
СОМАТИЧЕСКИЕ МУТАЦИИ В ПРИРОДЕ. Мутации растений
Мутации | Биология
Что такое мутация?
Под мутацией понимают изменение количества и структуры ДНК в клетке или у организма. Другими словами, мутация — это изменение генотипа. Особенностью изменения генотипа является то, что это изменение в результате митоза или мейоза может быть передано следующим поколениям клеток.
Чаще всего под мутациями понимают небольшое изменение в последовательности нуклеотидов ДНК (изменения в одном гене). Это так называемые генные мутации. Однако кроме них существуют хромосомные и геномные, когда изменения затрагивают крупные участки ДНК, или меняется количество хромосом.
В результате мутации у организма внезапно может появиться новый признак.
Мысль, что именно мутация является причиной появления новых передающихся через поколения признаков, была впервые высказана Гуго де Фризом в 1901 году. Позже мутации у дрозофилы были изучены Т. Морганом и сотрудниками его школы.
Мутация — вред или польза?
Мутации, происходящие в «незначащих» («молчащих») участках ДНК, не изменяют признаки организма и могут спокойно передаваться из поколения в поколение (на них не будет действовать естественный отбор). Такие мутации можно считать нейтральными. Также нейтральными являются мутации, когда участок гена заменяется на синонимичный. При этом, хотя последовательность нуклеотидов в определенном участке и будет отличаться, но синтезироваться будет такой же белок (с той же последовательностью аминокислот).
Однако мутирование может затронуть значащий ген, изменить аминокислотную последовательность синтезируемого белка, а, следовательно, вызвать изменение признаков организма. В последствии, если концентрация мутации в популяции достигнет определенного уровня, то это приведет к изменению характерного признака всей популяции.
В живой природе мутации возникают как ошибки в ДНК, поэтому все они априори вредны. Большинство мутаций понижают жизнеспособность организма, вызывают различные заболевания. Мутации, возникающие в соматических клетках, не передаются следующему поколению, но в результате митоза образуются дочерние клетки, составляющие ту или иную ткань. Нередко соматические мутации приводят к образованию различных опухолей и других заболеваний.
Мутации, возникающие в половых клетках, могут быть переданы следующему поколению. В стабильных условиях внешней среды почти все изменения генотипа оказываются вредными. Но если условия среды изменились, то может оказаться, что ранее вредная мутации станет полезной.
Например, мутация, вызывающая образование коротких крыльев у какого-нибудь насекомого, скорее всего будет вредна в популяции, живущей в местах, где нет сильного ветра. Данная мутация будет сродни уродству, заболеванию. Обладающие ею насекомые с трудом будут находить партнеров для спаривания. Но если на местности начнут дуть более сильные ветры (например, в результате пожара участок леса был уничтожен), то насекомых с длинными крыльями будет сносить ветром, им будет тяжелее перемещаться. В таких условиях преимущество могут получить короткокрылые особи. Они чаще длиннокрылых будут находить партнеров и пищу. Через некоторое время в популяции окажется больше короткокрылых мутантов. Таким образом, мутация закрепится и превратится в норму.
Мутации лежат в основе естественного отбора и в этом их основная польза. Для организма же подавляющее число мутаций — это вред.
Почему возникают мутации?
В природе мутации возникают случайно и спонтанно. То есть любой ген в любой момент времени может мутировать. Однако частота мутаций у разных организмов и клеток различна. Например, она связана с продолжительностью жизненного цикла: чем он короче, тем мутации возникают чаще. Так у бактерий мутации возникают намного чаще, чем у организмов-эукариот.
Кроме спонтанных мутаций (случающихся в естественных условиях) бывают индуцированные (человеком в лабораторных условиях или неблагоприятными условиями среды) мутации.
В основном мутации возникаю в результате ошибок при репликации (удвоении), репарации (восстановлении) ДНК, при неравном кроссинговере, неправильном расхождении хромосом в мейозе и др.
Так в клетках постоянно происходит восстановление (репарация) поврежденных участков ДНК. Однако если в следствие различных причин механизмы репарации нарушаются, то ошибки в ДНК будут оставаться и накапливаться.
Результатом ошибки при репликации становится замена одного нуклеотида в цепочке ДНК на другой.
Что вызывает мутации?
Повышенный уровень мутаций вызывает рентгеновское излучение, ультрафиолетовые и гамма-лучи. Также к мутагенам относятся α- и β-частицы, нейтроны, космическое излучение (все это частицы, обладающие высокой энергией).
Мутаген — это то, что способно вызывать мутацию.
Кроме различных излучений, мутагенным действием обладают многие химические вещества: формальдегид, колхицин, компоненты табака, пестициды, консерванты, некоторые лекарственные препараты и др.
biology.su
СОМАТИЧЕСКИЕ МУТАЦИИ В ПРИРОДЕ | Институт радиобиологии НАН Беларуси
Поразительное свойство живых организмов — это высокая степень мутабильности генов.
Мутации – это стойкие внезапно возникшие изменения структуры наследственного материала на различных уровнях его организации, приводящие к изменению тех или иных признаков организма. Термин «мутация» введен в науку Де Фризом.
Мутации возникают в клетках любых тканей многоклеточного организма и на различных стадиях его развития. По типу клеток, в которых мутации произошли, различают: генеративные и соматические мутации [1].
Генеративные мутации возникают в половых клетках, не влияют на признаки данного организма, проявляются только в следующем поколении.
Соматические мутации возникают в соматических клетках, проявляются у данного организма и не передаются потомству при половом размножении. Мутации, возникающие в соматических клетках, наследуются дочерними клетками, которые образуются в процессе митотических делений. Фенотипические последствия таких изменений проявляются только у самой мутантной особи и только в том случае, если возникшие мутации препятствуют осуществлению специфических функций, свойственных данной клетке. Соматические мутации могут содержаться не во всех клетках организма, т.е. нормальные и мутантные клетки сосуществуют у одного индивидуума, что приводит к мозаицизму — наличию в организме клеток, отличающихся по своему генотипу и его фенотипическим проявлениям от других клеток этого же организма. Сохранить соматические мутации можно только путем бесполого размножения (прежде всего вегетативного).
Первая информация о соматических мутациях (они же почковые вариации или спорты (sport)) у многолетних, главным образом, древесных (плодово-ягодных и декоративных) растений появилась в первой половине 19-го века. Обобщивший ее Чарлз Дарвин (1868) посвятил этому вопросу целую главу в своем знаменитом труде «The Variation of Animals and Plants under Domestication»,
По адаптивному значению выделяют: полезные, вредные (летальные, полулетальные) и нейтральные мутации [2]. По характеру проявления мутации могут быть доминантными и рецессивными. Если доминантная мутация является вредной, то она может вызвать гибель ее обладателя на ранних этапах онтогенеза. Рецессивные мутации не проявляются у гетерозигот, поэтому длительное время сохраняются в популяции в «скрытом» состоянии и образуют резерв наследственной изменчивости. При изменении условий среды обитания носители таких мутаций могут получить преимущество в борьбе за существование.
В зависимости от того, выявлен ли мутаген, вызвавший данную мутацию, или нет, различают индуцированные и спонтанные мутации. Обычно спонтанные мутации возникают естественным путем, индуцированные — вызываются искусственно. В зависимости от уровня наследственного материала, на котором произошла мутация, выделяют: генные, хромосомные и геномные мутации.
Соматические мутации по своей природе ничем не отличаются от генеративных. Различие состоит лишь в проявлении и методах их обнаружения. Чем раньше в онтогенезе возникает соматическая мутация, тем больше оказывается участок ткани, несущие данную мутацию, и чем позднее — тем меньше. Соматическая мутация проявляется мозаично. Особи, несущие участки мутантной ткани, называют мозаиками, или химерами. В силу диплоидности набора хромосом в клетках соматической ткани, проявление мутации возможно только в тех случаях, когда мутантная аллель оказывается доминантной или будет рецессивна и будет находиться в гомозиготном состоянии.
Различий в частоте возникновения соматических и генеративных мутаций не обнаружено. Однако есть факты, что ряд генов мутирует с разной скоростью на разных стадиях онтогенеза. Так если у растений дельфиниума ген лавандовой окраски мутирует на поздней стадии развития цветка в лепестках встречаются одиночные клетки с измененной окраской, а при возникновении той же мутации на ранней стадии она может затрагивать большие по размеру участки — половину лепестка или даже целый цветок.
Ярким примером соматической мутации является окраска шерстного покрова у овцы: черное пятно на фоне коричневой окраски. Эта мутация могла проявиться либо как доминантная, либо как рецессивная при потере части или всей гомологичной хромосомы.
Подобные явления часто встречаются у самцов дрозофилы, у которых иногда часть глаза имеет красные фасетки, а часть — белые. Эта мозаичность связана с возникновением рецессивной мутации в локусе white половой хромосомы во время развития имагинальных дисков глаз. Но появляется она не только у самцов, у которых этот ген в Х-хромосоме находится в гомизиготном состоянии, но и у гетерозиготных самок в силу утраты целой хромосомы, несущей доминантную аллель w+, или потери части хромосомы (дефишенси), несущей ту же аллель. В этом случае рецессивная аллель, теперь уже находясь в гомозиготном состоянии, также может проявиться, но эта химерность будет не следствием мутации гена, а следствием изменений в числе или структуре хромосом.
Исследование соматических мутации в настоящее время приобретает важное значение для изучения причин возникновения рака у человека и животных. Предполагают, что ряд злокачественных опухолей возникает по типу соматических мутаций. Соматические мутации имеют прямое отношение также к выяснению причин старения человеческого организма, так как с возрастом может происходить накопление физиологических мутаций в популяции соматических клеток различных органов. Считают, что очень высокая частота заболеваний раком у людей старшего возраста обусловлена в какой-то мере накоплением соматических мутаций.
Наследственные заболеваний человека связаны с мутациями, приводящими к изменению уровней экспрессии генов [3]. Мутации в определенных генах нарушают функционирование биохимических систем, что приводит к развитию соответствующих патологических состояний организма (рисунок 1).
![Рисунок 1 - Генетические последствия мутаций, происходящих в геноме соматических клеток человека на разных стадиях эмбриогенеза Черным цветом закрашены места лока-лизации клонов мутантных клеток разных размеров в организме человека [3].](/800/600/http/www.irb.basnet.by/wp-content/uploads/2017/05/1.png) Рисунок 1 — Генетические последствия мутаций, происходящих в геноме соматических клеток человека на разных стадиях эмбриогенеза Черным цветом закрашены места лока-лизации клонов мутантных клеток разных размеров в организме человека [3].Если мутации происходят в геноме клеток зародышевой линии человека, все соматические клетки организма-потомка, который развивается из мутантной зиготы, образовавшейся от слияния мутантных гамет, будут содержать эту мутацию. Чем позже в онтогенезе возникает соматическая мутация, тем меньше размер клона мутантных клеток во взрослом организме. Если мутация доминантна, то возникает наследственное заболевание. Если мутация рецессивна, можно говорить о предрасположенности организма — гетерозиготы к соответствующему заболеванию и носительстве мутантного гена.
Рисунок 1 — Генетические последствия мутаций, происходящих в геноме соматических клеток человека на разных стадиях эмбриогенеза Черным цветом закрашены места лока-лизации клонов мутантных клеток разных размеров в организме человека [3].Если мутации происходят в геноме клеток зародышевой линии человека, все соматические клетки организма-потомка, который развивается из мутантной зиготы, образовавшейся от слияния мутантных гамет, будут содержать эту мутацию. Чем позже в онтогенезе возникает соматическая мутация, тем меньше размер клона мутантных клеток во взрослом организме. Если мутация доминантна, то возникает наследственное заболевание. Если мутация рецессивна, можно говорить о предрасположенности организма — гетерозиготы к соответствующему заболеванию и носительстве мутантного гена.
Организм, у которого действие рецессивной мутации маскируется функционированием полноценного аллеля, фенотипически выглядит нормальным, однако имеет больше шансов дать больное потомство в браке с носителем такого же мутантного гена. Кроме того, может произойти соматическая мутация в соответствующем аллельном гене соматических клеток, что станет причиной развития приобретенного генетического заболевания. Примером таких заболеваний может служить ретинобластома [3].
У организмов, размножающихся исключительно половым путем и имеющих раннее обособление зачаткового пути, соматические мутации не играют роли в эволюции и не представляют какой-либо ценности для селекции. Но у тех организмов, у которых есть бесполое размножение, соматические мутации могут иметь огромное значение, особенно в селекции, так как у таких форм из соматической ткани развиваются половые клетки. Так, например у плодовых и ягодных вегетативно размножаемых растений любая соматическая мутация может дать растение и целый клон с новым мутантным признаком.
Одним из видов соматических мутаций у растений являются почковые мутации, возникающие в меристемных клетках точки роста стебля. В этом случае весь побег, развившийся из этой клетки, будет нести мутантный признак. Многие сорта плодовых растений, винограда, картофеля являются соматическими мутантами. Эти сорта сохраняют свои свойства, если их воспроизводят вегетативным путем, к примеру, прививая обработанные мутагенами почки (черенки) в крону немутантных растений; таким путем размножают, к примеру, бессемянные апельсины. Почковые мутации были известны давно и назывались спортами; от такого спорта И. В. Мичурин получил сорт яблони, названный им Антоновка 600-граммовая
Соматические мутации используют для изучения частоты возникновения видимых мутаций. По мозаичности проявления мутантного признака в тканях можно обнаруживать соматические мутации вплоть до одиночных мутантных клеток. Растения чая в природных условиях характеризуются высокой пластичностью. При этом отмечаются как отдельные модификации, проявляющиеся на кусте, так и целиком измененные формы [4]. Спектр соматических мутаций представлен морфологическими (отклонения по длине, ширине, форме, морфологии листа, длине междоузлий), физиологическими (изменения окраски листа, вегетационного периода, генеративной активностью и урожайности) и пластидными (секториальные и периклинальные химеры) модификациями. Интересны формы с измененной фотосинтетической активностью, улучшенными биохимическими и органолептическими показателями [5]. При анализе растений сухумского района был установлен сорт с наибольшей частотой как соматических, так и структурных мутаций [6]. При этом кариологический анализ сорта и мутантных форм имели стандартный диплоидный набор хромосом (2n=30). В итоге в Сухумском районе были отобраны целиком измененные формы и выделены 23 мутантные формы чая с комплексом ценных признаков (урожайность и биохимические показатели).
Высокой пластичностью характеризуется также карельская береза, которая представлена группой переходных морфологических форм и не имеет четкого дендрологического описания. В естественных условиях B. pendula var. carelica представлена различными вариантами, включая деревья высотой до 25 м или сильно ветвящиеся кустарники с приподнимающимися стволиками до 3 м высотой. Биологические особенности карельской березы связаны с наличием аномальных процессов при делении и дифференцировки камбиальных клеток [7].
При выращивании в условиях Беларуси плантационные культуры березы карельской, представляют собой совокупность морфологических форм: высокоствольные (1а — крупноузорчатая и 1б — шаровидноутолщенная), короткоствольные (11а — пятнистоузорчатая и 11б — лироствольная), кустовидная, кустарниковая и безузорчатая. Соотношение узорчатых и безузорчатых растений в насаждениях березы карельской примерно одинаковое 50:50. Такое широкое формовое разнообразие по росту в высоту, диаметру, а, соответственно, и узорчатости древесины указывает на высокую пластичность данной березы, ее способность произрастать в различных условиях [7].
ВЕДЬМИНЫ МЕТЛЫ, СУВЕЛИ И КАПЫ У ДРЕВЕСНЫХ РАСТЕНИЙ
Ведьмины мётлы — фрагменты кроны растения с аномальным морфогенезом (рисунок 2). Проявляется как образование многочисленных тонких побегов, чаще бесплодных, прорастающих из спящих почек. Обильное ветвление приводит к образованию множества укороченных ветвей с недоразвитыми листьями, которые часто формируют плотные скопления в виде шара или бесформенные. Дерево с ведьминой метлой представляет собой химеру [9]. При этом, отношения между компонентами химеры всегда конкурентные. Метлы, образующиеся в кронах взрослых, вполне сформировавшихся, деревьев обычно недолговечны, а возникшие в верхней части кроны сравнительно молодых растений часто подавляют произведшую их крону и полностью замещают ее собой [10].
Рисунок 2 – Ведьмины метлы на сосне обыкновенной (А), лиственнице европейской (Б), берёзе (В) и ели обыкновенной (Г) [9]
Классической точкой зрения на происхождение ведьминой метлы является заражение растений ржавчинными грибами, микоплазмами, грибами рода Тафрина (на вишне, сливе, березе) или вирусами (на картофеле). По другой версии появление ведьминой метлы связано с инфицированием растений группой бактерий — фитоплазмой. Переносчиком которых являются насекомые. Однако кроме патологических встречаются ведьмины метлы с нормальной жизнеспособностью, высокой долговечностью и полным отсутствием каких-либо патогенов или следов их жизнедеятельности (рисунок 3). Причинами их возникновения являются соматические мутации. Такие нарушения играют решающую роль в видообразовании и выведении сортов.
![Рисунок 3 – 5-летние ветви Abies sibirica: НК-П – контроль к паразитарной ведьминой метле; ВМ-П – ведьмина метла паразитарная, ВМ-М – ведьмина метла мутационная; НК-М – контроль к мутационной ведьминой метле [11].](/800/600/http/www.irb.basnet.by/wp-content/uploads/2017/05/3.png) Рисунок 3 – 5-летние ветви Abies sibirica: НК-П – контроль к паразитарной ведьминой метле; ВМ-П – ведьмина метла паразитарная, ВМ-М – ведьмина метла мутационная; НК-М – контроль к мутационной ведьминой метле [11].Клоны ведьминых мётел хвойных растений используются в селекции для создания декоративных сортов так как отличаются ценными свойствами: высокой жизнеспособностью, замедленным ростом, скороплодностью, обильным плодоношением [12]. При этом в семенном потомстве ведьминых мётел, наблюдается расщепление сеянцев на растения с обычным габитусом и обильно ветвящимися карликами [13]. Установлено, что у растений сосны обыкновенной с ведьминой метлой число геномных и хромосомных мутаций выше, чем у нормальных деревьев. Они содержат триплоидные и тетраплоидные клетки, отличаются большей активностью ядрышкообразующих зон, а также нарушением структуры и функций ядрышек в интерфазных ядрах. В митозе отмечены отстающие и хаотически расходящиеся хромосомы, мосты, с-митоз, а в метафазе митоза обнаружены аномальные формы ядрышек [14].
Рисунок 3 – 5-летние ветви Abies sibirica: НК-П – контроль к паразитарной ведьминой метле; ВМ-П – ведьмина метла паразитарная, ВМ-М – ведьмина метла мутационная; НК-М – контроль к мутационной ведьминой метле [11].Клоны ведьминых мётел хвойных растений используются в селекции для создания декоративных сортов так как отличаются ценными свойствами: высокой жизнеспособностью, замедленным ростом, скороплодностью, обильным плодоношением [12]. При этом в семенном потомстве ведьминых мётел, наблюдается расщепление сеянцев на растения с обычным габитусом и обильно ветвящимися карликами [13]. Установлено, что у растений сосны обыкновенной с ведьминой метлой число геномных и хромосомных мутаций выше, чем у нормальных деревьев. Они содержат триплоидные и тетраплоидные клетки, отличаются большей активностью ядрышкообразующих зон, а также нарушением структуры и функций ядрышек в интерфазных ядрах. В митозе отмечены отстающие и хаотически расходящиеся хромосомы, мосты, с-митоз, а в метафазе митоза обнаружены аномальные формы ядрышек [14].
Специфичная, оригинальная форма растений придает им декоративный вид, что используется в селекции, а получаемые сорта используются в ландшафтном дизайне и при озеленении населенных пунктов (рисунок 4 и 5).
Рисунок 4 – Сорта кедра, созданные на основе прививки ведьминой метлой: А –сорт «Биосфрера», Б – сорт «Идеал», В – сорт «Рекордистка», Г – сорт «Тамагочи» [15]
![Рисунок 5 – Прививки «ведьминой метлы» сосны обыкновенной (слева) и сосны горной «Мопс» (справа) [16]](/800/600/http/www.irb.basnet.by/wp-content/uploads/2017/05/5.png) Рисунок 5 – Прививки «ведьминой метлы» сосны обыкновенной (слева) исосны горной «Мопс» (справа) [16]Подобно ведьминым метлам на древесных растениях наблюдаются шаровидные разрастания локальных участков стебля, возникающие при неспецифическом нарушении ростовых процессов в камбии [17]. К их числу относятся сувели – гладкие, шаровидные или близкие к шаровидным наросты на стволах и ветвях древесных растений
Рисунок 5 – Прививки «ведьминой метлы» сосны обыкновенной (слева) исосны горной «Мопс» (справа) [16]Подобно ведьминым метлам на древесных растениях наблюдаются шаровидные разрастания локальных участков стебля, возникающие при неспецифическом нарушении ростовых процессов в камбии [17]. К их числу относятся сувели – гладкие, шаровидные или близкие к шаровидным наросты на стволах и ветвях древесных растений
Широко распространены в природе капы – тоже шаровидные или почти шаровидные наросты на стволах (чаще в комлевой части) и ветвях деревьев, поверхность капов покрыта многочисленными спящими почками. Скопление придаточных спящих почек и является причиной образования наплывов древесины. Анатомическое строение древесины капов, как и сувелей, сохраняет черты видоспецифичности. Способность образовывать капы, по мнению большинства исследователей [18], является адаптацией к условиям, затрудняющим семенное возобновление. Спящие почки, дающие в определенных условиях побеги, обеспечивают вегетативное возобновление.
Сувели, капы, ведьми метлы являются аномальностью строения, что проявляется во внешней морфологии, в то время как анатомические особенности проводящих элементов в основном сохраняют видовую специфику, меняются преимущественно их количественные и топологические особенности. Например, во вторичной ксилеме сувелей, капов и множества других наростов, образующихся на стеблях древесных растений под влиянием различных агентов и факторов среды, структурные изменения касаются в основном пространственной организации как паренхимных, так и прозенхимных клеток [9].
Анатомические исследования [19] показали, что микроструктурные изменения инициируются в первичных лучах возле сердцевины и внешнее морфологическое проявление соматической мутации зависит от положения мутантной клетки в апикальной меристеме побега. Если мутация произошла на верхушке меристематической зоны, вблизи апикальной инициали, — она, вероятнее всего, «породит» ведьмину метлу; если же мутантная клетка не начнет сразу делиться или возникнет ниже самой верхушки конуса нарастания побега, то в процессе роста и дифференциации она с большой вероятностью окажется в прокамбии и будет служить началом образования нароста на стебле [9].
Подобные аномалии описаны у растений в зоне отчуждения вследствие аварии на Чернобыльской атомной станции в первые годы [20] и наблюдаются в настоящее время (рисунок 6).
Рисунок 6 – Изменение морфологических форм растений, произрастающих на лугу ур. Учитель (внп. Масаны): А – изменение хлорофильной пигментации хвои сеянцев сосны, Б-Г – морфозы сосны, Д – искривление стебля Oenothera biennis L.(фотографии из личного архива Н.В. Шамаль)
Литература
- http://www.activestudy.info/generativnye-i-somaticheskie-mutacii-2/
- http://studopedia.info/5-45142.html
- http://humbio.ru/humbio/genexp/001b2690.htm
- Керкадзе И.Г. Некоторые вопросы генетики чайного растения. Сообщение VIII. Соматическая мутация и клоновая селекция чая. // Субтропические культуры, 1983. № 1. С. 42-45.
- Борисова Е.М. Роль сортов и новых технологий в интенсивном садоводстве. // Сельскохозяйственная биология, 2003, № 5: С. 124-126.
- Гвасалия М.В. Частота и спектр хромосомных аберраций у сортов и форм чая при спонтанном мутагенезе. // Сельскохозяйственная биология. 2014. № 3. С. 65-69
- Новицкая Л.Л. Карельская береза: механизмы роста и развития структурных аномалий. Петрозаводск: Verso, 2008. 144 c.
- Барсукова Т.Л. Изменчивоть березы карельской по морфологическим формам в культурах разного возраста в условиях Беларуси // Структурные и функциональные отклонения от нормального роста и развития растений под воздействием факторов среды: Материалы Международной конференции. Петрозаводск, Карельский научный центр РАН. 2011. С. 29-33.
- Коровин В.В. Структурные аномалии стебля древеных растений: итоги наблюдений // Структурные и функциональные отклонения от нормального роста и развития растений под воздействием факторов среды: Материалы Международной конференции. Петрозаводск, Карельский научный центр РАН. 2011. С. 131-136.
- https://ru.wikipedia.org/wiki/
- Ямбуров М.С. Морфологические особенности мутационной и паразитарной «ведьминых мётел» пихты сибирской //Вестник Томского государственного университета. Биология. – 2009. № 329. – С. 246-250.
- Ямбуров М. С., Горошкевич С. Н. «Ведьмины мётлы» кедра сибирского как спонтанные соматические мутации: встречаемость, свойства и возможности использования в селекционных программах // Хвойные бореальной зоны. — 2007. — Т. XXIV, № 2—3.
- Ямбуров М. С., Горошкевич С. Н., Третьякова И. Н., Хихлова О. В., Лукина А. В. Ведьмины мётла мутационного типа как перспективный источник для получения новых декоративных форм хвойных растений // Вестник Иркутской сельскохозяйственной академии. — 2011. — Т. 44, № 4. — С. 153-160.
- Муратова Е. Н., Седельникова Т. С. Геномные и хромосомные мутации у сосны обыкновенной (Pinus sylvestris L.) в экстремальных условиях произрастания // Хвойные бореальной зоны. — 2004. — С. 128—140.
- http://kedr.forest.ru
- Усольцев В/А. «Ведьмину метлу» – на улицы наших городов // Экопотенциал. – 2013. № 3–4. – С. 111-115.
- Коровин В.В., Курносов Г.А. Структурные аномалии: случайность или … // Лесной вестник. 2009. — № 1. С. 26-31.
- James Lignotubers and burls/ Their structure, function and ecological significance in Mediterranean ecosystem. // Bot. Rev., 1984. V. 50, N 3. P. 225–266
- Коровин В.В. Аномальные, предположительно мутантные, изменения морфологии сосны обыкновенной в ленточных борах Алтая // Тез.докл. Междун.научно-практической конф. Воронеж. 1996. С. 13-14.
- Радиоактивное загрязнение растительности Беларуси (в связи с аварией на Чернобыльской АЭС) / отв. ред. В.И Парфенов, Б.И. Якушев // Мн.: Навука i тэхнiка, 1995 – 581 с.
 Шамаль Наталья Владимировна
Шамаль Наталья Владимировна© Наталья Шамаль, старший научный сотрудник лаборатории моделирования и минимизации антропогенных рисков
e-mail: [email protected]
www.irb.basnet.by
Мутации у растений - Справочник химика 21
В последнее время химия проникает и в область селекции растений и семеноводства с.-х. культур. Здесь уже получают применение химические средства, вызывающие мутации у растений (см. Мутагены химические), средства, вызывающие полиплоидию у растений, стерилизаторы пыльцы нри гибридном семеноводстве кукурузы и других культур, основанном на явлении гетерозиса. В практическом семеноводстве можно отметить приемы химической сушки семян и их очистки. [c.335] Летальные мутации у растений выражаются, например, в неспособности образовывать корни, гибели зародыша, альбинизме и т. д. [c.204]Хорошо известно, что мутации, происходившие в процессе эволюции, обусловили индивидуальность разных видов растений и животных. Эволюция - это процесс постепенного изменения, в результате которого появляются новые виды, лучше приспособленные к условиям окружающей среды, чем их предки. При помощи процесса спонтанных мутаций виды способны адаптироваться к изменениям окружающей среды. Неспособность к адаптации приводит к вымиранию вида и замене его другим, более приспособленным. Спонтанные или естественно возникающие мутации являются важнейшими движущими силами эволюционного процесса. Причины этих мутаций еще изучены не до конца, хотя известно, что химические вещества, включая половые гормоны, УФ- и ионизирующее излучение, а также высокая температура могут повышать частоту возникновения мутаций у растений и животных. Относительный вклад этих факторов в возникновение спонтанных мутаций пока неизвестен. [c.100]
Мутации у растений могут происходить скачками, сразу создавая различия на уровне семейств, а мутации у насекомых могут привести к появлению дополнительных конечностей, т. е. признаков, свойственных представителям других классов беспозвоночных [c.371]
Далее Г. А. Надсон отмечает, что в 1920 г. им была обнаружена изменчивость микробов под влиянием радиевых и рентгеновых лучей, происходящая скачкообразно. Эти скачкообразные изменения наследственны, и для отличия от мутаций у растений и животных автор предложил называть их сальтациями (от латинского saltus — скачок). Этот термин не удержался в литературе, и явление внезапной наследственной изменчивости микроорганизмов считается мутационной изменчивостью. Мутанты, возникшие под влиянием обработки культуры радиацией или химическими реагентами, относятся к категории индуцированных мутантов в отличие от возникающих естественно при неучитываемом действии среды. [c.98]
В ряде работ было показано, что восстановление физиологических процессов в растениях, нарушенных под влиянием ионизирующей радиации, может быть достигнуто созданием определенных условий питания и применением физиологически активных веществ (Ильина и др., 1964, 1965 Батыгин, Мисюк, 1965 Крюкова и др., 1967). Восстановление процессов обменного характера осуществляется более легко, чем нарушения генетического типа. Некоторые исследователи отмечают, что в первую очередь важпо защитить физиологические процессы, протекающие в растениях. Это дает возможность увеличить дозу радиации, а при более высокой дозе получить и более широкий спектр мутаций у растений. Такой взгляд можно считать правильным лишь при условии конкретной задачи, связанной с радиоселекцией растений. [c.28]
Мутагенность пестицидов характеризуется частотой появления мутаций у растений, животных и у дрозофилы. По этому признаку выделяют пять групп пестицидов супермутагены — вещества, вызывающие 100% и более мутаций у растений и животных (за 100% принимается 100 мутаций на 100 хромосомах) сильные мутагены — вещества, вызывающие у дрозофилы 5—10% мутаций средние мутагены — вызывают у дрозофилы 2—5% мутаций слабые мутагены — вызывают у дрозофилы 1—2% мутаций очень слабые мутагены — вызывают у дрозофилы 0,5—1,0% мутаций. [c.83]
Мутагенность — способность пестицидов вызывать мутации у растений, животных и дрозофиллы. [c.89]
О значении цитогенетических методов в селекции свидетельствует следующий пример. На симпозиуме, посвященном npnpio-де мутаций у растений, их получению и использованию (США, Пуллман, 1969), Рэ мед ж сообщил о том, что в США министерством земледелия и Университетом Аризоны выпущен в производство первый гибридный ячмень. Для его получения использована линия с транслокацией, обеспечивающая наличие мужской стерильности у растений, служащих материнской формой. У этих растений добавлена к диплоидному набору лишняя хромосома, состоящая из участков хромосом 2—7. В этой лишней хромосоме локализован рецессивный ген мужской стерильности и ген-маркер, позволяющий отбирать растения с лишней хромосомой без цитсуюгического анализа. Лишняя хромосома передается только через яйцеклетки и должна наследоваться вместе с одной из хромосом диплоидного набора. Авторы рекомендуют пользоваться подобной сбалансированной системой для получения гибридных форм у ряда других культур. [c.130]
Эти открытия доказывали, что наследственные изменения — мутации у растений, животных и микроорганизмов — можно вызывать в эксперименте, воздействуя внешними условиями. Те№ самым устанавливались причины возиикиовения мутаций и открывались возможности для получения нужных наследственных изменений. Процесс возникновения мутаций -— мутагенез становится одной из важнейших проблем генетики. [c.189]
Школами Баура и Штуббе проведены обширные исследования мутаций у растения Antirrhinum (львиный зев). У этого айда мутация одного гена изменяет двустороннюю симметрию [c.192]
chem21.info
Мутации
Количество просмотров публикации Мутации - 422
Мутации (от лат. mutatio — перемена) — это изменение генов и хромосом, фенотипически проявляющиеся в изменении свойств и признаков организмов. Впервые их описал в 1901 ᴦ. голландский ученый Г. Де Фриз (1848-1935). Он же заложил основы и теории мутаций. Процесс образования мутаций во времени и пространстве называют мутагенезом. Мутации характерны для всех живых существ, включая человека. Определяя мутации и мутагенез, следует также определить и такие понятия, как дикий тип организма или гена и мутантный организм (мутант). Под диким типом понимают организмы со свойствами, находимыми в природе. Диким типом обозначают также наборы генов или отдельные гены, также являющиеся природными для организма. Образно говоря, дикий тип — это ʼʼсправочныйʼʼ тип, своего рода эталон, с которым сравнивают мутантные организмы и мутантные гены.
Мутантные организмы (мутанты) — это организмы, содержащие в своем геноме одну или несколько мутаций. Мутантные организмы могут отличаться от исходных (организмов дикого типа) по самым различным свойствам — морфологическим, физиологическим, биохимическим и другим. К примеру, у микроорганизмов мутации сопровождаются изменением формы колоний, питательных потребностей, отношения к лекарственным веществам и т. д. У высших растений мутации сопровождаются изменениями качественных и количественных признаков (рис. 115). У насекомых мутанты отличаются от исходных организмов по форме и окраске тела, крыльев, конечностей, глаз, реакции на свет, серологическим свойствам и т. д. (рис. 116). У млекопитающих мутации также ведут к изменению многих качественных и количественных признаков (рис. 117). У человека мутации приводят к различным отклонениям от нормы и сопровождаются наследственной патологией (см. гл. XIII). В целом можно сказать, что мутации являются вредными, полезными или нейтральными для организмов. Но важно помнить, что мутанты остаются организмом того же вида, что и организм дикого типа, из которого они произошли.
Мутации следует отличать от фенокопий, которые продуцируются факторами среды. Мимикрируя действие генов, они не передаются по наследству. К примеру, в случае если беременных мышей экспонировать к пониженному атмосферному давлению, то некоторые из особей в пометах таких мышей будут иметь повреждения в мочеполовом тракте, не передающиеся, однако, по наследству. В случае человека фенокопиями можно считать, к примеру, ненаследуемые повреждения скелета и слепоту, вызываемые ядами и другими вредными факторами воздействия.
Мутации возникают на всех стадиях индивидуального развития организмов и поражают гены и хромосомы как в половых клетках, причем до оплодотворения и после оплодотворения (после первого деления оплодотворенных яйцеклеток), так и в соматических, причем в любой фазе клеточного цикла. По этой причине по типу клеток, в которых возникают мутации, различают генеративные и соматические мутации (соответственно).
Генеративные мутации происходят в генитальных и половых клетках.
В случае если мутация (генеративная) происходит в генитальных клетках, то мутантный ген могут получить сразу несколько гамет, что увеличит потенциальную способность наследования этой мутации несколькими особями (индивидуумами) в потомстве. В случае если мутация произошла в гамете, то, вероятно, лишь одна особь (индивид) в потомстве получит данный ген. На частоту мутаций в половых клетках оказывает влияние возраст организма.
Соматические мутации встречаются в соматических клетках организмов. У животных и человека мутационные изменения будут сохраняться только в этих клетках. Но у растений из-за их способности к вегетативному размножению мутация может выйти за пределы соматических тканей. К примеру, знаменитый зимний сорт яблок ʼʼДелишесʼʼ берет начало от мутации в соматической клетке, которая в результате деления привела к образованию ветви, имевшей характеристики мутантного типа. Далее следовало вегетативное размножение, позволившее получить растения со свойствами этого сорта.
По типу наследования различают доминантные, полудоминантные, кодоминантные и рецессивные мутации. Доминантные мутации характеризуются непосредственным эффектом на организм, полудоминантные мутации заключаются в том, что гетерозиготная форма по фенотипу является промежуточной между формами АА и аа, а для кодоминантных мутаций характерно то, что у гетерозигот A1A2 проявляются признаки обоих аллелей. Рецессивные мутации не проявляются у гетерозигот.
В случае если доминантная мутация встречается в гаметах, ее эффекты выражаются непосредственно в потомстве. Многие мутации у человека являются доминантными. Οʜᴎ часты у животных и растений. К примеру, генеративная доминантная мутация дала начало анконской породе коротконогих овец.
Примером полудоминантной мутации может служить мутационное образование гетерозиготной формы Аа, промежуточной по фенотипу между организмами АА и аа. Это имеет место в случае биохимических признаков, когда вклад в признак обоих аллелей одинаков.
Примером кодоминантной мутации являются аллели IA и IB, детерминирующие группу крови IV.
В случае рецессивных мутаций их эффекты скрыты в диплоидах. Οʜᴎ проявляются лишь в гомозиготном состоянии. Примером являются рецессивные мутации, детерминирующие генные болезни человека.
Τᴀᴋᴎᴍ ᴏϬᴩᴀᴈᴏᴍ, главными факторами в детерминировании вероятности проявления мутантного аллеля в организме и популяции являются не только стадия репродуктивного цикла, но и до-минантность мутантного аллеля.
Учитывая зависимость отлокализации в клетках различают генные (точечные) и хромосомные мутации (рис. 118).
Генные мутации заключаются в изменениях индивидуальных генов. По этой причине их еще называют точечными мутациями и классифицируют на односайтовые и многосайтовые. Односайтовая мутация затрагивает один сайт, многосайтовая — несколько сайтов генного локуса. Некоторые сайты являются ʼʼгорячими точкамиʼʼ, т. к. в них происходят концентрированные мутации, что связано с наличием в нуклеотидных последовательностях модифицированных оснований. Последние подвергаются частому дезаминированию, а это ведет к изменениям в последовательностях оснований.
Генные мутации классифицируют также на прямые и обратные (реверсивные) мутации, которые одинаково встречаются у организмов всех систематических групп.
Прямые мутации — это мутации, инактивирующие гены дикого типа, т. е. мутации, которые изменяют информацию, закодированную в ДНК, прямым образом, благодаря чему изменение от организма исходного (дикого) типа идет прямым образом к организму мутантного типа.
Обратные мутации представляют из себяреверсии к исходным (диким) типам от мутантных. Эти реверсии бывают двух типов. Одни из реверсий обусловлены повторными мутациями аналогичного сайта или локуса с восстановлением исходного фенотипа и их называют истинными обратными мутациями. Другие реверсии представляют из себямутации в каком-то другом гене, которые изменяют выражение мутантного гена в сторону исходного типа, т. е. повреждение в мутантном гене сохраняется, но он как бы восстанавливает свою функцию, благодаря чему восстанавливается фенотип. Такое восстановление (полное или частичное) фенотипа вопреки сохранению первоночального генетического повреждения (мутации) получило название супрессии, а такие обратные мутации назвали супрессор-ными (внегенными). Как правило, супрессии происходят в результате мутаций генов, кодирующих синтез тРНК и рибосом.
Большинство генов довольно устойчива по отношению к мутациям, однако известны гены, которые мутируют очень часто. Как правило, мутантный фенотип организмов является одинаковым, независимо от того, наследуется ли генная мутация от отца или матери. Тем не менее небольшое число генов у человека и мышей экспрессируется дифференциально, причем исходя из источника наследования мутантного гена (от отца или матери). Во всех случаях один из родительских аллелей полностью инактиви-руется, тогда как другой аллель является активным. Напоминая инактивацию Х-хромосом, это явление получило название генетического импринтинга. Следовательно, генетический импринтинг представляет собой явление неэквивалентности генных аллелей, активность которых зависит от родительского происхождения. К примеру, патологический синдром Прадела-Вилли у человека обязан мутации, которая встречается только в отцовском хромосомном гомологе, а синдром Ангельмана у человека возникает в результате мутации в материнском хромосомном гомологе.
Хромосомные мутации связаны с изменениями числа и структуры хромосом. Изменения в числе хромосом определяются добавлением или уменьшением всего набора хромосом, ведущим к полиплоидии или гаплоидии (соответственно), часто называемые геномными мутациями, а также добавлением или удалением одной или больше хромосом из набора, что ведет к гетероплоидии или анеуплоидии (моносомии, трисомии и др. Размещено на реф.рфполисемии), тогда как изменения в структуре хромосом определяются перестройками (аберрациями) их структуры.
Полиплоидия — это хромосомная мутация в виде увеличения числа полных гаплоидых наборов хромосом. Известны триплоидия (Зп), тетраплоидия (4п), пентаплоидия (5п) и т. д. Наиболее часто полиплоидия встречается у растений, поскольку для них характерны гермафродитизм и апомиксис. Почти третья часть всех видов диких цветковых растений представлена полиплоидами. Типичными полиплоидами являются виды пшеницы, у которых соматические числа 2n = 14,28 и 42, при основном гаметном числе n = 7, картофель, табак, белый клевер, люцерна и другие растения. Родственные виды, наборы хромосом которых представляют ряд возрастающего увеличения (кратного) основного числа хромосом, составляют полиплоидные ряды.
Полиплоидные растения с нечетными наборами хромосом характеризуются пониженной плодовитостью и снижением уровня количественных признаков. Напротив, для растений-полиплоидов с четными наборами хромосом характерен повышенный уровень количественных признаков.
Полиплоиды, происходящие от диплоидных организмов аналогичного вида, называют автополиплоидами. Примером автопо-липлоидов служат растения мягкой пшеницы, у которых 2п == 42 и которые являются гексаплоидами. Искусственные полиплоиды, полученные из гибридов диплоидных растений, относящихся к далеко отстоящим друг от друга видам, называют аллополипло-идами.
В природе встречаются как автополиплоиды, так и аллополип-лоиды, однако географическое распределение полиплоидов обычно отличается от распределения их диплоидиых ʼʼродственниковʼʼ. К примеру, флора о. Шпицберген содержит очень высокий процент полиплоидных рядов, тогда как в других местах количество их меньше по сравнению с диплоидами.
Полиплоидия растений имеет хозяйственную ценность (повышенные размеры плодов, большая сахаристость, лучшая сохранность и др.). По этой причине полиплоиды используют в селекционной работе для получения новых сортов сельскохозяйственных растений.
У животных полиплоидия очень редка. Она встречается у земляных червей, размножающихся партеногенетически (обнаружены полиплоидные ряды с основными числами 11, 16, 17, 18 и 19 хромосом), у некоторых членистоногих, рыб и земноводных. В частности, она описана у морских креветок Artemia salivana. Женские особи саламандр отдельных видов, которые имеют крупные эритроциты с ядрами, продуцируют триплоидные личинки с 42 хромосомами, тогда как саламандры с малыми ядерными эритроцитами продуцируют диплоидные личинки с 28 хромосомами. Все тихоокеанские лососевые рыбы являются полиплоидами.
Сирийский хомячок (Mesocricetius aurantus), у которого 2 n = 44, является аллополиплоидом, возникшим в результате естественной гибридизации между европейским хомяком (Cricetus cricetus, 2n == 22) и хомяком, принадлежащим к одному из азиатских видов (Cricetus griseus, 2n = 24).
Полиплоидия встречается также у человека в пренатальном периоде развития. В частности известно много сообщений об обнаружении триплоидии и тетраплоидии в клетках абортусов. Имеющиеся данные позволяют считать, что триплоидия встречается у 20% абортусов, а тетраплоидия — у 6% абортусов.
Редкость полиплоидии у раздельнополых животных, по-видимому, определяется тем, что она нарушает нормальные соотношения аутосом и половых хромосом в зиготах.
Кроме авто- и аллополиплоидии, в соматических клетках ряда многоклеточных организмов установлена эндополиплоидия, характеризующаяся увеличением числа хромосом в покоящемся ядре (при отсутствии митоза). От полиплоидии следует отличать псевдополиплоидию отдельных растений и насекомых, возникающую в результате однократного или многократного деления компонентов генома, когда центромеры имеют диффузный характер.
Гаплоидия — это мутация в виде уменьшения всего набора хромосом. Она найдена в основном также у растений. Известны гапло-иды свыше 800 видов растений, включая такие культуры, как пшеница и кукуруза. У животных она очень редка, а у человека совсем неизвестна.
Мутации в виде нарушений нормального количества хромосом из-за добавления или удаления одной или более хромосом в какой-то хромосомной паре называют гетероплоидией или анеуп-лоидией. Среди гетероплоидов известны полисемии, когда какая-то пара хромосом становится большей на один (трипликатом) и более гомологов, моносомии, когда какая-либо хромосомная пара теряет один гомолог, и нулесомии, когда теряется вся хромосомная пара. Эти мутации широко распространены как у животных, так и у растений. В частности трисомии и моносомии обнаружены у человека, собак и других животных, а также у многих плодовых, зерновых и овощных растений. Часто трисомии, как и моносомии, обнаруживают в клетках абортированных эмбрионов или плодов человека. Гетероплоидия сопровождается многими фенотипическими эффектами, которые, к примеру, у человека неблагоприятны для его здоровья.
Мутации, поражающие структуру хромосом, называют хромосомными перестройками или чаще аберрациями. Эти мутации классифицируют на внутрихромосомные и межхромосомные перестройки. Οʜᴎ возникают как у животных, включая человека, так и у растений.
Внутрихромосомными перестройками являются делении (нехватки, дефишенсы), дубликации, инверсии, тогда как межхромосомные перестройки представлены транслокациями (рис. 119). Внутрихромосомные перестройки исходя из ʼʼразломаʼʼ плеч хромосом бывают парацентрическими (ʼʼразломʼʼ затрагивает одно хромосомное плечо) и перицентрическими (ʼʼразломʼʼ проходит по обе стороны центромеры, т. е. захватывает оба плеча). Межхромосомные перестройки обычно захватывают две или более негомологичные хромосомы. Степень изменений бывает различной и часто доходит до изменения группы сцепления.
Делеции представляют из себяпотери сегмента хромосомы, несущего один или несколько генов. Οʜᴎ являются наиболее частой и опасной для человека формой генетических макроповреждений. Большие делеции заключаются в потере одного или нескольких генов или даже блоков генов. У гаплоидных организмов крупные делеции детальны. Эффект делеции у диплоидных организмов зависит от количества делегированных генов, количественных потребностей в продуктах пораженных генов, позиции генов среди функционально координированных групп и других факторов. Гомозиготные делеции для диплоидных клеток или организмов детальны.
Дупликации (добавления) представляют из себядобавления (удлинения) какого-либо сегмента хромосомы, несущего один или несколько генов, в результате того, что один и тот же сегмент хромосомы должна быть повторен в процессе биосинтеза несколько раз. Этот повтор должна быть малым, касаясь одиночного гена, или большим, затрагивая большое количество генов. Дупликации часто безвредны для их носителей. Предполагают, что они способствуют формированию полигенов или являются способом введения новых генов в геномы. Некоторые дуп-ликации, однако, вредны и даже детальны (см. гл. IX).
Инверсии заключаются в поворотах на 180° сегментов, освобождающихся в результате парных разрывов в хромосомах. В случае если инвертированный сегмент не содержит центромеру, эту мутацию называют парацентрической инверсией, в случае если же такой сегмент содержит центромеру, такую мутацию называют перицентрической. Инверсии оказывают влияние на мейоз, что приводит к пониженной фертильности гибридов. Описаны отдельные наследственные аномалии, вызываемые этой мутацией (см. гл. XIII).
Транслокация — это обмен частями (сегментами) гомологич-ной и негомологичной хромосом, образованными разрывами по длине последних. Транслоцируемые сегменты могут иметь разные размеры — от небольших до значительных.
Хромосомные перестройки, как и гетероплоидия, также сопровождаются различными фенотипическими эффектами. У человека они ведут к нарушению здоровья. У растений они ведут к изменению их продуктивности.
При этом, обсуждая значение хромосомных перестроек, нельзя не заметить, что для отдельных организмов они безразличны. К примеру, у пионов, дурмана и других растений присутствие в гено-ме транслокаций является нормальным явлением.
Учитывая зависимость отпроисхождения различают спонтанные и индуцированные генные и хромосомные мутации, которые возникают у организмов независимо от уровня их организации.
Спонтанными называют те мутации, которые возникают у организмов в нормальных (природных) условиях на первый взгляд без видимых причин, тогда как индуцированными называют те мутации, которые возникают в результате обработки клеток (организмов) мутагенными факторами. Главное отличие спонтанных мутаций от индуцированных состоит по сути в том, что мутация может возникнуть в любой период индивидуального развития. Что касается случайного характера мутаций в пространстве, то это означает, что спонтанная мутация произвольно может поразить любую хромосому или ген.
Длительное время считали, что спонтанные мутации являются беспричинными, однако теперь по этому вопросу существуют другие представления, сводящиеся к тому, что спонтанные мутации не являются беспричинными, что они являются результатом естественных процессов, протекающих в клетках. Οʜᴎ возникают в условиях природного радиоактивного фона Земли в виде космического излучения, радиоактивных элементов на поверхности Земли, радионуклидов, инкорпорированных в клетки организмов, которые вызывают эти мутации или в результате ошибок репликации ДНК. Факторы естественного радиоактивного фона Земли вызывают изменения в последовательности оснований или повреждения оснований подобно тому, как это имеет место в случае индуцированных мутаций (см. ниже).
Ошибки репликации ДНК заключаются в том, что какой-либо нуклеотид ошибочно включается в ДНК при ее репликации и при наличии недостатка в ʼʼредактирующемʼʼ механизме, что исключает коррекцию ошибок. Ошибки в репликации ДНК, т. е. ʼʼнеаккуратностьʼʼ ДНК-полимеразы оказывают влияние на частоту спонтанных мутаций. Ошибки репликации ДНК бывают также связаны с химической нестабильностью нуклеотидов, сопровождающейся изменением спаривающей способности азотистых оснований. К примеру, цитозин должна быть дезаминирован в урацил, который затем распознается ДНК-полимеразой как тимин в течение репликации ДНК.
Определение частот спонтанных мутаций организмов разных видов проводят с помощью разных способов, один из которых связан с определением частоты таких мутаций на репликацию пары азотистых оснований в молекулах ДНК. Этот способ оказался предпочтительным в случае бактерий и других низших организмов.
Определенные к настоящему времени частоты мутаций на репликацию пары оснований и общие частоты генных мутаций у разных организмов показаны в табл. 13.
В случае человека частоты спонтанных мутаций определяют измерением прямых мутаций в пределах разных генов, которые очень чувствительны к мутациям независимо от того, являются ли условия для организмов ограничивающими или селективными.
Рассмотрим конкретный пример определения частоты спонтанных мутаций, к примеру, среди родившихся за один год 242 257 детей 7 оказались больными ахондоплазией. Следовательно, 7 : 242 257´1 : 2 (два аллеля на зиготу) = 1,4´10-6. Таким образом частота ахондоплазии составляет 1,4 х 10-5. Одни гены вообще устойчивы к спонтанным мутациям, другие спонтанно мутируют чаще, третьи так часто, что их носители являются мозаиками мутантных (мутировавших) и немутантных (немутировавших) генов.
Средние частоты мутаций по многим генам у человека и домашних животных составляют примерно 1´10-9, что значительно выше частоты мутаций микроорганизмов. Больше того, между частотами спонтанных мутаций по разным генам человека или домашних млекопитающих существуют значительные различия, достигающие 100 раз, а то и более. Подлинные причины этих различий неизвестны, хотя для их объяснения и предложено несколько гипотез. Одна из них состоит по сути в том, что наиболее чувствительны к мутациям гены больших размеров, поскольку в них содержится много азотистых оснований и существует большая вероятность мутации отдельных из них. По другой гипотезе наиболее чувствительными к мутациям являются гены, располагающиеся в районах хромосом, являющихся ʼʼгорячимиʼʼ точками.
Таблица 13
referatwork.ru
материалы для подготовки к ЕГЭ по Биологии
Автор статьи - Л.В. Окольнова.
Сразу на ум приходят Люди Х… или Человек - Паук …
Но это в кино, в биологии тоже так, но немного более научно, менее фантастично и более обыденно.
Мута́ция (в переводе — изменение) — устойчивое, передающееся по наследству изменение ДНК, происходящее под влиянием внешних или внутренних изменений.
Мутагенез - процесс появления мутаций.
Обыденность в том, что эти изменения (мутации) происходят в природе и у человека постоянно, почти каждодневно.
В первую очередь, мутации подразделяются на соматические - возникают в клетках тела, и генеративные - появляются только в гаметах.
| Соматические мутации | Генеративные мутации |
| Не всегда передаются при половом размножении. Передаются при вегетативном (бесполом размножении). | Передаются по наследству. |

Разберем сначала виды генеративных мутаций.
Генные мутации
Что такое ген? Это участок ДНК (т.е. несколько нуклеотидов), соответственно, это и участок РНК, и участок белка, и какой-либо признак организма.
Т.е. генная мутация - это выпадение, замена, вставка, удвоение, изменение последовательности участков ДНК.
Вообще, это не всегда ведет к болезни. Например, при удвоении ДНК случаются такие “ошибки”. Но они возникают редко, это очень малый процент от всего количества, поэтому они незначительны, что практически не влияют на организм.
Бывают и серьезные мутагенезы:- серповидно-клеточная анемия у человека;- фенилкетонурия - нарушение обмена веществ, вызывающее довольно серьезные нарушения умственного развития- гемофилия- гигантизм у растений
Геномные мутации
Вот классическое определение термина “геном”:
Геном-
- совокупность наследственного материала, заключенного в клетке организма;- геном человека и геномы всех остальных клеточных форм жизни, построены из ДНК;- совокупность генетического материала гаплоидного набора хромосом данного вида в парах нуклеотидов ДНК на гаплоидный геном.
Для понимания сути мы очень сильно упростим, получится такое определение:
Геном - это количество хромосом
Геномные мутации - изменение числа хромосом организма. В основном, их причина - нестандартное расхождение хромосом в процессе деления.
- синдром Дауна - в норме у человека 46 хромосом (23 пары), однако при этой мутации образуются 47 хромосомрис. синдром Дауна
- полиплойдия у растений (для растений это вообще норма - большинство культурный растений - полиплойдные мутанты)
Хромосомные мутации - деформации самих хромосом.
Примеры (некоторые перестройки такого рода есть у большинства людей и вообще никак не отражаются ни внешне, ни на здоровье, но есть и неприятные мутации):- синдром кошачьего крика у ребенка- задержка в развитиии т.д.
Цитоплазматические мутации - мутации в ДНК митохондрий и хлоропластов.
Есть 2 органеллы со своими собственными ДНК (кольцевыми, в то время как в ядре - двойная спираль) - митохондрия и растительные пластиды.
Соответственно, есть мутации, вызванные изменениями именно в этих структурах.
Есть интересная особенность - этот вид мутации передается только женским полом, т.к. при образовании зиготы остаются только материнские митохондрии, а “мужские” отваливаются с хвостом при оплодотворении.
Примеры:- у человека - определенная форма сахарного диабета, туннельное зрение;- у растений - пестролистность.
Соматические мутации.
Это все описанные выше виды, но возникают они в клетках тела ( в соматических клетках).Мутантных клеток обычно намного меньше, чем нормальных, и они подавляются здоровыми клетками. (Если не подавляются, то организм перерождаться или болеть).
Примеры:- у дрозофилы глаз красный, но может иметь белые фасеты- у растения это может быть целый побег, отличающийся от других (И.В. Мичурин таким образом выводил новые сорта яблок).
- раковые клетки у человека
Примеры вопросов ЕГЭ:
Синдром Дауна является результатом мутации
1))геномной;
2) цитоплазматической;
3)хромосомной;
4) рецессивной.
Ответ: 1.
Генные мутации связаны с изменением
А) числа хромосом в клетках;
Б) структуры хромосом;
B) последовательности генов в аутосоме;
Г) нуклеогидов на участке ДНК.
Ответ: Г.
Мутации, связанные с обменом участками негомологичных хромосом, относят к
А) хромосомным;
Б) геномным;
В) точковым;
Г) генным.
Ответ: А.
Животное, в потомстве которого может появиться признак, обусловленный соматической мутацией
А) гидра
Б) волк
В) еж
Г) выдра
Ответ: А.
Звоните нам: 8 (800) 775-06-82 (бесплатный звонок по России) +7 (495) 984-09-27 (бесплатный звонок по Москве)
Или нажмите на кнопку «Узнать больше», чтобы заполнить контактную форму. Мы обязательно Вам перезвоним.
ege-study.ru
Классификация мутаций по фенотипу:Классификации мутаций: по Мёллеру
 . .

По генотипу:
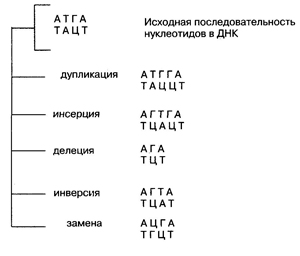
 Замены оснований приводят к появлению трех типов мутантных кодонов: с измененным смыслом (миссенс-мутации), с неизмененным смыслом (нейтральные мутации) и бессмысленных, или терминирующих кодонов (нонсенс-мутации). — мутации, изменяющие последовательность нуклеотидов в гене, т. е. структуру самого гена.
2. Генные инсерции — вставка пары или нескольких нар нуклеотидов (вставка пары Г—Ц между А—Т и Т—А). 3. Генные делеции — выпадение нуклеотидов (выпадение комплементарной пары Т—А между А—Т и Г—Ц). 4. Генные инверсии — перестановка фрагмента гена (во фрагменте исходная последовательность нуклеотидов Т—А, Г—Ц заменяется на обратную Г—Ц, Т—А). 5. Замены нуклеотидов — замена пары нуклеотидов на другую; при этом общее число нуклеотидов не меняется (замена Т—А на Ц—Г). Один из наиболее частых типов мутаций. Дупликации, инсерции и делеции могут приводить к изменению рамки считывания генетического кода. Рассмотрим это на примере. Возьмем следующую исходную последовательность нуклеотидов в ДНК (для простоты будем рассматривать только одну ее цепь): АТГАЦЦГЦГА... Она будет считываться следующими триплетами: АТГ, АЦЦ, ГЦГ, А... Допустим, произошла делеция, и в самом начале последовательности между А и Г выпал нуклеотид Т. В результате этой мутации получится измененная последовательность нуклеотидов: АГАЦЦГЦГА, которая уже будет считываться совершенно иными триплетами: АГА, ЦЦГ, ЦГА. Поэтому в полипептидную цепь будут соединяться совершенно другие аминокислоты и, таким образом, будет синтезироваться мутантный белок, совершенно непохожий на нормальный. Кроме того, в результате генных мутаций, приводящих к сдвигу рамки, могут образовываться терминирующие кодоны ТАА, ТАГ или ТГА, прекращающие синтез. Выпадение целого триплета приводит к менее тяжелым генетическим последствиям, чем выпадение одного или двух нуклеотидов. Рассмотрим ту же нуклеотидную последовательность: АТГАЦЦГЦГА... Допустим, произошла делеция, и выпал целый триплет АЦЦ. Мутантный ген будет иметь измененную последовательность нуклеотидов АТГГЦГА, которая будет считываться следующими триплетами: АТГ, ГЦГ, А... Видно, что после выпадения триплета рамка считывания не сдвинулась, синтезированный белок хоть и будет на одну аминокислоту отличаться от нормального, но в целом будет весьма на него похож. Однако это отличие в аминокислотном составе может привести к изменению третичной структуры белка, которая в основном и определяет его функции, и функция мутантного белка, скорее всего, будет снижена по сравнению с нормальным белком. Этим и объясняется тот факт, что мутации, как правило, рецессивны. Генные мутации проявляются фенотипически в результате синтеза соответствующих белков:
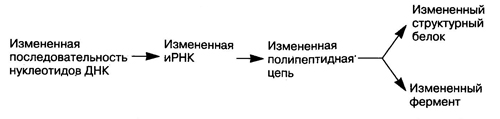
Генные мутации приводят к изменению строения молекул белков и к появлению новых признаков и свойств (например, альбиносы у животных и растений, махровость у цветков за счет преобразования тычинок в лепестки и снижение их плодовитости, образование летальных и полулетальных генов, вызывающих гибель организма, и т. д.). Генные мутации происходят пoд влиянием мутагенных факторов (биологических, физических химических) или спонтанно (случайно). Генные мутации свойственны и генетической РНК вирусов.
Геномные мутации характеризуются изменением числа хромосом. У человека известны полиплоидия (в том числе тетраплоидия и триплоидия) и анеуплоидия. Полиплоидия — увеличение числа наборов хромосом, кратное гаплоидному (Зn, 4n, 5n и т.д.). Причины: двойное оплодотворение и отсутствие первого мейотического деления. У человека полиплоидия, а также большинство анеуплоидий приводят к формированию леталей. Исключительно велика роль полиплоидии в происхождении культурных растений и их селекции. Полиплоидными являются все или большинство культивируемых сортов пшеницы, овса, риса, сахарного тростника, арахиса, свеклы, картофеля, сливы, яблони, груши, апельсина, лимона, земляники, малины. К этому перечню следует добавить тимофеевку, люцерну, табак, хлопчатник, розы, тюльпаны, хризантемы, гладиолусы и многие другие, возделываемые человеком, культуры. Аутополиплоидные мутанты растений обычно крупнее исходной формы. Тетраплоиды, как правило, имеют большую вегетативную массу. Однако у них может резко уменьшиться плодовитость из-за нерасхождения поливалентов в мейозе. Триплоиды - крупные и мощные растения, но полностью или почти полностью стерильные, поскольку продуцируемые ими гаметы содержат неполный набор хромосом. Аутополинлоидные виды размножают вегетативным способом, поскольку плоды таких растений не содержат семян. Анеуплоидия — изменение (уменьшение — моносомия, увеличение — трисомия) числа хромосом в диплоидном наборе, т.е. не кратное гаплоидному (2n+1, 2n-1 и т.д.). Механизмы возникновения: нерасхождение хромосом (хромосомы в анафазе отходят к одному полюсу, при этом на каждую гамету с одной лишней хромосомой приходится другая — без одной хромосомы) и «анафазное отставание» (в анафазе одна из передвигаемых хромосом отстаёт от всех других). *Трисомия — наличие трёх гомологичных хромосом в кариотипе (например, по 21-й паре, что приводит к развитию синдрома Дауна; по 18-й паре — синдрома Эдвардса; по 13-й паре — синдрома Патау). *Моносомия — наличие только одной из двух гомологичных хромосом. При моносомии по любой из аутосом нормальное развитие эмбриона невозможно. Единственная совместимая с жизнью моносомия у человека — по хромосоме X — приводит к развитию синдрома Шерешевского—Тернера (45,Х0). *Тетрасомия и пентасомия:Тетрасомия (4 гомологичные хромосомы вместо пары в диплоидном наборе) и пентасомия (5 вместо 2-х) встречаются чрезвычайно редко. Примерами тетрасомии и пентасомии у человека могут служить кариотипы XXXX, XXYY, XXXY, XYYY, XXXXX, XXXXY, XXXYY, XYYYY и XXYYY. Как правило, с нарастанием количества "лишних" хромосом увеличивается тяжесть и выраженность клинических симптомов.Гаплоидия,- противоположное полиплоидии явление, заключающееся в кратном уменьшении числа хромосом у потомства в сравнении с материнской особью. Гаплоидия, как правило, - результат развития зародыша из редуцированных (гаплоидных) гамет или из функционально равноценных им клеток путём апомиксиса, т. е. без оплодотворения.Гаплоидия редко встречается в животном мире, но распространена у цветковых растений: зарегистрирована более чем у 150 видов растений из 70 родов 33 семейств (в т. ч. из семейства злаков, паслёновых, орхидных, бобовых и др.). Известна у всех основных культурных растений: пшениц, ржи, кукурузы, риса, ячменя, сорго, картофеля, табака, хлопка, льна, свёклы, капусты, тыквы, огурцов, томатов; у кормовых трав: мятликов, костра, тимофеевки, люцерны, вики и др. Гаплоидия генетически детерминирована и встречается у некоторых видов и сортов с определённой частотой (например, у кукурузы - 1 гаплоид на 1000 диплоидных растений). В эволюции видов Гаплоидия служит своеобразным механизмом, снижающим уровень плоидности. Гаплоидия пользуются для решения ряда генетических проблем: выявления эффекта дозы гена, получения анеуплоидов, для исследования генетики количественных признаков, генемного анализа и др. В селекции растений Гаплоидия пользуются для получения из гаплоидов путём удвоения у них числа хромосом гомозиготных линий, равноценных самоопылённым линиям при производстве гибридных семян (например, у кукурузы), а также для перевода селекционного процесса с полиплоидного на диплоидный уровень (например, у картофеля). Особая форма Гаплоидия - андрогенез, при котором ядро спермия замещает ядро яйцеклетки, используется для получения мужских стерильных аналогов у кукурузы.
Хромосомные мутации. Виды хромосомных мутаций.Хромосомные мутации (аберрации) характеризуются изменением структуры отдельных хромосом. При них последовательность нуклеотидов в генах обычно не меняется, но изменение числа или положения генов при аберрациях может привести к генетическому дисбалансу, что пагубно сказывается на нормальном развитии организма. Виды аберраций и их механизмы представлены на рисунке.  Различают внутрихромосомные, межхромосомные и изохромосомные аберрации. Хромосомные аберрации (хромосомные мутации, хромосомные перестройки) — тип мутаций, которые изменяют структуру хромосом. Классифицируют делеции (утрата участка хромосомы), инверсии (изменение порядка генов участка хромосомы на обратный), дупликации (повторение участка хромосомы), транслокации (перенос участка хромосомы на другую), а также дицентрические и кольцевые хромосомы. Известны также изохромосомы, несущие два одинаковых плеча. Если перестройка изменяет структуру одной хромосомы, то такую перестройку называют внутрихромосомной (инверсии, делеции, дупликации, кольцевые хромосомы), если же двух разных, то межхромосомной (дупликации, транслокации, дицентрические хромосомы). Хромосомные перестройки подразделяют также на сбалансированные и несбалансированные. Сбалансированные перестройки (инверсии, реципрокные транслокации) не приводят к потере или добавлению генетического материала при формировании, поэтому их носители, как правило, фенотипически нормальны. Несбалансированные перестройки (делеции и дупликации) меняют дозовое соотношение генов, и, как правило, их носительство сопряжено с клиническими отклонениями от нормы. Внутрихромосомные аберрации — аберрации в пределах одной хромосомы. К ним относятся делеции, инверсии и дупликации. *Делеция — утрата одного из участков хромосомы (внутреннего или терминального), что может стать причиной нарушения эмбриогенеза и формирования множественных аномалий развития (например, делеция в регионе короткого плеча хромосомы 5, обозначаемая как 5р-, приводит к недоразвитию гортани, ВПР сердца, отставанию умственного развития). Этот симптомокомплекс обозначен как синдром кошачьего крика, поскольку у больных детей из-за аномалии гортани плач напоминает кошачье мяуканье. *Инверсия — встраивание фрагмента хромосомы на прежнее место после поворота на 180°. В результате нарушается порядок расположения генов. *Дупликация — удвоение (или умножение) какого-либо участка хромосомы (например, трисомия по короткому плечу хромосомы 9 приводит к появлению множественных ВПР, включая микроцефалию, задержку физического, психического и интеллектуального развития). Межхромосомные аберрации — обмен фрагментами между негомологичными хромосомами. Они получили название транслокаций. Различают три варианта транслокаций: реципрокные (обмен фрагментами двух хромосом), нереципрокные (перенос фрагмента одной хромосомы на другую), робертсоновские (соединение двух акроцентрических хромосом в районе их центромер с потерей коротких плеч, в результате образуется одна метацентри-ческая хромосома вместо двух акроцентрических). *Реципрокные скрещивания — два эксперимента по скрещиванию, характеризующиеся прямо противоположным сочетанием пола и исследуемого признака. В одном эксперименте самца, имеющего определенный доминантный признак, скрещивают с самкой, имеющий рецессивный признак. Во втором, соответственно, скрещивают самку с доминантным признаком и самца с рецессивным признаком.Реципрокные транслокации являются сбалансированной хромосомной перестройкой, при их формировании не происходит потери генетического материала. Они являются одной из самых распространенных хромосомных аномалий в человеческой популяции, частота носительства варьирует от 1/1300 до 1/700[1][2]. Носители реципрокных транслокаций, как правило, фенотипически нормальны, при этом имеют повышенную вероятность бесплодия, сниженной фертильности, спонтанных выкидышей и рождения детей с врождёнными наследственными заболеваниями, так как половина гамет у них генетически несбалансирована из-за неравновесного расхождения перестроенных хромосом в мейозе. Изохромосомные аберрации — образование одинаковых, но зеркальных фрагментов двух разных хромосом, содержащих одни и те же наборы генов. Это происходит в результате поперечного разрыва хроматид через центромеры (отсюда другое название — центрическое соединение).
(аберрации, перестройки) — изменения положения участков хромосом; приводят к изменению размеров и формы хромосом. В эти изменения могут быть вовлечены как участки одной хромосомы, так и участки разных, негомологичных хромосом, поэтому хромосомные мутации (перестройки) подразделяются на внутри- и межхромосомные. А. Внутрихромосомные мутации 1. Хромосомные дупликации — удвоение участка хромосомы. 2. Хромосомные делеции — утрата хромосомой какого-либо участка. Хромосомные инверсии — разрыв хромосомы, переворачивание оторвавшегося участка на 180° и встраивание его на прежнее место. Б. Межхромосомные мутации 1.Транслокация — обмен участками между негомологичными хромосомами (в мейозе).тип хромосомных мутаций, при которых происходит перенос участка хромосомы на негомологичную хромосому. Отдельно выделяют реципрокные транслокации, при которых происходит взаимный обмен участками между негомологичными хромосомами, и Робертсоновские транслокации, или центрические слияния, при которых происходит слияние акроцентрических хромосом с полной или частичной утратой материала коротких плеч.Транслокации, также как и другие хромосомные перестройки, играют роль в видообразовании, в снижении фертильности, в онкологических и врождённых наследственных заболеваниях. Различные транслокации в соматических клетках приводят к развитию лимфом, сарком, лейкозов. 2.Транспозиция — включение участка хромосомы в другую, негомологичную хромосому без взаимного обмена. Оценка за работу: 5 |



Генеративные и соматические мутации
Мутации возникают в клетках любых тканей многоклеточного организма и на различных стадиях его развития.
Мутации, возникающие в клетках полового зачатка и в половых клетках, называют генеративными. Мутации, возникающие в клетках других тканей тела, называют соматическими. Необходимость такого разделения вызвана тем, что эволюционная ценность генеративных и соматических мутаций различна и определяется типом размножения организма.
Мутации генов в половых клетках обнаруживаются на стадии зиготы следующих поколений. Если исключить такие мутации, которые действуют на гаметы, затрагивая их физиологию и оплодотворяющую способность, то доминантная мутация проявляется в зиготе первого же поколения F1 а рецессивная — лишь в F2, F3 и т. д. при переходе мутации в гомозиготное состояние.
Если генеративная мутация возникает в одной клетке на ранней стадии зачаткового пути или в период размножения сперматогониев и оогониев, то такая мутация размножится в количестве, пропорциональном числу, прошедших клеточных делений. В этом случае часть половых клеток будет нести мутантную аллель, а у части генотип останется неизменным. Особь с такими половыми клетками будет нести «пучок» идентичных мутаций, которые обнаруживаются генетически при скрещивании.
Мутация, возникшая на стадии сперматозоида или яйцеклетки, останется, как правило, единичной.
Соматические мутации по своей природе ничем не отличаются от генеративных. В отношении растений, которые вообще не имеют зачаткового пути и у которых половые клетки развиваются из меристемы точки роста, деление мутаций по указанному принципу не имеет большого значения. Различие состоит лишь в проявлении и методах их обнаружения. Чем раньше в онтогенезе возникает соматическая мутация, тем больше оказывается участок ткани, несущие данную мутацию, и чем позднее — тем меньше. Соматическая мутация проявляется мозаично. Особи, несущие участки мутантной ткани, называют мозаиками, или химерами. В силу диплоидности набора хромосом в клетках соматической ткани, проявление мутации возможно только в тех случаях, когда мутантная аллель оказывается доминантной или будет рецессивна и будет находиться в гомозиготном состоянии.
Показан пример соматической мутации окраски шерстного покрова у овцы: черное пятно возникло на фоне коричневой окраски. Эта мутация могла проявиться либо как доминантная, либо как рецессивная при потере части или всей гомологичной хромосомы.

Соматическая мутация (темное пятно) окраски шерстного покрова у каракульской овцы
Подобные явления часто встречаются у самцов дрозофилы, у которых иногда часть глаза имеет красные фасетки, а часть — белые. Эта мозаичность обязана возникновению рецессивной мутации в локусе white половой хромосомы во время развития имагинальных дисков глаз. Но появляется она не только у самцов, у которых этот ген в Х-хромосоме находится в гомизиготном состоянии, но и у гетерозиготных самок в силу утраты целой хромосомы, несущей доминантную аллель w+, или потери части хромосомы (дефишенси), несущей ту же аллель. В этом случае рецессивная аллель, теперь уже находясь в гомозиготном состоянии, также может проявиться, но эта химерность будет не следствием мутации гена, а следствием изменений в числе или структуре хромосом.
У организмов, размножающихся исключительно половым путем и имеющих раннее обособление зачаткового пути, соматические мутации не играют роли в эволюции и не представляют какой-либо ценности для селекции. Но у тех организмов, у которых есть бесполое размножение, соматические мутации могут иметь огромное значение, особенно в селекции, так как у таких форм из соматической ткани развиваются половые клетки. Так, например у плодовых и ягодных вегетативно размножаемых растений любая соматическая мутация может дать растение и целый клон с новым мутантным признаком.
Одним из видов соматических мутаций у растений являются почковые мутации, возникающие в меристемных клетках точки роста стебля. В этом случае весь побег, развившийся из этой клетки, будет нести мутантный признак. Почковые мутации были известны давно и назывались спортами; от такого спорта И. В. Мичурин получил сорт яблони, названный им Антоновка 600-граммовая.
Исследование соматических мутации в настоящее время приобретает важное значение для изучения причин возникновения рака у человека и животных. Предполагают, что ряд злокачественных опухолей возникает по типу соматических мутаций. Соматические мутации имеют прямое отношение также к выяснению причин старения человеческого организма, так как с возрастом может происходить накопление физиологических мутаций в популяции соматических клеток различных органов.
Соматические мутации используют для изучения частоты возникновения видимых мутаций. По мозаичности проявления мутантного признака в тканях можно обнаруживать соматические мутации вплоть до одиночных мутантных клеток.
Различий в частоте возникновения соматических и генеративных мутаций не обнаружено. Однако есть факты, указывающие на то, что ряд генов мутирует с разной скоростью на разных стадиях онтогенеза. Так, например, у растения дельфиниум ген левандовой окраски мутирует как на очень ранней, так и на поздней стадии развития цветка; в случае мутирования гена на поздней стадии в лепестках встречаются одиночные клетки с измененной окраской; при возникновении той же мутации на ранней стадии она может затрагивать большие по размеру участки — половину лепестка или даже целый цветок.
Таким образом, генеративные и соматические мутации по своему возникновению не отличаются друг от друга.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info










