишках, представляющих собой утолщенные боковые или придаточные корни (георгин, ятрышник, чистяк и др.), как и в корнеплодах, хорошо развита запасающая ткань, среди которой проходят очень мелкие проводящие пучки.
40. Побег: определение, функции, отличие от корня; составляющие побега; разнообразие побегов зависимости от длины междоузлий, способов нарастания, степени и типа ветвления, положения в пространстве, формы поперечного разреза стебля т.д..
Побег (как и корень) - основной орган растения. Вегетативные побеги в типичном случае выполняют функцию воздушного питания, но имеют ряд других функций и способны к разнообразным метаморфозам. Спороносные побеги (в том числе и цветок) специализированы как органы репродуктивные, обеспечивающие размножение.
Побег образуется верхушечной меристемой как единое целое и, поэтому, представляет собой единый орган того же ранга, что и корень. Однако по сравнению с корнем побег имеет более сложное строение. Вегетативный побег состоит из осевой части – стебля, имеющего цилиндрическую форму, и листьев – плоских боковых органов, сидящих на стебле. Кроме того, обязательной частью побега являются почки – зачатки новых побегов, обеспечивающие нарастание побега и его ветвление, т.е. образование системы побегов. Главную функцию побега – фотосинтез – осуществляют листья; стебли – преимущественно несущие органы, выполняющие механическую и проводящую функции.
Главная черта, отличающая побег от корня, - его облиственность. Участок стебля, от которого отходит лист (листья) называется узел. Участки стебля между соседними узлами – междоузлия. Узлы и междоузлия повторяются вдоль оси побега. Таким образом, побег имеет метамерное строение, метамером (повторяющимся элементом) побега являются узел с листом и пазушной почкой и нижележащее междоузлие.
П о б е г — надземный листостебельный орган высших растений, формирующий внешний облик растения — габитус, обеспечивающий воздушное питание. Элементы побега — стебель, почки, листья. При редукции листьев развиваются безлистные побеги. Весь побег или его элементы могут видоизменяться.
Почка (gemma) — зачаточный побег, находящийся в состоянии относительного покоя. Она состоит из главной оси с конусом нарастания на верхушке, зачаточных листьев — примордиев, цветков и почек. Почки различаются по строению, функциям, месту положения на побеге, длительности существования, форме, величине, окраске, опушенности, наличию кроющих чешуй и другим признакам. Верхушечная почка образуется на верхушке побега и обеспечивает его рост. Над узлами в пазухах листьев расположены пазушные, или боковые почки. Их развитие обусловливает ветвление, а если верхушечная почка отмирает, то и нарастание побега. Придаточные почки, или почки возобновления, образуются в любых местах стеблей, на листьях, корнях, корневищах, клубнях, соцветиях и служат для вегетативного возобновления. Закрытые почки защищены почечными чешуями, а открытые, или голые, почки — нет. В зависимости от строения и специализации различают: вегетативные почки — зачатки вегетативных побегов, генеративные почки — зачатки соцветий, цветочные почки, или бутоны — зачатки цветков ивегетативно-генеративные почки, состоящие из серии узлов и междоузлий, а вместо конуса нарастания— зачаток цветка или соцветия. Спящие почки замирают на долгое время и трогаются в рост при особых условиях, а зимующие почки остаются в покое лишь на зиму. Выводковые, или опадающие, почки специализируются на вегетативном размножении, например, путем «живорождения». Они могут также видоизменяться в луковички и клубеньки. Характер расположения почек и листьев на стебле аналогичен.
Стебель (caulis) — осевая, обычно надземная, радиально симметричная часть побега, которая ориентирует растение в пространстве, несет все другие надземные органы, является магистралью для передвижения веществ. На стеблях выделяются узлы — места прикрепления листьев и междоузлия — участки стеблей между двумя соседними узлами. Пространство между листом и находящимся над ним междоузлием — пазуха листа. След от места прикрепления опавшего листа — листовой рубец.
Характерными признаками побега являются: неограниченный рост и заложение новых органов благодаря деятельности верхушечной, вставочной и боковых меристем; радиальная симметрия, метамерность, или продольная симметрия — последовательное повторение вдоль оси метамера — узла и нижележащего междоузлия.
Главный побег — ось первого порядка, которая закладывается в зародыше спорофита у высших растений. При развитии проростка последующие метамеры главного побега формируются верхушечной почкой, а из боковых вырастают боковые побеги второго и последующих порядков. Они обусловливают ветвление, увеличение размеров вегетативного тела растения. По степени ветвления различают побеги слабо- и сильноветвистые. Существует несколько типов ветвления и способов нарастания побега. Наиболее примитивное — верхушечное ветвление и дихотомическое, или вильчатое, нарастание: главная ось рано прекращает расти, верхушечная меристема раздваивается, дает пару боковых осей, ветвящихся затем равно- или неравновильчато. Так нарастают водоросли, грибы, лишайники, мхи, плауны. При боковом ветвлении образование ветвей происходит из боковых почек, расположенных ниже точки роста главной оси. Если верхушечная почка сохраняется и обеспечивает постоянный рост главной оси, а боковые оси развиты слабее и не превышают главную ось, то нарастание моноподиальное. Если верхушечная почка прекращает свое развитие, а из ближайших пазушных почек развиваются боковые побеги второго порядка, растущие в направлении главной оси, как бы замещая ее, то это симподиальное нарастание. У травянистых растений ветвление наиболее хорошо выражено в соцветиях. В зависимости от числа замещающих боковых осей и их расположения различают симподиальные побеги типа монохазия, дихазия и плейохазия. У деревьев ветвление ствола явно выражено и влияет на тип и форму кроны. Для злаков, некоторых кустарников и кустарничков, имеющих прикорневую зону ветвления, характерно кущение — развитие боковых побегов у основания главного побега, ведущее к формированию куста или дерновины. Колоновидный, не ветвящийся стебель, характерен для пальм. Эволюция и приспособительная адаптация растений привели к морфолого-физиологическому разнообразию побегов. Помимо вегетативных ассимилирующих побегов, существуют специализированные генеративные спороносные побеги, обеспечивающие размножение (стробил, соцветие, цветок). Кроме надземных, имеются подземные побеги, приспособленные у многолетних травянистых растений к накоплению питательных веществ и вегетативному возобновлению. Побег, развивающийся в течение одного вегетационного периода — годичный, травянистый побег, а в течение нескольких лет — многолетний, одревесневший побег, или ветка. По положению в пространстве и характеру роста среди вертикальных, или ортотропных, побегов различают прямостоячие, изогнутые, цепляющиеся, лазящие, вьющиеся. Горизонтальные, или плагиотропные, побеги могут быть лежачими (плети), ползучими, укореняющимися (усы), стелющимися. Плагиотропно-ортотропные побеги называются приподнимающимися, или восходящими.
В зависимости от развитости междоузлий различают побеги укороченные и удлиненные. Укороченные побеги могут быть: вегетативными — со сближенными листьями, генеративными, или плодушками — с цветками и плодами; розеточными, имеющими редуцированные междоузлия и прикорневую розетку листьев. Удлиненные побеги подразделяются на хорошо олиственные вегетативные, или ростовые, ,и безлистные цветоносные, или цветочную стрелку — сильно удлиненное верхнее междоузлие розеточного побега. Разнообразие побегам придают и такие морфологические признаки, как форма стебля и его очертание в поперечном сечении. Продолжительность существования побега, наличие почек возобновления, размеры, степень одревеснения стебля и другие показатели габитуса учитываются при определении биоморфы, или жизненной формы растения. В соответствии с эколого-морфологической классификацией выделяют три категории жизненных форм — древесную, полудревесную и травянистую. Лист (folium) — боковой, обычно уплощенный, временный орган, растущий ограниченно своим основанием и краем. Симметрия билатеральная: различается верхняя, или внутренняя (брюшная, адаксиальная) сторона, обращенная к стеблю, инижняя, илинаружная(спинная, абаксиальная) сторона, обращенная от стебля. Главные функции листьев — фотосинтез, транспирация и газообмен. Дополнительные функции выполняют видоизмененные листья. При наземном прорастании семян у части двудольных семядоли выносятся над землей, зеленеют и служат первичными листочками. Зачатки настоящих листьев — примордии, закладываются в конусе нарастания почек. В полном листе различают, помимо главной его части — листовой пластинки, также основание, черешок и прилистники. Основание — нижняя часть листа, сочлененная со стеблем, бывает слабо различимым, в виде небольшого утолщения — листовой подушки, разросшимся во влагалище, или иным. Черешок — стеблеобразная часть листа между основанием и пластинкой, выполняющая функции опоры, проведения, регулирования положения листа в пространстве и создания «листовой мозаики». Листья с длинным или коротким черешком называются длинно- или короткочерешковыми, без черешка — сидячими, которые в свою очередь бывают низбегающими, пронзенными, стеблеобъемлющими и полустеблеобъемлющими. Черешки листьев могут иметь каллус — резкое утолщение в верхней или нижней части, могут разрастаться в пленчатое влагалище, охватывающее стебель (злаки, зонтичные). Прилистники — парные боковые листоподобные выросты основания листа. Они могут быть свободными, приросшими к черешку, пазушными, разрастающимися в крупные фотосинтезирующие листочки, редуцированными или опадающими (тогда листья без прилистников). Иногда прилистники срастаются между собой в раструб, охватывающий стебель (гречишные), видоизменяются в пленочки, колючки, волоски и пр. Прилистники, образующие почечные чешуи, выполняют защитную роль. Листовая пластинка приспособлена к выполнению основных функций листа и отличается большим морфологическим разнообразием. Форма пластинки определяется соотношением длины и ширины, сходством с геометрическими фигурами, предметами и др. При описании листьев обычно указывают не только форму самой пластинки, но и ее частей — верхушки, основания, края, а также отмечают тип жилкования. При характеристике края листа, кроме основного названия, часто добавляются приставки-уточнения, например: мелко-, крупно-, неравномерно-, двояко-, трояко-, глубоко-, неясно-, слабо- и др. Довольно часто край листа на своем протяжении неоднороден, например: пильчато-городчатый, зубчато-пильчатый, зубчато-выемчатый и др. Жилкование листа — это характер прохождения жилок (проводящих пучков) в листовой пластинке. Для класса двудольных характерно перистое и пальчатое жилкование, а для класса однодольных — дуговидное и параллельное. Перистое жилкование: вдоль листовой пластинки по центру проходит четко выраженная главная, или центральная, жилка, от которой в обе стороны равномерно отходят боковые жилки. Они доходят до края пластинки (перисто-краевое жилкование), не доходят до края, дуговидно изгибаются и соединяются с вышележащими жилками (перисто-петлевое жилкование) или многократно ветвятся, образуя густую сеть жилок (перисто-сетчатое жилкование). Пальчатое жилкование: от основания листовой пластинки веерообразно отходит несколько одинаковых жилок с боковыми неветвящимися или многократно ветвящимися жилками (пальчато-сетчатое жилкование), доходящими до края (пальчато-краевое жилкование), или петлевидно соединяющимися жилками (пальчато-петлевое жилкование). Дуговидное и параллельное жилкование: жилки равноценны, параллельны краю листа, дуговидные или прямые, если листовая пластинка линейная. Встречаются и сочетания двух типов жилкования, например: пальчато-дуговидное, перисто-дуговидное, пальчато-перистое и др. Различают два типа листьев — простые и сложные. Простые листья имеют одну пластинку. Она может быть цельной или изрезанной. Лист считается цельным, если края пластинки цельные или их изрезанность не превышает 1/3 половины пластинки. Лист считается изрезанным(расчлененным), если изрезанность превышает 1/3 половины пластинки. У цельных листьев, как правило, хорошо различимы и описываются все морфологические признаки: форма пластинки, ее основание, верхушка, край, тип жилкования. У расчлененных листьев очертания верхушки и основания не всегда можно определить, а характеристика края относится уже не к пластинке, а к свободным ее частям — лопастям, долям, сегментам. По форме пластинки, расположению и числу свободных частей различают изрезанные листья перистые, тройчатые и пальчатые. По степени изрезанности и относительным размерам свободных частей эти листья подразделяются на лопастные (свободные части — лопасти; изрезанность составляет более 1/3, но менее 1/2 полупластинки), раздельные, или разделенные(свободные части — доли равны 1/2 полупластинки и более), и рассеченные (изрезанные на сегменты до основания пальчатой пластинки или до рахиса перистой пластинки). При описании изрезанных листьев отмечается форма свободных частей и характер их края (например: доли ланцетные, по краю пильчатые). Изрезанность может быть двойной, тройной или многократной и тогда, при описании листовой пластинки, характеризуется форма, верхушка и край долей и сегментов, или перышек листа, второго, третьего и последующих порядков. Сложные листья состоят из нескольких черешковых или сидячих листочков. Заметно место их сочленения с черешком или рахисом, а после опадания обычно остается листовой рубец. От количества и расположения листочков зависит название сложного листа. У листьев тройчато-сложных и пальчато-сложныхлисточки прикрепляются к верхушке черешка, а у перисто-сложных — к рахису, или стержню, который является продолжением общего черешка. Если рахис разветвляется, листья становятся дважды- или трижды-перисто-сложными. Для сложных листьев характерно то, что при листопаде опадают отдельные листочки сложного листа (видны листовые рубцы), в то время как у рассеченных листьев, похожих внешне на сложные, отделяется от стебля весь лист, а не его сегменты. Для большинства растений характерна гетерофиллия, или разнолистность, заключающаяся в том, что различные по местоположению на стебле листья отличаются друг от друга размерами, формой, степенью изрезанности, развитостью частей и др. В этой связи выделяют такие листовые формации: прикорневые, или розеточные, листья; стеблевые низовые; стеблевые срединные; стеблевые верховые — это прицветники и прицветнички. Характер взаимного расположения листьев на стебле называется листорасположением. Оно аналогично расположению почек на стебле и бывает очередным, супротивным, мутовчатым, розеточным. Тип листа, листорасположение, наличие или отсутствие и развитость его частей — пластинки, черешка, прилистников,— наследственные, исторически закрепленные признаки, которые учитываются при систематизации растений. Существует также большое количество описательных признаков, имеющих значение при видовой характеристике, макроскопическом анализе лекарственных растений и лекарственного растительного сырья. К ним относятся: окраска, размеры, консистенция (листья сухие, мясистые, пленчатые, чешуйчатые), форма свободных частей, наличие и характер опушения листа или его частей, а также другие особенности.
studfiles.net
Вторичное строение корня
Категория: Анатомия растений
Вторичное строение корня
У большинства двудольных растений первичное строение в корнях сохраняется недолго, так как в них рано начинается вторичное утолщение, связанное с заложением и деятельностью камбия. По сравнению со стеблем вторичное утолщение корня имеет некоторые особенности, обусловленные спецификой его первичной структуры.
Тонкостенные живые клетки, находящиеся в центральном цилиндре с внутренней стороны флоэмных тяжей, между лучами первичной ксилемы, вытягиваются в радиальном направлении и делятся тангентальными перегородками, образуя клетки камбия. На поперечном срезе он располагается вогнутыми дугами, концами упирающимися в перицикл. В результате его деятельности возникают вторичные проводящие ткани, поэтому этот камбий условно можно назвать пучковым. При делении клеток пучкового камбия и последующей дифференцировке их производных ковнутри от него, между лучами первичной ксилемы, возникают элементы вторичной ксилемы. Постепенно дуги пучкового камбия становятся выпуклыми. Кнаружи камбий откладывает клетки, превращающиеся в элементы вторичной флоэмы. Число его дуг всегда равно числу лучей первичной ксилемы. Меристематическими вскоре становятся и паренхимные клетки перицикла, соприкасающиеся с группами элементов протоксилемы. В результате их деления образуются клетки межпучков о(по положению) камбия. Участки пучкового и межпучкового камбия смыкаются в непрерывный камбиальный слой.
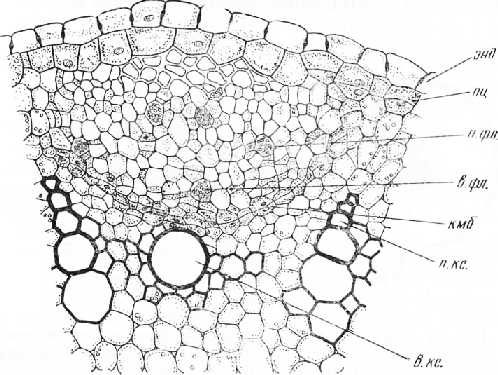
Рис. 1. Заложение и начало деятельности камбия в корне проростка тыквы: энд — эндодерма, пи, — перицикл, л. фл. — первичная флоэма, в. фл. — вторичная флоэма, кмб— камбий, я. КС. — первичная ксилема, в. кс.— вторичная ксилема
Таким образом, в центральном цилиндре корня формируются открытые коллатеральные проводящие пучки в числе, равном числу лучей первичной ксилемы. Первичная флоэма оттесняется вторичными тканями к периферии пучков, сдавливается и становится плохо заметной.
Клетки межпучкового камбия, заложившиеся в пе-рицикле, образуют широкие лучи паренхимы, располагающиеся между пучками вторичных проводящих тканей и имеющие вид светлых полос. Так как эти лучи возникают в самом начале вторичного утолщения, их можно называть первичными. На этой стадии развития корня они обеспечивают связь центральной части корня с первичной корой, поэтому по аналогии со стеблем их нередко называют сердцевинными лучами, хотя в корне нет типичной сердцевины. Кроме первичных могут быть и типичные вторичные лучи, образованные лучевыми инициалями пучкового камбия и входящие в состав вторичных ксилемы и флоэмы.
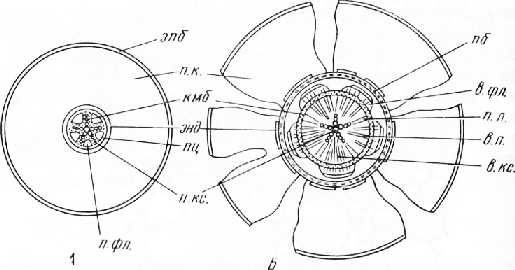
Рис. 2. Схема вторичного утолщения корня. А — первичное строение и заложение камбия; Б — образование вторичных
Вследствие быстрого нарастания изнутри вторичных тканей, обусловливающего сильное утолщение корня, первичная кора нередко разрывается. К этому времени клетки перицикла, энергично делясь по всей окружности центрального цилиндра, образуют широкую зону паренхимных клеток, во внешней части которой закладывается феллоген, откладывающий наружу пробку, а внутрь— многослойную феллодерму. Пробка изолирует первичную кору от проводящих тканей, кора отмирает и сбрасывается. Тонкостенные клетки феллодермы и остатки перицикла в дальнейшем разрастаются и составляют крупноклеточную паренхим-ную зону, окружающую с периферии проводящие ткани.
Корень тыквы (Cucurbita реро L.)
Первичное строение и начало вторичного утолщения
Для изучения первичного строения и последующих вторичных изменений необходимо иметь корни проростков.
Семена тыквы проращивают в течение одной-двух недель в чашках Петри. В связи с тем, что свежие тонкие корни проростков режутся плохо, их следует предварительно фиксировать спиртом с добавлением глицерина.
Поперечные и продольные срезы обрабатывают фло-роглюцином с соляной кислотой и рассматривают в глицерине.
На серии поперечных срезов корня проростка в зоне проведения на расстоянии 1,5—2 см от кончика можно ознакомиться с первичным строением, а также проследить заложение и начало деятельности камбия. Под экзодермой видна широкая паренхимная первичная кора, заканчивающаяся слоем тонкостенных клеток эндодермы с поясками Каспари, которые перерезаны поперек и на радиальных стенках имеют вид небольших вздутий (пятна Каспари). После проведения реакции с флоро-глюцином и соляной кислотой они приобретают красный цвет. Под эндодермой расположен однорядный перицикл. Первичная ксилема обычно четырех-, реже трех-и пятилучевая. Середина корня занята участком тонкостенной ткани, клетки которой на продольных срезах имеют удлиненную форму, типичную для прокамбия.
На более поздних стадиях развития, т. е. ближе к основанию корня, в центре его формируется крупный сосуд метаксилемы. Группы элементов первичной флоэмы отделены от первичной ксилемы узким слоем живых тонкостенных клеток, сохраняющих способность к делению. При большом увеличении видно, что они несколько вытянуты в радиальном направлении и плотно примыкают одна к другой. Клетки заполнены густой цитоплазмой, имеют крупные ядра. В некоторых местах эти клетки делятся тангентальными перегородками, вычленяя узкие клетки пучкового камбия. На срезах, сделанных с более старого участка корня, можно видеть, что плоские, вытянутые в тангентальном направлении расположены вогнутыми дугами. О начале деятельности камбия можно судить по появлению между двумя соседними лучами первичной ксилемы одного-двух сосудов вторичной ксилемы, которые более широкопро-светны, чем сосуды метаксилемы.
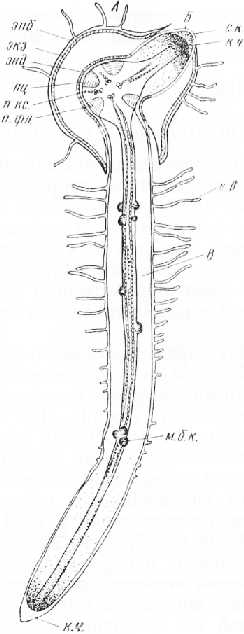
Рис. 3. Заложение боковых корней у тыквы. А — материнский корень с двумя боковыми корнями второго порядка (Б, В)\ Б — сформировавшийся боковой корень, пробившийся сквозь первичную кору; В — функционирующий боковой корень второго порядка с меристематическими зачатками боковых корней (м. б. к.) третьего порядка: эпб — эпиблема, экз — экзодерма, энд — эндодерма, пц — перицикл, п. кс. — первичная ксилема, п. фл. — первичная флоэма, с. к. — секреторный кармашек, к. ч. — корневой чехлик, к. е. — корневые волоски таблитчатые клетки камбия
На этих же срезах можно видеть, что клетки перицикла, находящиеся над тяжами первичной ксилемы, также разделились тангенталь-ными перегородками. Это начало развития межпучкового камбия, который в дальнейшем образует па-ренхимные клетки лучей.
На поперечных срезах корней проростка в зоне поглощения или в начале зоны проведения можно проследить также заложение и последовательность развития боковых корней. Боковой корень возникает эндогенно из группы клеток перицикла против лучей первичной ксилемы. Апикальная меристема материнского корня в образовании боковых корней не участвует. Путем деления клеток перицикла продольными и поперечными перегородками формируется бугорок мери-стематической ткани («кор-неродная дуга»), слегка вдающийся в первичную кору. Затем деление распространяется на клетки эндо-
дермы, окружающие бугорок снаружи. Они делятся преимущественно радиальными перегородками, образуя на поверхности меристематического зачатка корня секреторный кармашек, который выделяет ферменты, растворяющие клетки первичной коры, и таким образом облегчает рост бокового корня в ее толще. В апексе корневого зачатка дифференцируются те же слои инициалей, как в апексе материнского корня. Зона интенсивно делящихся клеток прикрыта корневым чехли-ком. В центральной части заложившегося бокового корня, близ его основания, формируется прокамбий, дающий начало проводящим тканям. Трахеальные элементы ксилемы бокового корня непосредственно присоединяются к сосудам тяжа первичной ксилемы материнского корня, против которого образовался зачаток бокового корня, а элементы флоэмы примыкают к двум соседним участкам первичной флоэмы. Так устанавливается связь проводящего аппарата бокового корня с центральным цилиндром материнского корня. Сформированный боковой корень, нарастая в длину своей верхушкой, пробивается наружу.
Заложение боковых корней против тяжей первичной ксилемы определяет их расположение на материнском корне продольными рядами.
Задание. 1. Зарисовать схему первичного строения корня. Показать заложение камбиальной дуги между участком первичной флоэмы и двумя соседними лучами первичной ксилемы. 2. При большом увеличении микроскопа изобразить часть центрального цилиндра с клетками пучкового камбия и первыми сосудами вторичной ксилемы.
Вторичное строение
С особенностями вторичного строения взрослого корня удобнее ознакомиться на поперечных срезах корней толщиной 2,5—5 мм.
При изучении препарата в центре среза следует прежде всего отыскать первичную ксилему. Она представлена четырьмя, изредка тремя или пятью короткими радиальными цепочками узкопросветных мелких сосудов, сходящихся к одному более широкому сосуду мета-ксилемы.
Между лучами первичной ксилемы находятся четыре или три — пять крупных открытых коллатеральных проводящих пучков камбиального происхождения. Большую часть сечения каждого пучка занимает вторичная ксилема, состоящая из широкопросветных сосудов, волокон и мелких клеток паренхимы.
На срезах из материала, собранного в конце вегетационного периода, внутри крупных сосудов можно обнаружить пузыревидные образования, частично или полностью заполняющие их просвет. Это тиллы, возникающие в результате врастания паренхимных клеток в расположенный рядом с ними сосуд. Оболочки их иногда тонкие, иногда утолщенные, одревесневшие, с многочисленными порами. Заполненный тиллами сосуд утрачивает проводящую функцию.
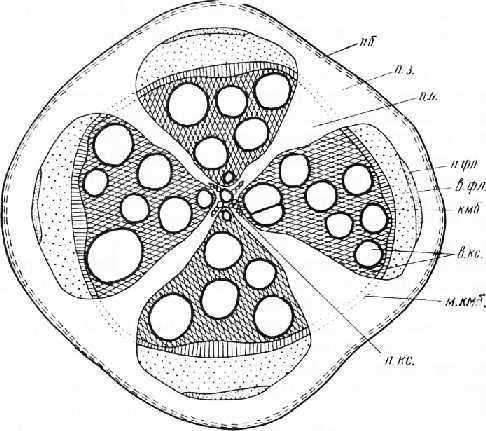
Рис. 4. Поперечный срез вторично утолщенного корня тыквы: лб — пробка, л. з. — паренхимная зона, п. л. — первичный луч, л. фл. — первичная флоэма, в. фл. — вторичная флоэма, кмб — камбий, в. кс. — вторичная ксилема, л. кс. — первичная ксилема, м. кмб. — межпучковый камбий
Снаружи элементы вторичной ксилемы огибает камбиальная зона, имеющая слегка волнистые очертания. Она представляет собой широкий слой прозрачных мелких узких таблитчатых клеток, расположенных правильными радиальными рядами. К периферии от камбия в пучке находится вторичная флоэма, состоящая из широ-копросветных ситовидных трубок с простыми горизонтальными ситовидными пластинками, сопровождающих клеток и паренхимы. Первичная флоэма оттеснена к периферии пучка, деформирована и вследствие ранней облитерации (сдавливания) ситовидных трубок плохо различима.
От элементов протоксилемы, несколько изгибаясь между проводящими пучками, расходятся светлые, расширяющиеся к периферии первичные лучи. Они состоят из крупных удлиненных в радиальном направлении тонкостенных клеток, образованных межпучковым камбием. Межпучковый камбий у корней со вторичным строением не всегда заметен.
С поверхности корень покрыт перидермой. На тонком участке среза старого корня в перидерме можно различить несколько наружных слоев сплюснутых пустых клеток пробки с бурыми стенками. Под пробкой находится слой узких клеток феллогена, заполненных мелкозернистой цитоплазмой, и феллодерма. Крупные клетки внутренних слоев феллодермы вместе с производными перицикла, возникшими еще до начала деятельности пробкового камбия, составляют паренхимную зону, окружающую снаружи проводящие пучки и лучи.
Корень вторичного строения всегда можно отличить от стебля по следующим признакам: 1) первичная ксилема экзархная, ее элементы располагаются в центре корня радиальными тяжами, типичной сердцевины нет; 2) промежутки между тяжами первичной ксилемы заняты вторичной ксилемой; 3) первичные лучи внутренними концами упираются в проводящие элементы первичной ксилемы; 4) первичной коры в корнях двудольных растений, имеющих вторичное утолщение, обычно нет.
У тыквы нередко встречаются корни не с тремя — пятью проводящими пучками, а с большим числом. Центральную часть таких корней составляют тонкостенные клетки. По строению эти корни кажутся сходными со стеблем, однако у основания первичных лучей между пучками вторичных проводящих тканей при внимательном рассмотрении всегда можно обнаружить участки экзархной первичной ксилемы, которая характерна только для корня (в стебле первичная ксилема эндархная).
Задание. При малом увеличении зарисовать схему строения корня, отметив: а) первичную и вторичную ксилему; б) пучковый камбий; в) вторичную и облитери-рованную первичную флоэму; г) лучи; д) периферическую паренхимную зону, окружающую проводящие пучки; е) пробку.
Вторичное строение можно изучать также на корнях русских бобов (Faba vulgaris Moench ,) или фасоли (Pha-seolus vulgaris L.). Корни этих растений отличаются от корней тыквы присутствием в первичной флоэме, непосредственно под перициклом, механических элементов — волокон — и небольшого участка тонкостенных клеток в центре.
Корень лютика ползучего (Ranunculus repens L.)
Активная деятельность камбия, вследствие которой в корне образуется большое количество вторичных тканей, наблюдается не у всех травянистых двудольных растений. Очень слабое вторичное утолщение корней харак терно для многих травянистых представителей семейства лютиковых, в том числе лютика ползучего.
На поперечных срезах корня, обработанных раство ром иода в водном растворе иодистого калия, а затем флороглюцином и соляной кислотой, можно видеть широкую первичную кору и узкий центральный цилиндр (рис. 74).
В центре корня четырьмя-пятью радиальными тяжами расположены элементы первичной ксилемы, чередующиеся с группами клеток флоэмы (рис. 74, А). С внутренней стороны каждый участок флоэмы огибают доходящие до перицикла дуги камбия, образовавшие небольшое число элементов вторичных проводящих тканей.
Вторичная флоэма содержит ситовидные трубки с простыми ситовидными пластинками и сопровождающие клетки, вторичная ксилема — пористые сосуды и паренхиму.
Перицикл представлен одним рядом крупных тонкостенных паренхимных клеток. Делясь тангентальными перегородками, клетки перицикла, расположенные против элементов протоксилемы, образуют клетки межпучкового камбия, дающего начало коротким и широким паренхимным лучам.
Центральный цилиндр окружен однослойной эндодермой из слегка вытянутых в тангентальном направлении клеток с утолщенными одревесневшими вторичными оболочками. Против лучей первичной ксилемы в эндодерме, находятся пропускные клетки с пятнами Каспари. Так как вторичное утолщение незначительно, первичная кора сохраняется без заметных изменений. Наружные и внутренние слои ее состоят из довольно плотно сомкнутых клеток с утолщенными целлюлозными оболочками. В этих клетках можно видеть крупные ядра и зерна запасного крахмала. В некоторых клетках встречаются кристаллы оксалата кальция. В средней части коры расположены крупные воздухоносные полости рексигенного происхождения, т. е. возникшие вследствие разрывов клеток. Наличие полостей объясняется повышенной влажностью естественного местообитания лютика.
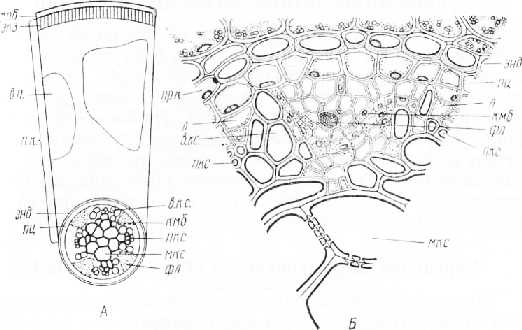
Рис. 5. Строение корня лютика ползучего. А — схема строения; Б — сектор центрального цилиндра корня лютика при большом увеличении (поперечный срез): эпб — эпиблема, экз — экзодерма, в. п. — воздухоносная полость, п. к. — первичная кора, энд — эндодерма, пц — перицикл, в. кс. — вторичная ксилема, кмб — камбий, пкс — протоксилема, мкс — метаксилема, фл — флоэма, пр. к. — пропускная клетка, л — луч паренхимы
Перидермы нет. Функцию защитной периферической ткани выполняют однослойная экзодерма из сравнительно тонкостенных клеток и остатки отмершей эпиблемы.
Задание. При малом увеличении микроскопа зарисовать схему строения корня, отметив первичные флоэму и ксилему, участки вторичной ксилемы, короткие парен-химные лучи, перицикл, эндодерму, широкую первичную кору с воздухоносными полостями, экзодерму.
Анатомия растений - Вторичное строение корняgardenweb.ru
| АрхеологияАрхитектураАстрономияАудитБиологияБотаникаБухгалтерский учётВойное делоГенетикаГеографияГеологияДизайнИскусствоИсторияКиноКулинарияКультураЛитератураМатематикаМедицинаМеталлургияМифологияМузыкаПсихологияРелигияСпортСтроительствоТехникаТранспортТуризмУсадьбаФизикаФотографияХимияЭкологияЭлектричествоЭлектроникаЭнергетика | Метаморфозы корня: корни ходульные, дыхательные, контрактильные, воздушные, опорные. Микориза, клубеньки на корнях бобовых, их значение в природе и в сельском хозяйстве. Анатомическое строение корня. Первичное строение корня характеризуется наличием трех систем тканей — покровно-всасыпающей эпидермы с корневыми волосками (эпиблемы), первичной коры и центрального цилиндра Эпиблема Выполняет всасывающую и защитную функции, состоит из одного слоя тонкостенных, плотно сомкнутых клеток с корневыми волосками, не имеет устьиц, толстой кутикулы и трихом. Первичная кора, лежащая под элиблемой, подразделяется на экзодерму, мезодерму и эндодерму. Экзодерму составляет 3—4 слоя крупных, плотно сомкнутых клеток с частично опробковевшими оболочками. Выполняет защитную и опорно-пропускную функции. Мезодерма — многослойная запасающая часть, составляет основную массу первичной коры. Мезодерма выполняет также транспортную и воздухоносную функции. Эндодерма — самый внутренний, обычно однорядный слой первичной коры, граничащий с центральным цилиндром, выполняющий опорно-пропускную функцию. Центральный, Или Осевой, цилиндр Занимает срединную часть корня, окружен кольцом перицикла, из которого формируются боковые корни, В корне нет настоящей сердцевины. В центре могут находиться сосуды, склеренхима, иногда — запасающая паренхима, образующая Ложную сердцевину Вторичное строение корня формируется благодаря появлению и функционированию вторичных боковых меристем — камбия и феллогена. Камбий образуется из перицикла над лучами первичной ксилемы и из прокамбия или паренхимы под флоэмой. К наружи деляшийся камбий откладывает вторичную флоэму, а к центру — вторичную ксилему. Первичная ксилема оттесняется к оси корня, а первичная флоэма — к периферии центрального цилиндра. Вторичное строение корня может быть пучкового типа или беспучкового Пучковый тип формируется в том случае, если межпучковый камбий, образовавшийся над лучами первичной ксилемы, продуцирует паренхиму сердцевинных лучей, а пучковый камбий, лежащий под флоэмой,— вторичную флоэму и ксилему коллатерального или биколлатерального проводящего пучка. При Беспучковом типе Строения кольцо камбия образует сплошные кольца ксилемы и флоэмы. Вторичные преобразования в первичной коре корня связаны с деятельностью феллогена. Если он возникает из перицикла, то пробка, образующаяся кнаружи, изолирует первичную кору, и она отторгается, слущивается, если феллоген образуется из паренхимы мезодермы, что встречается редко, то часть первичной коры, лежащая под феллогеном, сохраняется. У древесных растений корень обычно беспучкового типа и в древесине формируются кольца годичного прироста. Годичное Кольцо — это совокупность тканей, образованных камбием за один вегетационный период. Покровной тканью корней древесных может быть перидерма или корка. Наличие в центре органа радиального проводящего пучка, склеренхимы или сосудов первичной ксилемы отличает корень от стебля и корневища, у которых центральную часть занимает настоящая сердцевина.. 3. Метаморфозы корня. В процессе эволюции корни отдельных растений приобретали дополнительные функции. Одни из них стали резко отличаться по внешнему виду от типичных корней - это и есть метаморфозы корней. Другие сохранили типичное строение. Это корни с особыми функциями. К метаморфозам корней относятся мясистые корни - корнеплоды и корнеклубни. Корнеклубни - это видоизменения придаточных корней. Корнеклубни на вершине несут придаточные почки. Корнеклубни образуются у георгин, чистяка, земляных орешков, бататов. Корнеплоды известны у моркови, петрушки, редьки, свеклы и др. Но следует помнить, что «корнеплод» у них в морфологическом смысле представлен укороченным стеблем - частью несущей листья, подсемядольным коленом и, наконец, собственно корнеплодом - видоизмененным мясистым корнем. Ходульные корни, тип корней, растущих от стебля растения и дающих растению дополнительную поддержку. Эти корни продолжают расти по мере того, как тонкий стебель растения становится выше, и часто являются необходимыми для того, чтобы растение не упало. Эти мощные надземные корни часто поддерживают ствол деревьев над уровнем воды и укрепляют растения в иле или в песчаном грунте приливной полосы морских побережий. Встречаются у некоторых травянистых растений, например, у маиса, и у деревьев, таких как мангровые деревья. Кроме того, многим тропическим растениям присущ другой тип поддерживающих корней - плоские, расположенные на земле, корни, которые растут от основания ствола и помогают держаться деревьям с неглубокими корнями. Воздушные корни водяных и болотных растений находят в окружающей среде мало кислорода или не находят его вовсе. Так как ткани высших растений неспособны к "нормальному" анаэробиозу, т. е. неспособны нормально жить без доступа свободного кислорода, то понятно, что этот последний должен быть доставлен корням растения тем или иным путем. По большей части достигается это при посредстве сильноразвитой воздухоносной системы (см.) с громадными межклетниками, проводящими кислород от листьев, заимствующих его в свой черед из атмосферы или накопляющих внутри своих тканей в процессах ассимиляции. |
studopedya.ru
Формирование вторичного строения корня
В корнях древесных растений в процессе роста и развития корневого окончания между его ксилемой и флоэмой из ткани перицикла формируется камбий.
Сначала он представляет разобщенные между собой прослойки, затем в результате тангенциального деления клеток перицикла они соединяются между собой. Образуются вторичные (боковые) меристемы — камбий и феллоген. В начальные периоды своего образования положение камбия напоминает звездчатообразный характер, которое довольно быстро принимает цилиндрическую форму. В результате деления клеток камбия внутрь от него кольцеобразно откладываются ткани ксилемы, а в наружную сторону — клетки флоэмы.
Формирование камбия между участками флоэмы и ксилемы является первым признаком перехода корня от первичного строения ко вторичному. Вторым признаком вторичного строения является образование феллогена из всего кольца клеток перицикла в результате их тангенциального деления. Клетки феллогена делятся также тангенциально и отлагают наружу клетки пробки, а внутрь — несколько рядов живых клеток феллодермы.
Феллоген образуется еще в перицикле, т. е. под слоем первичной коры. В результате его деятельности первичная кори постепенно изолируется от внутренних живых тканей корня, отмирает и отлущивается. Зона отлущивания первичной коры является зоной перехода корня от первичного строения к вторичному. Здесь корень, сбросив первичную кору, утончается. Характер анатомического строения его становится стабильным. Сбрасывание первичной коры — это третий характерный признак перехода от первичного строения корня к вторичному. Поверхность корня к этому периоду покрыта опробковевшим слоем клеток (коркой), в результате чего в этой зоне корневое окончание теряет функцию всасывания и в качестве основной приобретает функцию проведения растворов питательных веществ. Этот процесс смены функций происходит постепенно, поскольку в начальном его периоде среди опробковевших клеток покрова имеется более или менее значительное количество живых неопробковевших клеток, через которые почвенный раствор и вода продолжают поступать внутрь корня. Эту зону корня в отличие от сосущей зоны корневых окончаний и последующих более старых зон, полностью утративших функцию всасывания, называют полупроводящими корнями. Последующие более старые части корня называются проводящими корнями.
Таким образом, по функциональным признакам в корневой системе формируются всасывающие полупроводящие и проводящие зоны корня. Корни древесных пород обладают способностью к вторичному утолщению и в процессе роста увеличиваются в диаметре. Поэтому параллельно указанной классификации корней по функциональным признакам применяют классификацию по морфологическим признакам, выделяя корни тонкие, полускелетные и скелетные. Обычно к тонким корням относят корни и части диаметром менее 2 мм, к полускелетным — 2—3 мм, к скелетным — участки корней диаметром более 3 мм.
Рост корневого окончания и его развитие (в физиологическом аспекте) — это одновременный процесс. Собственно рост является интегральным результатом деления клеток первичной меристемы, приобретения ими тургора и процесса растягивания. Это явление происходит в небольшой по величине зоне, в вершине которой — группа инициальных клеток. Согласно законам гидравлики возникающее здесь давление распределяется равномерно во все стороны, т. е. наружу (по радиусу) и в оба конца по оси корня. Площадь, на которую распределяется радиальное давление, во много раз больше площади сечения оси корня, следовательно, на единицу боковой поверхности корня приходится значительно меньшее давление, чем на единицу площади его сечения. К тому же строение оболочек клеток обеспечивает скольжение составляющих их молекул по оси клеток, что обеспечивает эффект растяжения стенок клеток вдоль ее оси. Все это создает значительное давление, направленное вдоль оси корня от зоны растяжения, где с одной стороны находится неподвижная, скрепленная с почвой базальная часть корня, а с другой — апикальная меристема, прикрытая корневым чехликом. Апикальная меристема и корневой чехлик вдавливаются между частицами почвы и раздвигают их, продвигаясь вперед. В результате создается своего рода движущаяся в пространстве биологическая система, перемещение частей которой по законам физики направлено в сторону меньшего сопротивления, т. е. в сторону инициальных клеток. Группа инициальных клеток под прикрытием корневого чехлика и под влиянием давления постоянно продвигается в новые зоны почвы в направлении от ранее сформировавшегося и укрепившегося в почве участка корня. Таким образом, происходящий линейный рост корня сопровождается выносом группы инициальных клеток, сопутствующей им апикальной меристемы и всего корневого окончания в новые слои и зоны почвы. Энергия роста корня и побегов настолько велика, что пробивает своими растущими окончаниями асфальтовые покрытия и обеспечивает проникновение живых корней в плотные слои почвы, трудно поддающиеся разрушению лопатой, ломом, кайлой.
Процесс одновременного роста корневого окончания и изменения его анатомического строения можно представить рядом последовательных, постепенно переходящих одна в другую фаз, между которыми невозможно установить четкую границу. В процессе роста корня его зоны как бы продвигаются в глубь почвы вслед за первичной меристемой и корневым чехликом. На самом деле, постоянно обновляясь, перемещаются только клетки апикальной меристемы и корневой чехлик. Перемещение остальных зон по отношению к почвенному пространству носит элемент иллюзии, это эффект процесса их появления и трансформации в последующую зону.
Наличие камбия у корней древесных растений обусловливает их вторичное утолщение. Деятельность камбия корней аналогично камбию надземной части дерева имеет периодический характер соответственно временам года. В зимний период деятельность камбия прекращается, что приводит к отложению годичных колец корней. Особенно хорошо видны годичные кольца у хвойных пород. У лиственных они также хорошо различимы при соответствующей обработке срезов корней.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Биология для студентов - 21. Первичное анатомическое строение корней
Рассмотрим поперечный срез корешка, сделанный в зоне всасывания.
Такое первичное строение корня характерно большинству семенных растений.
Снаружи корня расположена ризодерма - всасывающая ткань, через которую происходит поглощение воды и минеральных веществ, а также взаимодействие с живым населением почвы. В ней различают два типа клеток: трихобласты, образующие корневые волоски, и атрихобласты.
Под ризодермой находится первичная кора, выполняющая защитную, проводящую, запасающую и другие функции. Наружный слой первичной коры после отмирания корневых волосков и сбрасывания ризодермы дифференцируется в экзодерму, а из внутреннего слоя развивается эндодерма.
Эндодерма в своем развитии может пройти три ступени. На первой ее клетки расположены плотно, без межклетников, и характеризуются наличием на радиальных и поперечных стенках утолщений в виде рамочек, опоясывающих клетки (поясков Каспари), в которых откладываются гидрофобное вещество, сходное с суберином, и лигнин. Пояски соседних клеток смыкаются - и создается их непрерывная цепь вокруг стелы. Пояски Каспари непроницаемы для растворов; следовательно, вещества из коры в стелу и обратно могут пройти только по симпласту, т.е. через живые протопласты клеток эндодермы.
Таким образом, функция эндодермы - контроль за проведением веществ. У многих растений эндодерма получает вторичное и третичное развитие. На второй ступени развития субериноподобные вещества откладываются по всей внутренней поверхности стенок эндодермы. Однако при этом эндодерма не становится абсолютно непроницаемой для растворов, поскольку в ней имеются пропускные клетки, сохранившие первичное строение.
Третья ступень характеризуется сильным утолщением и одревеснением боковых и внутренних стенок, и отмиранием протопластов. Пропускные клетки сохраняются и осуществляют физиологическую связь между первичной корой и центральным цилиндром.
В центре корня располагается центральный цилиндр (стела). Он имеет следующее строение. Сразу под эндодермой находится перицикл - ткань, образующая боковые корни. Клетки прокамбия формируют первичные проводящие элементы. Флоэма начинает развиваться раньше ксилемы. Клетки протофлоэмы примыкают к перициклу. Метафлоэма расположена ближе к центру.
Ксилема формируется позже. Протоксилема возникает вплотную к перициклу, метаксилема образуется ближе к центру корня, т. е. ксилема и флоэма в корне закладываются экзархно и развиваются центростремительно. На поперечном срезе ксилема выглядит, как звезда, число лучей которой - от одной (папоротники) до тридцати. Радиальные пучки, имеющие по одному тяжу ксилемы и флоэмы, называют:
- моноархными,
- по два - диархными,
- по три - триархными,
- по нескольку - полиархными.
Между этими лучами располагаются участки флоэмы. Такой тип проводящей системы облегчает поступление растворов в сосуды, т.к. лучи ксилемы подходят к периферии стелы.
vseobiology.ru
Анатомия корня однодольных и двудольных растений
1.Общая характеристика корня. Корневые системы.
2. Зоны корня.
3. Первичное анатомическое строение корня.
4. Вторичное анатомическое строение корня.
5. Метаморфозы корней.
Корень – осевой орган, име6ющий более или менее цилиндрическую форму и обладающий радиальной симметрией. Он способен к открытому росту до тез пор, пока сохраняется апикальная меристема. Морфологически корень отличается от побега тем, что на нём никогда не возникают листья, а апикальная меристема покрыта корневым чехликом.
Общая характеристика корня.
- главный корень, он всегда развивается из зародышевого корешка
- боковые корни, они образуются на главном корне или боковых корнях более высокого порядка из перецикла (боковая меристема)
- придаточные корни, они развиваются всегда на стеблях или листьях, но не на корнях. Воздушные корни и корни-подпорки также являются придаточными корнями. Придаточные корни могут развиваться при вегетативном размножении.
Типы корневых систем:
1. корневая система главного корня (стержневая корневая система). В этом случае главный корень всегда присутствует, и он имеет наибольшие размеры.
2. придаточная корневая система (мочковатая корневая система злаков). Главный корень, как правило, быстро отмирает, и начинают активно развиваться придаточные корни. Придаточная корневая система развивается у корневищных растений (ландыш, купена, вороний глаз, клевер).
Другой вариант классификации корневых систем:
а) стержневая корневая система и её разновидность – ветвистая. Все боковые корни семенных растений закладываются эндогенно, т. е. внутри корня предшествующего порядка, и развиваются чаще всего из перицикла. Такую систему имеют двудольные и голосеменные растения.
б) мочковатая корневая система. Этот тип характерен для однодольных однолетних растений. У многолетних растений (как двудольных, так и однодольных) часто формируется корневище, от которого отходят придаточные корни. В силу этого сам характер корневой системы может существенно изменяться.
Зоны корня:
1. Зона деления (1мм), или меристематическая зона. Она состоит из небольшого числа клеток апикальной меристемы.
Зона деления отличается жёлтой окраской. Она представлена апексом корня, состоящим из мелких многогранных клеток-инициалей и их производных, эти клетки отличаются густой цитоплазмой и крупными ядрами.
Снаружи зона деления прикрыта корневым чехликом, который выполняет несколько функций.
а) защищает апикальную меристему при продвижении корня через почву
б) его клетки в процессе жизнедеятельности выд6ляют слизь, которая облегчает продвижение корня
в) эти клетки представляют собой особую сенсорную зону, способную восприни-
мать действие сил гравитации.
Клетки чехлика живые, но поверхностные клетки периодически слущиваются и заменяются новыми.
Корневой чехлик состоит из живых паренхимных клеток, возникающих у большинства однодольных растений из особой меристемы (калиптрогена), а у двудольных и голосеменных растений – из верхушечной меристемы кончика корня. Водные растения обычно лишены корневого чехлика
2. Зона растяжения, или роста (2-3мм) – здесь клетки растягиваются, т. е. резко увеличиваются в размерах за счёт поступления воды и образования крупной центральной вакуоли. Наряду с процессами растяжения происходит и дифференцировка клеток. На уровне этой зоны будет формироваться проводящая ткань.
Зона роста выделяется светлой окраской и как бы прозрачна. Клетки этой зоны отличаются высоким тургором, что способствует раздвиганию частиц почвы.
3. Зона поглощения, или зона корневых волосков (1-2см) – все клетки полностью дифференцированы и закончили свой рост. Внутреннее строение этой зоны первичное, а снаружи она покрыта ризодермой. В состав ризодермы входят трихобласты и атрихобласты. Все клетки ризодермы активно поглощают воду и минеральные соли.
Корневые волоски появляются в виде небольших сосочков – выростов клеток эпиблемы. Рост волоска осуществляется у его верхушки, оболочка растягивается быстро. Продолжительность жизни корневых волосков 15– 20 дней.
4. Зона проведения – наиболее протяжённая зона, она начинается в месте исчезновения корневых волосков. У молодых корней в нижней части она покрыта экзодермой, а более старая часть покрыта перидермой (у древесных растений старые части корня могут быть покрыты пробкой). У большинства растений эта зона имеет вторичное анатомическое строение. Основная функция – вертикальное проведение в побег поглощённого из почвы водного раствора.
Первичное строение корня
У двудольных растений первичное строение корня отмечается на уровне ризодермы, здесь все ткани уже дифференцированы. В ниже расположенной зоне растяжения не все клетки достигли окончательной дифференцировки. В первичном строении корня отмечают три анатомо-топографических зоны: покровы, первичная кора, стель. Для первичного строения корня характерно хорошее развитие первичной коры. В центре стели располагаются прочные элементы: либо первичная ксилема, либо склеренхима.
Покровы представлены ризодермой, в состав которой входят трихобласты с корневыми волосками, а также атрихобласты.
Под покровами расположена первичная кора, в составе которой выделяют 3ткани:
- экзодерма – 1-2 слоя паренхимных плотно расположенных клеток. В зоне проведения, после того как клетки ризодермы отомрут и слущатся, зкзодерма будет выполнять функцию покровов.
- мезодерма – несколько слоёв паренхимных клеток, обеспечивающих радиальный транспорт воды и минеральных веществ и накопление запасных питательных веществ. Мезодерма (а также экзодерма) обеспечивают радиальный транспорт водного раствора от ризодермы к центральному цилиндру.
- эндодерма – внутренний слой коры корня, граничащий с центральным цилиндром. Она представлена одним слоем клеток, большая часть которых имеет пояски Каспари. Пояски Каспари – это формирующаяся вторичная клеточная стенка. Она пропитывается суберином, сначала суберин расположен пятнами, а потом пятна увеличиваются в размерах и формируют пояски. Клетки с поясками Каспари не способны проводить воду из мезодермы в центральный цилиндр, а их протопласт отмирает. Среди толстостенных отмирающих клеток эндодермы встречаются живые клетки с тонкими неодревесневшими стенками. Это пропускные клетки: через них осуществляется связь между первичной корой и осевым цилиндром.
В центральном цилиндре самая наружная ткань представлена перециклом (он даёт начало боковым корням). Проводящий пучок в стели корня имеет радиальное строение. В самом центре корня расположена первичная ксилема в форме 4-х лучевой звезды, а между лучами ксилемы находятся участки первичной флоэмы. У однодольных растений в самом центре проводящего пучка располагается склеренхима, а у двудольных – протоксилема. Такой тип строения, где водопроводящие и механические элементы располагаются в самом центре, обеспечивает достаточную прочность корня на разрыв. В состав центрального цилиндра также входят клетки основной паренхимы.
Особенности вторичного строения корня
У однодольных растений и папоротников первичное строение корня сохраняется на протяжении всей жизни. У двудольных растений и голосеменных достаточно быстро закладывается камбий и формируется вторичное строение корня. Его легко наблюдать на уровне зоны проведения. Особенностью развития камбия в корне является то, что он закладывается в виде лопастного кольца между участками первичной ксилемы и флоэмы. Частично он образуется из перецикла, а частично из клеток основной паренхимы. Деление клеток камбия происходит неравномерно и в результате лопастное кольцо выравнивается и принимает более или менее правильную фору.
У голосемянных и двудольных растений формируется вторичная структура корня, при которой радиальное расположение проводящих тканей заменяется на коллатеральное. Из паренхимных клеток формируется слой камбия. Его клетки интенсивно делятся и дифференцируются во вторичную флоэму (к периферии) и во вторичную ксилему (к центру). В итоге первичная флоэма оттесняется кнаружи, а «звезда» первичной ксилемы остаётся в центре корня.
При вторичном строении корня большой объём его занимает стель, который развивается за счёт деятельности камбия. Кнаружи от стели располагаются остатки первичной коры. Они представлены лишь несколькими рядами клеток.
Помимо вторичных изменений, происходящих в центральном цилиндре, существенные перемены происходят и в первичной коре. Из-за роста вторичных тканей корень утолщается и первичная кора разрывается. К этому времени клетки перицикла образуют феллоген. Феллоген откладывает наружу пробку, а внутрь феллодерму. Пробка изолирует первичную кору от проводящих тканей, кора отмирает и сбрасывается. Клетки феллодермы в дальнейшем разрастаются и окружают проводящие ткани. Иногда эту зону называют вторичной корой. Снаружи корни двудольных растений покрыты перидермой. Корка образуется редко, лишь на старых корнях деревьев.
Основные функции корня:
1. поглощение из почвы воды и минеральных веществ,
2. заякоривание растений,
3. биосинтетическая. В апикальной меристеме корня синтезируются цитокинины, которые оказывают значительное воздействие на рост и развитие корня. Они абсолютно необходимы для митотического цикла, и также для формирования и развития фотосинтетического аппарата. В корнях ещё синтезируются гиббереллины и абсцизовая кислота.
4. выделение веществ, стимулирующих развитие ризобиумных бактерий
5. запасание основными тканями корня питательных веществ,
6. вегетативное размножение (корневые отпрыски).
Метаморфозы корня:
1. корнеплоды – разрастание паренхимных тканей главного корня для усиления запасающей функции его (морковь, петрушка, сельдерей).
2. корневые шишки – разрастание боковых корней, в которых откладываются запасные питательные вещества (георгин, батат, орхидные)
3. корни – присоски (гаустории) – развиваются на кончиках корней растений полупаразитов, таких как погремок, марьянник. С их помощью полупаразиты прикрепляются и внедряются в корни растения – хозяина и поглощают из его тканей воду и минеральные вещества.
4. воздушные корни – придаточные корни, развившиеся на стебле. Если стебель опускается на землю, они участвуют в заякоривании его.
5. Корни – доски
Метаморфозы корней
1. Микориза, растения называются микоризными.
а) эктотрофная – гифы охватывают корень только снаружи, корневые волоски при этом отмирают; редко гифы проникают в межклетники коровой паренхимы (сосна, берёза, дуб, бук, ива, а также у злаков).
б) эндотрофная – встречается более широко (у большинства цветковых). Она не образует корневой чехол, корневые волоски не отмирают, но гифы проникают внутрь корня и внедряются в летки коровой паренхимы.
2. Воздушные корни (эпифитные тропические орхидеи). Они покрыты многослойной эпиблемой, называемой веламеном. Иногда веламен выполняет функцию фотосинтеза и поглощения атмосферной влаги.
3. Втягивающие, или контрактильные корни (луковичные и корневищные растения). Укорачиваясь у основания, такие корни способны втягивать луковицы или корневища в почву на оптимальную для их сохранения глубину в период летней засухи или зимних морозов.
4. Дыхательные корни, или пневматофоры (растения болот и бедных кислородом почв). Растения затопляемых побережий образуют мангровые заросли.
5. Досковидные корни – выполняют функции опоры для огромной кроны тропических деревьев (баньян).
6. Запасающие корни: корнеплод и корнеклубень.
У корнеплода запасные питательные вещества откладываются в главном корне (морковь, петрушка, сельдерей). Однако у части корнеплодов большая их часть образована основанием стебля и разросшимся гипокотилем, а собственно корень представляет собой самую нижнюю часть корнеплода, несущую боковые корни.
Корнеклубни возникают при изменении мочковатой и ветвистой корневых систем при разрастании боковых корней (георгин, орхидные).
7. Клубеньки (бобовые и ряд видов других семейств). В них поселяются бактерии из рода Ризобиум или реже цианобактерии.
students-library.com
Анатомическое строение корня
Биология Анатомическое строение корня
просмотров - 544
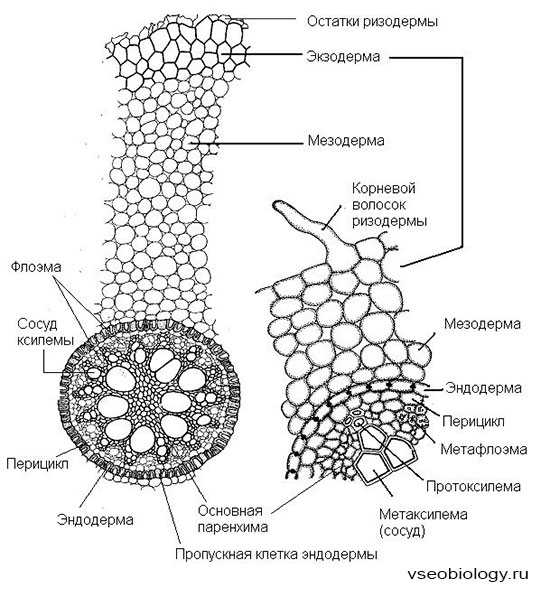
Первичное строение корня.Оно характерно для молодых корней всех растений. У плаунов, хвощей, папоротников и однодольных растений оно сохраняется в течение всей жизни. Первичное строение возникает в результате дифференциации меристемы конуса нарастания. На поперечном срезе корня в зоне всасывания можно различить три части: эпиблему, первичную кору и центральный осевой цилиндр (стелу).
Эпиблема (ризодерма), или кожица – первичная покровная ткань корня. Состоит из одного ряда плотно сомкнутых клеток, имеющих выросты – корневые волоски.
Первичная кора состоит из живых тонкостенных клеток в периферической части корня. Представлена тремя четко отличающимися друг от друга слоями: экзодермой, мезодермой и эндодермой.
| Рис. 1 Поперечный срез корня ириса (Iris germanica): 1 - ризодерма, 2 - экзодерма, 3 - основная паренхима (мезодерма), 4 - эндодерма, 5 - пропускная клетка эндодермы, 6 - перицикл, 7 - луч первичной ксилемы, 8 - участок первичной флоэмы (2-5 - первичная кора, 6-8 - центральный цилиндр). |
• Экзодерма располагается непосредственно под эпиблемой, являясь наружной частью первичной коры. Ее клетки многоугольные, плотно сомкнутые, располагаются в один или несколько рядов. По мере отмирания корневых волосков экзодерма оказывается на поверхности корня и выполняет роль покровной ткани, при этом происходит утолщение и опробковение клеточных оболочек и отмирание содержимого клеток.
• Мезодерма, или основная паренхима, располагается кнаружи от эндодермы. Состоит из рыхло расположенных клеток с системой межклетников, по которым идет интенсивный газообмен. Здесь происходит синтез и передвижение в другие ткани пластических веществ, накапливаются питательные вещества, располагается микориза.
• Эндодерма – самый внутренний слой коры, непосредственно прилегающий к стеле. У двудольных растений он состоит из одного ряда клеток, имеющих утолщения на радиальных стенках, – пояски Каспари, непроницаемые для воды. У однодольных растений образуются подковообразные утолщения клеточных стенок. В эндодерме встречаются живые тонкостенные клетки, которые называют пропускными клетками. Эти клетки также имеют пояски Каспари. Клетки эндодермы контролируют поступление воды и растворенных в ней минеральных веществ из коры в центральный цилиндр и обратно.
Центральный цилиндр, или осевой цилиндр, или стела, занимает центральную часть корня. Наружный слой стелы, примыкающий к эндодерме, принято называть перициклом. Его клетки долго сохраняют способность к делению. Здесь закладываются боковые корешки. В центральной части осевого цилиндра находится сосудисто-волокнистый пучок. Для корней характерно чередование в стеле участков первичной ксилемы и первичной флоэмы. Ксилема образует звезду, а между ее лучами располагается флоэма. Количество лучей ксилемы различно – от двух до нескольких десятков, у двудольных – до пяти, у однодольных – более пяти. В самом центре цилиндра могут находиться элементы ксилемы, склеренхима или тонкостенная паренхима.
Вторичное строение корня.У двудольных и голосеменных растений первичное строение корня сохраняется недолго. Примерно через 10 дней после прорастания семян происходят изменения, в результате которых возникает вторичное строение корня.
Процесс вторичных изменений начинается с появления прослоек камбия под участками первичной флоэмы, внутрь от нее. Камбий возникает из слабо дифференцированной паренхимы центрального цилиндра. Внутрь он откладывает элементы вторичной ксилемы (древесины), наружу – элементы вторичной флоэмы (луба). Сначала прослойки камбия разобщены, затем смыкаются, образуя сплошной слой. Это происходит благодаря делению клеток перицикла напротив лучей ксилемы. Камбиальные участки, возникшие из перицикла, образуют только паренхимные клетки серцевинных лучей, остальные клетки камбия образуют проводящие элементы – ксилему и флоэму. При делении клеток камбия исчезает радиальная симметрия, характерная для первичного строения корня.
В перицикле возникает и пробковый камбий (феллоген). Он откладывает наружу слои клеток вторичной покровной ткани – пробки. При этом первичная кора отмирает.
Читайте также
На протяжении корня от кончика к стеблю его строение неодинаково. В кончике корня (его верхушке) находится конус нарастания, покрытый корневым чехликом. Роль его — защищать глубже расположенные меристематические клетки от механических повреждений. Продвигаясь между... [читать подробенее]
oplib.ru







