
Обоеполые и однополые цветки. Растения обоеполые
Однополые и обоеполые цветки.
Обоеполым называют цветок, в котором есть как пестики, так и тычинки (андроцей и гинецей). Иногда к обоеполому цветку также применяют термины совершенный или однодомный цветок.
Цветок, который имеет только тычинки (андроцей) или только пестики (гинецей), называется однополым. Однополые цветки с тычинками — это тычиночные, или мужские цветки; соответственно цветки только с пестиками – пестичные, или женские цветки.
Мужские и женские однополые цветки могут вырастать на одном и том же растении, тогда растение называют однодомным, или обоеполым, например: дуб, береза, молочай, кукуруза. В этом случае опыление может происходить между цветками в пределах одного растения.
Если мужские и женские цветки вырастают на разных растениях, то имеем дело с двудомным растением. Двудомное растение с тычиночными цветками называют мужским, а с женскими – женским растением, например: тополь, ива, конопля, крапива. Для оплодотворения двудомных видов необходимо наличие как минимум двух разнополых растений – мужского и женского.
Растение, на котором встречаются как обоеполые, так и однополые цветки, называется полигамным,например, такое соседство встречается в соцветиях сложноцветных.
Цветки, лишенные спороносных органов – это стерильные, или бесполые цветки, как, например, язычковые цветки в соцветиях сложноцветных.
Пример полигамного растения: в соцветии герберы на фото имеются мужские цветки (с желтыми пыльниками), женские (с белыми пестиками) и стерильные язычковые цветки по краю.
Цветки злаковых, осоковых.
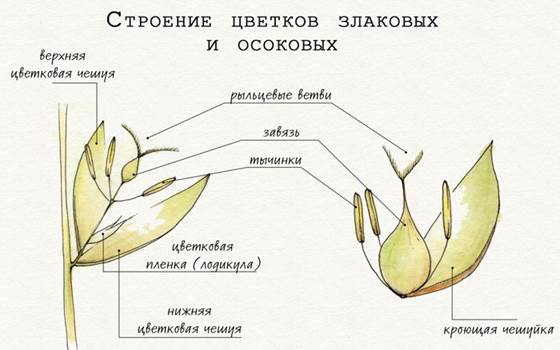
Цветки злаковых, осоковых.
Цветки злаковых, как правило, мелкие и неприметные. Они приспособлены для опыления ветром, и поэтому лишены околоцветника, так как им не зачем привлекать насекомых. Цветки злаков располагаются на боковых побегах колоска и состоят из тычинок и завязи с рыльцевыми ветвями. Цветок защищен верхней и нижней цветковыми чешуями. Выше цветковых чешуй вырастают две мелкие бесцветные чешуйки – так называемые цветковые пленки, или лодикулы. Во время цветения длинные тычинки выступают за пределы чешуй, распространяя пыльцу по ветру. Цветки злаковых могут быть обоеполыми или однополыми, иногда в пределах одного соцветия.
Цветки осоковых также мелкие и невзрачные, собраны в разнообразные колоски и сидят в пазухах прицветников, называемых кроющими чешуями. Сам цветок осоки состоит из тычинок и завязи с рыльцевыми ветвями. Цветки бывают обоеполые и однополые, с околоцветником и без. Околоцветник осоковых может состоять из набора чешуй, волосистых или бахромчатых щетинок или шелковистых волосков, и чаще присутствует у обоеполых или у женских цветков.
АНДРОЦЕЙ
(греч. «дом мужчины»): совокупность микроспорофиллов, тычинок состоящих из тычиночной нити с разделенным на две половинки пыльником, содержащим четыремикроспорангия (пыльцевых мешка). Тычинки располагаются в один-два круга. Тычинки делятся на свободные и сросшиеся.
Существуют разные типа андроцея, различимые по числу сросшихся групп тычинок:
-однобратственный (тычинки в одной группе, люпин, камелия),
-двубратственный (две группы тычинок),
-многобратственный (несколько групп, магнолия, зверобой),
-братственный (несросшиеся тычинки).
Также тычинки различаются по длине: равные, неравные, двусильные (из четырех тычинок две длинные), трехсильные (из шести тычинок три длинные),четырехсильные (из шести тычинок четыре длинные).
Тычинка состоит из тычиночной ткани, на верхнем конце которой расположенпыльник, а нижним концом прикрепленной к цветоложу. Основной тканью тычиночной нити является паренхима. В пыльнике происходят важные процессы –микроспорогенез (образование микроспор в микроспорангиях) и микрогаметогенез(образование из микроспор мужского гаметофита). Стерильная тычинка называетсястаминодией.
Рис.3 Тычинка и развитие пыльника
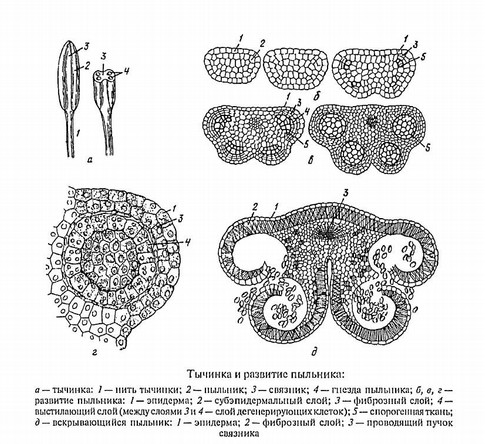
Пыльник состоит из однородных клеток, окруженных эпидермой.
Диаграмма – это схематическая проекция цветка на плоскости, при которой цветок пересекается поперек, перпендикулярно его оси. Правило оформления диаграммы: ось соцветия вверху, кроющий лист внизу. Условные обозначения диаграммы: дугами обозначаются части околоцветника, чашелистики – с выступом на середине дуги, лепестки – без выступа. Тычинки обозначаются в форме поперечного разреза пыльника или тычиночной нити. Гинецей - в виде поперечного разреза завязи. Если отдельные члены срастаются, это указывается на диаграмме дугами.
Андроцей
Андроцей представляет собой совокупность тычинок, в которых происходит микроспорогенез, микрогаметофитогенез и формирование мужских спор.
В онтогенезе тычинки могут закладываться в виде бугорков конуса нарастания как в акропетальной (т. е. от основания к верхушке), так и в базипетальной (от верхушки к основанию) последовательности. В первом случае самые молодые тычинки располагаются ближе к центру цветка, а старые ближе к его периферии, а во втором – наоборот. Тычинки могут быть свободными либо срастаются различным образом и в разной степени. Например, в тропическом семействе мелиевых все 10 тычинок срастаются своими нитями в трубку (однобратственный андроцей). У зверобоя тычинки срастаются в пучки, для сложноцветных характерно склеивание пыльников. У многих представителей семейства бобовые срастаются 9 тычинок, а одна остается свободной (так называемый двубратственный андроцей).
Каждая тычинка состоит из суженной нитевидной или редко лентовидной или лепестковидной части – тычиночной нити и обычно расширенной части – пыльника. Пыльник имеет две половинки, соединенные друг с другом связником, являющимся продолжением тычиночной нити. Связник иногда продолжен в надсвязник, заметный в виде небольшого выступа над пыльником.
Формирование нити начинается позже пыльника, и ее дальнейшее удлинение осуществляется за счет вставочного роста. Число формирующихся бугорков иногда меньше числа тычинок, в дальнейшем бугорки расщепляются, и тычинок может быть довольно много (мимозовые). Длина тычиночных нитей у разных растений варьирует. Чаще они более или менее равны по длине околоцветнику, но иногда значительно короче или во много раз его превышают, как, например, у известного тропического лекарственного растения почечный чай, или кошачьи усы, из семейства губоцветных. На поперечном срезе через тычиночную нить видно, что большая ее часть состоит из паренхимной ткани, а в центре проходит один сосудистый пучок.
Каждая половинка пыльника несет два (реже одно) гнезда – микроспорангия. Гнезда пыльников иногда называют пыльцевыми мешками. В зрелом пыльнике перегородки между гнездами по большей части исчезают. Снаружи пыльник покрыт эпидермой. Непосредственно под эпидермой располагается слой клеток так называемого эндотеция, имеющего вторично утолщенные клеточные оболочки. При подсыхании оболочек эндотеция вскрываются гнезда пыльника. Глубже залегают 1-3 слоя некрупных тонкостенных клеток. Самый внутренний слой, выстилающий полость пыльцевых мешков, получил название тапетума. Считается, что содержимое его клеток служит питанием для развивающихся материнских клеток микроспор (микроспороцитов) и способствует их дифференциации. Гнезда пыльника обычно заполнены материнскими клетками микроспор, микроспорами и зрелой пыльцой. Микроспоры, как известно, возникают из микроспороцитов в результате мейоза, а сами микроспороциты – из немногих клеток археспория (образовательной ткани, функционирующей на ранних стадиях развития гнезд пыльника). Созревший пыльник вскрывается разнообразно: продольными трещинами, дырочками, клапанами и др. При этом пыльца высыпается.
Признаки строения, формы, положения, числа тычинок, а также сам тип андроцея имеют большое значение для систематики цветковых и познания их филогении.
У некоторых видов часть тычинок теряет свою первоначальную функцию, становится стерильной и превращается в так называемые стаминодии. Иногда пыльники преобразуются в нектарники – секреторные части цветка, выделяющие нектар. Превратиться в нектарники или осмофоры могут также лепестки, их части, части пестика и даже выросты цветоложа. Нектарники имеют разнообразную форму, располагаются обычно в глубине цветка и нередко выделяются своей блестящей поверхностью.
Гинецей.
Совокупность плодолистиков одного цветка, образующих один или несколько пестиков, называют гинецеем. Плодолистики, или карпеллы, – это структуры, связанные, как считают, по происхождению с листом. Однако функционально и морфологически плодолистики соответствуют не вегетативным листьям, а листьям, несущим мегаспорангии, т. е. мегаспорофиллам. Большинство морфологов считают, что в ходе эволюции из плоских и открытых плодолистиков возникли сложенные вдоль (кондупликатно) плодолистики. Затем они срослись краями и образовали пестик с его наиболее существенной частью – завязью, несущей семязачатки. Таким образом, сформировалась уникальная, более ни у каких групп растений не встречающаяся структура, напоминающая замкнутый сосуд, в котором развиваются надёжно защищённые семязачатки. Структура пестика идеально приспособлена к опылению и оплодотворению. В семязачатках, находящихся в завязи, осуществляются процессы мегаспорогенеза и мегагаметофитогенеза.
Пестик, а точнее завязь, выполняет функцию влажной камеры, предохраняющей семязачатки от высыхания. Это сделало покрытосемянные растения практически независимыми от уровня влажности окружающей среды и явилось одним из факторов широкого освоения ими аридных территорий. Кроме того, пестик надежно укрывает семязачатки от поедания их насекомыми и отчасти от колебаний температуры.
Пестик, образовавшийся из одного плодолистика, называют простым, из двух и более сросшихся плодолистиков – сложным. Простой пестик обычно одногнездный; сложный может быть разделен на гнезда или бывает также одногнездным. Многогнёздность возникает либо в результате срастания плодолистиков, либо вследствие образования дополнительных перегородок – выростов стенок завязи.
Рыльце пестика – уникальная и характерная только для цветковых растений структура, предназначенная для восприятия пыльцы. Оно развивается на верхушке столбика или непосредственно на завязи – сидячее рыльце; реже (у архаичных видов) – вдоль сросшихся краев плодолистика. Форма и величина рыльца различны у разных видов. Поверхность рыльца очень часто неровная, бугорчатая и покрыта липкой жидкостью, что способствует более эффективной фиксации и улавливанию пыльцы. Кроме того, рыльцевая поверхность несет тонкий белковый слой – пелликулу, которая, взаимодействуя с белками спородермы пыльцевого зерна, обеспечивает прорастание пыльцевой трубки или препятствует ему.
Столбик состоит из рыхлой паренхимной ткани. Он как бы приподнимает рыльце вверх, что необходимо при некоторых механизмах процесса опыления. Морфология столбиков довольно разнообразна и служит важным систематическим признаком. Для многих архаичных семейств (особенно из подкласса магнолиид) характерно отсутствие или слабое развитие столбика. Столбики часто неразвиты и у многих специализированных ветроопыляемых форм, например у многих злаков. В крупных ветроопыляемых цветках (у некоторых видов лилий) столбики достигают значительной длины, рыльце выносится высоко вверх, и тем самым облегчается опыление. Однако при этом значительно удлиняется путь пыльцевой трубки.
Завязь – наиболее существенная часть пестика, несущая семязачатки. Она разнообразна по форме и внешнему виду, что в значительной степени определяется типом гинецея.
Место прикрепления семязачатков в завязи называется плацентой. Плацента обычно имеет вид небольшого вздутия, выроста или выступа, образованного тканями завязи.
В зависимости от особенностей прикрепления семязачатков к стенке завязи различают несколько типов плацентации.
· постенная, или париентальная, когда семязачатки располагаются внутри завязи по ее стенкам или в местах срастания плодолистиков.
· осевой, или аксиальной, когда семязачатки находятся на центральной колонке завязи, расчлененной на гнезда по числу плодолистиков.
· свободная центральная плацентация, когда семязачатки развиваются на свободной центральной колонке, не связанной перегородками со стенкой завязи.
· базальная, когда единственный семязачаток находится в самом основании одногнездной завязи.
Типы гинецеев:
1. апокарпный – плодолистики между собой не срастаются, и каждый плодолистик формирует отдельный пестик (лютик, роза)
а) мономерный – гинецей состоит из 1 пестика, а он образован 1 плодолистиком (горох, слива, вишня)
б) полимерный – пестиков много, но каждый состоит из одного плодолистика
(лютик, земляника, шиповник)
1. ценокарпный – пестик формируется из сросшихся между собой плодолистиков
а) синкарпный – плодолистики срастаются боковыми поверхностями, образуется несколько колец (тюльпан). Внутри плода образуется несколько гнёзд.
б) паракарпный – плодолистики срастаются краями и образуют одно кольцо (мак), или центральную камеру.
в) лизикарпный – плодолистики срастаются краями, образуя одну камеру или полость, а со дна завязи выступает колонка, на которой располагается семяпочка, а затем и семена (гвоздичные).
13. Семяпочка - относительно сложное образование, состоящее из семяножки (фуникулуса), несущей нуцеллус, заключенный в один или два интегумента. В зависимости от вида на плацентахразвиваются от одного до многих семязачатков. Развивающаяся семяпочка первоначально состоит целиком из нуцеллуса, но вскоре возникают один или два покровных слоя (интегумента) с небольшим отверстием, микропиле, на одном конце (рис.6).

Рис. 6. Схема формирования семязачатка и зародышевого мешка.
1, 2, 3, 4 – развитие нуцеллуса, обособление и мейоз клетки археспория, отмирание трех мегаспор; 5, 6, 7, 8 – развитие из мегаспоры (оставшейся) женского гаметофита – зародышевого мешка.
На ранней стадии развития семязачатка в нуцеллусе возникает единственный диплоидный мегаспороцит. Он делится митотически, давая четыре гаплоидные мегаспоры, обычно расположенные линейной тетрадой. Этим завершается мегаспорогенез. Три мегаспоры обычно разрушаются, а четвертая, наиболее удаленная от микропиле, развивается в женский гаметофит.
Функциональная мегаспора вскоре начинает увеличиваться за счет нуцеллуса, а ее ядро трижды делится митотически. В конце третьего митоза восемь дочерних ядер располагаются по четыре двумя группами – вблизи микропилярного конца мегагаметофита, а также на противоположном, халазальном, конце. По одному ядру из каждой группы мигрирует к центру восьмиядерной клетки; они называются полярными. Три оставшихся у микропилярного конца ядра образуют яйцевой аппарат, состоящий из яйцеклетки и двух клеток-синергид. У халазального конца также идет формирование вокруг расположенных здесь ядер клеточных оболочек, и возникают так называемые клетки-антиподы. Полярные ядра остаются в двуядерной центральной клетке. Такая восьмиядерная семиклеточная структура является зрелым женским гаметофитом, называемым зародышевым мешком.
stydopedia.ru
Обоеполые и однополые цветки
Цветки бывают обоеполые (с андроцеем и гинецеем) или однополые (только с андроцеем или только с гинецеем). Однополые цветки могут быть или на одном и том же растении, как у дуба, березы, молочая, кукурузы (и тогда растение как целое обоеполое), или же на разных растениях, как у тополя, ивы, конопли (тогда мы имеем мужские и женские растения). В связи с этим в ботанической литературе уже давно существуют два термина -однодомный и двудомный. Со времен Линнея многие ботаники эти термины применяют к самим растениям и говорят о растениях двудомных и однодомных. Если на растении встречаются как обоеполые, так и однополые цветки, как у многих сложноцветных, то о них говорят, что они полигамные (от греч. poly — много и gamos — брак). Однако, начиная с О. П. де Кандолля, С. Л. Зндлихора, Д. Вептама и Дж. Д. Хукера и кончая А. Энглером, Р. Веттгатейном, А. Б. Репдлом и Дж. Хатчинсоном, многие авторы применяют термины «двудомный» и. «однодомный» только к цветкам, а не к целым растениям. Возникающие иногда споры о том, какое из двух этих применений терминов более правильно, в сущности, лишены смысла. О конопле или об иве можно с равным основанием говорить, что они двудомные или что цветки у них двудомные. В зависимости от контекста может быть удобнее то одно, то другое употребление этих терминов и ни в одном случае это не вызовет недоразумения.
Имеются все основания считать, что однополые цветки возникли из обоеполых, а у однополых цветков двудомность — явленно более позднее, чем однодомность. Начиная со второй половины прошлого века многочисленные исследования по сравнительной морфологии и биологии опыления привели к выводу, что однополые цветки возникли из обоеполых в результате недоразвития или полного подавления в одних случаях тычинок, а в других — плодолистиков. В однополых цветках многих родов и целых семейств часто сохраняются редуцированные остатки (рудименты) тычинок и плодолистиков (так называемые стаминодии и карпеллодии). Такие остаточные образования можно видеть в цветках представителей самых разных семейств, в том числе у платана, некоторых тутовых, крапивных и ореховых. Основной биологической причиной перехода обоеполых цветков в однополые является более надежное перекрестное опыление, на что в свое время указал еще Чарлз Дарвин.
Прочитав эти строки, читатель может задать вопрос: а можно ли говорить о поле цветка, ведь цветок — это часть спорофита, или бесполого поколения, и тем самым лишен пола? Некоторые ботаники так и считают, и вместо терминов «мужской», «женский» и «обоеполый» предпочитают употреблять термины «тычиночный», «пестичный» и «совершенный» (совершенный в том смысле, что имеются как тычинки, так и плодолистики). Однако большинство ботаников продолжают пользоваться терминами «обоеполый» и «однополый», «мужской» и «женский», и притом с полным основанием. Морфологически цветок является, несомненно, частью спорофита, но функционально он имеет самое непосредственное отношение к половому процессу.
Когда мы говорим о мужских и женских цветках, то имеем в виду их роль в подготовке полового размножения, а не принадлежность к половому поколению (гаметофиту). Все дело в том, что генетическая и физиологическая дифференциации между мужским и женским полом переходит и на бесполое поколение, происходит определенная сексуализация спорофита. Это особенно хорошо выражено у двудомных растений (у растений с двудомными цветками). Мужские и женские растения конопли различаются генетически и физиологически, и можно даже сказать, что мужская конопля не в меньшей степени мужская, чем самцы у животных. С таким же основанием можно считать тычинку мужской структурой, а плодолистик — женской.
Поделитесь на страничкеslovar.wikireading.ru
Обоеполые и однополые цветки - это... Что такое Обоеполые и однополые цветки?
Цветки бывают обоеполые (с андроцеем и гинецеем) или однополые (только с андроцеем или только с гинецеем). Однополые цветки могут быть или на одном и том же растении, как у дуба, березы, молочая, кукурузы (и тогда растение как целое обоеполое), или же на разных растениях, как у тополя, ивы, конопли (тогда мы имеем мужские и женские растения). В связи с этим в ботанической литературе уже давно существуют два термина -однодомный и двудомный. Со времен Линнея многие ботаники эти термины применяют к самим растениям и говорят о растениях двудомных и однодомных. Если на растении встречаются как обоеполые, так и однополые цветки, как у многих сложноцветных, то о них говорят, что они полигамные (от греч. poly — много и gamos — брак). Однако, начиная с О. П. де Кандолля, С. Л. Зндлихора, Д. Вептама и Дж. Д. Хукера и кончая А. Энглером, Р. Веттгатейном, А. Б. Репдлом и Дж. Хатчинсоном, многие авторы применяют термины «двудомный» и. «однодомный» только к цветкам, а не к целым растениям. Возникающие иногда споры о том, какое из двух этих применений терминов более правильно, в сущности, лишены смысла. О конопле или об иве можно с равным основанием говорить, что они двудомные или что цветки у них двудомные. В зависимости от контекста может быть удобнее то одно, то другое употребление этих терминов и ни в одном случае это не вызовет недоразумения.
Имеются все основания считать, что однополые цветки возникли из обоеполых, а у однополых цветков двудомность — явленно более позднее, чем однодомность. Начиная со второй половины прошлого века многочисленные исследования по сравнительной морфологии и биологии опыления привели к выводу, что однополые цветки возникли из обоеполых в результате недоразвития или полного подавления в одних случаях тычинок, а в других — плодолистиков. В однополых цветках многих родов и целых семейств часто сохраняются редуцированные остатки (рудименты) тычинок и плодолистиков (так называемые стаминодии и карпеллодии). Такие остаточные образования можно видеть в цветках представителей самых разных семейств, в том числе у платана, некоторых тутовых, крапивных и ореховых. Основной биологической причиной перехода обоеполых цветков в однополые является более надежное перекрестное опыление, на что в свое время указал еще Чарлз Дарвин.
Прочитав эти строки, читатель может задать вопрос: а можно ли говорить о поле цветка, ведь цветок — это часть спорофита, или бесполого поколения, и тем самым лишен пола? Некоторые ботаники так и считают, и вместо терминов «мужской», «женский» и «обоеполый» предпочитают употреблять термины «тычиночный», «пестичный» и «совершенный» (совершенный в том смысле, что имеются как тычинки, так и плодолистики). Однако большинство ботаников продолжают пользоваться терминами «обоеполый» и «однополый», «мужской» и «женский», и притом с полным основанием. Морфологически цветок является, несомненно, частью спорофита, но функционально он имеет самое непосредственное отношение к половому процессу.
Когда мы говорим о мужских и женских цветках, то имеем в виду их роль в подготовке полового размножения, а не принадлежность к половому поколению (гаметофиту). Все дело в том, что генетическая и физиологическая дифференциации между мужским и женским полом переходит и на бесполое поколение, происходит определенная сексуализация спорофита. Это особенно хорошо выражено у двудомных растений (у растений с двудомными цветками). Мужские и женские растения конопли различаются генетически и физиологически, и можно даже сказать, что мужская конопля не в меньшей степени мужская, чем самцы у животных. С таким же основанием можно считать тычинку мужской структурой, а плодолистик — женской.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
dic.academic.ru
Обоеполые и Однополые Цветки - Биологическая энциклопедия
Цветки бывают обоеполые (с андроцеем и гинецеем) или однополые (только с андроцеем или только с гинецеем). Однополые цветки могут быть или на одном и том же растении, как у дуба, березы, молочая, кукурузы (и тогда растение как целое обоеполое), или же на разных растениях, как у тополя, ивы, конопли (тогда мы имеем мужские и женские растения). В связи с этим в ботанической литературе уже давно существуют два термина -однодомный и двудомный. Со времен Линнея многие ботаники эти термины применяют к самим растениям и говорят о растениях двудомных и однодомных. Если на растении встречаются как обоеполые, так и однополые цветки, как у многих сложноцветных, то о них говорят, что они полигамные (от греч. poly - много и gamos - брак). Однако, начиная с О. П. де Кандолля, С. Л. Зндлихора, Д. Вептама и Дж. Д. Хукера и кончая А. Энглером, Р. Веттгатейном, А. Б. Репдлом и Дж. Хатчинсоном, многие авторы применяют термины «двудомный» и. «однодомный» только к цветкам, а не к целым растениям. Возникающие иногда споры о том, какое из двух этих применений терминов более правильно, в сущности, лишены смысла. О конопле или об иве можно с равным основанием говорить, что они двудомные или что цветки у них двудомные. В зависимости от контекста может быть удобнее то одно, то другое употребление этих терминов и ни в одном случае это не вызовет недоразумения.
Имеются все основания считать, что однополые цветки возникли из обоеполых, а у однополых цветков двудомность - явленно более позднее, чем однодомность. Начиная со второй половины прошлого века многочисленные исследования по сравнительной морфологии и биологии опыления привели к выводу, что однополые цветки возникли из обоеполых в результате недоразвития или полного подавления в одних случаях тычинок, а в других - плодолистиков. В однополых цветках многих родов и целых семейств часто сохраняются редуцированные остатки (рудименты) тычинок и плодолистиков (так называемые стаминодии и карпеллодии). Такие остаточные образования можно видеть в цветках представителей самых разных семейств, в том числе у платана, некоторых тутовых, крапивных и ореховых. Основной биологической причиной перехода обоеполых цветков в однополые является более надежное перекрестное опыление, на что в свое время указал еще Чарлз Дарвин.
Прочитав эти строки, читатель может задать вопрос: а можно ли говорить о поле цветка, ведь цветок - это часть спорофита, или бесполого поколения, и тем самым лишен пола? Некоторые ботаники так и считают, и вместо терминов «мужской», «женский» и «обоеполый» предпочитают употреблять термины «тычиночный», «пестичный» и «совершенный» (совершенный в том смысле, что имеются как тычинки, так и плодолистики). Однако большинство ботаников продолжают пользоваться терминами «обоеполый» и «однополый», «мужской» и «женский», и притом с полным основанием. Морфологически цветок является, несомненно, частью спорофита, но функционально он имеет самое непосредственное отношение к половому процессу.
Когда мы говорим о мужских и женских цветках, то имеем в виду их роль в подготовке полового размножения, а не принадлежность к половому поколению (гаметофиту). Все дело в том, что генетическая и физиологическая дифференциации между мужским и женским полом переходит и на бесполое поколение, происходит определенная сексуализация спорофита. Это особенно хорошо выражено у двудомных растений (у растений с двудомными цветками). Мужские и женские растения конопли различаются генетически и физиологически, и можно даже сказать, что мужская конопля не в меньшей степени мужская, чем самцы у животных. С таким же основанием можно считать тычинку мужской структурой, а плодолистик - женской.
На нашем сайте Вы найдете значение "Обоеполые и Однополые Цветки" в словаре Биологическая энциклопедия, подробное описание, примеры использования, словосочетания с выражением Обоеполые и Однополые Цветки, различные варианты толкований, скрытый смысл.
Первая буква "О". Общая длина 53 символа
my-dict.ru









