
Оптимизация минерального питания – значительный резерв в растениеводстве. Оптимизация питания растений
Фундамент оптимизации минерального питания растений
Мы можем в 4 раза увеличить урожай, если нам удастся вдвое ускорить физиологические процессы», – авторитетно заявляют современные ученые. И это действительно возможно. Сегодня они готовы открыть для каждого из нас эти секреты, именно те моменты, которые помогут управлять процессами ассимиляции, через рассматриваемые нами факторы. Одним, из которых является наиболее действенный и проверенный временем – это оптимизация питания растений.
Хотя для большинства, и даже опытных агрономов, понятие оптимизация питания почему-то ассоциируется как дополнительные капиталовложения, мы же рассматриваем – как поиск такого варианта, который позволяет правильно и грамотно использовать свет, воду, тепло, пищу, максимально использовать генетический потенциал растения. Как снизить затраты и избежать недобора урожая, повысить продуктивность растения в условиях неблагоприятного воздействия климатических условий и других негативных факторов – вопросы, волнующие каждого. Реализация этих замыслов возможна за грамотного и творческого подхода к разработке системы питания, что является целенаправленным влиянием элементов питания на биохимические и физиологические процессы, в частности: рост, развитие и, соответственно, уровень урожайности. Минеральные соединения при этом является не только строительным материалом, но и регуляторами соответствующих процессов, составной частью ферментов и аминокислот.
 Фундамент оптимизации питания растений заложил еще в середине XIX века немецкий профессор Юстус фон Либих, который первым сформулировал правило, которое сегодня широко известно под названием Закон минимума Либиха. Это правило расширил американский ученый Виктор Шелфорд, благодаря чему оно и получило название Закон толерантности Шелфорда. Корифей отечественной агрохимии Дмитрий Прянишников убедительно доказал, что недостаток знаний нельзя заменить избытком удобрений. Этот постулат особенно актуален сегодня, когда агрономы изыскивают возможность как обеспечить полноценное минеральное питание и при этом сэкономить средства на приобретение удобрений, не рискуя жертвовать показателями урожайности.
Фундамент оптимизации питания растений заложил еще в середине XIX века немецкий профессор Юстус фон Либих, который первым сформулировал правило, которое сегодня широко известно под названием Закон минимума Либиха. Это правило расширил американский ученый Виктор Шелфорд, благодаря чему оно и получило название Закон толерантности Шелфорда. Корифей отечественной агрохимии Дмитрий Прянишников убедительно доказал, что недостаток знаний нельзя заменить избытком удобрений. Этот постулат особенно актуален сегодня, когда агрономы изыскивают возможность как обеспечить полноценное минеральное питание и при этом сэкономить средства на приобретение удобрений, не рискуя жертвовать показателями урожайности. Таким образом, оптимизация питания – это подбор, установка и применение таких приемов, как оптимальные уровни, соотношения и формы элементов минерального питания, способы внесения удобрений для конкретных почвенно-климатических условий. Также учтены физиологические и сортовые особенности растений, приемы агрономических технологий и ряд других факторов. В тесном взаимодействии между собой эти факторы определяют величину урожая. Вместо этого главным условием выживания в современном земледелии является умение получить высокие и устойчивые урожаи с минимальной себестоимостью единицы продукции.
Придерживаться этих принципов – главная задача каждого агронома, технолога и руководителя. Сортовой состав постоянно меняется – появляются новые сорта растений, гибриды, растут требования к производительности сельхозкультур и качества продукции при таких условиях, как рискованное земледелие. И на первый план из всех агроприемов получается минеральное питание растений, ведь при рациональном подборе элементов можно корректировать, как минеральное корневое питание так и фотосинтез. Семена, минеральные удобрения, микроудобрения, стимуляторы роста, средства защиты растений (гербициды, инсектициды, фунгициды) является неотъемлемой частью аграрных технологий.
Следовательно, оптимизация питания – это наиболее благоприятное для растений сочетание воздушного и почвенного питания и экономически выгодный для предприятия вариант их обеспечения. Сделать правильный выбор помогут специалисты Ecowonder.org, которые понимают необходимость и стремятся обеспечить сочетание науки с производством.
ecowonder.org
ОПТИМИЗАЦИЯ УСЛОВИЙ ПИТАНИЯ ТЕПЛИЧНЫХ РАСТЕНИЙ — КиберПедия
Основой оптимизации питания тепличных культур, — овощных, цветочных и других, — является, с одной стороны, использование стандартных по периодам выращивания растворов с соответствующими сбалансированными соотношениями макро- и микроэлементов, уровнями ЕС, рН, поддержание необходимых условий микроклимата (температура, влажность субстрата и воздуха, освещённость, подкормка углекислотой, защита растений от вредителей и болезней.
В процессе выращивания тепличных культур постоянно возникают явления, которые необходимо учитывать для оптимизации питания:
— антагонизм элементов питания в связи с фактической концентрациейотдельных элементов питания в субстратном растворе, вследствие чего нарушается усвоение растениями отдельных элементов питания, несмотря на использование сбалансированных питательных растворов;
— нарушение питания в связи с недостатком или избытком элементов,необходимых для правильного роста и развития растений, что имеет местопри неблагоприятных агротехнических условиях, или вследствие недостатка-избытка элементов питания.
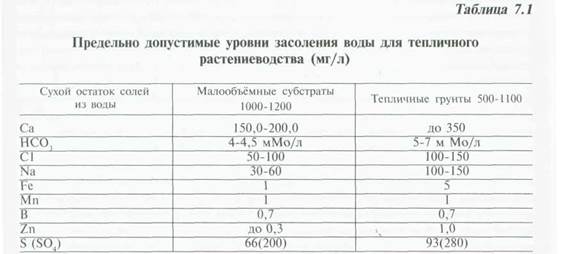
Рассмотрим более подробно факторы питания и методы регулирования условий питания растений, обеспечивающих оптимизацию условий выращивания. Прежде всего, необходимо в течение всей вегетации иметь достоверную, часто меняющуюся в период вегетации, информацию о химическом составе воды. Повторные анализы проводят 1 раз в 2—3 месяца, особенно если это вода из открытых водоёмов. Анализ проводят по следующим показателям: рН , HCO3,Na, Cl, Nh5, NO3, К, Са, Mg, Fe, Mn, Zn, В, Си.
Для малообъёмного выращивания в отдельных регионах, вода без дополнительной доработки непригодна для непосредственного использования, если имеют место следующие параметры: ЕС — 1-1,5 мСм/см, Na — 70-100 мг/л, С1 — 100—160 мг/л и более высокое.
Допустимые предельные уровни элементов в воде для приготовления рабочих растворов удобрений должны быть в следующих максимальных пределах или менее их (мг/л) (табл. 7.1)
Рассмотрим проблемы корректировки питательных растворов с учётом анализа воды.
При малообъёмном методе выращивания необходимо держать под посто-
янным контролем буферность воды и дренажа, т. е. содержание свободных ионов НСО3 ', общее количество которых в растворах не должно превышать суммы ионов Са+2 и Mg+2, обычная норма гидрокарбонатов 0,5-1 мМо/л.
Необходимо учитывать жёсткость воды — общее содержание солей Са(НСО3)2, Mg(HCO3)2, СаС12, MgCl2, CaSO4, MgSO4. Г жёсткости означает концентрацию катионов Са и Mg, эквивалентную 10 мг/л СаО.
Содержание ионов Са и Mg в используемой воде должно быть ниже расчётного количества этих ионов в рабочем растворе, в противном случае нарушается оптимальное соотношение ионов К и Са +Mg проявляется их антагонизм и сокращение поглощения К растениями.
Часто вода имеет избыток гидрокарбонатов, ионов Na, CI, Mg, S, Zn, Fe. Только избыток Mg не является токсичным; но имеет место дисбаланс элементов питания. Избыток Са, Mn, Fe, HCO3-' также создаёт дисбаланс, нарушается оптимальное соотношение этих элементов в рабочем растворе. Кроме того, избыток Cl, Mn, S может быть токсичным, т. е. дисбаланс и токсичность — результат неконтролируемого количества этих элементов. К токсичным гидрокарбонатам в воде относятся NaHCO3 и А1(НСО3)3. Вот почему гидрокарбонаты нейтрализуют частично, а иногда и полностью. При рН раствора удобрений равным 5,5, обычно остаётся в воде 1 мМо/л НСО3, при рН = 5, в воде остаётся 0,3 мМо/л и менее гидрокарбонатов.
Избыток Na в рабочих растворах более 60 мг/л опасен для тепличных культур, так как имеет место постепенное накопление Na в корневой зоне. Установлено, что концентрация Na+ 30-60 мг/л ощутимо снижает интенсивность роста томата, огурцов и других тепличных растений. Кроме того, Na — антагонист Са, Mg, К, что будет рассмотрено ниже.
Против негативного действия повышенного количества Na следует увеличивать в питательных растворах и в дренаже норму Са, Mg, К, выдерживая соотношение этих элементов.
Следующая проблема — это вода с повышенным количеством серы, S — > 60 мг/л (SO4 — > 150 мг/л). Повышенное количество серы в почвенном раст-
воре усиливает усвояемость Na и одновременно уменьшает усвояемость Са. Избыточные количества серы в воде снижают предварительной обработкой воды активным хлором (Са(НОС1)2, хлорной известью, жидким хлором). Норма активного хлора составляет 0,6 мг на 1 мг серы. Этой же нормой активного хлора дезактивируют избыточное количество Fe2 и Мп. Предварительное осаждение избыточного количества серы в воде можно осуществлять, добавляя в воду мел СаСО3 с активным перемешиванием воды (фонтанированием).
В связи с необходимостью регулировать качество воды для приготовления рабочих растворов, особенно в регионах, где используют воду с повышенными количествами в воде Са, Mg, S, Na ,C1 учитывают следующие факторы:
1) Показатель рН воды и рабочего раствора. Летом вода открытых водоёмов имеет более высокую щёлочность, чем весной и осенью. Это явление связано с деятельностью сине-зелёных водорослей в открытых водоёмах и разложением гидрокарбонатов на СО2 и ОН. Поэтому летом необходимо чаще анализировать воду из открытых водоёмов. Предварительная кислотная обработка воды в бассейнах-накопителях летом до рН = 6 — важный технологический приём подготовки воды для малообъёмного выращивания т. к. при такой кислотности предотвращается осаждение солей Са и Mg на трубах-магистралях. Жёсткость воды и осаждение солей на магистралях капельного полива и капельницах также связана с избытком Fe, Mn, Al, Zn, S.
Гидрокарбонаты воды представлены солями карбоновой кислоты — Н2СО3, которая диссоциируется в воде на Н+ и НСО3~'. Ионы НСО3' вступая в реакцию с ионами металлов, находящихся в воде, образуют следующие соли: Са (НСО3)2, Mg (HCO3)2, NaHCO3, KHCO3. Это основные соли по их количеству в воде. Кроме того, в воде могут присутствовать: Nh5HCO3 , А1(НСО3)3, Zn(HCO3)2, Cu(HCO3)2, MnHCO3 из них только NaHCO3 и А1(НСО3)3 токсичны для растений. Нейтрализация гидрокарбонатов кислотами с оставлением 0,5-1 мМо/л для создания определённой буферности раствора имеет место при показателе кислотности раствора в пределах рН = 5,5-5,3. Кислотная обработка бикарбонатов приводит к выделению в раствор ионов металлов. Усвоение этих ионов растениями имеет место при их соотношениях в растворах, не связанных с антагонистическими количествами.
Предварительная химводоочистка также необходима в водах с высокими показателями гидрокарбонатов 3,5-4 мМо/л и более (214-244 и > мг/л НСО3). Для предварительной химводоочистки можно рекомендовать установку сборной ёмкости объёмом около 55 м3 (1 шт. на 1 га), так как основное водопот-ребление имеет место в летние месяцы. Для удаления из воды ионов С1~, Fe 2+, S хорошо использовать бассейны-отстойники большой ёмкости с аэра-торными установками, где можно использовать активный С1. Это баллонный хлор или гипохлорит кальция, с содержанием активного хлора от 30 до 70%. Активный хлор нейтрализует: h3S — 1 мг на 1 мг Cl, Fe2+ — 1 мг на 0,6 мг С1, Мп — 1 мг на 0,6 мг С1.
Весьма эффективным решением является активное аэрирование воды с последующей фильтрацией её через песчано-гравийный фильтр. После этого необходимо провести анализ воды для определения рН, ЕС, количества катионов и анионов, что следует учитывать при приготовлении рабочих растворов.
В процессе выращивания необходимо регулярно контролировать коли-
чество макро- и микроэлементов в почвенном растворе, выжимке из субстрата, что позволит корректировать в нужную сторону показатели питания растений.
Одновременно следует учитывать и другие факторы, влияющие на усвоение элементов питания, а также темпы роста и развития растений.
Часто усвоение связано с неблагоприятными для культур климатическими условиями: слишком низкой или слишком высокой температурой, интенсивностью света, агротехническими условиями, в том числе недостаточным или избыточным водоснабжением, ненадлежащими удобрениями или использованием для полива воды плохого качества. Недостаток питания (фактическое отсутствие, неусвоение, ненадлежащая реакция кислотности почвенного раствора), избыток питания, неразвитая корневая система, неправильное орошение, высокие концентрации катионов и анионов, особенно Na и С1, также негативно влияют на усвоение элементов питания растениями.
Признаки нарушения нормального роста и развития растений обычно проявляются на всём растении, но чаще всего это наблюдается на листьях и плодах у овощных культур, на листьях и цветках у цветочных культур. На листьях это проявляется изменением окраски, деформацией, уменьшением размеров, пятнистостью и некрозами листьев, целиком или частично и их дальнейшим засыханием, пожелтением, побурением краев листовых пластинок, деформацией цветков и соцветий; обесцвечиванием, пятнистостью, некротическим растрескиванием, пятнами на плодах томатов, огурцов и других тепличных культур.
Признаки недостатка питательных веществ могут появляться на разных частях растений: на молодых и старых листьях, точках роста. Признаки недостатка элементов питания на молодых листьях и конусах роста чаще информируют о недостатке В, Са, Си, Fe, Mn, Zn, т.е. кальция и микроэлементов, а на старых листьях — N, Mg, К, S, Мо. В условиях использования полноценных по составу питательных растворов разные пятнистости, некрозы, хлорозы свидетельствуют о невозможности их усвоения — как реакция на низкую или высокую температуру субстрата, слабое развитие корневой системы, либо на фитотоксичность химических средств защиты или питания. Пожелтение нижних листьев томатов, (когда главная жилка листа зелёная), — это обычное проявление недостатка магния, что может быть связано с условиями повышенной потребности растений (но и не только!) в магнии, недостаточной освещённостью, неправильным (обычно недостаточным) во-дополивом, высокой ночной температурой, избытком калия (антагонистом магния), недостаточной аэрацией. Это часто связано с антагонизмом между калием и магнием, вследствие чего имеет место не только магниевый хлороз, но и опадание листьев без внешних признаков хлороза, например, на розах. А у томатов на самых молодых листьях темно-зелёная окраска — признак избытка азота и недостатка йода, а светло-зелёная окраска свидетельствует о недостатке азота. Побурение и отмирание верхушечных листьев у томатов вызывается недостатком фосфора.
Антоциановый оттенок нижней части листа томатов, роз — результат недостатка фосфора из-за слишком низкой температуры субстрата, избытка азота или серы, слабой корневой системы, слишком высокого, — (рН- более 6,2),
— показателя кислотности почвенного раствора. Пятнистость листьев и усы-хание их краёв происходит при недостатке калия. Скручивание молодых листьев томатов и других культур связано с недостатком марганца и меди, высоким показателем рН почвенного раствора (щелочная реакция вместо обычной срН 5,3-5,8). Изменение конуса нарастания связано с недостатком кальция ибора при слишком высоком показателе рН почвенного раствора — более 6,5.
Тепличные растения отличаются очень разной способностью поглощения и усвоения азота и калия. У томатов имеет место более трудное поступление фосфора в растения. Часто это не исправляет повышение доз фосфора, а скорее улучшается при сбалансированном количестве N, Р, К, Са, Mg в почвенном растворе и поддерживанием кислотности на уровне рН 5-6. У томатов поглощение Са и Mg в течение вегетации обычно равномерное, а потребление N систематически возрастает до пика плодоношения. Недостаточное питание приводит к нарушениям роста: недостаток N — к медленному росту корней, К
— к снижению жизнеспособности растений, Mg — к неблагоприятным физиологическим изменениям, Са — к слабой корневой системе и тонким побегам, Fe — к задержке роста, В — к растрескиванию листьев, хрупкости побега,сбрасыванию завязей. Избыточное питание также влияет отрицательно, усиливается антагонизм между усвоением растениями ионов. Этому способствуетнеравномерное потребление ионов и их накопление в почвенном растворе.
Необходим не только постоянный мониторинг элементов питания в почвенном растворе и в почве (субстрате), но и осуществление мероприятий по поддержанию необходимых уровней и соотношения элементов питания. Это достигается кратковременной корректировкой питательного раствора, более широким использованием дренажа.
Рассмотрим взаимодействие между элементами питания в почвенном растворе и их доступности растениям, в зависимости от их количества, факторов рН, ЕС, микроклимата.
Показатель кислотности почвенного раствора подлежит постоянному контролю и корректировке, в связи с сильным влиянием рН на доступность многих элементов питания, так как при возрастающем показателе рН против оптимума, лежащего в пределах 5,1—5,9. снижается доступность таких элементов, как Р, В, Си, Fe, Mn, Zn, а в кислой среде доступность Мо. Часто причиной неправильного или недостаточного питания является не недостаток элементов питания в субстратном растворе, а невозможность их поглощения при щелочной реакции почвенного раствора, использовании воды с щелочной реакцией, высокое содержание в ней Na и С1. При рН почвенного раствора выше 7,0 в почвенной культуре микроэлементы и Р становится менее доступными, а макроэлементы — К, S, поглощаются в избыточных количествах.
На супесчаных, легко- и среднесуглинистых почвах известкование, в случае его необходимости, проводят до уровня рН 5,5-6, в связи с низким уровнем поглощающего комплекса. На суглинистых и глинистых почвах с малым содержанием гумуса известковать почву можно до показателя рН 6,5. Количество легкодоступного Са в почве должно составлять не менее 1500—2000 мг/л в кислотной вытяжке. Нельзя допускать переизвесткования, что уменьшает доступность микроэлементов. С ростом рН от 5,5 до 6,7 концентрация
доступного фосфора быстро снижается, в том числе содержание Р в листьях падает до 30% от нормы. Также снижается содержание в листьях В, Си, Мп, Zn. В зависимости от используемого субстрата показатель рН субстрата и почвенного раствора следует поддерживать до показателя рН в пределах 5,1—5,9.
Для регулирования уровня рН > 6, соль КН2РО4 в рабочем растворе заменяют ортофосфорной кислотой. Низкий показатель рН < 5 наблюдается при использовании большого количества аммония (фосфат аммония, амсе-литра). В этом случае рН рабочего раствора повышают до рН 6, а содержание Nh5 в растворе дают не более 10 мг/л. Следует в этом случае уменьшить дозу К, уменьшают разовую дозу капельного полива до 70 мл. При высоком рН несколько увеличивают дозу Nh5 до 20 мг/л, норму Fe увеличивают до 2-2,5 мг/л раствора.
При использовании малообъемных субстратов применяют систематический и многоразовый в течение дня полив растений раствором минеральных удобрений соответствующей концентрации. В зависимости от вида растений, а в пределах культуры — от вида выращивания и состава питательного раствора используют определенный уровень общей концентрации солей. Последняя связана с солеустойчивостью культуры, т. е. способностью усваивать из почвенного раствора воду и элементы питания.
Раньше солеустойчивость растений оценивали в атмосферах осмотического давления (ОД) и учитывали способность корневой системы поглощать из почвенного раствора элементы питания при определённой предельно допустимой засоленности субстрата. Сейчас в практике тепличного растениеводства оперируют понятием электропроводимости рабочего и почвенного раствора с показателем в миллисименсах на 1 см (mSm/см, мСм/см). Имеются рекомендации по средним показателям для различных культур (табл. 7.2).
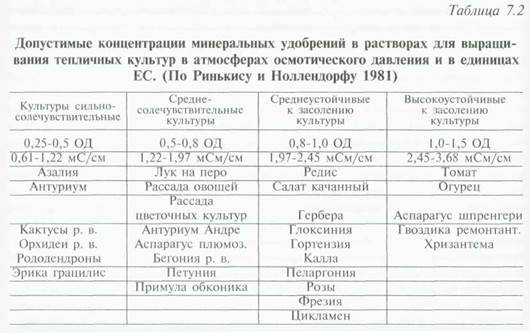
Оптимальные концентрации солей в рабочих растворах варьируют по фазам роста и развития культур, по уровням температуры и освещённости, плодовой нагрузки. Но с учётом устойчивости к засолению почвенного раствора всегда требуется контролировать и корректировать его.
У томата при малообъемном выращивании, оптимальными показателями рН и ЕС в период вегетации являются следующие:
— пропитка матов: снижение рН в дренаже до 4,8—5;ЕС питательного раствора около 2,8 мСм/см;
— установка растений на маты: рН питательного раствора 5,5;
ЕС питательного раствора — 2,7 мСм/см. Одноразовая доза полива около 200 мл раствора на одно растение;
— установка растений в отверстия для укоренения: рН питательногораствора 5,5, ЕС питательного раствора около 2,6 мСм/см;
— цветение 1-3 кисти: рН питательного раствора 5,5, ЕС питательногораствора 2,8-3 мСм/см, водопотребление 0,8—1,2 л/растение;
— цветение 4-5 кисти: рН — 5,5-5,8, ЕС — 2,6-2,8 мСм/см,водопотребление 1,2-1,8 л/растение;
— массовое плодоношение: рН — 5,5-5,8, ЕС — 2,5-2,6 мСм/см,водопотребление 1,8-2,5 л/растение и более;
— осенний период: рН — 5,5, ЕС — 2,7-2,8 мСм/см.
Для каждой культуры по периодам выращивания оптимальны определенные показатели ЕС. В период выращивания не допускают рН в дренаже выше 6,2, для чего можно использовать ортофосфорную кислоту в количестве нормы Р в растворе. На пике плодоношения томатов ЕС раствора может быть на уровне 2,8—4,2 мСм/см. Но нельзя допускать накопления солей в матах, при котором доступность катионов и анионов резко снижается, имеет место повреждение корневых волосков. Допустимое превышение показателя ЕС в почвенном растворе на 0,5 мСм/см, в сравнение с подаваемым рабочим раствором. Следует увеличить норму дренажа и несколько понизить показатель ЕС рабочего раствора. В летние месяцы норму ЕС можно снизить до 3,2 мСм/см.
Нельзя допускать рН дренажа ниже 5. Следует уменьшить количество Nh5 в рабочем растворе до 7—10 мг/л, уменьшить дозу К, рН рабочего раствора повысить до 6, за счет уменьшения количества кислоты при подщела-чивании раствора — увеличить количество Nh5 временно до 20—25 мг/л, увеличить норму хелата железа на 20%, но не более 2 мг/л.
Нормирование питания растений при малообъёмной технологии базируется на агрохимическом мониторинге дренажа, выжимки из субстрата, субстрата, в растительном материале (листовая диагностика). При проведении анализа грунта в теплицах одна смешанная проба берётся с площади не более 400 кв. м., отбирая по 1 индивидуальной пробе с каждых 100 кв. м. При малообъёмном выращивании, в процессе интенсивного роста или массового созревания концентрация элементов питания быстро изменяется, в связи с уровнем поглощения элементов питания, их вымыванием с дренажем, сорбцией и т.п. явлениями. Поэтому полный агрохимический анализ проводят каждые 3—4 недели, в период интенсивного роста — каждые 2 недели, показатели рН и ЕС — 2—3 раза в неделю.
Наиболее интенсивное поглощение воды и минеральных удобрений из почвенного раствора наблюдается при начальных показателях концентрации солей в почвенном растворе и постоянно снижается при его максимальных показателях. Поэтому в программу оптимизации питания входит использование удобрений и воды наименее засоляющих почвенный раствор.
Количество азота подлежит постоянному контролю раздельно нитратного и аммиачного азота в почвенном растворе (дренаже). Избыток азота способствует чрезмерному развитию вегетативной массы, рыхлости растений, ослаблению и запаздыванию получения продукции, завязыванию плодов у овощных растений. Чрезмерное количество азота относительно легко удаляется промыванием субстрата, правильным нормированием дренажа.
В процессе выращивания различных культур необходимо контролировать уровни соотношения различных катионов и анионов в рабочих и особенно почвенных растворах, и с помощью корректировки рабочих растворов примерное количество катионов и анионов должно быть равным.
Для каждой культуры, прежде всего, необходимо следить за соотношением — азот: калий в почвенном растворе (выжимка из субстрата, дренаж в начале его выделения), чтобы поддерживать необходимое соотношение. У молодых растений до начала завязывания плодов или бутонизации поддерживают соотношения N : К=1 : 1-1,2. По мере роста плодовой нагрузки, например у томатов, соотношение постепенно изменяются N : К < 1-1,2 до 1,5 затем до 1,8, иногда выше — до 3.
Дефицит магния наблюдается на различных тепличных растениях, но более распространён у томатов при высоких уровнях N и К. Низкий уровень азота при известковании субстрата чаще проявляется в почвенной культуре. Средний уровень N в этом случае предпочтителен. При низких уровнях К при среднем и высоком количестве азота в почвенном растворе нарушает созревание томатов. В условиях низкой освещенности количество пустотелых плодов увеличивается при чрезмерном применении фосфора, и снижается при высоких уровнях калия. Повышение уровня азота снижает содержание в листьях калия, хотя он может находиться в почвенном растворе в умеренном количестве. Увеличение количества К в растворе снижает потребление магния, т.к. проявляется антагонизм несбалансированных количеств К и Mg. Поэтому при малообъёмной культуре томата применяют следующие соотношения N : К до образования 1-го соцветия — 1 : 1,1, от 1-ой до 3-ей кисти — 1 : 1,3 , от 3-ей до 5-ой кисти 1 : 1,5, в период плодоношения — 1 : 1,8. При плохом освещении весной и осенью поддерживают более высокий уровень Mg. Повышенное количество магния в почвенном растворе не отражается отрицательно на росте растений. При повышенном количестве фосфора в почвенном растворе выше нормы сдерживается поступление магния в растение, его место занимает калий, например на розах в этом случае наблюдается у чувствительных сортов опадение листьев. Высокие концентрации аммонийного азота (норма до 10-14 мг/л) снижают поступление в растения из почвенного раствора Са и Mg. При возрастании выше допустимого количества Na и CI в почвенном растворе также увеличивается поступление в растения фосфора, калия, снижается содержание Са. С другой стороны если много хлоридов в почвенном растворе, то увеличение количества азота заметно снижает поступление хлора
в листья. Повышенные дозы фосфора в почвенном растворе снижают поступление Mn ,Zn в листья. Возрастающие количества Мп в почвенном растворе требуют повышение количества вносимого железа, чтобы соотношение Fe : Мп = 2 — 5 :1. У культур с повышенным потреблением железа (розы, гербе-ры) его количество повышают до 2,5 мг/л раствора, одновременно снижая количество Мп до 0,2-0,3 мг/л. У томата некоторые сорта требуют до 0,7 мг/л Мп в этом случае количество Fe также следует повысить до 2 мг/л. Следует учитывать, что высокие уровни Mg, Co, Zn в почвенном растворе снижают поступление Fe в растения. Постоянный контроль содержания макро- и микроэлементов в почвенном растворе — путь к оптимизации условий выращивания высокоурожайных растений.
Интенсивность усвоения элементов питания из почвенного раствора зависит и от таких факторов, как температура и освещённость, влажность воздуха и субстрата.
Низкие температуры субстрата и воздуха сдерживают рост растений и поглощение питательных элементов. Хотя увеличение уровня азота в почвенном растворе до 220 мг/л при температуре в корневой зоне 22—27°С увеличивает сырой вес плодов томата при постоянной оптимальной температуре воздуха в 20—25°С, более высокие дозы азота в корневой зоне не дают прироста урожая. При температуре субстрата 13°С и выше вес растений увеличивается при вышеуказанном уровне N, при более низкой, чем 13°С температуре высокие дозы азота не действуют положительно. По мере роста температуры необходимо повышать уровень азота, но не более 220 мг/л. Если температура возрастает, а уровень азота низкий, у томатов наблюдается сбрасывание цветков. Низкая температура субстрата (8°С и ниже) не способствует транспортировке N и К в надземную часть растений, ведёт к накоплению их в корневой системе. Рост растений замедляется. При 10—13°С в зоне корней замедляется поступление фосфора в растение. Поступление К, Са, Mg снижается при температуре в зоне корней в 13°С, а по мере роста температуры в корневой зоне увеличивается поступление в листья Р, К, Mg, Си, Fe, Mn. Решение этой проблемы в устройстве подсубстратного обогрева, что позволяет несколько снижать температуру воздуха в теплице, экономя энергетические затраты. Эта система "тёплые ноги и холодная голова" применяется при недостаточном уровне обогрева теплицы по техническим или другим причинам. С другой стороны высокие температуры воздуха ночью (2 ГС и более) увеличивают поступление в листья Са, Na, но уменьшают уровень фосфора в листьях.
Использование экранов для защиты теплиц от перегрева летом может дать сильное затенение (до 65%), что приводит к снижению поглощения N, Р, К, Са, Mg, при условии, что азотное питание на 90—95% ведётся за счёт NO3. Досвечивание растений в осенне-зимне-весенний период увеличивает поглощение N, К, Са, Mg, Mn. Очень важен в это время сбалансированный уровень К в субстратном растворе.
Длительная искусственная освещённость в условиях зимних коротких дней, при высоком уровне кальция в почвенном растворе, даёт эффект снижения сухого веса растений, а по мере увеличения длины дня весной увеличивается накопление сухого вещества.
Поглощение N и К увеличивается от низкого уровня ночью, до максимального днём, снова снижаясь в течение вечера и ночи. Поэтому необходимо брать пробу почвенного раствора на анализ в 13—14 часов дня. Поглощение N и К тесно связано не только с уровнем освещённости и температурой воздуха, но и с увеличением количества потребляемой воды.
Влажность воздуха так же важный фактор усвоения из почвенного раствора катионов и анионов. Содержание Са в молодых листьях при относительной влажности воздуха 95% значительно ниже, чем при 50%, так как поступающий кальций находится в транспирационном потоке.
Проявление вершинной гнили томатов усиливается даже при низкой влажности, при высоком уровне ЕС почвенного раствора. Кроме того, повышенная высокая влажность ночью благоприятствует движению Са к молодым листьям и плодам, а при низкой влажности в течение дня приводит к накоплению Са в зрелых листьях. У сортов и растений с высокой устойчивостью к вершиной гнили плодов наблюдается более интенсивное усвоение растениями К. У чувствительных к вершинной гнили сортов наблюдается недостаток К в растениях, а также значительное накопление Са в листьях, а не в плодах. Недостаток Са связан и с другими ионами — антагонистами из почвенного раствора.
Большое количество Na в почвенном растворе, в присутствии повышенных доз серы снижает поступление Са, в связи с чем, в такой ситуации следует повышать количество Са в почвенном растворе, регулируя одновременно соотношение К : Са.
Избыток К сдерживает поступление Са, так же, как Na, Nh5. При количестве Nh5 в почвенном растворе более 10 мг/л блокируется поступление Са. Одновременно такой уровень Nh5 повреждает корневые волоски, особенно в зимне-весеннем и осеннем периодах. В случае первых признаков вершинной гнили томатов при начале налива плодов соотношение К : Са поддерживают на уровне 0,8-1,5 : 1. Улучшение вкусовых качеств томатов за счет повышенных количеств К в почвенном растворе требуют в свою очередь повышение концентрации Са в растворе для поддержания соотношения К : Са.
На молодых листьях растений с недостатком Са образуется пожелтение, а края листьев становятся бурыми, точка роста может отмереть, а плоды становятся черными вокруг рубца столика (начало вершинной гнили). Вершинная гниль часто является результатом низкой влажности субстрата, нарушением водоснабжения растений томата, засоленностью субстрата. При первых признаках недостатка Са в растениях следует провести несколько опрыскиваний раз в 4—5 дней 0,4—0,5% раствором Ca(NO3)2. При возникновении вершинной гнили опрыскивают тщательно нанося раствор по всей их поверхности. Одновременно увеличивают норму Ca(NO3)2 в питательном растворе, количество Са в субстратном растворе может в 10 раз превосходить нормальное содержание Mg.
Первые признаки недостатка Mg проявляются на старых листьях в виде светлых мраморных пятен, обесцвечиванием листьев между жилками, хотя главная и боковые жилки остаются зелеными. При длительном магниевом голодании наблюдается приостановка роста плодов. При недостатке магния проводят каждые 5—7 дней опрыскивание растений 0,5—0,7% раствором сульфата магния или магниевой селитры. На сортах с повышенной чувствитель-
ностью к недостатку Mg внекорневые подкормки практикуют 1 раз в неделю профилактически начиная от времени завязывания плодов на 4-ой кисти. Проявление хлороза усиливает большая плодовая нагрузка. Следим за соотношением К : Mg, т.к. высокий уровень К сдерживает поглощение Mg. Нормы Mg повышают.
Для регулирования соотношения К : Са для усиления вегетативного или генеративного развития томатов и др. культур применяют изменения соотношения Са : К=1 : 1,2-1до 7—10 дней . Для генеративного развития используют соотношения Са : К 1,2-1 при концентрации катионов около 235 мг/л К и 200 мг/л Са. Для усиления вегетативного развития оптимально соотношение 1 : 1,6-1,7 при концентрации катионов около 235 мг/л К и 320-400 мг/л Са. Кроме указанных соотношений К : Са необходимо поддерживать соответствующий уровень относительной влажности воздуха для оптимальной транспирации.
Усвоение фосфора в оптимальном постоянном количестве до 40-45 мг/л рабочего раствора и в почвенном растворе до 50 мг/л у томатов стимулирует усвоение Са. Кроме оптимального количества фосфора и кальция необходим показатель рН раствора от 5 до 5,9. Одновременно следим, чтобы фосфор не накапливался в более высокой концентрации, что тормозит поступление Mg и вызывает магниевый хлороз. Если показатель рН > 6,2-6,7 то Р становится труднодоступным.
Присутствие в почвенном растворе CI в количестве 1-3 мМо/л т.е. 30-90 мг/л и максимально до 150 мг/л способствует усвоению Са. Так же усиливается усвоение Са и других элементов питания при оптимальном показателе ЕС почвенного раствора до 2,6-2,8 мСм/см и при максимально допустимом в период плодоношения томатов — до 3,5 мСм/см, или чуть более.
Микроэлементы.Отклонение рН в кислую сторону приводит к повышению растворимости соединений Mn, Fe, A1, находящихся в грунте. Высокие концентрации этих ионов могут повредить корневую систему. При значительном повышении рН Fe, Mn, Си образуют нерастворимые соединения — гидрооксиды, которые не усваиваются корневой системой, поэтому вместо сернокислых солей лучше применят хелаты — органические соединения доступные для растений в этих условиях. На щелочных грунтах также резко снижается растворимость бора. Поэтому длительный полив водой с рН выше 7 приводит к подщелачиванию грунтов и снижает доступность вносимых микроэлементов.
При избытке фосфора (зафосфачивание грунтов), Zn и Fe вступают в реакцию с фосфором и образуют трудно растворимые фосфаты этих элементов, которые выпадают в осадок. Так как Zn является антагонистом Fe при повышенном количестве Zn норму железа увеличивают до 2-2,5 мг/л рабочего раствора.
При малообъемном выращивании доступность ионов микроэлементов в корнеобитаемом слое определяется теми же факторами, что и при выращивании на грунтах. Важным является поддержание всех питательных веществ в растворимом состоянии за счет хелатирования таких элементов, как Fe, Zn, Cu, Mn. Так как хелаты более доступны для растений их расход небольшой в сравнении с сернокислыми окислами, которые еще часто рекомендуются к применению.
Учитывая высокую стабильность Fe — ДТРА при рН от 1,5 до 7 в рабочих растворах (в том числе концентрированных) его предпочтительно использовать как основной хелат железа, в сравнении с хелатом Fe — ЕДТА, эффективным при рН ниже 6, т.е. рабочие растворы должны иметь рН ниже 6, например рН 5,5.
При использовании полихелатов в форме ЕДТА необходимо поддерживать показатель рН в почвенном (дренажном) растворе на уровне ниже рН 6, в противном случае норму Fe — ДТРА повышать на 0,4-0,5 мг/л с учетом вносимого Fe — ЕДТА вместе с остальными полихелатами ЕДТА, которые стабильны при рН 3,5-10.
ТРЕБОВАНИЯ К КАЧЕСТВУ ВОДЫ ДЛЯ КАПЕЛЬНОГО ПОЛИВА, МЕТОДИКА КОРРЕКТИРОВКИ ПИТАТЕЛЬНОГО РАСТВОРА В ЗАВИСИМОСТИ ОТ СОСТАВА ВОДЫ
При использовании капельного полива качество воды является одним из основных факторов, обеспечивающих успех тепличного производства. Поэтому так важно знать химический состав используемой в хозяйстве воды. Это необходимо, как для расчета количества солей и кислот в питательном растворе, так и при его коррекции. В каждом тепличном комбинате используемая поливная вода имеет определенный химический состав, что следует учитывать еще на стадии проектирования систем капельного полива, с учетом пригодности конкретной воды.
Различная по происхождению вода представляет собой сложный раствор, содержащий все известные химические элементы в виде простых и сложных ионов, комплексных соединений, растворенных или газообразных молекул, стабильных и радиоактивных изотопов, а также определенный бактериальный состав.
Сложность состава определяется присутствием большого числа химических элементов, различным содержанием и разнообразием форм каждого из них.
В воде отмечается 6 основных групп химических компонентов:
1) главные ионы (макроэлементы) К+, Na+, Mg2+, Са2+, С1 ', SO42,НСО3',СО2;
2) растворенные газы — кислород, азот, сероводород, углекислота;
3) биогенные вещества — соединения N, P, Fe, Si;
4) органические вещества — органические кислоты, сложные эфиры,фенолы, гумусовые вещества;
5) микроэлементы;
6) загрязняющие вещества.
Суммарное содержание минеральных веществ называется минерализацией воды, которая выражается в мг/дм3, г/дм3, г/кг, % (промилле (от лат. Pro mille — за тысячу) тысячная часть числа, обозначается — %).
По степени минерализации вода бывает пресной до 1%, солоноватой (1—25%), соленой (25—50%), очень соленой (более 50%). Для капельного орошения лучше использовать воду с содержанием минеральных веществ до 0,5—1%.
Повышенное поступление солей с поливной водой приводит к засолению субстратов, что отрицательно сказывается на продуктивности растений. Томаты более солеустойчивы, чем огурцы, но на засоленных субстратах сильнее поражаются вершинной гнилью.
Химический состав воды для приготовления питательных растворов не должен превышать величины указанной в таблице 7.4.
На основе химического состава воды проводится коррекция питательного раствора. Существует правило, по которому концентрация элементов в поливной воде не должна превышать их содержание в стандартных питательных растворах. Особенно это касается микроэлементов, так как выращивание растений в ограниченном корневом объеме может привести к их накоплению и отравлению растений или к явлениям антогонизма элементов.
Питательный раствор обязательно корректируется на содержание присутствующих в воде К, Са, Mg, SO42~, NO3\ Реакция его доводится до оптимального уровня рН, который для большинства культур составляет 5,5-6,0. Так как вода чаще всего слабощелочная или щелочная для снижения рН используют ортофосфорную (Н3РО4) или азотную (HNO3) кислоты.
Количество кислоты рассчитывается по содержанию бикарбонатов (НСО3). В принципе на 1 мМоль НСО3 в воде нужен 1 мМоль кислоты. Однако в нейтрализации участвуют не все бикарбонаты, поэтому для обеспечения буферности раствора оставляют 1 мМоль НСО3= 61 мг без нейтрализации 0,5-1 мМоль/л.
Кислоты и бикарбонаты взаимодействуют в эквивалентных количествах:
1 мМ НСО3- = 61 мг/л
cyberpedia.su
Оптимизация питания: рациональное питание | Компетентно о здоровье на iLive
Оптимизация питания
Оптимизация питания - проблема, нуждающаяся в общем решении. Она представляется особенно важной, так как для настоящего времени характерны дефекты питания в глобальных масштабах. С дефектами питания человек сталкивается также в различных аварийных и экстремальных ситуациях и будет сталкиваться при создании искусственных микробиосфер и микротрофосфер в связи с исследованием космоса, океана и другими задачами. Поэтому необходимо понять, каким образом последствия различных дефектов питания могут быть предупреждены или ослаблены.
Прежде всего рассмотрим крайний случай - отсутствие продуктов питания. Долгое время считалось, что в таких условиях полное голодание предпочтительнее, чем поедание непитательных структур (в частности, листьев растений). В рамках теории сбалансированного питания это объяснялось тем, что при питании листьями происходит значительная работа пищеварительного аппарата и имеет место дополнительный расход энергии, а также размножение кишечной бактериальной флоры, отрицательно действующей на организм. Однако с позиций теории адекватного питания нормальная деятельность желудочно-кишечного тракта необходима для поддержания в норме многих сторон обмена, для функционирования эндокринного аппарата и, что чрезвычайно важно, для поддержания эндоэкологии кишечника. Сохранение эндоэкологии благодаря пищевым волокнам, по-видимому, более благоприятно для организма, чем абсолютное голодание. Такая точка зрения подтверждается известными примерами о поведении людей во время голода, добавляющих в пищу траву, листья, древесные опилки и т. д. Иной смысл, чем несколько лет назад, возможно, приобретает оценка поведения некоторых хищных животных, во время голода питающихся листвой, травой, ягодами и другими растениями. Понятными становятся также различия между полным голоданием и голоданием в сочетании с потреблением продуктов с незначительной энергетической ценностью, но существенно влияющих на соседние эндоэкологии организма. Использование пищевых волокон - лишь один из многочисленных примеров оптимизации питания в неблагоприятных условиях.
Использование нерафинированных пищевых продуктов (например, цельнозернового хлеба, неполированного риса и др.), что ценно само по себе, при оптимизации питания в условиях нехватки пищевых ресурсов приобретает особенно большое значение. По-видимому, нерафинированные продукты и в большинстве других случаев имеют преимущества перед рафинированными.
Справедливость ряда положений теории адекватного питания хорошо демонстрируется на примере диких животных, инстинкт которых помогает им поддерживать структуру своего тела с необычайной точностью. Что касается человека, то он, вероятно, в процессе формирования вида Homo sapiens, а также в результате воспитания (часто неправильного), традиций, предрассудков и т. д. в значительной мере утрачивает эти навыки и инстинкты, обеспечивающие адекватный выбор пищи. Следует подчеркнуть, что оптимизация питания была важной и полезной частью национальных, племенных и религиозных традиций. Однако в научной литературе часто обращается внимание лишь на недостатки этих традиций, многие из которых теперь утрачены. В то же время такая утрата создает вакуум, который нередко заполняется неправильными и неорганизованными действиями. Последние опираются на многочисленные модные концепции питания, иногда не имеющие теоретической базы и не подкрепленные вековой практикой.
По-видимому, при оптимизации питания следует иметь в виду его национальные особенности, ассортимент соответствующих продуктов (который зависим от окружающей среды, способа добывания и обработки пищи), уровень технологии и т. д. Известно, что относительное потребление энергии в виде жиров у эскимосов достигает 47 %, а у кикуйю составляет только 10 %. В отличие от эскимосов европейцы и американцы, как и кикуйю, потребляют значительно меньше жиров. Вместе с тем следует иметь в виду, что люди, как правило, довольно легко адаптируются к изменению рациона.
При оптимизации питания следует, по-видимому, учитывать сведения, что в ряде случаев некоторые следовые микроэлементы могут иметь важное нутритивное значение.
Оптимизация питания связана с решением еще нескольких проблем. Поскольку создание токсикантов, предназначенных для борьбы с вредителями сельского хозяйства и безвредных для человека, практически невозможно из-за универсальности функциональных блоков, то прежде всего должны быть получены такие соединения, которые выполняли бы основные регуляторные функции во внешней среде, но не попадали бы в пищевые продукты или пищу. Далее, следует искать соединения с такой степенью избирательности, при которой эти вещества и их метаболиты были бы по возможности индифферентны для человека. Важно также разработать пищевые технологии, в том числе кулинарные, при которых в процессе приготовления пищи токсические вещества разрушались бы или превращались в безвредные. Наконец, должна существовать достаточно полная и открытая информация о наличии в продуктах токсических соединений и о возможности чередования этих продуктов во избежание кумулятивных эффектов неблагоприятных воздействий и т. д.
Понятие адекватности позволяет оптимизировать питание в соответствии с возрастом и характером труда. Но и в этом случае пища не будет идеальной. Определенные перспективы для оптимизации питания открывает улучшение аминокислотного состава пищи путем введения в нее соответствующих пептидов вместо аминокислот. Как известно, в 1970-1980-х годах были разработаны питательные смеси на основе коротких пептидов и свободных аминокислот. Зарубежными фирмами выпущен ряд пептидных диет. При этом было продемонстрировано, что диета, содержащая короткие пептиды, утилизируется более эффективно, чем смесь свободных аминокислот. Рядом исследователей была показана высокая ценность белковых гидролизатов, содержащих короткие пептиды. Следует также отметить, что в отличие от неприятного вкуса аминокислотных смесей гидролизаты пищевых белков (в том числе смеси, состоящие из коротких пептидов) во многих случаях обладают довольно приятным вкусом. Пептидные гидролизаты могут быть рекомендованы для питания ослабленных организмов, при краткосрочных диетах, при больших физических нагрузках и т. д.
При оптимизации питания необходимо также учитывать пищевые добавки, которые находят все большее применение в питании человека и особенно сельскохозяйственных животных. В число пищевых добавок входят антибиотики, факторы (стимуляторы) роста, кокцидиостатики, гистомоностатики и др. Их полезные и побочные отрицательные эффекты в последние годы широко дискутируются. При этом постоянно предлагаются новые добавки, в том числе стимуляторы роста, антибиотики, анаболические агенты, дрожжевые культуры, микроорганизмы и т. д.
Основной вопрос - безопасность таких добавок для потребителя. В то же время необходимо помнить, что фактически во всех продуктах питания (как естественных, так и переработанных) возможно присутствие нежелательных веществ. И система строгого контроля всей трофической цепи, защищая человека, не всегда может гарантировать его здоровье. Например, избыток даже полноценной, абсолютно «здоровой» пищи может привести к ожирению, сердечно-сосудистым заболеваниям, развитию злокачественных опухолей и другим серьезным болезням. В последние годы для потери лишнего веса рекомендуется ряд более низкокалорийных диет (400-600 ккал в сутки), чем обычно, а также полуголодание. Поэтому при попытках оптимизировать питание следует помнить слова Гиппократа, сказанные более 2300 лет назад: «Диететика позволяет тем, кто имеет хорошее здоровье, сохранить его, а тем, кто потерял здоровье, его восстановить».
Различные категории пищевых добавок, рекомендованные ЕЭС (по: Vanbelle, 1989)
- Антибиотики (стимуляторы роста)
- Факторы роста (стимуляторы роста)
- Кокцидиостатики и гистомоностатики
- Ароматические и вкусовые вещества
- Эмульсии, стабилизирующие агенты, гели и уплотнители
- Цветовые агенты и пигменты
- Предохранители
- Витамины и витаминоподобные вещества
- Микроэлементы
- Ферменты, холин
Наконец, оптимизация питания необходима в связи с проблемой пищи будущего.
ilive.com.ua
Диссертация на тему «Оптимизация минерального питания растений» автореферат по специальности ВАК 06.01.04 - Агрохимия
1. Авакимова Л.Г., Королькова H.A. Сравнительная характеристика фотосинтетической деятельности различных сортов гороха // Докл. ТСХА. Вып. 206, 1975. - С. 91 - 96.
2. Агрохимические методы исследования почв. 5-е изд. - М.: Нау-ка,1975.- 656 с.
3. Агрохимия / Под ред. Б.А. Ягодина. М.: Колос, 1989. - 654с.
4. Алиев Н.Т. Оптимизация суммарной дозы и соотношения азота, фосфора и калия в удобрении под озимую пшеницу в условиях Апшерона: Ав-тореф. дис. канд. с.-х. наук-Баку, 1988. 23 с.
5. Алиев Н.Т., Марданов A.A. Раздельное определение оптимумов суммарной дозы N+P+K и соотношения N:P:K в удобрении для урожая зерна озимой пшеницы в условиях Апшерона //Агрохимия. — 1985. № 2-С. 47-50.
6. Алиев Н.Т., Марданов A.A. Раздельное определение оптимумов суммарной дозы N+P+K в удобрении. Продуктивность озимой пшеницы //Агрохимия.- 1986. № 5. - С. 36-45.
7. Арбузова И.Н., Ниловская Н.Т., Силецкая Л.А. Характер накопления элементов минерального питания яровой пшеницы в искусственных условиях выращивания // Агрохимия. 1978. - № 9. - С. 73.
8. Баланс азота в земледелии Украины // Круговорот и баланс азота в системе почва-удобрение-растение-вода. М.: Наука, 1979. С. 104-111.
9. Балашев JI.J1. Удобрение и сорт // Агрохимия, № 8, 1966.
10. Басистый В.П., Юрова С.Н., Кофан А.И. Известкование пахотных земель в Хабаровском крае: Рекомендации.- Новосибирск: Васхнил СО, Дальниисх, 1990.- 24 с.
11. Бентли М. Промышленная гидропоника. М.: Колос, 1965. - 376 с.
12. Блэк К.А. Растение и почва. М., 1973.
13. Болдырев H.K. Анализ листьев как метод определения потребности растений в удобрениях. (Листовая диагностика): Учебное пособие. Омск: ОмСХИ, 1970.
14. Булаткин Г.А., Шевцов Н.Д. Влияние минеральных удобрений на урожай и химический состав озимой тииеницы на выщелоченном черноземе // Агрохимия, № 11, 1974. С. 44 - 50.
15. Вахмистров Д.Б. Сортовые различия в поглощении и транспорте ионов культурными растениями // С. х. за рубежом, растениеводство, № 1, 1967.
16. Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости // Классики советской генетики. JL, 1968. - С. 9 - 50.
17. Васильев А.И. Поглощение и использование азота, фосфора, калия кукурузой в зависимости от густоты стояния и уровня минерального питания // Агрохимия. 1979. - № 8. - С. 46-50.
18. Ватагин A.B., Сергеева М.Е. Влияние удобрений на динамику питательных веществ в почве, поступление их в растения и урожай ячменя // Агрохимия. 1971. - № 10. - С. 88 - 93.
19. Вахмистров Д.Б. Исследования механизмов поглощения солей растениями //Агрохимия.- 1968. № 1. - С. 151-163.
20. Вахмистров Д.Б. О двух механизмах избирательности при поглощении растениями элементов минерального питания // Физиология растений. -1966. Т. 13, вып. 5. - С. 807-812.
21. Вахмистров Д.Б. Потребление проростками ячменя элементов питания из эндосперма и наружной среды // Физиология растений. 1980. -27(3).- С. 551-559.
22. Вахмистров Д.Б. Пространственная организация ионного транспорта в корне. М.: Изд-во АН СССР, 1991. - 48 с. - (49-е Тимирязявское чтение).
23. Вахмистров Д.Б. Раздельное определение оптимумов суммарной дозы Ы+Р+К и соотношения №Р:К в удобрении. Сообщение 1. Постановка проблемы //Агрохимия. 1982. - № 4. - С. 3-12.
24. Вахмистров Д.Б. Распределительная функция корневой системы растений // Агрохимия. 1966. - № 2.- С. 49 - 55.
25. Вахмистров Д.Б. Современные представления о механизмах первичного поглощения солей растениями // Агрохимия. 1966. - № 11. - С. 130145.
26. Вахмистров Д.Б. Сортовые различия в поглощении и транспорте ионов культурными растениями // С.-х. за рубежом. Растениеводство. 1967. -№ 1.
27. Вахмистров Д.Б., О Эн До Генетические особенности соотношения Ы:Р:К в злаковых растениях //Агрохимия. 1992. - № 2. - С.52-62.
28. Вахмистров Д.Б., Вильяме М.В., Шарма Г., Ягодин Б.А. Соотношение №Р:К в среде и урожай: Анализ формы купола отклика // Агрохимия. -1986.-№ 11. -С. 30-37.
29. Вахмистров Д.Б., Воронцов В.А. Избирательная способность растений не направлена на обеспечение их максимального роста // Физиология растений. 1997. - Т. 44, № 3. - С. 404-412.
30. Вахмистров Д.Б., Воронцов В.А. Соотношение элементов минерального питания в среде и рост растений. 1. Исследование купола отклика // Физиология растений. 1994, Т.41, - № 1. - С. 56-63.
31. Вахмистров Д.Б., Воронцов В.А. Соотношение элементов минерального питания в среде и рост растений. 2. Уточнение формы купола отклика // Физиология растений. 1994. - Т. 41, № 1. - С. 64-69.
32. Вахмистров Д.Б., Воронцов В.А. Соотношение элементов минерального питания в среде и рост растений. 3. Корреляционное зондирование купола отклика // Физиология растений. 1994. - Г. 41, № 3.- С. 425-429.
33. Вахмистров Д.Б., Воронцов В.А. Соотношение элементов минерального питания в среде и рост растений. 4. Минимизация объема опыта для определения оптимального соотношения элементов // Физиология растений. 1994. - Т. 41, № 3. - С. 430-435.
34. Вахмистров Д.Б., Воронцов В.А. Соотношение элементов минерального питания в среде и рост растений. 5. Оптимизация N:P:K для водной культуры кукурузы и его сезонная динамика // Физиология растений. -1995. Т. 42, № 2. - С. 248-253.
35. Вахмистров Д.Б., Воронцов В.А. Соотношение элементов минерального питания в среде и рост растений. 6. Зависит ли оптимум соотношения от суммарной концентрации и наоборот? // Физиология растений. 1995. -Т. 42, №2.-С 254-261.
36. Вахмистров Д.Б., Журбицкий З.И. О диапазоне избирательной способности растений к поглощению растениями элементов минерального питания //Докл. АН СССР, 1963.-Т. - 151, № 5. - С. 1228-1231.
37. Вахмистров Д.Б., Пахомова Л.М., Балахонцев E.H., Исхаков Ф.М., Ис-хаков Ф.Ф. Раздельное определение оптимумов суммарной дозы N+P+K и соотношения N:P:K в удобрении. Сообщение 7. Продуктивность сахарной свеклы //Агрохимия. 1983. - № 8. - С. 15-27.
38. Вахмистров Д.Б., Смирнова В.В. Зависимость полегаемости озимой пшеницы от суммарной дозы N+P+K, соотношения N:P:K в удобрении и распределения годовой дозы азота между тремя сроками внесения // Агрохимия 1992. - № 1. - С. 40 - 49.
39. Вахмистров Д.Б., Смирнова B.B. Зависимость полегаемости озимой пшеницы от суммарной дозы N+P+K, соотношения N:P:K в удобрении и распределения годовой дозы азота между тремя сроками внесения // Агрохимия. 1992. - № 1.- С. 40-49.
40. Вахмистров Д.Б., Смирнова В.В. Оптимизация соотношения N:P:K в удобрении: сравнение методов // Агрохимия. 1990.-№ 11. — С. 128-139.
41. Вахмистров Д.Б., Смирнова В.В. Оптимизация суммарной дозы N+P+K и соотношения N:P:K в удобрении озимой пшеницы для лесостепи Украины // Агрохимия. 1991. - № 4. - С. 25-34.
42. Вахмистров Д.Б., Смирнова В.В. Применение метода корреляционного зонда для оптимизации сроков внесения азота под озимую пшеницу // Агрохимия.- 1991. № 10. - С. 19-28.
43. Вахмистров Д.Б., Федоров A.A. Анализ сортовых различий минерального состава растений ячменя в зависимости от их продуктивности // Агрохимия. 1978. - № 7. - С. 46 - 56.
44. Вахмистров Д.Б., Федоров A.A. Раздельное определение оптимумов суммарной дозы N+P+K и соотношения М:Р:К в удобрении. Сообщение 2. Предварительный анализ //Агрохимия. 1982. - № 5. - С. 3-11.
45. Вахмистров Д.Б., Федоров A.A. Раздельное определение оптимумов суммарной дозы N+P+K и соотношения N:P:K в удобрении. Сообщение
46. Качество урожая сои //Агрохимия. 1982. - № 7. - С. 3-10.
47. Вахмистров Д.Б., Федоров A.A. Раздельное определение оптимумов суммарной -дозы N+P+K и соотношения N:P:K в удобрении. Сообщение
48. Выход белка и жира с урожаем сои //Агрохимия. 1982.-№8. - С. 3-11.
49. Вахмистров Д.Б., Федоров A.A. Раздельное определение оптимумов суммарной дозы N+P+K и соотношения N:P:K в удобрении. Сообщение
50. Связь между величиной и качеством урожая сои //Агрохимия. 1983. -№> 4. -С. 3-10.
51. Вахмистров Д.Б., Федоров A.A. Раздельное определение оптимумов суммарной дозы N+P+K и соотношения N:P:K в удобрении. Сообщение 9. Продуктивность кукурузы //Агрохимия. 1985. - № 12. - С. 78-88.
52. Вахмистров Д.Б., Федоров A.A., Дворникова Н.Е Раздельное определение оптимумов суммарной дозы N+P+K и соотношения N:P:K в удобрении. Сообщение 3. Урожай семян сои //Агрохимия. 1982. - № 6. - С. 311.
53. Вахмистров Д.Б., Шарма Г., Вильяме М.В., Ягодин Б.А. Корреляционный зонд как метод оптимизации соотношения N:P:K в питательной смеси // Агрохимия. 1986. - № 12. - С. 54-62.
54. Вильяме М.В., Шарма Г., Ягодин Б.А., Вахмистров Д.Б. Оптимизация соотношения N:P:K в питательной смеси для песчаной культуры ячменя // Физиология и биохимия культурных растений. 1986. - Т. 18, № 4. -С. 222-231.
55. Виноградов А.П. Развитие идей В.И. Вернадского // Почвоведение.1963.-№ 8.-С. 1-10.
56. Воробьев С.А., Буров Д.И., Туликов А.М. Земледелие. М.: Колос, 1981. - 479 с.
57. Гинзбург К.Е. Методы определения фосфора в почве // Агрохимические методы исследования почв. 5-е изд. - М.: Наука, 1975. - С. 106-109.
58. Годунова К.Н. Сорт и удобрения // Селекция и семеноводство, № 2,1964.
59. Горбунов H.H. Минералогия и физическая химия почв. М.: Наука, 1978.-294 с.
60. Грицун А.Т. Основы возделывания сои в Приморье. Владивосток, 1981.- 159с.
61. Грицун А.Т. Основы возделывания сои в Приморье. Владивосток: Дальиздат, 1981. - 159 с.
62. Грицун А.Т. Роль бобовых культур (сои, клевера) в накоплении биологического азота и повышении плодородия почвы в условиях Дальнего Востока // Агрохимия. 1975. - № 5. - С. 17-23.
63. Гуревич Г.М., Бородина И.И. Поступление питательных веществ в растение ячменя и вынос их с урожаем в зависимости от уровня питания // Агрохимия. 1965. - № 12.
64. Доспехов Б.И. Методика полевого опыта. М., 1985. - 351 с.
65. Егоров В.В. Органическое вещество почвы и ее плодородие // Вестник с.-х. науки.- 1978.- №5. С. 15.
66. Егорова Л.А. Изучение минерального питания гвоздики голландской как основание для разработки системы внесения удобрений: Автореф. дис. канд. с.-х. наук. -М., 1969. 16 с.
67. Енкен В.Б. Соя. -М.: Сельхозгиз, 1952. С. 180.
68. Ермохин Ю.И. Изучение сока черешков листьев как способ контроля питания // Науч. тр. ОмСХИ. 1970. - Т. 87.
69. Жуков Ю.П. О рациональном удобрении орошаемых пастбищ // Известия TCXA.-1978.-X2l. С. 85-90.
70. Жуков Ю.П., Глухов Н.И. Определение оптимальных доз и соотношений удобрений с учетом использования питательных элементов из удобрений и почвы // Известия ТСХА.- 1977.- №4.- С. 68-76.
71. Жуков Ю.П., Загоняйло Л.И. Продуктивность овощного севооборота при использовании систем удобрения, расчитанных балансовым методом // Известия ТСХА.- 1979.- №4.- С. 120-128.
72. Журбицкий З.И. Питательные смеси для разных культур // Агрохимия. -1964.-№2.-С. 18.
73. Журбицкий З.И. Физиологические и агрохимические основы применения удобрений. М.: Изд-во АН СССР, 1963.- 328 с.
74. Журбицкий 3.И.Обеспечение помидоров 1ЧРК по этапам роста в связи с избирательной способностью растений // Физиологическое обоснование системы питания растений. М, 1964. - С. 15-21.
75. Журбицкий 3.И.Теория и практика вегетационного метода. М.: Наука, 1968.-С. 18-142.
76. Журбицкий З.И., Вахмистров Д.Б. Удовлетворение потребности томатов в азоте, фосфоре и калии по этапам роста в связи с избирательной способностью растений // Физиологическое обоснование системы питания растений. М., 1964. - С. 15-21.
77. Журбицкий З.И., Лавриченко В.М. Определение потребности растений в питании и удобрении по соотношению ИРК: Обзорн. информ. / ВНИИТЭИСХ. М., 1982.-63 с. - (Обзоры по важнейшим науч.-техн. проблемам).
78. Ильин С.С. Роль удобрения и бобовых культур в повышении плодородия почв // Органические удобрения: Материалы науч.-методич. совещания стран-участниц СЭВ.- М., 1972.- С. 327-342.
79. Катутис К.Ю., Вахмистров Д.Б. Оптимизация соотношения .М:Р:К в удобрении польдерских дамб. Сообщ. 1. Плотность травостоя (сравнение методов) // Агрохимия. 1992. - № 4. - С.41-50.
80. Катутис К.Ю., Вахмистров Д.Б. Оптимизация соотношения 1М:Р:К в удобрении польдерских дамб. Сообщ. 2. Урожай сухой массы побегов и корней многолетних трав // Агрохимия. 1992. - № 5. - С. 78-89.
81. Климашевский Э.Л. Генотипические реакции растений на уровни корневого питания. Физиология и биохимия культурных растений. Т.1. Вып. 3, 1969
82. Климашевский Э.Л. Сорт удобрение - урожай // Вестн. с.-х. науки, №2, 1974.
83. Климашевский Э.Л. Специфика генотипических реакций растений на удобрения // Сибирский вестн. с.-х. науки,№ 2, 1972
84. Климашевский Э.Л. Теория агрохимической эффективности растений // Агрохимия. 1990. - № 1. - С 131.
85. Климашевский Э.Л. Физиологические особенности корневого питания разных сортов кукурузы в нечерноземной полосе. — М., 1966.
86. Климашевский Э.Л. Физиология генотипической специфики корневого питания растений // Физиология растений в помощь селекции. М., 1974.-С. 226.
87. Климашевский ЭЛ., Писецкая Н.Ф. О физиологической разнокачественности сортов кукурузы в связи с применением удобрений // Агрохимия, № 8, 1964.
88. Кнаппе С., Рерихт., Рауэ К. Изменение содержания гумуса в почве в зависимости от типа севооборота и системы удобрений // Органические удобрения: Материалы науч.-методич. совещания стран-участниц СЭВ.-М., 1972.-С. 343-350.
89. Колларжик И. Анализ растений как метод научного исследования правильного питания растений II За социалистическую с.-х. науку. 1959. -№ 6. - С. 75 - 79.
90. Колларжик И. Вопросы питания и удобрения кукурузы. // Международный с.-х. журнал. 1961. - № 3. - С. 57 - 63.
91. Король В.Э. Соя. Культура и использование. М.: СКХГИЗ, 1931. -С.155.
92. Кук Дж. У. Регулирование плодородия почв. М.: Колос, 1970. - 520 с.
93. Куркаев В.Т. Ускоренное определение №>К в растениях из одной навески // Почвоведение. 1959. - № 9. - С. 114-117.
94. Куртесов А.П. Вопросы улучшения плодородия дерново-подзолистых почв Суйфуно-Ханкайской равнины. Владивосток, 1949. - 110 с.
95. Къдрев Т., Андонова П. Установяване на оптимални съотношения и концентрация на микроэлемента при отглеждането на царевица // Физи-ол. растен. 1979. - № 5. - Кн. 2. - С. 71 - 80.
96. Лавриченко В.М. Соотношение элементов питания в растении как видовое генотипическое понятие // Вестник с.-х. науки. 1971. - № 7. - С. 129- 134.
97. Лавриченко В.М., Журбицкий З.И. Соотношение элементов питания в растениях и видовое генотипическое явление // Агрохимия. 1976. - № 9.-С. 135-140.
98. Либих Ю. Химия в приложении к земледелию и физиологии. М.- Л., 1936.-393 с.
99. Лигум С.Т. Балансовый коэффициент использования растениями питательных веществ из удобрений и почвы и его применение // Агрохи-мия.-1977.-№5.- С. 128-133.
100. Лыков A.M. Гумус и плодородие почвы.- М.: Моск. рабочий, 1985.-191 с.
101. Магницкий К.П. Диагностика потребности растений в удобрениях. М.: Московский рабочий - 1972.
102. Максимов И.Л., Понаморев В.И. Повышение качества зерна пшеницы: Обзорн. информ. / ВНИИТЭИСХ. М., 1981. - 67 с. - (Обзоры по важнейшим науч.-техн. проблемам).
103. Методические указания по проведению исследований в длительных опытах с удобрениями. Ч. 2 (анализ растений). М., 1976. - 166 с.
104. Методические указания по проведению полевых опытов с удобрениями географической сети на 10-ю пятилетку. М.: Изд-во ВИУА, 1976. -139с.
105. Минеев В.Г. Агрохимия. -М.: Изд-во МГУ, 1990. 486 с.
106. Митчерлих Э.А. Определение потребности почвы в удобрении. М.; Л.: Сельхозгиз, 1931. - 103 с.
107. Михайлов H.H., Книпер В.П. Определение потребности растений в удобрениях. М.: Колос, 1971. - 255 с.
108. Мишустин E.H., Шильникова В.К. Биологическая фиксация атмосферного азота. М.: Наука, 1967. С. 56-58.
109. Мишустин E.H., Шильникова B.K. Клубеньковые бактерии и инокуля-ционный процесс. М.: Наука, 1973. - 118 с.
110. Мишустина Н.Е. Влияние соотношений минеральных элементов в питательной среде на их поглощение и рост кукурузы. // Автореф. дис. канд. биол. наук. -М., 1970. 32 с.
111. Мишустина Н.Е. О двух основных методах построения системы минерального питания растений // Известия АН СССР. Сер. биол. 1972. -№1.-С.88-95.
112. Момот Я.Г. Соя в СССР. М.;Л.: Госсельхозиздат, 1930. С.47.
113. Мочалова А.Д. Спектрофотометрический метод определения содержания серы в растениях // С.-х. за рубежом. 1975. - № 4.
114. Нарциссов В.Н. Некоторые итоги и задачи изучения севооборотов в условиях Нечерноземной полосы Европейской части СССР// Окультуривание дерново-подзолистых почв: Тр. Горьковского СХИ . Горький, 1973 - С. 33-45.
115. Научные отчеты отдела агрохимии Приморского НИИ сельского хозяйства за 1974-1992 гг.
116. Ниловская Н.Г., Арбузова И.Н. Об оптимальном соотношении элементов минерального питания для растений // Агрохимия. 1978. - № 5 - С. 138.
117. Ниловская Н.Т., Арбузова И.Н. О соотношении элементов питания в среде и продуктивность растений // Агрохимия. 1982. - № 2. - С. 126 -131.
118. Ниловская Н.Т., Арбузова И.Н., Осипова Л.В. Продуктивность яровой пшеницы в зависимости от соотношения элементов минерального питания: Обзорн. информ. / ВНИИТЭИСХ. М., 1984. - 51 с. - (Обзоры по важнейшим науч.-техн. проблемам).
119. Новак А.Г. Соя на Дальнем Востоке. Владивосток, 1960. 303с.
120. О Эн До. Генотипические особенности накопления элементов минерального питания злаковыми растениями: Автореф. дис. канд. биол. наук. -М, 1991.-21 с.
121. Орлов Н.В. Влияние почвенных условий на урожай и химический состав различных сортов гречихи // Агрохимия, № 6, 1967.
122. Отзывчивость сортов зерновых культур на удобрения. М., 1970.
123. Пандев С. Влияние на соотношението и концентрацията на йоните на азота, фосфора и цинка в хранителния раствор въерху натрупването или в пшеничните растении и върху братенето // Растениевъдни науки. -1979.-Т. 16, №8.-С. 5-10.
124. Пандев С., Къдрев Т., Станев В. Влияние на преобладаването на отдел-ните хранителни элемента (N, S, Р, К, Ca, Mg) в хранителния и качест-вото на добива при слънчоглода // Физиол. растен. 1981. - Т. 7. - Кн. 3.-С. 46-51.
125. Перегудов В.Н. Метод наименьших квадратов и его применение в исследованиях. М.: Колос, 1965. - 182 с.
126. Перегудов В.Н. Планирование многофакторных полевых опытов с удобрениями и математическая обработка их результатов. М.: Колос, 1978.-182 с.
127. Пестриков В.К. Окультуривание дерново-подзолистых почв (в северозападной зоне).- Л.: Колос, 1970. -120 с.
128. Петербургский A.B. Корневое питание растений. М.: Россельхозиздат, 1964. - 254 с.
129. Петербургский A.B. Практикум по агрономической химии. М.: Колос, 1963.
130. Петербургский A.B. Практикум по агрономической химии. М.: Сель-хозгиз, 1954.-С. 72.
131. Плешков Б.П., Неттевич Э.Д., Ракипов Н.Г. Биохимическая характеристика белков ярового ячменя Московского 121 икороткостебельного мутанта, выделенного из этого сорта // Докл. ВАСХНИЛ. Вып. 4, 1975.
132. Плешков Б.П., Ракипов Н.Г. Аминокислотный состав белков зериа возделываемых сортов ячменя // Изв. ТСХА. Вып. 1, 1977.
133. Прево П., Оланье М. Закон минимума и сбалансированное минеральное питание // Анализ растений и проблемы удобрений. М., 1964. - С.247-270.
134. Прошляков A.A. Бубнов A.A. О выносе азота, фосфора и калия урожаем ячменя на дерново-подзолистой легкосуглинистой почве // Агрохимия. 1971. -№ 10.-С.69-78.
135. Прошляков A.A. О высокой эффективности калия на дерновоподзоли-стых почвах левобережья Березины //Агрохимия. 1970. - №1. - С.50-55.
136. Прянишников Д.Н. Азот в жизни растений и в земледелии СССР // Собр. соч. М.: Сельхозгиз, 1965. Т. 3.- С. 283-468.
137. Прянишников Д.Н. Избр. соч. Т.1 Агрохимия.- М.: Колос, 1965. - 345с.
138. Ринькис Г.Я. Оптимизация минерального питания растений. Рига: Зи-натне, 1972. - 355 с.
139. Ринькис Г.Я. Основные итоги исследований по оптимизации минерального питания растений // Физиология растений. 1976. - Т. 23, вып. 6. -С. 1128- 1133.
140. Ринькис Г.Я., Нолпендорф В.Ф. Оптимизация минерального питания полевых и тепличных культур. Рига: Знание, 1977. - 167 с.
141. Рокицкий П.Ф. Основы вариационной статистики для биологов. -Минск: Изд-во Белорусского ун-та, 1961. 145 с.
142. Романенкова М.М., Лубяков B.C. Влияние минеральных удобрений на урожай разных сортов ячменя // Агрохимия, № 11, 1974. С. 82 - 89.
143. Рюбензам Э.,Рауэ Н. Земледелие.- М.: Колос, 1965.- 518 с.
144. Сабинин Д.А. Диагностирование потребности растений в удобрениях по физиологическим признакам // Химизация социалистического земледелия. 1932. -№ 1.
145. Сальников В.К. Внесение минеральных удобрений в целях получения запланированных урожаев (обзорная информация). М., 1979. - 48 с.
146. Самцевич С.А, Активные выделения корней растений и их значение // Физиология растений.- 1965.- Т. 12, вып 5.- С. 837.
147. Сдобникова О.В. Условия эффективного применения фосфорных удобрений.- М., 1979.- Вып 57.- С. 3-20.
148. Смирнова В.В. Оптимизация суммарной нормы и соотношения азота, фосфора, калия и сроков внесения азота под озимую пшеницу в лесостепной зоне Украины: Автореф. дис. канд. с.-х. наук- М., 1992.-12с.
149. Соколов A.B. Вегетационный метод //Агрохимические методы исследования почв. 5-е изд. - М.: Наука, 1975. - С. 585-604.
150. Соколов A.B. Использование азота бобовых трав в земледелии // Тр. Почв, ин-таим. В.В. Докучаева. М., 1957. Т.50.
151. Соколов A.B. Предисловие к русскому изданию книги Э.Хьюитта «Песчаные и водные культуры в изучении питания растений». М.: Иностранная литература, 1960. - С. 8.
152. Соколов A.B. Регулирование круговорота веществ в земледелии// Агрохимия.-1974.-№1.-С. 3-7.
153. Соколов A.B. Состояние и задачи теоретических и методических работ в области агрохимии // Агрохимия. 1967. - № 8. - С. 3.
154. Сорт и удобрение. Иркутск, 1974.
155. Стребков И.М. Оптимизация калийного питания // Бюл. ВИУа.- 1983.- № 90.- С. 48-55.
156. Стребков И.М., Холанская Т.П. Гумус и эффективность удобрений // Бюл. ВИУа.- 1988.- С. 55-59.
157. Сусляков B.C. Кущение сортов яровой пшеницы в лесостепной части Сибири // Тр. СибНИИСХ. Т. 1 ( 16). - Омск, 1971. - С. 45 - 47.
158. Тильба В.А. Аборигенная популяция ризобий сои основной соесеющей зоны России: Диссертация в виде научного доклада доктора биол. наук Владивосток, 1998. - 47 с.
159. Титлянова A.A. Вариабельность химического состава растений // Изв. СО АН СССР. 1972. - Вып. 1, № 5. - С. 21 - 32.
160. Трепачев Е.П. О методах исследования азотфиксирующей способности бобовых культур // Агрохимия. 1981. - № 12. - С. 129-141.
161. Трепачев Е.П., Атрашкова H.A., Хабарова А.И. О методах определения и размерах фиксации атмосферного азота бобовыми растениями // Биологический азот в земледелии Нечерноземной зоны СССР. М.: Колос, 1970. С. 27-74.
162. Трофимовская А.Я. Генофонд ячменя с повышенным содержанием белка. Тр. по прикладной .ботанике, генетике и селекции. Т. 49. Вып. 3, 1973.
163. Тюрин И.В. Почвообразовательный процесс, плодородие почв и проблема азота //Почвоведение. 1956. - № 3. - С. 1-17.
164. Удобрение и применение удобрений. Термины и определения. ГОСТ 20432-75. М.: Издательство стандартов, 1975. - 6 с.
165. Фарафонтов В.Ф., Новохатин В.В. Взаимосвязь элементов структуры урожая различных сортов яровой пшеницы в условиях Северного Зауралья // Сибирск. вестн. с.-х. науки, № 1, 1977.
166. Федоров A.A. Использование питательных веществ почвы и удобрений сельскохозяйственными культурами // Сиб. вестник с.-х. науки. 1989. -№1. - С.7-11.
167. Федоров A.A. Новый подход к определению реально доступных растениям элементов питания в почве // Агрохимия. 2002. -№7.-С.32-39.
168. Федоров A.A. Оценка содержания в почве элементов минерального питания, доступных растениям // Агрохимия. 2002. - № 3.- С. 15 — 22.
169. Федоров A.A. Повышение плодородия почв и улучшение использования земель // Система ведения сельского хозяйства в Приморском крае. -Новосибирск, 1981. С. 26-54.
170. Федоров A.A. Принцип определения реально доступных растениям питательных элементов в почве. Уссурийск, 2001. - 47 с.
171. Федоров A.A. Система применения удобрений (практикум). Уссурийск: ПГСХА, 1998. - 169 с.
172. Федоров A.A. Система удобрения в севооборотах и агрохимическое обслуживание сельского хозяйства // Система ведения сельского хозяйства в Приморском крае на 1986 1990 годы. - Новосибирск, 1987. - С. 2139.
173. Федоров A.A. Удобрение для сельскохозяйственных культур // Система земледелия в Приморском крае. Новосибирск, 1990. - С. 30 - 37.
174. Федоров A.A. Фосфоритование почв юга Российского Дальнего Востока. Уссурийск, 2001. - 201 с.
175. Федоров A.A., Вахмистров Д.Б. Влияние соотношений между питательными элементами в среде на минеральный состав растений в связи с их избирательной способностью // Агрохимия. 1980. - № 8. - С. 93-101.
176. Федоров A.A., Вахмистров Д.Б. Раздельное определение оптимумов суммарной дозы N+P+K и соотношения N:P:K в удобрении. Сообщение 9. Продуктивность кукурузы // Агрохимия. 1985. - № 12. - С. 78-89.
177. Федоров A.A., Зайковская H.A., Хавкина Н.В. Азотфиксирующая способность сои и клевера // Пути повышения производства растительного белка.- М.-1984.- С. 83-92.
178. Федоров A.A., Зайковская H.A., Хавкина Н.В. Азотфиксирующая способность сои и клевера // Пути повышения производства растительногобелка: Тр. университета Дружбы народов им. Лумумбы. М., 1984. С. 83-92.
179. Федоров A.A., Басистый В.П. Теория и практика известкования кислых почв юга российского Дальнего Востока. Уссурийск, 2001. 164 с.
180. Федоров М.В. Биологическая фиксация азота атмосферы. М.: Сельхоз-гиз, 1952. 503 с
181. Федоровский Д.В. Расчет дозы удобрений по выносу питательных веществ урожаем //Химия в сел. хоз-ве. 1964. - № в.- С. 45-52.
182. Фрид A.C. Методологические подходы к оценке доступности веществ-почвы корням растений с помощью миграционной концепции // Агрохимия. -1996.-№5.- С. 89-99.
183. Фрид A.C. Миграционная концепция доступности веществ почвы кор-ням растений // Агрохимия. -1996.-№3.-С. 29-37.
184. Фрид A.C. Опыт экспериментальной оценки доступности веществ поч-вы корням растений на основе миграционной концепции // Агрохимия. -1996.-№6.-С. 36-46.
185. Церлинг В.В. Диагностика питания растений по их химическому анализу // Агрохимические методы исследования почв. М.: Наука, 1965.
186. Цильке P.A., Новикова В.А., Бородин М.Ф. Изменчивость признака открытого цветения у пшеницы в зависимости от сорта и метеорологических условий//Тр. СибНИИСХ. Т. 1 (16). - Омск, 1971.-С. 79-83.
187. Чесноков В.А. и др. О системе питания растений в гидропонике // Вопросы корневого питания растений. Л., 1968. - С.6 - 8.
188. Чижевский М.Г. Развитие теоретических основ земледелия // Земледелие.- 1967.-№ п.-С. 18-27.
189. Чичкин А., Тарабрин Н.В. Изучение эффективности минеральных удобрений в севообороте на обыкновенном черноземе при орошении /7 Результаты исследований в длительных опытах с удобрениями по зонам страны / Тр. ВИ-УА. М., 1976. - Вып 1. - С.42-48.
190. Шамра Гирдж Влияние соотношения N.P.K в питании ячменя на урожай и избирательную способность растений: Автореф. дис. канд. с.-х. наук-М., 1986.-С. 13.
191. Шарахимбаев Ж., Тазебеков Г.Т. Соотношение N:P:K в почве и растениях на разных горизонтах темно-каштановой почвы // Вести с.-х. науки Казахстана. 1974. - Вып. 12.
192. Шенявский A.JI. Оценка плодородия почвы методом гумусового баланса,-М.- 1979.- 27с.
193. Шконде Э.И. Роль сои в накоплении азота почв // Удобрение и урожай. -1957. -№> 1, С.31-35.
194. Dijkshoorn W.Jn.: 1-st Red. Conf. Of Jnt. Potash Jnst., Wexford, 1963. P. 43-62.
195. Homes M. Alimentation monirele ig uilibrie des vigetaux wetteren: Universa, 1961, v 1. P. 65.
196. Homes M. La solution de culture hudroponique. Agroctimica, 1963. V. 7, N2.- 101 p.
197. Homes M. Effect de la fumure complètement equilibree sur la production de plantes de grande culture. Pontif. ACAD. Sci. Ser. Varia, 1973, N 38, part. 2.-P. 1257- 1281.
198. Homes M., Van Schoor G.H.I. Rendement maximum susceptible d'etre obtenu dans des conditions definies de culture. Agroctimica, 1976. V. 20, N 415.-P. 339-347.
199. Richard L. problèmes de fumure minerale. Coton Fibres trop., 1958. - V. 13. N2.-P. 293-329.
200. Said J.M. Thesis Wageningen, 1959.
201. Scheiner O., Skinner J.J. Bot. Gaz.- 1911. -v. 50. -№ 1. P. 1.
202. Stout P.R. Owerstreet R. Soil chemistry in relation to inorganic nutrition of plants // Ann. Rew/ Plant Physio. 1950, P. 305-342.
www.dissercat.com
Оптимизация режима почвенного питания растений — Сельское хозяйство
Территория рекламы
Общий прирост урожайности с/х культур от внесения удобрений на 2/3 зависит от минеральных и на 1/3 от органических удобрений. Внесение минеральных удобрений - это прежде всего прямая компенсация потерь минеральных веществ почвы после извлечения их урожаем. Внесение минеральных удобрений непосредственно отражается на физических, химических и биологических свойствах почвы и может привести к ее оздоровлению или наоборот, к заболеванию.
Основное требование к подбору системы удобрений с/х культур - это оптимизация режима питания(, а не минимализация применения удобрений), удовлетворение потребностей растений в макро- и микроэлементах в течение вегетации, особенно в критические периоды роста и развития растений. Питательные вещества должны находиться в почве в доступном состоянии и в сбалансированном количестве. Это достигается поэтапным внесением удобрений: основного - до посева, стартовой дозы - при посеве и последующих подкормок в течение вегетации. Систему применения удобрений следует подбирать к каждой экосистеме с/х культур и каждой почвенной разности.
В настоящее время известно 17 химических элементов абсолютно необходимых растениям. Кроме углерода (C), кислорода (O) и водорода (H), которые растения получают из воздуха и воды, остальные 14 питательных элементов растения получают из почвы. Каждый почвенный питательный элемент жизненно важен и имеет свое агроэкологическое значение, но условно принято подразделять элементы приблизительно по потребляемому растениями количеству. Наиболее важными питательными элементами - макроэлементами - являются азот (N), фосфор (P) и калий (K), далее следуют кальций (Ca), магний (Mg), сера (S), и уже к микроэлементам относят железо (Fe), марганец (Mn), цинк (Zn), медь (Cu), бор (B), молибден (Mo), кобальт (Co), хлор (Cl). Перечисленные элементы нельзя заменить другими и при острой нехватке хотя бы одного из них, растения существовать не могут, поэтому их запасы в почве следует восстанавливать. Кроме перечисленных элементов растения используют в малых количествах многие другие элементы, необходимые для их физиологических процессов. В растениях обнаружено более 70 химических элементов.
При выращивании с/х культур наиболее часто и ярко проявляется дефицит макроэлементов: азота, фосфора и калия. Тем не менее, в настоящее время отмечается повсеместное обеднение пахотных земель микроэлементами и эта проблема все более обостряется. Применение микроэлементов способствует нормализации физиологических процессов в растениях, повышает их урожайность и снижает возможность поражения растений болезнями, вредителями и сорняками. Максимальная эффективность минеральных удобрений достигается при применении их вместе с органическими. Органические удобрения ослабляют отрицательное действие высоких концентраций минеральных удобрений и обеспечивают питание растений в течение всей вегетации.
ifreestore.net
Оптимизация минерального питания – значительный резерв в растениеводстве
Сегодня нередко приходится слышать от сельхозпроизводителей, что ради снижения себестоимости продукции они вынуждены экономить на необходимых для растениеводства ресурсах. Но как при этом вырастить урожай достаточного количества и, самое главное, качества? Огромный резерв для улучшения этих двух показателей – оптимизация минерального питания сельхозкультур.
Использование этого резерва позволяет уйти от практики, когда сельхозпроизводители вносят традиционные макроудобрения без сбалансированного внесения микроэлементов и в итоге зачастую не получают ожидаемого эффекта.Важно отметить, что оптимизация минерального питания сельхозкультур как агроприём до сих пор остается недооцененной в отечественном сельском хозяйстве. Если же обратиться к опыту зарубежных стран, то там питание растений балансируется не только по азоту, фосфору и калию, а по десятку макро- и микроэлементов и использование микроэлементов является обязательным приемом при выращивании практически всех сельскохозяйственных культур.В настоящее время на рынке имеется большое количество перспективных сортов растений. Однако в ходе их возделывания зачастую выясняется, что они не в состоянии полностью реализовать свой потенциал. Почему? Потому что почти половина сельхозугодий нашей страны деградирует по целому комплексу показателей. Снижение содержания микроэлементов в почве, например, связывают с сокращением объемов вывоза органических удобрений на поля. При недостатке микроэлементов в растениях нарушаются ключевые биологические процессы – дыхание, фотосинтез и другие, на 30% снижается усвоение растениями основных макроэлементов. Всё это приводит к снижению продуктивности полей, ухудшению качества урожая и, следовательно, к снижению рентабельности сельхозпредприятий.Рынок предлагает огромное множество препаратов, которые, как гласят аннотации, способны увеличить урожайность культур и качество получаемой продукции. Сельхозпроизводителям трудно определиться с выбором того или иного препарата. Основное действие большинства из них – обычная стимуляция роста растений.Основное количество комплексных микроудобрений, имеющихся сегодня на рынке, представлены западными производителями.Тем не менее и на рынке отечественных препаратов есть компании, которые предлагают качественные удобрения, содержащие микроэлементы. Например, компания «Волски Биохим», член Национального агрохимического союза РФ, отечественный разработчик, производитель и поставщик инновационных препаратов в сегментах растениеводства и животноводства. Собственные разработки компании сконцентрированы в сфере микроэлементных и гуминовых продуктов. Ключевая продукция «Волски Биохим» – жидкие комплексные микроэлементные удобрений Микромак® иМикроэл®. Их составы разработаны с учетом именно российских почвенно-климатических условий. Акцент сделан на высокие дозы микроэлементов и ассортимент, которые ориентированы на активацию ферментов растений, почвы и микроорганизмов. В состав удобрений входят 17 элементов питания: Cu, Zn, B, Mn, Fe, Mo, V, Co, Mg, Cr, Se, Ni, Li, S, N, P, K. Указанные препараты способны оптимизировать питательный режим растений на протяжении всей вегетации. Удобрение Микромак® предназначено для предпосевной обработки семян,Микроэл® – для некорневых подкормок.
Наука и практикаНаучно-исследовательская деятельность «Волски Биохим» осуществляется исследовательским подразделением и лабораторией компании совместно с ведущими биологическими ВУЗами и НИИ РФ. Удобрения Микромак® и Микроэл® разработаны на основе фундаментальных исследований, проводимых с 1961 г. на базе специальной научной исследовательской лабораторией по усвоению атмосферного азота (СНИЛУА) под руководством профессора М.И. Волского.Масштабность проведения научно-практической работы «Волски Биохим» трудно переоценить, как трудно найти какие-либо другие инновационные препараты, опыт применения которых был бы изучен на столь широкой территории, в разных природно-климатических зонах и на разных культурах. К настоящему времени компания сотрудничает с 30 научно-исследовательскими учреждениями и многими сельхозпредприятиями в разных регионах страны. При поддержке этих организаций проведено более 150 научных и производственных экспериментов, доказывающих положительный эффект действия препаратов.Один из масштабных экспериментов проводился в 2009 г. в Краснодарском крае на территории землепользования ООО «Холдинговая компания «Агрофирма Россия» под эгидой ВНИИ агрохимии имени Д.Н. Прянишникова (г. Москва) совместно с сотрудниками Кубанского государственного агроуниверситета. Зафиксировано, что в результате обработки семян препаратом Микромак® и двух некорневых подкормок препаратомМикроэл® урожайность озимой пшеницы повысилась на 12,2 ц/га. При этом увеличилось содержание клейковины, что способствовало повышению класса зерна с 4-го до 3-го.Специалисты «Волски Биохим» обобщили результаты всех испытаний с комплексными микроудобрениями, получив положительные выводы по разным показателям. Прибавка урожая в среднем составляет 25%. Улучшается и качество. В частности, содержание клейковины в пшенице возрастает на 3-4 процента.
Удобрения Микромак® и Микроэл® успешно используются во многих регионах страны. Сегодня общая площадь применения удобрений превышает полмиллиона гектаров.Мы начали эту публикацию с разговора о том, что в связи с постоянным ростом цен на азотные и другие макроудобрения внесение их в традиционных нормах становится малодоступным и экономически неоправданным для многих хозяйств. Комплекс удобрений Микромак® и Микроэл® при правильном использовании повышает эффективность работы фосфорных и калийных удобрений и снижает традиционное применение азотных удобрений благодаря повышению эффективности их использования, а также активизации процесса азотфиксации – источника дополнительных 30-40 кг биологического азота, усваиваемого на 100% на каждом гектаре посевов. В рамках апробации предлагаемого агротехнического приема были также проведены многочисленные испытания, которые доказали, что при использовании комплекса удобрений Микромак® и Микроэл® дозу азотных удобрений можно уменьшить на 30-50% без снижения урожайности (график 3). Метод сокращения внесения азотных удобрений при применении Микромак® и Микроэл® активно внедряется в крупнейших хозяйствах России.
В структуре всех затрат на агрохимикаты, а это в среднем более 2000 рублей на гектар, средства на покупку и использование инновационных препаратов составляют всего 282 рублей на га. Сюда входит стоимость предпосевной обработки семян преператом Микромак® и двух некорневых обработок препаратом Микроэл® по вегетации. Кроме того, микроудобрения очень технологичны в использовании – имеют жидкую препаративную форму и не требуют затрат на внесение, так как применяются совместно с пестицидами.Учитывая рост количественных и качественных показателей в производстве зерна, каждый рубль затрат на препараты Микромак® и Микроэл® приносит от 10-15 рублей дохода.Это и есть ответ на вопрос, который мы поставили в начале статьи. Перспектива сельского хозяйства заключается в применении тех инноваций, которые доступны любому сельхозпроизводителю и при этом высокоэффективны.
http://agroobzor.ru/ahim/a-123.html
Дата: 12.11.2012
www.agromaniya.ru
Янчевская Т. Г.. Оптимизация минерального питания растений
Монография посвящена физиолого-биохимической оптимизации комплекса факторов, обеспечивающих рост, развитие и продуктивность растений в агрофитоценозах (1-я часть), и интенсивной светокультуре в модельных системах на ионообменных субстратах (2-я часть).
Приведен обзор результатов по оптимизации минерального питания растений в полевых условиях с помощью динамического моделирования и на их теоретической основе созданы оптимальные искусственные корнеобитаемые среды с использованием природных и синтетических ионообменных материалов нового поколения. Рассмотрены физико-химические свойства современных ионитов, обеспечивающих избирательность обмена и интенсивный трансмембранный перенос ионов в клетках корней. Исходя из анализа многофакторных экспериментов, обоснованы научные представления о необходимых и достаточных условиях оптимизации минерального состава субстрата, произведены расчеты дозы и соотношения элементов с использованием куполообразной зависимости доза–эффект, отражающие достижение максимального параметра целевой функции – клубневого коэффициента размножения на примере культуры картофеля.
Предназначена для научных сотрудников в области физиологии растений, агрономов и сельскохозяйственных работников, аспирантов и студентов соответствующих вузов.
Содержание:
Принятые сокращения...... 3 Предисловие...... 4 Введение...... 7 ЧАСТЬ 1. ОПТИМИЗАЦИОННАЯ МОДЕЛЬ ВЫРАЩИВАНИЯ РАСТЕНИЙ В АГРОФИТОЦЕНОЗАХ...... 11 Глава 1. Принципы оптимизации условий развития растений...... 12 1. 1. Оптимизационные модели...... 12 1. 1. 1. Классификация моделей...... 15 1. 1. 2. Методы создания моделей. Метод переменных состояния...... 16 1. 1. 3. Практичность модели...... 18 1. 2. Определение границ системы и уровни продуктивности...... 20 1. 3. Перспективность экологизации моделей развития агрофитоценозов...... 23 Глава 2. Агрофизические свойства почв, обусловливающие оптимальное минеральное питание растений...... 25 2. 1. Компоненты почвенных субстратов как аккумуляторы элементов питания растений...... 25 2. 1. 1. Особенности минерального питания растений в зависимости от гранулометрического состава природных почв...... 26 2. 1. 2. Ионообменные процессы в почвах и их плодородие...... 27 2. 1. 3. Азот в питании растений и особенности его связывания частицами почвы различных размеров...... 32 2. 1. 4. Фосфор в питании растений и связывание его частицами почвы в зависимости от гранулометрического состава...... 40 2. 1. 5. Связывание калия частицами почвы и его роль в питании растений...... 43 2. 1. 6. Стабилизирующая роль ионов кальция и магния в клетках растений...... 50 Глава 3. Технические средства оптимизационной модели развития растений в агрофитоценозе...... 54 3. 1. Создание информационной базы модели...... 57 3. 1. 1. Субмодель корневого питания растений...... 58 3. 1. 2. Субмодель вегетативного развития (фенофаз)...... 62 3. 1. 3. Субмодель водного режима агрофитоценоза...... 64 3. 1. 4. Субмодель урожайности...... 65 3. 2. Экспертная система расчета доз удобрений под планируемый урожай...... 68 3. 3. Диалоговые компьютерные системы для оптимизации минерального питания растений...... 79 3. 3. 1. Реализация метода систематических вариантов Омеса на IBM PC...... 80 3. 3. 2. Реализация системы сбалансированного минерального питания на IBM PC (модель ARMAGRO)...... 81 3. 3. 3. Расширение возможностей метода систематических вариантов Омеса (модель BIOTOP)...... 84 Заключение к части 1...... 87 ЧАСТЬ 2. ФИЗИОЛОГО-БИОХИМИЧЕСКОЕ ОБОСНОВАНИЕ ОПТИМИЗАЦИИ УСЛОВИЙ ВЫРАЩИВАНИЯ РАСТЕНИЙ В ЗАЩИЩЕННОМ ГРУНТЕ...... 89 Глава 4. Оптимизация транспортных потоков минеральных ионов в клетках корней растений...... 91 4. 1. Роль соотношения минеральных ионов в среде культивирования для оптимального развития растений...... 91 4. 2. Концентрация минеральных ионов в питательной среде, их поглощение и усвоение растениями...... 94 4. 2. 1. Апопластный и симпластный путь транспорта ионов...... 95 4. 2. 2. Транспорт ионов в клетки в зависимости от их концентрации в наружной среде...... 96 4. 3. Трансмембранный перенос ионов клетками корней растений...... 102 4. 3. 1. Транспортные системы клеток для неорганических ионов...... 106 4. 3. 2. Ацидофицирующая способность растений...... 111 4. 3. 3. Корреляция между ацидофицирующей способностью растений, биологической продуктивностью и кислотоустойчивостью...... 112 4. 4. Транспорт ионов в клетки корней картофеля при клональном микроразмножении...... 129 4. 4. 1. Характеристики транспортных систем окисленной и восстановленной форм азота в растительных клетках...... 130 4. 4. 2. Трансмембранный перенос ионов водорода и транспорт азота через клетки корней пробирочных растений картофеля...... 134 Выводы к главе 4...... 146 Глава 5. Теоретические основы оптимизации роста и развития растений в искусственных условиях...... 151 5. 1. Система оптимизации минерального питания на инертном субстрате...... 156 Выводы к главе 5...... 186 Глава 6. Выращивание растений на искусственных корнеобитаемых средах...... 187 6. 1. Композиционный состав искусственных почв и культура растений...... 188 6. 1. 1. Культура растений на почвенных грунтах...... 189 6. 1. 2. Беспочвенная культура растений...... 191 6. 1. 3. Культура растений на водных растворах...... 192 6. 1. 4. Воздушно-капельная культура растений...... 195 6. 1. 5. Культура растений на субстратной гидропонике – агрегатопоника...... 196 6. 1. 6. Культура растений на ионитопонике...... 200 Выводы к главе 6...... 202 Глава 7. Ионообменные субстраты как универсальная искусственная почва...... 203 7. 1. Ионообменные свойства ионитных компонентов субстрата...... 204 7. 1. 1. Общие свойства ионообменных смол...... 207 7. 1. 2. Ионообменное равновесие...... 214 7. 1. 3. Сильнокислотные катионообменные смолы...... 218 7. 1. 4. Слабокислотные катионообменные иониты...... 220 7. 1. 5. Сильноосновные анионообменные смолы...... 221 7. 1. 6. Слабоосновные анионообменные иониты...... 223 7. 1. 7. Недостатки ионообменных субстратов старого образца...... 223 7. 2. Теоретические подходы к созданию оптимизированного ионообменного субстрата...... 224 7. 2. 1. Оптимизация ионообменных процессов...... 225 7. 2. 2. Оптимизация ионообменных технологических задач...... 227 7. 2. 3. Экспериментальные подходы к разработке оптимального субстрата, сбалансированного по минеральным элементам питания...... 228 7. 3. Модификация синтетических ионообменных почв природными компонентами...... 230 7. 3. 1. Природные цеолиты – высокопродуктивные компоненты ионообменных субстратов...... 230 7. 3. 2. Инертные наполнители как компонент ионообменных субстратов...... 235 7. 4. Получение искусственной почвы с заданными соотношениями ионов...... 236 7. 5. Верификация продукционного потенциала ионообменных субстратов нового поколения...... 246 7. 5. 1. Развитие овощных зеленных культур и их продуктивность на ионообменных субстратах...... 246 7. 5. 2. Развитие безвирусных растений картофеля на ионообменном субстрате нового поколения...... 251 7. 5. 2. 1. Общая характеристика корнеобитаемой среды Триона...... 251 7. 5. 2. 2. Развитие безвирусных растений картофеля на ионообменном субстрате Триона и Триона-2...... 252 Выводы к главе 7...... 261 Глава 8. Оптимизация вегетативного размножения картофеля в искусственных условиях...... 262 8. 1. Теоретические основы моделирования и оптимизации фитоонтогенеза в искусственных условиях...... 263 8. 1. 1. Вегетативное размножение путем микроклонирования...... 267 8. 1. 1. 1. Традиционный способ размножения пробирочных растений in vitro...... 267 8. 2. Оптимизация процесса микроклонирования растений картофеля в условиях in vivo...... 271 8. 2. 1. Морфофизиологические особенности развития микроклонов пробирочных растений in vitro...... 274 8. 2. 2. Особенности развития регенерантов из черенков различных участков стебля пробирочных растений in vivo...... 281 8. 3. Теоретические подходы к оптимизации процесса развития рассады картофеля при разном уровне минерального питания и интенсивности света...... 295 8. 3. 1. Оптимизация светового режима для получения рассады картофеля в условиях in vivo...... 304 8. 3. 2. Световой режим для оптимизации морфообразовательного процесса картофеля in vivo на ионообменных субстратах...... 322 8. 4. Ретардантный эффект светового режима...... 330 Выводы к главе 8...... 344 Глава 9. Оптимизация влажности ионообменного субстрата...... 345 9. 1. Влияние водного потенциала и влажности субстрата на процессы ризогенеза in vivo картофеля...... 348 9. 2. Зависимость коэффициента размножения картофеля от характеристик влажности субстрата...... 354 Выводы к главе 9...... 364 Глава 10. Технологические аспекты оптимизации развития растений картофеля для первичного семеноводства...... 365 10. 1. Биотехнические устройства, оптимизирующие условия выращивания растений in vivo...... 366 10. 2. Промышленный способ ускоренного размножения исходного материала для первичного семеноводства картофеля...... 369 10. 2. 1. Получение рассады...... 376 10. 2. 2. Получение мини-клубней...... 381 10. 2. 3. Сортовые особенности развития и продуктивность картофеля разной спелости, реализованные по ионообменной технологии...... 389 10. 2. 4. Биотехнологические приемы регуляции физиологических процессов при размножении картофеля как элемент технологии...... 392 10. 2. 5. Частичный съем мини-клубней в процессе вегетации как элемент технологии...... 405 10. 3. Иммуноферментная и гель-электрофоретическая характеристики регенерантов...... 408 10. 4. Контроль продуктивности и микрофлоры в ионообменном субстрате при многоразовом использовании...... 411 Выводы к главе 10...... 416 Заключение к части 2...... 417 Литература...... 421
Издательство: "Белорусская наука"
ISBN: 9789850817686
dic.academic.ru









