ФИТОГОРМОНЫ Фитогормоны v Соединения с помощью которых. Основное место синтеза гормона цитокинина у вегетирующих растений
| Достижения молекулярной биологии и генетики позволили по новому подойти к исследованию механизма действия гормонов. Широко использовались модифицированные растения, как мутанты, так и трансгенные. Особенно большие успехи были достигнуты в изучении рецепции и трансдукции гормонов, а также их взаимодействия на транскрипционном уровне. Большинство ученых считают, что подобно гормонам животных первичной реакцией фитогормонов является взаимодействие с рецептором. Рецепторы— это молекулы белковой природы, которые распознают гормон, специфически связываются с ним, меняя свою конфигурацию с образованием гормон-рецепторного комплекса. Гормон-рецепторный комплекс передает гормональный сигнал, необходимый для «запуска» ответной физиологической реакции клетки. Рецепторы располагаются как на мембранах, так и в цитозоле. Один и тот же гормон может связываться с разными рецепторами, тем самым вызывая различные ответные физиологические реакции. Именно это является одной из причин многофункциональности в действии фитогормонов. Необходимо учитывать, что фитогормоны оказывают влияние только на восприимчивые к ним (компетентные) клетки. Одним из важнейших условий компетентности как раз и является наличие специфических рецепторов. Для идентификации белков-рецепторов с успехом используется генетический подход. Объектом наибольшего количества исследований служит крошечное растение сем. Крестоцветные Arabidopsis thaliana — арабидопсис (резушка Таля). Это связано с быстрым размножением этого растения, коротким жизненным циклом (около 6 недель) и изученностью его генома. Однако используются мутанты и трансгенные особи таких растений как кукуруза, табак, соя, томаты. Трудность этих исследований заключается в том, что, как правило, фитогормоны связываются со многими белками, но не все из них обладают рецепторными функциями, т. е. участвуют в дальнейшей передаче гормонального сигнала. Первый белок, обладающий рецепторной функцией, был обнаружен для ауксинов — AБП1— ауксинсвязывающий протеин. Предполагается, что этот протеин связан с гормоном при помощи иона металла. AБП1локализован на эндоплазматическом ретикулуме и частично на плаз-малемме. Именно та часть белка, которая расположена на плазмалемме, служит для восприятия ауксинового сигнала. Белок, находящийся на ЭПР является запасным. После связывания ауксина с рецептором следует активирование группы факторов транскрипции, входящих в сигнальную цепь. Это вызывает транскрипцию ранних генов (гены первичного ответа). Ингибиторы белкового синтеза, такие как циклогексимид, не тормозят экспрессию ранних генов. Время для экспрессии ранних генов ограничено и колеблется от нескольких минут до часа. Исследования гормонов животных показали, что гены прямого ответа выполняют несколько задач, в том числе кодируют белки, регулирующие транскрипцию генов вторичного ответа (поздних генов). В 1996 г. Абель с сотрудниками идентифицировали 5 групп ранних ауксин-зависимых генов. Гены первой группы кодируют образование маленьких полипептидных цепочек молекулярной массой 19—36 кДа. Эти цепочки являются короткоживущими факторами, регулирующими транскрипцию, и служат репрессорами или активаторами для экспрессии поздних генов. Поздние гены кодируют белки, вызывающие физиологическую реакцию. Например, в случае ауксина фактор регуляции транскрипции вызывает экспрессию позднего гена, который кодирует протеазы и рибонуклеазы, участвующие в дифференциации клеток и образовании элементов ксилемы. Поиск рецепторов цитокининов в течение ряда лет осуществлялся в разных лабораториях. Мембранный рецептор цитокининов был выделен в 2001 г. Этому предшествовало обнаружение мутантов арабидопсиса, нечувствительных к цитокинину. Был выделен соответствующий ген и кодируемый этим геном белок-рецептор. Этот рецептор локализован в плазмалемме. Предполагается, что он принадлежит к классу бикомпонентных регуляторов. Один домен этого белка-рецептора, расположенный на внешней стороне мембраны, узнает и связывает цитокинин. Связывание приводит к фосфорилированию, и как следствие к изменению конфигурации и активации рецептора. Далее включается сигнальная цепь, представляющая каскад фосфорилирования, в конце которого фосфат переносится на факторы регуляции транскрипции генов. В настоящее время в арабидопсисе идентифицировано семь генов первичного ответа на цитокинины. Нельзя не отметить, что имеются данные о регуляции цитокининами синтеза белков не только на уровне транскрипции, но и независимым от транскрипции путем (О.Н. Кулаева, В.В. Кузнецов). Это может происходить за счет регуляции цитокининами фосфорилирования рибосомальных белков и белковых факторов трансляции. Рецепторы этилена по своей структуре сходны с рецепторами цитокининов. Исследования проводились на мутантах арабидопсиса, нечувствительных к этилену. Был выделен соответствующий ген ETR1(триплетный ответ на этилен). Этот ген кодирует сенсорную гистидинкиназу, сходную с рецептором цитокининов. Связывание этилена с рецептором происходит с участием меди. Дальнейшие исследования показали, что этилен может связываться с пятью аналогичными белками. Все они относятся к его рецепторам. Распределение различных рецепторов этилена тканеспецифично. Сигнал от всех рецепторов передается на про-теинкиназу и далее через МАР-киназный каскад. Это путем фосфорилирования активирует фактор регуляции транскрипции и индукцию генов. Вместе с тем есть данные, что в передаче этиленового сигнала от рецептора на MAP — киназы участвуютG-белки. О рецепторе гиббереллинов известно мало, но предполагают, что, по-видимому, он находится на поверхности клеток. Наиболее изучен механизм действия гиббереллинов на образование ос-амилазы и других гидролитических ферментов в алейроновом слое прорастающих семян. Показано, что гиббереллин влияет на синтез а-амилазы de novo на уровне транскрипции. Фактор транскрипции связывается с ДНК на особом гиббереллинчувствительном участке — промоторе. Мутация в этом участке приводит к потере способности экзогенного гиббереллина индуцировать экспрессию а-амилазы. Установлена зависимость секреции уже образовавшегося фермента от содержания кальция и кальцийсвя-зывающего белка — кальмодулина. Рецептор АБК не идентифицирован. Но опыты с инъекцией АБК в плазма-лемму показали, что рецептор находится на поверхности мембраны. Не исключено наличие второго рецептора, расположенного внутри мембраны. Имеются данные, что АБК оказывает влияние на фактор регуляции транскрипции. Таким образом, взаимодействие с гормоном меняет конформацию рецептора, переводя его в активированное состояние. Активированные мембранные рецепторы передают сигнал внутрь клетки с помощью тех или иных каскадных механизмов с участием вторичных посредников. Передаваемый сигнал непосредственно взаимодействует с промоторной областью ДНК и при участии фактора регуляции транскрипции вызывает экспрессию генов. Влияние фитогормонов на новообразование белков-ферментов может осуществляться и на постгранскрипционном уровне. В частности, показано значение в этом процессе гиббереллина. Фитогормоны могут регулировать время жизни мРНК, а также процесс ее поступления в цитоплазму. Ряд исследований показывает, что под влиянием цитокинина повышается функциональная активность рибосом, а также их число, что и приводит к возрастанию синтеза белков-ферментов. Имеются прямые экспериментальные доказательства взаимосвязи фитогормонов с синтезом белков-ферментов. Действие фитогормонов не проявляется при введении ингибиторов синтеза мРНК и белка. В тоже время показано влияние гормонов на увеличение новообразования мРНК и белка. При неблагоприятных условиях гормоны участвуют в индукции генов, кодирующих образование стрессовых белков. Так, ауксины стимулируют биосинтез таких ферментов, как целлюлаза, цел-люлозосинтетаза, пектинметилэстераза, АТФаза и др. Гиббереллины активируют образование ряда гидролаз и ферментов, катализирующих синтез липидов, входящих в состав мембран. Четкие данные по индуцированию синтеза ферментов, таких, как нитратредуктаза, РБФ-карбоксилаза/оксигеназа и др., получены для цитокининов. Фитогормоны, ингибирующие рост (АБК), тормозят синтез ряда белков. В то же время АБК, накапливаясь в семенах в период их естественного обезвоживания, индуцирует синтез белков в позднем эмбриогенезе. Этилен активирует образование белков-ферментов, участвующих в процессах, стимулирующих созревание плодов. Однако наряду с действием на дифференциальную активность генома большое значение имеет влияние фитогормонов на мембраны. Действие на уровне мембран, по-видимому, характерно для всех фитогормонов. Так, существуют данные, что под влиянием ауксина изменяется толщина мембран. Гормоны регулируют проницаемость мембран. Ауксины и цитокинины усиливают поступление ионов через мембраны. АБК и этилен индуцируют выход ионов из клетки. Под влиянием гиббереллинов возрастает проницаемость мембран для Сахаров. Показано также, что гиббереллин вызывает новообразование мембран эндоплаз-матического ретикулума, способствуя синтезу их липидных компонентов. Показано, что под влиянием ауксина происходит выход протонов на наружную сторону плазмалеммы, развивается гиперполяризация мембран, возрастает ΔμН+(В.В. Полевой). Как известно, ΔμН+возникает либо благодаря работе редокс-цепи, либо за счет распада АТФ в результате действия АТФ-фазы: ΔμН+-> АТФ -> ΔμН+. Имеются данные, что ИУК активизирует АТФазу, локализованную в плазмалемме. Это и может быть причиной активации выхода протонов (Н+-помпа). Вместе с тем ауксины повышают интенсивность дыхания, ускоряя работу дыхательной цепи (редокс-цепь), следствием чего также может быть усиление выброса протонов через мембрану митохондрий. Этот механизм является одной из основ действия ИУК на рост растяжением. В настоящее время признано, что способность к росту определяется энергетическим потенциалом клетки (отношение АТФ/АДФ). При взаимодействии с мембранами фитогормоны влияют именно на поток энергии. Усиливаются процессы окислительного фосфорилирования (ауксины), фотосинтетического фосфорилирования (гиббереллины, цитокинины). Образовавшаяся после взаимодействия фитогормонов с мембранами АТФ, наряду с использованием на различные синтезы, может служить источником образования циклической АМФ (цАМФ). Последняя образуется из АТФ при участии связанного с мембранами фермента аденилатциклазы. Под влиянием цАМФ активируются ферменты протеинкиназы, катализирующие фосфорилирование белков, участвующих в сигнальных цепях и транскрипции. Изложенный материал показывает, что под влиянием условий среды, а также в зависимости от свойств данной клетки (ее местоположения) в ней создается определенное соотношение гормонов. Это, в свою очередь, определяет развертывание генетической программы, темпы роста, дифференциацию и развитие. Надо только учитывать, что клетка должна быть восприимчива, или компетентна, к действию фитогормонов, в частности, должна обладать соответствующими рецепторами. |
studfiles.net
| Действие цитокининов, как и других фитогормонов, многофункционально. Цитокинины в первую очередь оказывают влияние на деление клеток, хотя в некоторых случаях могут регулировать и их растяжение. Особенно ярко влияние цитокининов на процессы деления проявляется на культуре изолированных тканей. На листьях целого растения показано соответствие активности цитокининов и скорости клеточных делений (А.Т. Мокроносов). Высказывается предположение, что цитокинины регулируют последнюю стадию деления, а именно цитокинез (деление самой клетки). Показано, что цитокинины стимулируют экспрессию специфического циклина и ускоряют переход от фазы g2 к митозу. Кроме того, они активируют рост растяжением изолированных листьев и семядолей у двудомных растений. Цитокинины также оказывают влияние на направление дифференциации клеток и тканей. Так, на стеблевом каллусе табака показано их участие в органообразовании. Влияние цитокининов тесно связано с присутствием ауксинов. Взаимосвязь в действии этих фитогормонов проявляется по-разному. В некоторых случаях действие цитокининов требует определенной концентрации ауксинов. В других случаях проявляется антагонизм этих фитогормонов. Цитокинины способствуют пробуждению и росту боковых почек. В опытах К. Тиманна обработка пазушных почек растворами кинетина устраняла тормозящее влияние верхушечных почек, вызывая их рост. В этом случае цитокинины являются как бы антагонистами ауксинов, которые задерживают рост боковых побегов (апикальное доминирование). Цитокинины задерживают старение листьев. Одним из показателей процесса старения является разрушение хлорофилла. Так, если срезанные листья выдерживать во влажной атмосфере, они постепенно желтеют. Анализы показывают, что в них происходит разрушение хлорофилла и белка. Обработанные цитокининами листья остаются в течение долгого времени зелеными. Цитокинины не только задерживают распад белка и хлорофилла, но и стимулируют синтез этих соединений (омолаживающее влияние). Показано усиление под действием цитокининов синтеза хлоропластных белков, в т. ч. тех, которые кодируются геномом ядра. Нельзя не отметить, что такое же омолаживающее влияние на листья проявляется при их укоренении. Это является еще одним доказательством, что цитокинины вырабатываются в корнях. Цитокинины оказывают влияние на ультраструктуру хлоропластов. Опыты показали, что обработка кинетином ускоряет дифференциацию пластид, образование в них мембран и гран (И.Н. Свешникова, О.Н. Кулаева). Цитокинины повышают содержание хлорофилла, ускоряя образование его предшественника — протохлорофиллида. Под влиянием цитокининов усиливается интенсивность фотофосфорилирования, возрастает активность сопрягающего фактора (АТФ-синтаза) в хлоропластах, содержание АТФ увеличивается (Н.И. Якушкина, С.М. Похлебаев). Вместе с тем цитокинины активируют синтез ключевого фермента темновых реакций фотосинтеза — РБФ-карбоксилазы/оксигеназы (С.С. Повар). Интенсивность фотосинтеза у обработанных цитокинином листьев возрастает. Возможно, отчасти повышению интенсивности фотосинтеза способствует усиление оттока ассимилятов, а также изменение степени открытости устьиц. Цитокинины, подобно ауксинам, усиливают передвижение веществ к обогащенным ими тканям (аттрагирующее влияние), Так, в опытах немецких физиологов К. Мотеса и Л. Энгельбрехт показано, что аминокислоты и углеводы перетекают из необработанной половины листа к половине, которая обогащена кинетином. Это проявляется как на изолированных листьях, так и на интактных растениях. Имеются данные о стимуляции кинетином флоэмного транспорта. Усиливая поглощение калия замыкающими клетками устьиц, цитокинины способствуют их открыванию. Цитокинины влияют на азотный обмен растений, стимулируя синтез белка-фермента нитратредукатазы. В ряде исследований отмечается, что обработка растений цитокининами повышает устойчивость к различным неблагоприятным условиям среды. Подобно гиббереллинам, цитокинины способствуют выходу из покоящегося состояния спящих почек, семян, клубней. Как уже говорилось, цитокинины образуются в корнях и у клубненосных растений, передвигаясь акропетально, вызывают утолщение столонов и образование клубней. |
fizrast.ru
Цитокинины — WiKi
Цитокинины (греч. κύτταρο ячейки + греч. κίνηση движение), класс гормонов растений 6-аминопуринового ряда, стимулирующих деление клеток (цитокинез). С этой способностью цитокининов связаны их основные функции в развитии растений — например, поддержание апикальной меристемы побега. Кроме того, к физиологическим функциям цитокининов относятся стимуляция транспорта питательных веществ в клетку, ингибирование роста боковых корней, замедление старения листьев[1]. Молекулярная масса (~ 5-20 кДа).
Цитокинины вовлечены в рост растительных клеток и другие физиологические процессы. Эффект цитокининов впервые был открыт на табаке в 1948 году Фольком Скугом.[2]
Кроме природных цитокининов — производных 6-аминопурина, представленных изопентинилом, зеатином и 6-бензиламинопурином, известны и синтетические приозводные фенилмочевины, стимулирующие цитокинеза у растений — N,N'-дифенилмочевина и тидиазурон (N-фенил-N'-(1,2,3-тиадиазол-5-ил)мочевина). Цитокинины синтезируется в основном в корнях, а также в стеблях и листьях. Камбий и другие активно делящиеся ткани растений также являются местом синтеза цитокининов.[3] Не показано, что цитокинины типа фенилмочевины естественно встречаются в тканях растений.[4] Цитокинины участвуют в местной передаче сигнала, а также в передаче сигнала на расстоянии, причем последний механизм также используется для транспорта пуринов и нуклеозидов.[5]
История открытия
Открытие цитокининов связано с рядом экспериментов (Ф. Скуг), направленных на получение растительной культуры клеток. Сердцевинную паренхиму стеблей табака помещали на искусственные среды, содержащие минеральные вещества, сахар, витамины, аминокислоты, ИУК.В среды начали добавлять ДНК из молок сельди. Сначала это не привело к успеху, но, благодаря ошибке при проведении автоклавирования (среда с ДНК перегрелась), паренхима начала активно делиться. Оказалось, что в перегретом препарате ДНК содержался фурфуриладенин (кинетин), который на фоне ауксина, вызывает деление клеток[6]. Первый натуральный цитокинин — зеатин — был выделен в 1974 году[1].
Механизм действия
Цитокинины участвуют во многих физиологических процессах растений, регулируют деления клеток, морфогенез побега и корня, созревание хлоропластов, линейный рост клетки, образование добавочных почек и старение.[7] Соотношение ауксинов к цитокининам является ключевым фактором деления клеток и дифференцировки тканей растения.
В то время, как эффект цитокининов на сосудистые растения является плейотропным, цитокинины вызывают изменения роста протонемы у мхов. Образование почек можно считать вариантом дифференцировки клеток и этот процесс является очень специфическим эффектом цитокининов.[8]
Биосинтез
Предшественниками биосинтеза цитокининов в растениях являются свободные АТФ и АДФ, а также тРНК. Первая стадия биосинтеза цитокининов — синтез изопентил-нуклеотидов из АТФ или АДФ и диметилаллилпирофосфата — катализируется ферментом изопентенилтрасферазой (IPT). Кроме IPT, у растений выявлены ферменты тРНК-IPT, использующие в качестве субстрата тРНК — они используются для синтеза цис-зеатина. В дальнейшем изопентенил-нуклеотиды могут превращаться в зеатин-нуклеотиды с помощью фитохром P450-монооксигеназ. Наконец, последней стадией является получение активных цитокининов из цитокининовых нуклеотидов путём дефосфорилирования и дерибозилирования — это реакция катализируется ферментом 5’монофосфат-фосфорибогидролазой, который кодируется геном LOG.[1]
Фермент аденозинфосфатизопентилтрансфераза катализирует первую реакцию в биосинтезе изопреновых цитокининов, фермент использует АТР, ADP или AMP как субстрат и диметилаллилдифосфат или гидроксиметилбутенилдифосфат как донор пренильной группы.[9] Данная реакция является лимитирующей в биосинтезе цитокининов, субстраты—доноры пренильных групп образуются в пентилэритрол-фосфатном биохимическом пути.[9]
У растений и бактерий цитокинины также могут образовываться из продуктов распада тРНК.[9][10] Транспортные РНК, с антикодоном, начинающимся с уридина и имеющие пренилированные аденозины рядом с антикодоном, освобождают при деградации аденозины как цитокинины.[9] Пренилирование таких аденинов осуществляется тРНК-изопентилтрансферазой[10]
Показано также, что ауксины регулируют биосинтез цитокининов.[11]
По последним данным, разные этапы биосинтеза цитокининов осуществляются в разных тканях растения. Основным местом синтеза цитокининовых нуклеотидов является кончик корня, небольшое их количество синтезируется также в апексе побега, цветках и плодах. По ксилеме цитокининовые нуклеотиды доставляются в апекс побега, который является основным местом синтеза активных свободных цитокининов[1].
Катаболизм и инактивация цитокининов
Основные ферменты катаболизма цитокининов — цитокинин-оксидазы, которые локализованы в вакуолях и эндоплазматическом ретикуллуме (ЭР) и осуществляет расщепление цитокининов с образованием аденина. Субстратами цитокинин-оксидаз являются свободные цитокинины и их рибозиды. Помимо расщепления цитокинин-оксидазами возможна обратимая или необратимая инактивация цитокининов путём образования конъюгатов[1].
Транспорт цитокининов
Основной транспортной формой цитокининов является зеатин-рибозид (ксилемный транспорт). Кроме того, существует транспорт цитокининов по флоэме, благодаря которому свободные цитокинины и их конъюгаты могут перемещаться по растению в обоих направлениях.
Транспорт цитокининов между клетками растения осуществляют две группы белков:
- пуринпермеазы (PUP), которые транспортируют в клетку свободные цитокинины, а также аденин.
- равновесные транспортеры нуклеозидов (ENT), которые осуществляют транспорт в клетку цитокинин-рибозидов[1].
Функции цитокининов в развитии растений
Функции цитокининов в развитии растений очень многообразны:
- контроль пролиферации клеток;
- координация роста и развития растений в зависимости от доступности минерального и органического питания;
- поддержание апикальной меристемы побега и ингибирование развития корневой системы;
- предотвращение старения листьев.
В контроле большинства онтогенетических процессов цитокинины являются антагонистами ауксинов и гиббереллинов[1].
Примечания
- ↑ 1 2 3 4 5 6 7 Лутова Л.А., Ежова Т.А., Додуева И.Е., Осипова М.А. Генетика развития растений. / С.Г. Инге-Вечтомов. — Санкт-Петербург, 2011. — С. 432. — ISBN 978-5-94869-104-6.
- ↑ J.J. Kieber (2002): Tribute to Folke Skoog: Recent advances in our understanding of cytokinin biology. Journal of Plant Growth Regulation 21, 1-2. [1]
- ↑ Chen, C. et al. 1985. Localization of Cytokinin Biosynthetic Sites in Pea Plants and Carrot Roots. Plant Physiology 78:510-513.
- ↑ Mok, DWS and Mok, MC. 2001. Cytokinin metabolism and action. Annual Review of Plant Physiology and Plant Molecular Biology 52: 89-118
- ↑ Sakakibara, H. 2006. Cytokinins: Activity, Biosynthesis, and Translocation. Annual Review of Plant Biology 57: 431—449
- ↑ Физиология растений: учебник для студентов ВУЗов / под ред. И. П. Ермакова.
- ↑ Kieber JJ (2002 Cytokinins. In CR Somerville, EM Meyerowitz, eds, [www.aspb.org/publications/arabidopsis/ The Arabidopsis Book]. American Society of Plant Biologists, Rockville, MD, doi: 10.1199/tab.0009
- ↑ Eva L. Decker, Wolfgang Frank, Eric Sarnighausen, Ralf Reski (2006): Moss systems biology en route: Phytohormones in Physcomitrella development. Plant Biology 8, 397—406 [2]
- ↑ 1 2 3 4 Ildoo Hwang, Hitoshi Sakakibara (2006) Cytokinin biosynthesis and perception Physiologia Plantarum 126 (4), 528—538
- ↑ 1 2 Kaori Miyawaki, Miho Matsumoto-Kitano, Tatsuo Kakimoto (2004) Expression of cytokinin biosynthetic isopentenyltransferase genes in Arabidopsis: tissue specificity and regulation by auxin, cytokinin, and nitrate The Plant Journal 37 (1), 128—138
- ↑ Nordström, A. 2004. Auxin regulation of cytokinin biosynthesis in Arabidopsis thaliana: A factor of potential importance for auxin-cytokinin-regulated development. PNAS 101:8039-8044
Ссылки
ru-wiki.org
Ауксины — Википедия
Материал из Википедии — свободной энциклопедии
4-Хлориндолил-3-уксусная кислота Индолил-3-масляная кислотаАукси́ны (от др.-греч. αὔξω — увеличиваюсь, расту) — стимуляторы роста плодов (побегов) растений, апикальное доминирование, растет по фототропизму (к свету), стимулирует рост корней по гравитропизму (рост вниз), обладают высокой физиологической активностью.
Природные ауксины являются производными индола — 3-(3-индолил)пропионовая, индолил-3-масляная, 4-хлориндолил-3-уксусная и 3-индолилуксусная кислоты. Наиболее распространенным ауксином, широко применяющимся в растениеводстве, является гетероауксин — индолил-3-уксусная кислота.
- влияют на рост клетки в фазах растяжения
- стимулируют рост клеток камбия
- обуславливают взаимодействие отдельных органов
- регулируют коррелятивный рост
- перемещается со скоростью 10 мм в час
- уменьшение концентрации ауксина в растении приводит к увяданию листьев
- дифференцирует клетки
- помогает при росте придаточных корней
ru.wikipedia.org
ФИТОГОРМОНЫ Фитогормоны v Соединения с помощью которых
 ФИТОГОРМОНЫ
ФИТОГОРМОНЫ
 Фитогормоны v. Соединения, с помощью которых осуществляется взаимодействие клеток, тканей и органов и которые в малых количествах необходимы для запуска и регуляции физиологических и морфогенетических программ. v. Система регулирующих механизмов – осуществляется настройка процессов жизнедеятельности и реакция на внешние факторы. v. Низкомолекулярные органические вещества. v. Образуются в различных тканях и органах и действуют в очень низких концентрациях.
Фитогормоны v. Соединения, с помощью которых осуществляется взаимодействие клеток, тканей и органов и которые в малых количествах необходимы для запуска и регуляции физиологических и морфогенетических программ. v. Система регулирующих механизмов – осуществляется настройка процессов жизнедеятельности и реакция на внешние факторы. v. Низкомолекулярные органические вещества. v. Образуются в различных тканях и органах и действуют в очень низких концентрациях.
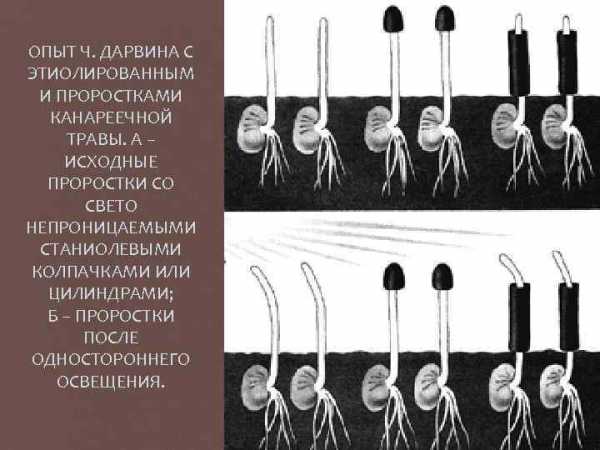 ОПЫТ Ч. ДАРВИНА С ЭТИОЛИРОВАННЫМ И ПРОРОСТКАМИ КАНАРЕЕЧНОЙ ТРАВЫ. А – ИСХОДНЫЕ ПРОРОСТКИ СО СВЕТО НЕПРОНИЦАЕМЫМИ СТАНИОЛЕВЫМИ КОЛПАЧКАМИ ИЛИ ЦИЛИНДРАМИ; Б – ПРОРОСТКИ ПОСЛЕ ОДНОСТОРОННЕГО ОСВЕЩЕНИЯ.
ОПЫТ Ч. ДАРВИНА С ЭТИОЛИРОВАННЫМ И ПРОРОСТКАМИ КАНАРЕЕЧНОЙ ТРАВЫ. А – ИСХОДНЫЕ ПРОРОСТКИ СО СВЕТО НЕПРОНИЦАЕМЫМИ СТАНИОЛЕВЫМИ КОЛПАЧКАМИ ИЛИ ЦИЛИНДРАМИ; Б – ПРОРОСТКИ ПОСЛЕ ОДНОСТОРОННЕГО ОСВЕЩЕНИЯ.
 Группы фитогормонов: 1. 2. 3. 4. 5. Ауксины. Гиббереллины. Цитокинины. АБК. Этилен.
Группы фитогормонов: 1. 2. 3. 4. 5. Ауксины. Гиббереллины. Цитокинины. АБК. Этилен.
 Ауксины • Ауксины были открыты Дарвином в результате изучения роста растяжением. • Вент выделил это ростовое вещество, накладывая отрезанные верхушки колеоптилей на агаровую пластинку. • В 1935 г. В лаборатории Ф. Кегля , это вещество было идентифицировано как индол-3 -уксусная кислота (ИУК). Соединения этой группы Кегль назвал ауксинами.
Ауксины • Ауксины были открыты Дарвином в результате изучения роста растяжением. • Вент выделил это ростовое вещество, накладывая отрезанные верхушки колеоптилей на агаровую пластинку. • В 1935 г. В лаборатории Ф. Кегля , это вещество было идентифицировано как индол-3 -уксусная кислота (ИУК). Соединения этой группы Кегль назвал ауксинами.
 Ауксины • Метаболизм и транспорт. У высших растений ее больше всего в развивающихся почках и листьях, в активном камбии, в формирующихся семенах, в пыльце. Синтез ауксина наиболее интенсивен в верхушке главного побега. Транспорт ИУК в растительных тканях осуществляется полярно со скоростью 10 -15 мм/ч от верхушки побега к корню. В стебле передвижение идет по живым клеткам проводящих пучков.
Ауксины • Метаболизм и транспорт. У высших растений ее больше всего в развивающихся почках и листьях, в активном камбии, в формирующихся семенах, в пыльце. Синтез ауксина наиболее интенсивен в верхушке главного побега. Транспорт ИУК в растительных тканях осуществляется полярно со скоростью 10 -15 мм/ч от верхушки побега к корню. В стебле передвижение идет по живым клеткам проводящих пучков.
 Ауксины • Физиология и биохимия действия. v Ауксин активирует деление и растяжение клеток, необходим для формирования проводящих пучков и корней, способствует разрастанию околоплодника. v Ткани, обогащенные ауксином, обладают аттрагирующим действием, т. е. способны притягивать питательные вещества. v В ряде случаев обработка ауксином задерживает процессы старения тканей и органов. v Ауксин обуславливает явление апикального доминирования, т. е. тормозящее влияние апикальной почки на рост пазушных почек. v Первостепенную роль играет ИУК в ростовых движениях – тропизмах и настиях.
Ауксины • Физиология и биохимия действия. v Ауксин активирует деление и растяжение клеток, необходим для формирования проводящих пучков и корней, способствует разрастанию околоплодника. v Ткани, обогащенные ауксином, обладают аттрагирующим действием, т. е. способны притягивать питательные вещества. v В ряде случаев обработка ауксином задерживает процессы старения тканей и органов. v Ауксин обуславливает явление апикального доминирования, т. е. тормозящее влияние апикальной почки на рост пазушных почек. v Первостепенную роль играет ИУК в ростовых движениях – тропизмах и настиях.
 Гиббереллины • Японский исследователь Е. Куросава в 1926 г. Установил, что культуральная жидкость фитопатогенного гриба Gibberella fujikuroi содержит химическое вещество, способствующее сильному вытягиванию стеблей у растений. • Т. Ябута (1938) выделил это вещество в кристаллическом виде и назвал его гиббереллином. • В 1954 г. Англичанин Б. Кросс расшифровал структуру гиббереллиновой кислоты – тетрациклического дитерпеноида. • Основная роль при прорастании, образовании цветков и росте плодов. • Образуется в верхушках стебля, кончике корня, в молодых листьях.
Гиббереллины • Японский исследователь Е. Куросава в 1926 г. Установил, что культуральная жидкость фитопатогенного гриба Gibberella fujikuroi содержит химическое вещество, способствующее сильному вытягиванию стеблей у растений. • Т. Ябута (1938) выделил это вещество в кристаллическом виде и назвал его гиббереллином. • В 1954 г. Англичанин Б. Кросс расшифровал структуру гиббереллиновой кислоты – тетрациклического дитерпеноида. • Основная роль при прорастании, образовании цветков и росте плодов. • Образуется в верхушках стебля, кончике корня, в молодых листьях.
 Метаболизм и транспорт • Наибольшее количество гиббереллинов у высших растений содержится в незрелых семенах. • Синтезируются главным образом в листьях, а также в корнях. • Транспорт их происходит пассивно с ксилемным и флоэмным током. • Синтезируются из ацетил-Со. А через мевалоновую кислоту и геранилгераниол; ближайший его предшественник – каурен. • Связанные в виде гликозидов ГА являются запасной и транспортной формами.
Метаболизм и транспорт • Наибольшее количество гиббереллинов у высших растений содержится в незрелых семенах. • Синтезируются главным образом в листьях, а также в корнях. • Транспорт их происходит пассивно с ксилемным и флоэмным током. • Синтезируются из ацетил-Со. А через мевалоновую кислоту и геранилгераниол; ближайший его предшественник – каурен. • Связанные в виде гликозидов ГА являются запасной и транспортной формами.
 Физиология и биохимия действия • Действие ГА на растения наиболее типично проявляется в удлинение их стебля. • Места действия ГА – апикальные и интеркалярные меристемы, где под влиянием этого фитогормона активируется деление клеток. Ускоряется также растяжение клеток, однако непосредственно на этот процесс ГА действует слабо. • Гиббереллины не стимулируют рост корня, а в повышенных концентрациях даже ухудшают его состояние.
Физиология и биохимия действия • Действие ГА на растения наиболее типично проявляется в удлинение их стебля. • Места действия ГА – апикальные и интеркалярные меристемы, где под влиянием этого фитогормона активируется деление клеток. Ускоряется также растяжение клеток, однако непосредственно на этот процесс ГА действует слабо. • Гиббереллины не стимулируют рост корня, а в повышенных концентрациях даже ухудшают его состояние.
 РУДБЕНИЯ, ВЕРХУШЕЧНАЯ ПОЧКА КОТОРОЙ ОБРАБОТАНА ГИББЕРЕЛЛИНОМ (СЛЕВА) И КОНТРОЛЬНОЕ РАСТЕНИЕ, ПОСАЖЕННЫЕ ОДНОВРЕМЕННО.
РУДБЕНИЯ, ВЕРХУШЕЧНАЯ ПОЧКА КОТОРОЙ ОБРАБОТАНА ГИББЕРЕЛЛИНОМ (СЛЕВА) И КОНТРОЛЬНОЕ РАСТЕНИЕ, ПОСАЖЕННЫЕ ОДНОВРЕМЕННО.
 Цитокинины • Это вещества, необходимые для индукции деления растительных клеток. • Впервые в чистом виде фактор клеточного деления был выделен из автоклавированного препарата ДНК спермы сельди. • Это вещество было идентифицированно как 6 фурфуриламинопурин (кинетин). • Природный цитокинин из незрелых зерновок кукурузы – зеатин – в 1963 г. Получил Д. Летам.
Цитокинины • Это вещества, необходимые для индукции деления растительных клеток. • Впервые в чистом виде фактор клеточного деления был выделен из автоклавированного препарата ДНК спермы сельди. • Это вещество было идентифицированно как 6 фурфуриламинопурин (кинетин). • Природный цитокинин из незрелых зерновок кукурузы – зеатин – в 1963 г. Получил Д. Летам.
 Метаболизм и транспорт • Наиболее богаты цитокинами развивающиеся семена и плоды, а также меристематически активные участки. • Основное место синтеза цитокинина у вегетирующих растений – апикальные меристемы корней. • Из корней цитокинины пассивно транстпортируются в надземные органы по ксилеме. Все природные цитокинины – производные изопентениладенина. • Связанные цитокинины в виде риботидов, рибозидов и гликозидов представляют собой транспортные и запасные формы.
Метаболизм и транспорт • Наиболее богаты цитокинами развивающиеся семена и плоды, а также меристематически активные участки. • Основное место синтеза цитокинина у вегетирующих растений – апикальные меристемы корней. • Из корней цитокинины пассивно транстпортируются в надземные органы по ксилеме. Все природные цитокинины – производные изопентениладенина. • Связанные цитокинины в виде риботидов, рибозидов и гликозидов представляют собой транспортные и запасные формы.
 Физиология и биохимия действия • Цитокинины индуцируют деление клеток, однако это их действие наблюдается лишь в присутствии ауксина. • Обработка цитокинином вместе с ИУК побуждает дифференцированные клетки растений снова перейти к делению. • К. Мотес и сотрудники показали, что обогащение тканей цитокинином предотвращает распад хлорофилла и деградацию внутриклеточных структур у изолированных листьев. • Цитокинин необходим для нормального развития листа и для поддержания его аттрагирующей способности. • На молекулярном уровне цитокинин в комплексе со специфическим белковым рецептором усиливает активность РНК-полимеразы и матричную активность хроматина; при этом увеличиваются количество полирибосом и синтез белков, в том числе некоторых ферментов, в частности нитратредуктазы. • Цитокинин действует на транспорт К, Н и Са.
Физиология и биохимия действия • Цитокинины индуцируют деление клеток, однако это их действие наблюдается лишь в присутствии ауксина. • Обработка цитокинином вместе с ИУК побуждает дифференцированные клетки растений снова перейти к делению. • К. Мотес и сотрудники показали, что обогащение тканей цитокинином предотвращает распад хлорофилла и деградацию внутриклеточных структур у изолированных листьев. • Цитокинин необходим для нормального развития листа и для поддержания его аттрагирующей способности. • На молекулярном уровне цитокинин в комплексе со специфическим белковым рецептором усиливает активность РНК-полимеразы и матричную активность хроматина; при этом увеличиваются количество полирибосом и синтез белков, в том числе некоторых ферментов, в частности нитратредуктазы. • Цитокинин действует на транспорт К, Н и Са.
 АБК • В 1961 г. В. Лью и Х. Карнс из сухих зрелых коробочек хлопчатника выделили в кристаллическом виде вещество, ускоряющее опадение листьев, и назвали его абсцизином. • Молекулярная структура абсцизина (абсцизовой кислоты) была установлена в 1963 г. Одновременно Окумой и др. и Корнфортом и др.
АБК • В 1961 г. В. Лью и Х. Карнс из сухих зрелых коробочек хлопчатника выделили в кристаллическом виде вещество, ускоряющее опадение листьев, и назвали его абсцизином. • Молекулярная структура абсцизина (абсцизовой кислоты) была установлена в 1963 г. Одновременно Окумой и др. и Корнфортом и др.
 Метаболизм и транспорт • Богаты АБК старые листья, зрелые плоды, покоящиеся почки и семена. • Синтезируется главным образом в листьях, а также в корневом чехлике. • В растительных тканях найдена связанная форма АБК – сложный эфир абсцизовой кислоты и D-глюкозы. • Перемещение АБК в растениях наблюдается как в базипетальном, так и в акропетальном направлениях в составе ксилемного и флоэмного сока.
Метаболизм и транспорт • Богаты АБК старые листья, зрелые плоды, покоящиеся почки и семена. • Синтезируется главным образом в листьях, а также в корневом чехлике. • В растительных тканях найдена связанная форма АБК – сложный эфир абсцизовой кислоты и D-глюкозы. • Перемещение АБК в растениях наблюдается как в базипетальном, так и в акропетальном направлениях в составе ксилемного и флоэмного сока.
 Физиология и биохимия действия • В большинстве случаев АБК тормозит рост растений. • Этот фитогормон может выступать антагонистом ИУК, цитокинина и гиббереллинов. • В некоторых случаях Аб. К функционирует как активатор: она стимулирует развитие партенокарпических плодов у розы, удлинение гипокотиля огурца, образование корней у черенков фасоли. • АБК – сильный ингибитор прорастания семян и роста почек и накапливается в них при переходе в состояние физиологического покоя. • АБК ускоряет распад нуклеиновых кислот, белков, хлорофилла. • АБК участвует в механизмах стресса. • Абсцизовая кислота может ингибировать синтез ДНК, РНК и белков.
Физиология и биохимия действия • В большинстве случаев АБК тормозит рост растений. • Этот фитогормон может выступать антагонистом ИУК, цитокинина и гиббереллинов. • В некоторых случаях Аб. К функционирует как активатор: она стимулирует развитие партенокарпических плодов у розы, удлинение гипокотиля огурца, образование корней у черенков фасоли. • АБК – сильный ингибитор прорастания семян и роста почек и накапливается в них при переходе в состояние физиологического покоя. • АБК ускоряет распад нуклеиновых кислот, белков, хлорофилла. • АБК участвует в механизмах стресса. • Абсцизовая кислота может ингибировать синтез ДНК, РНК и белков.
 Этилен • Обладает сильным морфогенетическим действием на растения. • Впервые физиологический эффект этилена на растения был описан Д. Н. Нелюбовым (1901), который обнаружил, что у этиолированных проростков гороха этилен вызывает «тройную реакцию» стебля: ингибирование растяжения, утолщение и горизонтальную ориентацию. • В 20 -х годах было показано, что этилен способен ускорять созревание плодов.
Этилен • Обладает сильным морфогенетическим действием на растения. • Впервые физиологический эффект этилена на растения был описан Д. Н. Нелюбовым (1901), который обнаружил, что у этиолированных проростков гороха этилен вызывает «тройную реакцию» стебля: ингибирование растяжения, утолщение и горизонтальную ориентацию. • В 20 -х годах было показано, что этилен способен ускорять созревание плодов.
 Метаболизм и транспорт • Наибольшая скорость синтеза этилена наблюдается в стареющих листьях и в созревающих плодах. • Выделение этилена растениями тормозится недостатком кислорода (кроме риса) и может регулироваться светом. • У высших растений этилен синтезируется из метионина. • Концентрация этилена в тканях контролируется скоростью его синтеза. • Газ свободно диффундирует по межклетникам в окружающую среду.
Метаболизм и транспорт • Наибольшая скорость синтеза этилена наблюдается в стареющих листьях и в созревающих плодах. • Выделение этилена растениями тормозится недостатком кислорода (кроме риса) и может регулироваться светом. • У высших растений этилен синтезируется из метионина. • Концентрация этилена в тканях контролируется скоростью его синтеза. • Газ свободно диффундирует по межклетникам в окружающую среду.
 Физиология и биохимия действия • Этилен ингибирует удлинение проростков, останавливает рост листьев (у двудольных) и вызывает задержку митозов. Все эти явления устраняются повышенными концентрациями СО 2. • Обработка этиленом индуцирует корнеобразование на стебле. • У некоторых растений этилен вызывает эпинастию (опускание) листьев. В то же время у многих видов он ускоряет прорастание пыльцы, семян, клубней и луковиц. • Механизм действия этилена изучен недостаточно. Возможно, он влияет на состояние цитоскелета на взаимосвязь мембран, микротрубочек и микрофиламентов.
Физиология и биохимия действия • Этилен ингибирует удлинение проростков, останавливает рост листьев (у двудольных) и вызывает задержку митозов. Все эти явления устраняются повышенными концентрациями СО 2. • Обработка этиленом индуцирует корнеобразование на стебле. • У некоторых растений этилен вызывает эпинастию (опускание) листьев. В то же время у многих видов он ускоряет прорастание пыльцы, семян, клубней и луковиц. • Механизм действия этилена изучен недостаточно. Возможно, он влияет на состояние цитоскелета на взаимосвязь мембран, микротрубочек и микрофиламентов.
 ВЛИЯНИЕ ЭТИЛЕНА В РАЗЛИЧНЫХ КОНЦЕНТРАЦИЯХ НА РОСТ ЭТИОЛИРОВАННЫ Х ПОБЕГОВ ГОРОХА ЗА 48 Ч ОБРАБОТКИ
ВЛИЯНИЕ ЭТИЛЕНА В РАЗЛИЧНЫХ КОНЦЕНТРАЦИЯХ НА РОСТ ЭТИОЛИРОВАННЫ Х ПОБЕГОВ ГОРОХА ЗА 48 Ч ОБРАБОТКИ
present5.com
Персональный сайт - Цитокинины
Цитокинины - гормоны корневого апекса. История открытия.
Первые стерильные культуры клеток животных удалось получить еще в начале ХХ века, однако вплоть до 1950-х годов получить устойчиво растущие культуры растительных клеток на искусственных питательных средах не удавалось.
Американец Ф.Скуг работал над проблемой получения стерильных культур клеток растений. Он работал в университете штата Висконсин поэтому его излюбленным объектом был один из сортов табака, который выращивали в этом штате - "Wisconsin-38". Из стеблей добывали сердцевинную паренхиму, помещали на искусственные среды, содержащие минеральные вещества, сахар, витамины, аминокислоты. Однако, клетки не делились. После открытия ауксина Скуг добавил ИУК в среды: вдруг растительным клеткам не хватает ауксина? Клетки паренхимы приступали к делениям, но рост быстро останавливался.
В лаборатории Скуга перепробовали самые различные добавки: экстракт дрожжей, томатный сок, сок листьев самого табака, и все безрезультатно. Небольшого успеха удалось добиться, добавив в среду кокосовое молоко. В 1950-х годах ученые обнаружили, что наследственная информация передается от родителей к потомкам с помощью ДНК. (Как устроено это вещество и какие свойства позволяют передать информацию тогда еще не было известно). И Скуг предположил, что растительным клеткам не хватает ДНК. В лаборатории появилась банка с ДНК, добытой из молок сельди (самый дешевый источник ДНК) и ее добавляли в среду наряду с ауксинами и питательными веществами. Клетки не делились.
Открытие цитокининов состоялось благодаря ошибке, допущенной в ходе эксперимента. Чтобы среда была стерильной, флаконы со средой помещали в автоклав, где раствор нагревается выше 100╟С. Случайно режим автоклавирования был нарушен и среда вместе с ДНК перегрелась. Именно на этой среде сердцевинная паренхима табака сорта "Wisconsin-38" начала интенсивно расти. При более детальной проверке выяснилось, что в перегретом препарате ДНК из молок сельди появлялось некоторое вещество, которое на фоне ауксина вызывает активные деления клеток. В 1953 году вышла первая статья, посвященная этому явлению, а затем удалось идентифицировать вещество, вызывающее рост клеток - фурфуриладенин. По физиологическому эффекту это вещество получило тривиальное название кинетин (от греч. "Кинезис" - "деление").
Вскоре из эндосперма кукурузы удалось выделить природное вещество, которое заставляет делиться клетки в организме растения - зеатин (Zea- кукуруза). Так было положено начало открытию новой группы растительных гормонов - цитокининов.
Выяснилось также, что природные цитокинины легко разрушаются при нагревании, что объясняет первые неудачи Скуга. В растительных экстрактах содержались природные цитокинины, но поскольку среды обязательно автоклавировали, эти гормоны разрушались и не оказывали действия на клетки.
Биосинтез и инактивация цитокининов. Биосинтез и инактивация цитокининов. В клетке цитокинины образуются из азотистого основания аденина. К аденину присоединяется боковая изопентильная группа, что приводит к образованию цитокининового скелета. Далее происходит последовательное удаление фосфатной группы и рибозы. Самый простой из цитокининов - изопентениладенин. Изопентениладенин проявляет большую физиологическую активность, чем изопентениладенинрибозид или изопентенил-АМФ. Другие цитокинины образуются за счет модификации изопентильного фрагмента (гидроксилирование, окисление, восстановление).
Цитокинины в клетке присутствуют в активной и неактивной форме. Неактивными формами цитокининов обычно являются N7- и N9-гликозиды, а также гликозиды по гидроксилу изопентильного фрагмента. Присоединяя или отсоединяя сахара, клетка регулирует концентрацию активных цитокининов. Возможно и необратимое разрушение цитокининов, поэтому по мере удаления от апекса корня концентрация цитокининов падает.
Из арабидопсис и кукурузы выделены гены, белковые продукты которых связываются с цитокинином и имеют характерную для рецепторов структуру. Так, у белка CRE 1 (от cytokinine receptor) есть фрагмент, выступающий на наружную поверхность плазмалеммы, который связывается с молекулой цитокинина. Далее в белке расположен гистидин-киназный домен и два домена, участвующие в переносе фосфатной группы (REC -Receiver domains). Предполагают, что рецептор цитокинина взаимодействует с МАР-киназной системой трансдукции сигнала.
Цитокинины способствуют синтезу новой ДНК в клетке и контролируют S-фазу клеточного цикла у растительных клеток. Интересно, что аденин с заместителями, похожими на радикал цитокининов, входят в состав некоторых РНК. Более того, если брать синтетические аналоги цитокининов (например, бензиламинопурин), то необычный радикал (бензил) появляется в тех же самых РНК у того же самого аденина. Эксперименты с мечеными атомами показывают, что цитокинин непосредственно не встраивается в молекулу РНК целиком. Происходит лишь "переброска" радикала с молекулы цитокинина на молекулу РНК.
Эффекты цитокининов от апекса корня до апекса побега. Цитокинины оказались во многом похожими на первую из известных групп растительных гормонов - на ауксины, однако были и существенные отличия. Главное - у цитокининов совершенно иная точка синтеза. Если ауксины синтезируются в апексе побега, то цитокинины - биохимический "маркер" кончика корня. Ауксин транспортируется по растению сверху вниз и активно, а цитокинин - снизу вверх и пассивно.
Образно цитокинины можно назвать " гормонами благополучия апекса корня". Аттрагирующий эффект. Кончик корня для своего роста нуждается в питательных веществах. Минеральных солей и воды у корня в достатке, поэтому необходимо "притягивать" продукты фотосинтеза: сахара, аминокислоты и др. Этот эффект проявляется в зоне деления (т.е. в апикальной меристеме) корня.
Иногда цитокинины называют гормонами "омоложения" растительных тканей. Если обработать цитокинином лист, готовящийся к листопаду, он еще долго будет оставаться зеленым. Этот эффект был настолько впечатляющим, что физиологи растений считали "омоложение" - главным эффектом цитокининов. Однако при более внимательном рассмотрении окажется, что это - всего лишь аттрагирующий эффект. В норме перед листопадом все белковые молекулы листа разрушаются и в виде аминокислот отправляются на зимнее хранение в корень. К моменту листопада в тканях листа почти не остается азота, который был бы доступен метаболизму. Оттекают из осеннего листа и другие органические вещества, следовательно, мы имеем дело не столько со старением, сколько с запрограммированной гибелью листа от истощения (заметим, что вечнозеленые листья тропических растений обычно остаются живыми в 3-5 раз дольше, чем листья растений умеренных широт) . Зимой большая листовая поверхность опасна, поэтому происходит листопад, а питательные вещества депонируются в корне.
Если ввести в лист радиоактивный глицин и обработать одну из частей листа цитокинином, метка быстро соберется в обработанной половине листа. Нормальная физиологическая реакция состоит в том, чтобы передать питательные вещества ближе к корню (т.е. источнику цитокининов). Так как в эксперименте источником цитокининов оказалась соседняя половина листа, метка переместилась именно туда.
Таким образом, цитокинин не омолаживает лист, а просто не дает ему погибнуть от истощения, притягивая и удерживая в тканях питательные вещества.
Цитокинин и дифференцировка клеток. В зоне дифференцировки корня цитокинины способствуют образованию проводящей системы. Поскольку корень нуждается в продуктах фотосинтеза, которые по растению разносит флоэма, цитокинины (гормоны корневого благополучия) вызывают образование преимущественно элементов флоэмы.
Цитокинин рассматривается тканями как запрос на фотоассимиляты. Если ткань в принципе способна образовать хлоропласты, то даже в темноте под действием цитокинина можно добиться синтеза хлорофилла. Так, в семялодях тыквы происходит дифференцировка фотосинтезирующих тканей, в клетках пластиды превращаются в хлоропласты.
Распространяется цитокинин вверх с ксилемным током. Поскольку ксилема - мертвая ткань, она не может обеспечить ни активного транспорта, ни полярности: для этого нужны живые мембраны. В отличие от ауксинов, цитокинины транспортируются пассивно и неполярно.
О том, что цитокинины содержатся в ксилеме, догадывался еще Скуг. Если сердцевинную паренхиму стебля не отделять от слоя ксилемы, то некоторое время клетки паренхимы могли делиться (это наблюдение было сделано еще до открытия цитокининов).
В зоне вторичного утолщения цитокинины стимулируют работу камбия и образование новых флоэмных элементов. Повышенная концентрация цитокининов говорит растению о благополучном развитии корневой системы. Это означает, что нет необходимости в новых корнях. Т.е. цитокинины подавляют рост боковых корней. С другой стороны, нужны побеги, которые образуют новые листья и позволят лучше снабжать растущие корни. Под действием цитокининов начинают расти боковые почки на побегах. Таким образом, цитокинины снимают апикальное доминирование, вызванное ауксинами. Заметим, что ауксины и цитокинины - антагонисты в процессе регуляции развития боковых почек. Однако, в другом процессе - клеточных делений - они синергисты (т.е. их совместное действие усиливаеся). Традиционно упоминается реакция устьичных клеток на цитокинин: если вода поступает в лист из корня (т.е. обогащена цитокининами), устьица открываются. Если вода поступает из других органов, она бедна цитокининами (богата абсцизовой кислотой) и происходит закрывание.
Цитокинины способствуют росту бессемянных плодов. Ситуация очень напоминает соответствующий эффект ауксина. В молодом зародыше очень рано появляется корневой полюс, который начинает синтезировать цитокинины. Плод с семенами, естественно, содержит больше цитокининов, чем бессемянный. При добавлении цитокининов извне, растение считает, что в бессемянном плоде зародыши есть и проявляется аттрагирующий эффект.
ВЗАИМОДЕЙСТВИЕ АУКСИНОВ И ЦИТОКИНИНОВ.
Физиологическое действие ауксинов и цитокининов в культуре in vitro. Открытие цитокининов положило начало эре культивирования растительных клеток in vitro. Первым типом ткани, полученным из сердцевинной паренхимы табака, был каллус. В природе каллусы образуются в местах повреждения, когда растению необходимо как можно быстрее зарастить клетками шрам, заполняя его бесформенной недифференцированной массой клеток. Лишь позднее происходит дифференцировка и восстанавливаются поврежденные сосуды, покровные и механические ткани.
Клетки каллуса, в отличие от клеток апикальных меристем или камбия, делятся, располагая веретено деления в случайном направлении. В результате получается рыхлая быстро растущая клеточная масса.
Для того, чтобы клетки быстро размножались in vitro в среду нужно добавить и ауксины, и цитокинины. Только в этом случае растительные клетки начинают делиться. Показано, что ауксины активируют СDK-протеинкиназы клеточного цикла (cycline dependent kinases), а цитокинины - соответствующие циклины. Комплекс СDK-циклин необходим для запуска клеточного деления.
Изменение соотношения ауксин/цитокинин в среде приводит к существенным изменениям в развитии клеток in vitro. При преобладании ауксинов (недостатке цитокининов) начинается процесс ризогенеза (от греч. "rhizo-" - "корень"; "genesis" - "рождение"). При преобладании цитокининов (недостатке ауксинов) образуются меристемы побегов: начинается геммагенез ("gemma" - "почка растения"). Такое поведение культур клеток хорошо согласуется с функцией ауксинов и цитокининов как "гормонов благополучия" побегов и корней соответственно. Недостаток ауксинов воспринимается клетками как недостаточное развити побегов, и служит сигналом для их образования. В дифференцированных побегах происходит синтез ауксинов и баланс гормонов восстанавливается. Аналогичный механизм срабатывает при недостатке цитокининов (формируются корни).
При удалении из среды и ауксинов, и цитокининов часто в культуре клеток начинается образование биполярных структур - зародышей. У каждого из них будет свой источник цитокининов (корневой полюс) и свой источник аусинов (побеговый полюс). Такие структуры, похожие на зародыши семян, называют эмбриоидами ("embryo" - "зародыш"; "eidos" - "похожий").
Такое поведение клеток in vitro используют для создания технологий учкоренного размножения растений.
Баланс между ауксинами и цитокининами в интактном растении. Итак, для нормальной жизни в растении всегда должен существовать баланс между ауксинами и цитокининами.
Процесс вегетативного роста растений можно моделировать, рассматривая только полярный транспорт и взаимодействие ауксинов и цитокининов. Представим, что главный стебель отделен от корня. В верхней части осталась точка синтеза ауксинов, цитокинины в дефиците. Ауксин из апекса транспортируется к основанию черенка, где создается избыток этого гормона. Чтобы сохранить гормональный баланс, нужны цитокинины: в основании закладываются придаточные корни.
У оставшейся нижней части растения есть точки синтеза цитокининов, а ауксинов не хватает. Цитокинины транспортируются наверх и накапливаются около среза. Это также вызывает нарушение гормонального баланса, что приводит к активации синтеза ауксинов. Пробуждаются спящие почки или даже возникают новые из каллусной массы.
В интактном растений активные деления клеток сосредоточены на кончике корня и на кончике побега. Согласно модели, чем дальше находится орган от кончика корня, тем меньше в нем цитокинина. Апексом побега транспорт заканчивается и происходит накопление цитокинина. Ауксин сам синтезируется в апексе побега. Таким обрзом в меристеме побега концентрация цитокининов и ауксинов оказывается достаточной для поддержания клеточных делений.
Обратная ситуация наблюдается в апексе корней: ауксин накапливается т.к. апекс корня является конечным пунктом транспорта. Вместе с синтезируемыми в кончике корня цитокининами, ауксины вызывают клеточные деления в меристеме корня.
Чем больше расстояние от кончика корня до верхушки побега преодолевают гормоны, тем меньше их содержание в меристеме. Наконец, наступает момент, когда цитокинины перестают поступать в апекс побега в нужной для делений концентрации. Рост стебля вверх останавливается. Аналогично регулируется рост корня вглубь. Таким образом, полярный рост растения можно моделировать на основании баланса ауксинов и цитокининов.
В зоне вторичного утолщения делящиеся клетки расположены не случайно. По флоэме вниз перемещается ауксин, а по ксилеме вверх - цитокинин. Между этими ткаными лежит камбиальное кольцо (слой делящихся клеток). Здесь клетки находятся на "перекрестке" потоков ауксинов и цитокининов (а также фотоассимилятов и минеральных веществ), необходимых для деления клеток.
Работу камбия также можно смоделировать исходя из теории поддержания баланса фитогормонов. Если ауксин преобладает, камбий откладывает больше ксилемных элементов (чтобы обеспечить приток цитокининов из корней). Наоборот, преобладание цитокининов заставляет камбий откладывать больше флоэмы. По ней поступят новые порции ауксина и снова возникнет равновесие между двумя типами гормонов.
Заметим, что поврежденные корни или ветки гораздо хуже утолщаются. Причина этого - в низкой способности этих органов к синтезу гормонов. С другой стороны, чем больший поток гормонов проходит через ветку или корень, тем сильнее утолщение, больше "транспортных магистралей" и больше механическая прочность.
Цитокинины и паразиты растений. Грибы и бактерии за миллионы лет совместного существования с растениями гораздо лучше усвоили физиологию растений, чем человек. И у этих организмов возникли блестящие приспособления для "эксплуатации" организма растений как источника пищи. Ведь достаточно "засесть" в любом органе растения и начать подавать запросы на питательные вещества. Растение "не сообразит" и направит продукты фотосинтеза вместе с минеральными веществами к месту инфекции, чего и надо бактериям и грибам-паразитам.
Многие из грибов, вызывающих заболевания растений, научились вырабатывать цитокинины. В местах поражения возникает опухоль, из которой во все стороны растут многочисленные тонкие побеги. В народе такую структуру прозвали "ведьминой метлой".
Аналогичные механизмы используют и бактерии. Псевдомонады (Pseudomonas savastanoi) способны синтезировать не только цитокинин, но и ауксин. Они поражают сирень, бирючину, оливу и другие маслинные растения. Пораженная псевдомонадами ткань начинает интенсивно делиться, образуя каллус, и питательные вещества со своего растения "притягиваются" к месту инфекции. А это и нужно псевдомонадам. Но если псевдомонад убить антибиотиком, то рост опухоли останавливается.
Самый тонкий и специализированный механизм нарушения гормонального баланса имеют агробактерии (Agrobacterium tumefaciens). Клетки этих бактерий умеют передавать свою ДНК в ядра растительных клеток. В передаваемом фрагмента ДНК содержится информация о биосинтезе ауксинов, цитокининов и особых веществ - опинов. Опины не могут утилизироваться клетками растения, но служат источником углерода и азота для роста бактерий. Клетки растений, в которые попала такая ДНК, начинают опухолевый рост. Даже при уничтожении бактерий (обработка антибиотиками), опухоль продолжает расти, т.к. клетки продолжают производить ауксины и цитокинины за счет встроенных бактериальных генов.
regrosta.ucoz.ru
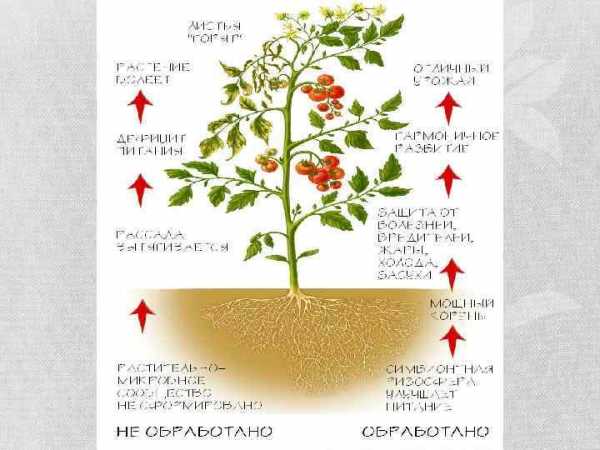
5. Семенная кожура семян у цветковых растений образуется из: а) зиготы б) центральной Первопричиной затруднения роста растений на почвах с высокой концентрацией солей Основное место синтеза гормона цитокинина у вегетирующих растений –верхушечная меристема.
Чтобы посмотреть этот PDF файл с форматированием и разметкой, скачайте его и откройте на своем компьютере.вопрос варианта Необходимо клетках отсутствуют вакуоли Имеется полость тела плоских червей первичная полость полость смешанная полость имеется неполным превращением относятся двукрылые полужесткокрылые стрекозы каких состоит грудная клетка амфибий грудины грудины позвонков грудной клетки млекопитающим Голарктической зоогеографической области относятся утконос волк сурок косуля ак верблюд суслик ягуар выхухоль 15. происходит переваривание пищи кишечной кишечной кишечной клетках Рабочие являются отложившими приступившими уходу самками отложить следующий Кольчатые черви отличаются круглых червей наличием выделительной системы системы варительной системы Сонную болезнь плазмодии 19. приведенных характеристик колбочек выберите неправильную осуществляют наибольшая плотность желтом пятне 20. Воздух пузырьках время сравнению наружным воздухом больше углекислого лекислого углекислого отличается Инсулин снижает уровень глюкозы крови счет обеспечивающих окисление выведения глюкозы организма активации клетки кровь поступает сердце через легочную вены легочные правильную последовательность процессов приводящих свертыванию крови разрушение появление сети белковых образование разрушение появление белковых появление разрушение желудка расщепляют жиры углеводы жиры углеводы жиры рганизме человекагемоглобина печени железа кишечнике Заболевание которое характерно жителей штатов США питающихся основном кукурузой дефицитом
енетический код это нуклеотидная последовательность хромосоме последовательностью нуклеотидов гене последовательностью белке клеток предоставить наи
docs4all.com