
Что такое митохондрии... Митохондрии: описание, строение и функции. Митохондрии растений
Митохондрии (хондриосомы) растительной клетки
Митохондрии — клеточные органоиды, которые находятся во всех живых клетках и рассеяны в цитоплазме.
Форма их при рассматривании в световом микроскопе очень разнообразна и изменчива, они имеют вид зернышек, цилиндриков, палочек или нитей. В электронном микроскопе они обычно имеют округленную или цилиндрическую форму.
Длина митохондрий — несколько микронов, толщина в среднем 0,6—1,0 мк. В световом микроскопе структура их не различается. Их можно прижизненно окрасить в клетке с помощью особой краски (януса зеленого). В зависимости от типа и состояния клетки количество митохондрий варьирует от нескольких десятков до нескольких тысяч, в среднем же их в клетке несколько сотен. Митохондрии отличаются от цитоплазмы несколько большей плотностью и вязкостью и химическим составом. Это очень чувствительные образования, легко разрушающиеся или изменяющие свою форму под действием неблагоприятных условий.

Клетка древесинной паренхимы белой акации
Значение митохондрий в жизнедеятельности клетки и их строение до последнего времени оставались неясными. С помощью электронного микроскопа была выявлена их структура. Снаружи они отграничены оболочкой, состоящей из двух параллельных мембран и светлого промежутка между ними, заполненного водянистым содержимым. Наружная мембрана обычно сплошная, а внутренняя дает выступы в виде складок или трубочек, увеличивающих внутреннюю активную поверхность митохондрии. Эти складки и трубочки называются гребнями, или кристами. Кристы у митохондрий растительных клеток определенно не ориентированы и располагаются в беспорядке. Все пространство между кристами заполнено относительно плотным по сравнению с цитоплазмой материалом, по-видимому, белкового происхождения, который называется матриксом, или основным веществом митохондрии. Матрикс обычно бесструктурен, но часто содержит электронноплотные зернышки, напоминающие рибосомы, и волокна (микрофибриллы). Возможно, что последние представляют собой структурный белок и содержат ДНК.
Строение митохондрии изменяется с возрастом клетки. В молодых, эмбриональных клетках тело митохондрии недифференцировано, внутренняя структура выражена слабо, основное вещество почти гомогенно, зернистость отсутствует, кристы немногочисленные и короткие. Во взрослой клетке кристы обычно становятся более многочисленными, их длина увеличивается, основное вещество приобретает зернистую структуру. Обилие крист наблюдается у клеток с высокоактивной жизнедеятельностью. У стареющей клетки кристы уменьшаются, превращаясь в неправильные трубочки, а зернистость матрикса исчезает. При повреждающем действии на растение низких и высоких температур или при его голодании наблюдается сильное набухание митохондрий (увеличение их размеров), разжижение матрикса, появление внутри митохондрий многочисленных пузырьков и, наконец, потеря внутренней мембранной структуры.

Строение митохондрий
Сложная внутренняя структура митохондрий является отражением их важной физиологической роли в обмене веществ клетки, которая была открыта совсем недавно. Оказалось, что каждая митохондрия — это своеобразная внутриклеточная лаборатория, где происходит дыхание, при котором потенциальная
энергия, заключенная в питательных веществах, освобождается и направляется на поддержание жизнедеятельности клетки. Поэтому клетки с высокой активностью освобождают большее количество энергии и содержат большое число митохондрий. Освобождение и перенос энергии осуществляется с помощью огромного числа ферментов. Предполагают, что эти ферменты сосредоточены в основном на мембранах митохондрий. Комплексы дыхательных ферментов входят в состав так называемых элементарных частиц митохондрий, которые расположены на наружной поверхности внешней мембраны и внутренней поверхности внутренней мембраны и на мембранах крист. Элементарные частица имеют обычно грибовидную форму и состоят из округлой «шляпки» диаметром 80—100 Å и цилиндрической ножки длиной около 55 Å. Материал, заключенный между мембранами, связывает оба слоя и поставляет «сырье» для синтеза ферментов мембран. Почти совершенно не выяснена роль матрикса. Предполагают, что в нем идут сложные процессы синтеза ряда органических соединений, в частности, белка.
До недавнего времени митохондрии рассматривались как замкнутые системы, имеющие сплошную наружную мембрану. Эта мембрана, обладая избирательной проницаемостью, контролирует поступление веществ в митохондрию и из нее. Некоторые ученые считают, что в наружной мембране митохондрии могут образовываться временные отверстия, открывающиеся непосредственно в гиалоплазму. Митохондрии довольно часто (особенно в период высокой активности клетки) соприкасаются с элементами эндоплазматической сети, а иногда прижимаются к ядерной оболочке. Отверстия, вероятно, позволяют осуществлять контакт между наружной мембраной митохондрии, мембраной эндоплазматической сети и ядерной оболочкой. Это дает возможность предполагать непосредственный перенос некоторых веществ из внутренней полости митохондриальных гребней. Митохондрии пассивно перемещаются в клетке током цитоплазмы и при этом постоянно меняют свою форму. В некоторых клетках, как предполагают, митохондрии способны и к активному движению, скапливаясь у тех участков цитоплазмы, которые в данный момент находятся в состоянии наиболее высокой активности.
В вопросе о происхождении митохондрий до сих пор нет полной ясности. Наиболее распространена точка зрения, согласно которой митохондрии являются постоянными клеточными органоидами, которые распределяются между дочерними клетками во время деления клетки. После этого число митохондрий увеличивается за счет их делений. Другая, более новая и завоевывающая все большее число сторонников точка зрения состоит в том, что митохондрии представляют собой непостоянные и недолго вечные органоиды, непрерывно образующиеся вновь и распадающиеся в течение жизни клетки. Одни ученые, придерживающиеся этой точки зрения, считают, что в образовании митохондрий принимает участие плазмалемма, другие считают, что ядерная оболочка, третьи — что аппарат Гольджи. С помощью микрокиносъемки удалось установить, что в отдельных случаях наружная мембрана хлоропластов способна вытягиваться и давать выросты, которые затем отрываются от тела хлоропласта и на глазах у наблюдателя образуют тельца, очень сходные с митохондриями. Некоторые ученые считают, что митохондрии могут дифференцироваться непосредственно в цитоплазме. Однако прямых доказательств различных способов возникновения митохондрий до сих пор нет. Деление митохондрий путем простой поперечной перетяжки можно наблюдать как в световой, так и в электронный микроскоп. Механизм деления и прежде всего поведение мембран и крист пока не выяснены. Митохондрии могут не только делиться, но и сливаться между собой.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
| Митохондрии — «силовые» станции клетки, в них локализована большая часть реакций дыхания (аэробная фаза). В митохондриях происходит аккумуляция энергии дыхания в аденозинтрифосфате (АТФ). Энергия, запасаемая в АТФ, служит основным источником для физиологической деятельности клетки. Митохондрии обычно имеют удлиненную палочковидную форму длиной 4—7 мкм и диаметром 0,5—2 мкм. Число митохондрий в клетке может быть различным от 500 до 1000 и зависит от роли данного органа в процессах энергетического обмена. Однако в некоторых организмах (дрожжах) имеется лишь одна гигантская митохондрия.
Электронно- микроскопическая фотография митохондрий (по П. В. Парамоновой) Химический состав митохондрий несколько колеблется. В основном это белково-липидные органеллы. Содержание белка в них составляет 60—65%, причем структурные и ферментативные белки содержатся примерно в равной пропорции, а также около 30% липидов. Очень важно, что митохондрии содержат нуклеиновые кислоты: РНК — 1 % и ДНК —0,5%. В митохондриях имеется не только ДНК, но и вся система синтеза белка, в том числе рибосомы. Митохондрии окружены двойной мембраной. Толщина мембран составляет 6—10 нм. Мембраны митохондрий на 70% состоят из белка. Фосфолипиды мембран представлены фосфатидтилхолином, фосфатидилэтаноламином, а также специфическими фосфолипидами, например, кардиолипином. Мембраны митохондрий не пропускают Н+ и служат барьером для их транспорта. Это важнейшее свойство лежит в основе образования электрохимического градиента. Между мембранами находится заполненное жидкостью перимитохондриальное пространство, равное 10 нм. Внутреннее пространство митохондрий заполняет матрикс в виде студнеобразной полужидкой массы. В матриксе сосредоточены ферменты цикла Кребса. Внутренняя мембрана дает выросты — кристы, расположенные перпендикулярно продольной оси органеллы и перегораживающие все внутреннее пространство митохондрий на отдельные отсеки. Однако, поскольку выросты-перегородки неполные, между этими отсеками сохраняется связь. Мембраны митохондрий обладают большой прочностью и гибкостью. Во внутренней мембране локализована дыхательная цепь (цепь переноса электронов). На внутренней поверхности внутренней мембраны митохондрий равномерно расположены грибовидные частицы. Каждая митохондрия содержит 104—105 таких грибовидных частиц, которые представляют собой фермент АТФ-синтазу, катализирующую образование АТФ.Митохондрии способны к движению. Это имеет большое значение в жизни клетки, так как митохондрии передвигаются к тем местам, где идет усиленное потребление энергии. Они могут ассоциировать друг с другом, как путем тесного сближения, так и при помощи связующих тяжей. Наблюдаются также контакты митохондрий с эндоплазматической сетью, ядром, хлоропластами. Известно, что митохондрии способны к набуханию, а при потере воды — к уменьшению объема. В растущих клетках в митохондриальном матриксе количество крист растет — это коррелирует с увеличением интенсивности дыхания. В процессе дыхания ультраструктура митохондрий меняется. В том случае, если в митохондриях протекает активный процесс преобразования энергии окисления в энергию АТФ, внутренняя часть митохондрий становится более компактной. Митохондрии имеют свой онтогенез. В меристематических клетках можно наблюдать инициальные частицы, которые представляют собой округлые образования, окруженные двойной мембраной. Диаметр таких инициальных частиц составляет 50 нм. По мере роста клетки инициальные частицы увеличиваются в размере, удлиняются и их внутренняя мембрана образует выросты, перпендикулярные оси митохондрий. Вначале образуются промитохондрии. Они еще не достигают окончательного размера и имеют мало крист. Из промитохондрии образуются митохондрии. Сформировавшиеся митохондрии делятся путем перетяжки или почкованием.
Схема строения митохондрии: 1 — наружная мембрана; 3 — матрикс; 2 — внутренняя мембрана; 4 — перимитохондриальное пространство. Свойства митохондрий (белки, структура) закодированы частично в ДНК митохондрий, а частично в ядре. Так, митохондриальный геном кодирует белки рибосом и частично систему переносчиков электронотранспортной цепи, а в геноме ядра кодирована информация о белках-ферментах цикла Кребса. Сопоставление размеров митохондриальной ДНК с числом и размером мито-хондриальных белков показывает, что в ней заложено информации почти для половины белков. Это и позволяет считать митохондрии, как и хлоропласты, полуавтономными, т. е. не полностью зависящими от ядра. Они имеют собственную ДНК и собственную белоксинтезирующую систему, и именно с ними и с пластидами связана так называемая цитоплазматическая наследственность. В большинстве случаев это наследование по материнской линии, так как инициальные частицы митохондрий локализованы в яйцеклетке. Таким образом, митохондрии всегда образуются от митохондрий. Широко обсуждается вопрос, как рассматривать митохондрии и хлоропласты с эволюционной точки зрения. Еще в 1921 г. русский ботаник Б.М. Козо-Полянский высказал мнение, что клетка — это симбиотрофная система, в которой сожительствует несколько организмов. В настоящее время эндосимбиотическая теория происхождения митохондрий и хлоропластов является общепринятой. Согласно этой теории, митохондрии — это в прошлом самостоятельные организмы. По мнению Л. Маргелис (1983), это могли быть эубактерии, содержащие ряд дыхательных ферментов. На определенном этапе эволюции они внедрились в примитивную, содержащую ядро, клетку. Оказалось, что ДНК митохондрий и хлоропластов по своей структуре резко отличается от ядерной ДНК высших растений и сходна с бактериальной ДНК (кольцевое строение, нуклеотидная последовательность). Сходство обнаруживается и по величине рибосом. Они мельче цитоплазматических рибосом. Синтез белка в митохондриях, подобно бактериальному, подавляется антибиотиком хлорамфениколом, который не влияет на синтез белка на рибосомах эукариот. Кроме того, система переноса электронов у бактерий расположена в плазматической мембране, что напоминает организацию электронтранспортной цепи во внутренней митохондриальной мембране. |


Митохондрия | Биология
Митохондрия – это двумембранный органоид эукариотической клетки, основная функция которого синтез АТФ – источника энергии для жизнедеятельности клетки.
Количество митохондрий в клетках не постоянно, в среднем от нескольких единиц до нескольких тысяч. Там, где процессы синтеза идут интенсивно, их больше. Также варьирует размер митохондрий и их форма (округлые, вытянутые, спиральные, чашевидные и др.). Чаще имеют округлую вытянутую форму, диаметром до 1 микрометра и длиной до 10 мкм. Могут перемещаться в клетке с током цитоплазмы или оставаться в одном положении. Перемещаются к местам, где больше всего требуется выработка энергии.
Согласно гипотезе симбиогенеза митохондрии произошли от аэробных бактерий, внедрившихся в другую прокариотическую клетку. Эти бактерии начали снабжать клетку дополнительным количеством молекул АТФ, а получать от нее питательные вещества. В процессе эволюции они потеряли автономность, передав часть своих генов в ядро и став таким образом клеточной органеллой.
В клетках новые митохондрии появляются в основном путем деления ранее существующих, т. е. они не синтезируются заново, что напоминает процесс размножения и говорит в пользу симбиогенеза.
Строение и функции митохондрии
Митохондрия состоит из
-
двух мембран — внешней и внутренней,
-
межмембранного пространства,
-
внутреннего содержимого — матрикса,
-
крист, представляющих собой выросты в матрикс внутренней мембраны,
-
собственной белок-синтезирующей системы: ДНК, рибосом, РНК,
-
белков и их комплексов, в том числе большого количества ферментов и коферментов,
-
других молекул и гранул различных веществ, находящихся в матриксе.

Внешняя и внутренняя мембраны выполняют разные функции, поэтому различается их химический состав. Расстояние между мембранами составляет до 10 нм. Внешняя мембрана митохондрий по строению схожа с плазмалеммой, окружающей клетку, и выполняет в основном барьерную функцию, отграничивая содержимое органоида от цитоплазмы. Через нее проникают мелкие молекулы, транспорт крупных избирателен. В некоторых местах внешняя мембрана соединена с ЭПС, каналы которой открываются в митохондрию.
На внутренней мембране, в основном ее выростах — кристах, располагаются ферменты, образуя мультиферментативные системы. Поэтому по химическому составу здесь преобладают белки, а не липиды. Количество крист варьирует в зависимости от интенсивности процессов. Так в митохондриях мышц их очень много.
В некоторых местах внешняя и внутренняя мембрана соединяются между собой.
У митохондрий, также как у хлоропластов, есть своя белоксинтезирующая система — ДНК, РНК и рибосомы. Генетический аппарат представляет собой кольцевую молекулу – нуклеоид, как у бактерий. Рибосомы митохондрий растений схожи с бактериальными, у животных митохондриальные рибосомы мельче не только цитоплазматических, но и бактериальных. Часть необходимых белков митохондрии синтезируют сами, другую часть получают из цитоплазмы, так как эти белки кодируются ядерными генами.
Главная функция митохондрий — снабжать клетку энергией, которая путем многочисленных ферментативных реакций извлекается из органических соединений и запасается в АТФ. Часть реакций идет с участием кислорода, в других выделяется углекислый газ. Реакции идут как в матриксе (цикл Кребса), так и на кристах (окислительное фосфорилирование).
Следует иметь в виду, что в клетках АТФ синтезируется не только в митохондриях, но и в цитоплазме в процессе гликолиза. Однако эффективность этих реакций невысока. Особенность функции митохондрий в том, что в них протекают реакции не только бескислородного окисления, но и кислородный этап энергетического обмена.
Другими словами, функция митохондрий – активное участие в клеточном дыхании, к которому относят множество реакций окисления органических веществ, переноса протонов водорода и электронов, идущих с выделением энергии, которая аккумулируется в АТФ.
Ферменты митохондрий
Ферменты транслоказы внутренней мембраны митохондрий осуществляют активный транспорт АДФ и АТФ.
В структуре крист выделяют элементарные частицы, состоящие из головки, ножки и основания. На головках, состоящих из фермента АТФазы, происходит синтез АТФ. АТФаза обеспечивает сопряжение фосфорилирования АДФ с реакциями дыхательной цепи.
Компоненты дыхательной цепи находятся в основании элементарных частиц в толще мембраны.
В матриксе находится большая часть ферментов цикла Кребса и окисления жирных кислот.
В результате активности электротранспортной дыхательной цепи ионы водорода поступают в нее из матрикса, а высвобождаются на наружной стороне внутренней мембраны. Это осуществляют определенные мембранные ферменты. Разница в концентрации ионов водорода по разные стороны мембраны приводит к возникновению градиента pH.
Энергию для поддержания градиента поставляет перенос электронов по дыхательной цепи. Иначе ионы водорода диффундировали бы обратно.
Энергия градиента pH используется для синтеза АТФ из АДФ:
АДФ + Ф = АТФ + h3O (реакция обратима)
Образующаяся вода ферментативно удаляется. Это, наряду с другими факторами, облегчает протекание реакции слева направо.
biology.su
структура, функции, особенности строения в связи с биологическими функциями.
⇐ ПредыдущаяСтр 4 из 13Следующая ⇒Митохондрии растительной клетки. Их структура и функции
Форма − округлые или гантелевидные тельца.
Размеры − длина 1-5 мкм, диаметром 0,4-0,5 мкм.
Количество в клетке − от десятков до 5 000.
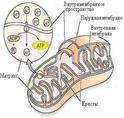
Структура. Состоят в основном из белка (60-65 %) и липидов (30 %). Это двухмембранные органоиды. Толщина наружной и внутренней мембран − 5-6 нм каждая. Перимитохондриальное пространство (промежуток между мемранами) заполнено жидкостью типа сыворотки. Внутренняя мембрана образует различной формы складки − кристы. На внутренней поверхности внутренней мембраны расположены грибовидные частицы − оксисомы, содержащие окислительные ферменты. Внутреннее содержимое митохондрий − матрикс. В матриксе содержатся рибосомы и митохондриальная ДНК (0,5 %), которая имеет кольцевое строение и отвечает за синтез белков митохондрий. Митохондрии имеют все типы РНК (1 %), делятся независимо от деления ядра, в клетке образуются от предсуществующих митохондрий путем деления или почкования. Полупериод жизни митохондрий − 5−10 дней.
Функции. Митохондрии являются центрами энергетической активности клеток. В митохондриях функционируют системы аэробного дыхания и окислительного фосфорелирования. Во внутренней мембране митохондрий локализованы компоненты электронтранспортной цепи и АТФ-синтетазные комплексы, осуществляющие транспорт электронов и протонов и синтез АТФ. В матриксе располагаются системы окисления ди- и трикарбоновых кислот, ряд систем синтеза липидов, аминокислот и др.
Митохондрии способны передвигаться к местам усиленного потребления энергии. Они могут ассоциировать друг с другом путем тесного сближения или при помощи тяжей. При анаэробном дыхании митохондрии исчезают.
Митохондрии имеют округлую и продолговатую форму диаметром 0,4–0,5 мкм и длиной 1–5 мкм (рис. 1.3).
 |
Количество митохондрий варьирует от единиц до 1 500–2 000 на растительную клетку.
Митохондрии ограничены двумя мембранами: наружной и внутренней, толщина каждой из них 5–6 нм. Наружная мембрана выглядит растянутой, а внутренняя образует складки, называемые гребнями (кристами), различной формы. Пространство между мембранами, в состав которого входит также внутреннее пространство крист, называется межмембранным (перимитохондриальным) пространством. Оно служит средой для внутренней мембраны и матрикса митохондрий.
Митохондрии в целом содержат 65–70 % белка, 25–30 % липидов и небольшое количество нуклеиновых кислот. 70 % от общего содержания липидов составляют фосфолипиды (фосфатидилхолин и фосфатидилэтаноламин). Жирнокислотный состав характеризуется высоким содержанием насыщенных жирных кислот, обеспечивающих «жесткость» мембраны.
В митохондриях локализованы системы аэробного дыхания и окислительного фосфорилирования. В результате дыхания расщепляются органические молекулы, и высвобождается энергия с передачей ее на молекулу АТФ.
Митохондрии содержат белки, РНК, тяжи ДНК, рибосомы, сходные с бактериальными, и различные растворенные вещества. ДНК существует в виде кольцевых молекул, располагающихся в одном или нескольких нуклеотидах.
Пластиды, наряду с вакуолями и клеточной оболочкой – характерные компоненты растительных клеток. Каждая пластида окружена собственной оболочкой, состоящей из двух элементарных мембран. Внутри пластид различают мембранную систему и более или менее гомогенное вещество – строму. Внутренняя структура хлоропласта довольно сложна. Строма пронизана развитой системой мембран, имеющих форму плоских пузырьков, называемых тилакоидами.Тилакоиды собраны в стопки – граны, напоминающие столбики монет.
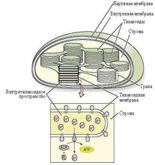 |
Хлоропласты, в которых протекает фотосинтез, содержат хлорофиллы и каротиноиды. Размер – 4–5 мкм. В одной клетке мезофилла листа может содержаться 40–50 хлоропластов, в мм2 листа – около 500 000. В цитоплазме хлоропласты обычно располагаются параллельно клеточной оболочке.
Хлорофиллы и каротиноиды встроены в тилакоидные мембраны. Хлоропласты зеленых растений и водорослей часто содержат зерна крахмала и мелкие липидные (жировые) капли. Крахмальные зерна – это временные хранилища продуктов фотосинтеза. Они могут исчезнуть из хлоропластов, находящихся в темноте всего лишь 24 ч и появиться вновь уже через 3–4 ч после переноса растений на свет.
В изолированных хлоропластах осуществляется синтез РНК, который обычно контролируется только хромосомной ДНК. Образование хлоропластов и синтез находящихся в них пигментов в значительной степени контролируется хромосомной ДНК, малопонятным образом взаимодействующей с ДНК хлоропластов. Тем не менее, в отсутствие собственной ДНК хлоропласты не формируются.
Хлоропласты могут считаться основными клеточными органеллами, поскольку первыми стоят в цепи преобразования солнечной энергии, в результате которого мы получаем пищу и топливо. В хлоропластах протекают не только фотосинтез. Они участвуют в синтезе аминокислот и жирных кислот, служат хранилищем временных запасов крахмала.
Хромопласты (от греческого сhroma – цвет) – пигментированные пластиды. Многообразные по форме хромопласты не содержат хлорофилла, но синтезируют и накапливают каротиноиды, которые придают желтую, оранжевую и другую окраску. Корнеплоды моркови, плоды томатов окрашены пигментами, которые находятся в хромопластах.
Лейкопласты являются местом накопления запасного вещества – крахмала. Особенно много лейкопластов в клетках клубней картофеля. На свету лейкопласты могут преобразовываться в хлоропласты (клубни картофеля зеленеют). Осенью хлоропласты преобразуются в хромопласты и зеленые листья, и плоды желтеют и краснеют.
Читайте также:
lektsia.com
описание, строение и функции :: SYL.ru
Что такое митохондрии? Если ответ на этот вопрос вызывает у вас затруднения, то наша статья как раз для вас. Мы рассмотрим особенности строения этих органелл во взаимосвязи с выполняемыми функциями.
Что такое органеллы
Но для начала давайте вспомним, что такое органеллы. Так называют постоянные клеточные структуры. Митохондрии, рибосомы, пластиды, лизосомы... Все это органеллы. Подобно самой клетке, каждая подобная структура имеет общий план строения. Органеллы состоят из поверхностного аппарата и внутреннего содержимого - матрикса. Каждую из них можно сравнить с органами живых существ. Органеллы также имеют свои характерные черты, обусловливающие их биологическую роль.

Классификация клеточных структур
Органеллы объединяют в группы по признаку строения их поверхностного аппарата. Различают одно-, дву- и немембранные постоянные клеточные структуры. К первой группе относятся лизосомы, комплекс Гольджи, эндоплазматический ретикулум, пероксисомы и различные виды вакуолей. Ядро, митохондрия и пластиды - двумембранные. А рибосомы, клеточный центр и органеллы движения полностью лишены поверхностного аппарата.

Теория симбиогенеза
Что такое митохондрии? Для эволюционного учения это не просто структуры клетки. Согласно симбиотической теории, митохондрии и хлоропласты являются результатом метаморфоз прокариот. Вполне возможно, что митохондрии произошли от аэробных бактерий, а пластиды - от фотосинтезирующих. Доказательством этой теории является тот факт, что данные структуры имеют собственный генетический аппарат, представленный кольцевой молекулой ДНК, двойную мембрану и рибосомы. Существует также предположение, что в дальнейшем от митохондрий произошли животные эукариотические клетки, а от хлоропластов - растительные.

Расположение в клетках
Митохондрии являются составляющей частью клеток преобладающей части растений, животных и грибов. Отсутствуют они только у анаэробных одноклеточных эукариот, обитающих в бескислородной среде.
Строение и биологическая роль митохондрий долгое время оставались загадкой. Впервые при помощи микроскопа их удалось увидеть Рудольфу Келликеру в 1850 году. В мышечных клетках ученый обнаружил многочисленные гранулы, которые на свету были похожи на пух. Понять, какова роль этих удивительных структур, стало возможно благодаря изобретению профессора Пенсильванского университета Бриттона Ченса. Он сконструировал прибор, который позволял видеть сквозь органеллы. Так была определена структура и доказана роль митохондрий в обеспечении энергией клеток и организма в целом.
Форма и размер митохондрий
Митохондрии могут иметь форму палочек, нитей или круглых телец. Длина их достигает от 0, 5 до 10 мкм. Количество данных органелл в клетке напрямую зависит от интенсивности протекающих в ней обменных процессов. К примеру, у одноклеточных паразитических жгутиконосцев - трипаносом - в клетке расположена единственная крупная митохондрия. Тогда как в амебе может находиться до пятисот тысяч этих структур.

Общий план строения
Рассмотрим, что такое митохондрии с точки зрения особенностей их строения. Это двумембранные органеллы. Причем наружная - гладкая, а внутренняя имеет выросты. Матрикс митохондрий представлен различными ферментами, рибосомами, мономерами органических веществ, ионами и скоплениями кольцевых молекул ДНК. Такой состав делает возможным протекание важнейших химических реакций: цикла трикарбоновых кислот, мочевины, окислительного фосфорилирования.
Значение кинетопласта
Эта структура наблюдается только у паразитических одноклеточных организмов - трипаносом или лейшманий. Кинетопласт внутри единственной гигантской митохондрии. Он представляет собой хорошо выраженное скопление ДНК. Эта структура практически всегда располагается у основания жгутика, эффективно обеспечивая его необходимой для движения в вязкой среде энергией. Кинетопласт имеет форму мини- и максиколец. Ученые доказали, что если паразит утрачивает свою специфичную ДНК, он не может существовать в организме насекомого. Все дело в том, что в его кинетопласте закодирована информация о единице фермента, который необходим для осуществления процесса фосфорилирования. Однако такие организмы способны развиваться в тканях позвоночных, где получают энергию в ходе процессе гликолиза.

Мембрана митохондрии
Мембраны митохондрий не одинаковы по своему строению. Замкнутая наружная является гладкой. Она образована бислоем липидов с фрагментами белковых молекул. Его общая толщина составляет 7 нм. Данная структура выполняет функции отграничения от цитоплазмы , а также взаимосвязи органеллы с окружающей средой. Последняя возможна благодаря наличию белка порина, который формирует каналы. По ним посредством активного и пассивного транспорта передвигаются молекулы.
Химическую основу внутренней мембраны составляют белки. Она образует внутри органоида многочисленные складки - кристы. Эти структуры в значительной степени увеличивают активную поверхность органеллы. Главной особенностью строения внутренней мембраны является полная непроницаемость для протонов. В ней не образуются каналы для проникновения ионов извне. В отдельных местах наружная и внутренняя соприкасаются. Здесь расположен особый рецепторный белок. Это своеобразный проводник. С его помощью митохондриальные белки, которые закодированы в ядре, проникают внутрь органеллы. Между мембранами находится пространство, толщиной до 20 нм. В нем расположены различные виды белков, которые являются обязательными компонентами дыхательной цепи.

Функции митохондрий
Строение митохондрии напрямую взаимосвязано с выполняемыми функциями. Основная из них заключается в осуществлении синтеза аденозинтрифосфата (АТФ). Это макромолекула, которая случит основным переносчиком энергии в клетке. В ее состав входит азотистое основание аденин, моносахарид рибоза и три остатка фосфорной кислоты. Именно между последними элементами заключено основное количество энергии. При разрыве одной из них максимально ее может выделиться до 60 кДж. В целом прокариотическая клетка содержит 1 млрд молекул АТФ. Эти структуры постоянно находятся в работе: существование каждой из них в неизменном виде не продолжается больше одной минуты. Молекулы АТФ постоянно синтезируются и расщепляются, обеспечивая организм энергией в тот момент, когда это необходимо.
По этой причине митохондрии называют "энергетическими станциями". Именно в них происходит окисление органических веществ под действием ферментов. Энергия, которая при этом образуется, запасается и хранится в виде АТФ. К примеру, при окислении 1 г углеводов образуется 36 макромолекул этого вещества.
Строение митохондрии позволяет им выполнять еще одну функцию. Благодаря своей полуавтономности они являются дополнительным носителем наследственной информации. Ученые установили, что ДНК самих органелл не могут функционировать самостоятельно. Дело в том, что они не содержат всех необходимых для своей работы белков, поэтому заимствуют их в наследственном материале ядерного аппарата.
Итак, в нашей статье мы рассмотрели, что такое митохондрии. Это двумембранные клеточные структуры, в матриксе которых осуществляется ряд сложных химических процессов. Результатом работы митохондрий является синтез АТФ - соединение, которое обеспечивает организм необходимым количеством энергии.
www.syl.ru
Особенности строения митохондрий растений. — КиберПедия
Митохондрии — «силовые» станции клетки, в них локализована большая часть реакций дыхания (аэробная фаза). В митохондриях происходит аккумуляция энергии дыхания в аденозинтрифосфате (АТФ). Энергия, запасаемая в АТФ, служит основным источником для физиологической деятельности клетки. Митохондрии обычно имеют удлиненную палочковидную форму длиной 4—7 мкм и диаметром 0,5—2 мкм. Число митохондрий в клетке может быть различным от 500 до 1000 и зависит от роли данного органа в процессах энергетического обмена. Однако в некоторых организмах (дрожжах) имеется лишь одна гигантская митохондрия. Химический состав митохондрий несколько колеблется. В основном это белково-липидные органеллы. Содержание белка в них составляет 60—65%, причем структурные и ферментативные белки содержатся примерно в равной пропорции, а также около 30% липидов. Очень важно, что митохондрии содержат нуклеиновые кислоты: РНК — 1 % и ДНК —0,5%. В митохондриях имеется не только ДНК, но и вся система синтеза белка, в том числе рибосомы. Митохондрии окружены двойной мембраной. Толщина мембран составляет 6—10 нм. Мембраны митохондрий на 70% состоят из белка. Фосфолипиды мембран представлены фосфатидтилхолином, фосфатидилэтаноламином, а также специфическими фосфолипидами, например, кардиолипином. Мембраны митохондрий не пропускают Н+ и служат барьером для их транспорта. Это важнейшее свойство лежит в основе образования электрохимического градиента. Между мембранами находится заполненное жидкостью перимитохондриальное пространство, равное 10 нм. Внутреннее пространство митохондрий заполняет матрикс в виде студнеобразной полужидкой массы. В матриксе сосредоточены ферменты цикла Кребса. Внутренняя мембрана дает выросты — кристы, расположенные перпендикулярно продольной оси органеллы и перегораживающие все внутреннее пространство митохондрий на отдельные отсеки. Однако, поскольку выросты-перегородки неполные, между этими отсеками сохраняется связь. Мембраны митохондрий обладают большой прочностью и гибкостью. Во внутренней мембране локализована дыхательная цепь (цепь переноса электронов). На внутренней поверхности внутренней мембраны митохондрий равномерно расположены грибовидные частицы. Каждая митохондрия содержит 104—105 таких грибовидных частиц, которые представляют собой фермент АТФ-синтазу, катализирующую образование АТФ.Митохондрии способны к движению. Это имеет большое значение в жизни клетки, так как митохондрии передвигаются к тем местам, где идет усиленное потребление энергии. Они могут ассоциировать друг с другом, как путем тесного сближения, так и при помощи связующих тяжей. Наблюдаются также контакты митохондрий с эндоплазматической сетью, ядром, хлоропластами. Известно, что митохондрии способны к набуханию, а при потере воды — к уменьшению объема. В растущих клетках в митохондриальном матриксе количество крист растет — это коррелирует с увеличением интенсивности дыхания. В процессе дыхания ультраструктура митохондрий меняется. В том случае, если в митохондриях протекает активный процесс преобразования энергии окисления в энергию АТФ, внутренняя часть митохондрий становится более компактной. Митохондрии имеют свой онтогенез. В меристематических клетках можно наблюдать инициальные частицы, которые представляют собой округлые образования, окруженные двойной мембраной. Диаметр таких инициальных частиц составляет 50 нм. По мере роста клетки инициальные частицы увеличиваются в размере, удлиняются и их внутренняя мембрана образует выросты, перпендикулярные оси митохондрий. Вначале образуются промитохондрии. Они еще не достигают окончательного размера и имеют мало крист. Из промитохондрии образуются митохондрии. Сформировавшиеся митохондрии делятся путем перетяжки или почкованием.
строения митохондрии: наружная мембрана; матрикс; внутренняя мембрана; перимитохондриальное пространство.
Свойства митохондрий (белки, структура) закодированы частично в ДНК митохондрий, а частично в ядре. Так, митохондриальный геном кодирует белки рибосом и частично систему переносчиков электронотранспортной цепи, а в геноме ядра кодирована информация о белках-ферментах цикла Кребса. Сопоставление размеров митохондриальной ДНК с числом и размером мито-хондриальных белков показывает, что в ней заложено информации почти для половины белков. Это и позволяет считать митохондрии, как и хлоропласты, полуавтономными, т. е. не полностью зависящими от ядра. Они имеют собственную ДНК и собственную белоксинтезирующую систему, и именно с ними и с пластидами связана так называемая цитоплазматическая наследственность. В большинстве случаев это наследование по материнской линии, так как инициальные частицы митохондрий локализованы в яйцеклетке. Таким образом, митохондрии всегда образуются от митохондрий. Широко обсуждается вопрос, как рассматривать митохондрии и хлоропласты с эволюционной точки зрения. Еще в 1921 г. русский ботаник Б.М. Козо-Полянский высказал мнение, что клетка — это симбиотрофная система, в которой сожительствует несколько организмов. В настоящее время эндосимбиотическая теория происхождения митохондрий и хлоропластов является общепринятой. Согласно этой теории, митохондрии — это в прошлом самостоятельные организмы. По мнению Л. Маргелис (1983), это могли быть эубактерии, содержащие ряд дыхательных ферментов. На определенном этапе эволюции они внедрились в примитивную, содержащую ядро, клетку. Оказалось, что ДНК митохондрий и хлоропластов по своей структуре резко отличается от ядерной ДНК высших растений и сходна с бактериальной ДНК (кольцевое строение, нуклеотидная последовательность). Сходство обнаруживается и по величине рибосом. Они мельче цитоплазматических рибосом. Синтез белка в митохондриях, подобно бактериальному, подавляется антибиотиком хлорамфениколом, который не влияет на синтез белка на рибосомах эукариот. Кроме того, система переноса электронов у бактерий расположена в плазматической мембране, что напоминает организацию электронтранспортной цепи во внутренней митохондриальной мембране.
cyberpedia.su
Митохондрии растений - Справочник химика 21
Дыхательная цепь митохондрий растений изучена гораздо хуже, чем дыхательная цепь животных митохондрий. Известно, однако, что [c.69] Ядра, митохондрии и хлоропласты растений, как сообщалось, также синтезируют белок. Ядра, тщательно выделенные из гороха, включают в белок меченый лейцин в присутствии других белковых аминокислот и АТФ. Синтез белка в митохондриях растений показан не очень убедительно, так как не было исключено загрязнение данной фракции рибосомами. Препараты хлоропластов, как обнаружено, также катализируют включение аминокислот в белок. [c.482]В митохондриях растений до сих пор не было осуществлено выделение отдельных компонентов фосфорилирования в дыхательной цепи. [c.254]
Окисление пировиноградной кислоты митохондриями растений многократно подтверждено, но попытки выделить растворимые препараты фермента оказались безуспешными. [c.193]
X. Дыхательная цепь в митохондриях растений........................69 [c.619]
И Цыбакова высказали предположение, что этот новый тип фосфорилирования сопряжен с переходом водорода от НАД-Нг к кислороду через реакции дыхательной цепи. Последующие работы подтвердили это предположение и показали, что при переходе каждой пары водородных атомов или электронов через реакции дыхательной цепи может происходить максимум три фосфорилирования. Прямое доказательство того, что фосфорилирование происходит не только на уровне субстратов, но и на уровне переносчиков водорода, было получено в 1951 г., когда Ленинджер показал, что при окислении НАД-Нг кислородом в митохондриях печени крысы отношение Р/0 близко к трем. Фосфорилирование в дыхательной цепи у растений впервые показали Миллер, Боннер, Аксельрод и Бандурский [22], которые использовали митохондрии, выделенные из проростков маша, и кислоты цикла Кребса в качестве субстратов. Эти авторы получили более низкое отношение Р/0 по сравнению с найденным для митохондрий животного происхождения. Поэтому возникло предположение, что митохондрии растений не эффективны как фосфорилирующие системы. Более поздние исследования показали, что митохондрии растений окисляют промежуточные продукты цикла Кребса, причем отношение Р/0 сравнимо с отношением, полученным для митохондрий животных. [c.243]
Данные, полученные с тканями и митохондриями растений, позволяют предполагать, что схема Р-окисления у растений такая [c.308]
На тонких срезах изолированных растительных митохондрий видно, что по структуре они очень похожи на митохондрии животных с той разницей, что в последних кристы образуют параллельные ряды, чего в растительных митохондриях не наблюдается. Кроме того, в митохондриях растений, в отличие от митохондрий животных, кристы часто соединяются друг с другом, образуя замкнутые петли. На фото 8 представлены ультратонкие срезы митохондрий, позволяющие рассмотреть их внутреннюю структуру. Мембраны растительных митохондрий были изучены Парсонсом и др. [77]. На внешней мембране совершенно отчетливо обнаруживаются углубления диаметром 25—30 А, неравномерно распределенные по ее поверхности (фото 9). На внутренней мембране видны мельчайшие субъединицы, похожие по виду на булавочные головки (фото 10) такие субъединицы ранее были обнаружены в митохондриях животных [34, 70]. В 1965 г. Парсонс и др. [77] показали на митохондриях тыквы, что эти субъединицы внутренних мембран имеют диаметр 90— 110 А нри толщине стебелька 35—40 А и длине 45 А (фото И). Функциональная роль этих субъединиц неизвестна. [c.57]
Цитохромоксидаза обнаружена ферментативным путем и оптическими методами во всех исследованных до сих пор растительных митохондриях. Оксидаза из митохондрий растений отличается по своему спектру поглощения от фермента животных смещением полос в сторону более коротких волн. [c.63]
Размеры рибосом, выделенных из различных органелл, варьируют. Митохондриальные рибосомы низших эукариот, таких, как грибы, имеют несколько большие размеры, чем рибосомы Е. соИ. В митохондриях растений рибосомы лишь ненамного меньше, чем в окружающей цитоплазме. Однако в митохондриях млекопитающих и амфибий их размеры еще меньше и составляют 60S. Эти рибосомы характеризуются более низким содержанием РНК (25-31%). Рибосомы хлоропластов приблизительно такого же размера, как и рибосомы бактерий, хотя с более высоким содержанием РНК. [c.104]
Регуляцию дыхания можно лучше всего показать на митохондриях животных (фиг. 66, А). Никакого поглощения кислорода не происходит, когда митохондрии инкубируют в присутствии субстрата (глутаминовой кислоты), фосфата и кислорода. Быстрое поглощение кислорода начинается только после добавления АДФ и продолжается до тех пор, пока концентрация АДФ не уменьшится до низкого уровня в результате фосфорилирования в дыхательной цепи. Эту последовательность явлений можно повторить, снова добавляя АДФ. В митохондриях растений (фиг. 66, Б) заметное поглощение кислорода происходит и в отсутствие АДФ. Это показывает, что использованные в данной работе митохондрии растений не имели таких прочно сопряженных систем, как препараты митохондрий из животных тканей. Добавление АДФ к митохондриям растений вызывает значительное усиление поглощения кислорода. Затем интенсивность поглощения кислорода постепенно уменьшается по мере того, как добавленный АДФ фосфорилируется в АТФ. Дальнейшие прибавки АДФ приводят к повторению всей последователь- [c.244]
X. ДЫХАТЕЛЬНАЯ ЦЕНЬ В МИТОХОНДРИЯХ РАСТЕНИЙ [c.69]
В растительных митохондриях обнаруживается зависящее от энергии восстановление НАД сукцинатом, по здесь в отличие от митохондрий животных для этого используется только энергия АТФ. В митохондриях растений в присутствии олигомицина сукцинат восстанавливает лишь незначительную часть всего пиридиннуклеотида [8], что не согласуется с характером аналогичного процесса в митохондриях животных. В митохондриях животных наблюдаются большие изменения скорости поглощения кислорода и окислительно-восстановительных уровней переносчиков при добавке двухвалентных катионов [14, 24]. В то же время в случае митохондрий корня свеклы и митохондрий маша добавление двухвалентных катионов сопровождается лишь небольшим ускорением дыхания (Боннер мл. и Робертсон, неопубликованные данные), а сдвиг окислительно-восстановительного состояния переносчиков при этом незначителен. [c.72]
Андреева И. Н. О состоянии митохондрий растений в связи с репарационными процессами после перенесения высоких температур // Физиология засухоустойчивости растений. М. Наука, 1971. С, 70—85. [c.247]
Неоднократно было обнаружено, что феназинметасульфат служит наиболее эффективным акцептором водорода для флавопротеид-ных дегидрогеназ. Сукцинатдегидрогеназа тесно связана с митохондриальными мембранами, но она была переведена в растворимое состояние и получена в чистом виде из митохондрий сердца быка. Очищенные препараты содержали двухвалентное железо и 1 моль флавина на 1 моль белка. По-видимому, флавин связан с белком ковалентными связями. Сравнительно недавно из митохондрий растений были получены препараты растворимой сукцииатдегидро-геназы [14]. [c.191]
Исходя из термодинамических соображений, можно предположить, что восстановление глутаминовой кислоты в ее полуальдегид скорее всего не является одноступенчатой реакцией, а требует какого-то активирования карбоксильной группы (ср. с превращением аспарагиновой кислоты в ее полуальдегид). Полуальдегид глутаминовой кислоты претерпевает спонтанную циклизацию до установления динамического равновесия с пирролин-5-карбо-новой кислотой. Восстановление пирролин-5-карбоновой кислоты в пролин наблюдалось у бактерий, грибов и животных. Окисление пролина осуществляют митохондрии растений, но механизм этого процесса не известен. [c.408]
Почти полная идентичность генетического кода у всех организмов служит убедительным доводом в пользу того, что все клетки произошли от общего предшественника. Как же в этом случае объяснить некоторые отличия генетического кода митохондрий Приблизиться к пониманию этого помогли недавно полученные данные о различии генетического кода в митохондриях разных организмов. Папример, триплет UGA, служащий в универсальном коде стоп-кодоном, в митохондриях млекопитающих, грибов и простейших кодирует триптофан, но в митохондриях растений используется как стоп-кодон. Аналогичным образом триплет AGG, обычно кодирующий аргинин, в митохондриях млекопитающих обозначает сигнал "stop", а у дрозофилы кодирует серин (табл. 7-4). Подобные отклонения указывают на то, что в генетическом коде митохондрий могут происходить случайные перемены. Вероятно, возможность появления и закрепления в потомстве случайных изменений в значении кодона связана с необычайно малым числом белков, кодируемых митохондриальным геномом в большом геноме подобные изменения привели бы к нарушению функции многих белков и, как следствие, к гибели клетки. [c.491]
Суммарные водорастворимые белки мицелия Р. ostreatus разобщают процессы окислительного фосфорилирования митохондрий растений в экспериментах in vitro. Белки из охлажденного мицелия Р. ostreatus обладают большей разобщающей активностью, чем из неохлажденного мицелия. [c.52]
Среди механизмов загциты растений от низкотемпературного стресса выделяется один механизм, связанный с термогенезом в митохондриях, который, как считалось раньгие, присущ только животным организмам. Однако в митохондриях растений были также обнаружены системы, способные диссипировать энергию с образованием тепла. [c.104]Третьей системой, вызывающей при своем функционировании разобщение оьсисления и фосфорилирования в митохондриях растений, является стрессовый белок БХШ 310. Молекулярная масса этого нативного белка составляет 310 ьсДа. Показано, что этот белок состоит из двух типов субъединиц с молекулярными массами 56 и 66 ьсДа (Колесниченко и др., [c.116]
Дыхательная цепь митохондрий растений и животных имеет сходную организацию, основные отличия касаются цианид (СК )-резистентного пути переноса электронов и строения НАДН-дегидрогеназного сегмента дыхательной цепи. Растительные митохондрии способны оьсислять как эндогенный, так и экзогенный НАДН (Dou e, 1985). Все переносчики электронов в митохондриях сгруппированы в четыре комплекса. [c.135]
Комплекс III (убихинон-цитохром с-оксидоредуктаза) переносит электроны от восстановленного убихинона к цитохрому с. Его структурная организация в митохондриях растений изучена пока слабо. Известно, что в его состав входят цитохромы bseo и bsee, цитохром i и железо-серный белок Риске. Этот комплекс чувствителен к антимицину А (Шугаев, 1991). [c.135]
chem21.info









