
Местообитание как экологическое понятие. Местообитание растения это
Местообитание
участок суши или водоёма, занятый организмом, группой особей одного вида, Биоценозом или синузией (См. Синузия) и обладающий всеми необходимыми для их существования условиями (климат, рельеф, почва, пища и др.). М. вида — совокупность отвечающих его экологическим требованиям участков в пределах видового Ареала, М. популяции — часть М. вида, обеспечивающая существование отдельной популяции (См. Популяция), М. особи — конкретный участок, занятый данным индивидом во всех фазах его развития. Различают также М. семьи, стада, стаи или колонии животных, а также зарослей, куртин и др. совокупностей растений. По широте использования М. выделяют Стенотопные организмы, занимающие только однотипные М., и Эвритопные организмы, проявляющие способность занимать в пределах своего ареала разнообразные М. У многих видов М. меняется в зависимости от стадии развития. Так, личинки земноводных обычно обитают в воде, взрослые животные — на суше. Многие паразиты имеют покоящуюся фазу, сохраняющуюся во внешней среде, и активную фазу, обитающую в теле хозяина, часто лишь в определённых его органах; разные фазы развития многих паразитных растений связаны с различными растениями-хозяевами. Часть М. вида, занимаемая им на ограниченный период (сезон, период суток), для определённой цели (питание, размножение), называемой стацией (См. Стация). М. биоценоза называемого Биотопом.
Лит.: Наумов Н. П., Экология животных, 2 изд., М., 1963; Основы лесной биогеоценологии, под ред. В. Н. Сукачева и Н. В. Дылиса, М., 1964.
Н. П. Наумов.
Поделитесь на страничкеslovar.wikireading.ru
местообитание вида
Местообитание вида - это пространственно ограниченная совокупность условий абиотической и биотической среды, обеспечивающая весь цикл развития особей (или группы особей) одного вида.[ ...]
Местообитание — это территория или акватория, занимаемая популяцией (видом), с комплексом присущих ей экологических факторов. Местообитание вида является компонентом его экологической ниши. Применительно к наземным животным местообитание вида называется стация, местообитание сообщества — биотоп.[ ...]
Ценный для науки вид Ареал его сокращается в связи с освоением территорий с местообитанием вида. Необходима организация отдельных заказников в Степи и Лесостепи.[ ...]
Стация - это часть местообитания вида, которая характеризуется особыми экологическими условиями, необходимыми для существования данного вида, и используется либо в ограниченное время (сезонно, суточно), либо для ог-раниченных целей (для питания, переживания неблагоприятных ситуаций, размножения и др.) .[ ...]
Так, например, летние местообитания видов серой утки приурочены к пойменным старицам тайги, бекасиных - к низинным евтрофным болотам пойм рек. Глухарь в весеннее время обитает вблизи мест токов по окраинам верховых торфяников. Рябчик питается и выводит птенцов в темнохвойных урёмах по днищам долин ручьев и таежных речек.[ ...]
Так, например, летние местообитания видов серой утки приурочены к пойменным старицам тайги, бекасиных - к низинным евтрофным болотам пойм рек. Глухарь в весеннее время обитает вблизи мест токов по окраинам верховых торфяников. Рябчик питается и выводит птенцов в темнохвойных урёмах по днищам долин ручьев и таежных речек.[ ...]
Роль конкуренции в выборе местообитания схематически иллюстрируется фиг. 103. Кривые характеризуют местообитания видов, лежащие в пределах крайних возможных для жизни границ. На графиках обозначены также оптимальные для жизни границы. При наличии конкуренции с другим близкородственным или экологически сходным видом диапазон местообитаний, возможных для данного вида, сокращается до оптимальных границ (т. е. вид распространяется в наиболее благоприятных для него местообитаниях, в которых он обладает теми или иными преимуществами по сравнению со своими конкурентами). Если же межвидовая конкуренция выражена слабо, то под влиянием внутривидовой конкуренции популяция расширяет границы своего местообитания. Тенденция к увеличению заселяемого пространства хорошо прослеживается на островах, на которых отсутствуют потенциальные конкуренты. Например, полевка М1сго1из часто заселяет лесные зоны на тех островах, где отсутствует ее лесной конкурент, полевка Clethrion.om.ys (см. Камерон, 1964). Кроуэлл (1962) обнаружил, что численность и краевые местообитания кардинала на Бермудских островах больше, чем на континенте, так как здесь отсутствуют многие его конкуренты.[ ...]
Таким образом, для сохранения редких видов часто не обязательно применение жестких форм охраны, достаточно сохранение видов-строителей сообществ через регулирование предельно допустимых нагрузок для сохранения типичных условий местообитания вида. Для успешного решения задачи сохранения биоразнообразия необходимы паспортизация сообществ, включающих редкие виды, определение лимитирующего фактора и предельно допустимой величины его влияния.[ ...]
Необходима полная охрана всех известных местообитаний вида и наблюдение за состоянием популяций.[ ...]
Территорию, объединяющую стации какого-либо вида-насекомого (и любого животного), называют его ареалом или географическим распространен и ем; границами ареала служат географические пункты, за пределами которых вид не ¡может размножаться. В пределах своего ареала любой ви обычно встречается не повсеместно, а лишь в свойственных ему биотопах, сумму этих биотопов называют местообитанием вида. Местообитание вида исторически слагается в экологической связи с другими компонентами биоценоза.[ ...]
Мы можем выбрать три разных аспекта зависимости вида от окружающей среды. Ареал вида — его географический ранг, его распространение в пространстве, которое можно показать на карте. Местообитание вида — тот тип среды, где вид встречается; эта среда может характеризоваться физическими и химическими показателями, высотой над уровнем моря, положением в рельефе и т. д. или типом сообщества. Вид в различных частях ареала может занимать ряд различных местообитаний, то есть более чем один специфический тип местообитания. В пределах каждого местообитания можно описать позицию вида в пространстве, времени и функциональные связи с природным сообществом, занимающим это местообитание. Позиция вида в сообществе в зависимости от других видов является его нишей.[ ...]
Образ жизни. На местах гнездования появляются ранней весной. Характерное местообитание вида — горные леса, преимущественно у их верхней границы. Есть предположения о гнездовании в негустых предгорных и равнинных лесах из лиственницы, кедра, ели, пихты, возможно — в смешанных лесах, на рямах, зарастающих гарях, вырубках.[ ...]
Альфа- и бета-разнообразие могу! рассматриваться как результат многообразия ниш и местообитаний видов. Оба типа могут параллельно изменяться Вдоль Некоторых климатических градиентов, например, оба нарастаю! в направлении от прибрежных секвойевых лесов в Глубь континента, к горным территориям, которые охарактеризованы верхней трансектой на рис. 4-2. При изучении изменения сообществ птиц по градиенту разнообразия от холодных климатов к тропикам Макартур обнаружил известную независимость этих двух типов. Альфа-разнообразие для птиц, тесно связанное, как мы уже отмечали, со структурой растительности, оказалось не намного выше в тропиках по сравнению с лесами умеренной полосы. В то же время бета-разнообразие возрастало в тропиках, и притом резким скачком. В ЭТОМ различии проявилось существование так называемой !очкй насыщения, которая означает максимально возможное разделение пространства ниши для птиц при данной структуре растительности. Эволюция в тропиках не ведет к увеличению альфа-разнообразия выше уровня насыщения, но обеспечивает внедрение дополнительного числа видов ВДоль градиентов среды путем дифференциации месМобйтаний и сужения распределения местообитаний, что й Приводит к возрастанию бета-разнообразия.[ ...]
Возрастание масштабов практически неуправляемого разрушения природных комплексов, местообитаний видов растений и животных ведет, как мы это уже обсуждали, к снижению численности все большего числа видов до критического уровня, обрекая их на исчезновение. Количество известных науке видов высших растений превышает 500 тысяч, а животных — 2 миллиона. Общее количество видов животных, по расчетам на основе специальных экспериментов с частотой встречаемости новых видов, составляет, по-видимому, не менее 10, а может быть и 30 миллионов. При этом значительная, если не большая, часть видов таких многочисленных и разнообразных классов, как круглые черви, паукообразные и насекомые, особенно обитатели все еще недостаточно изученных тропических лесов, до сих пор не описаны и остаются неизвестными науке. Поэтому речь идет о необходимости предотвращения вымирания сотен тысяч и миллионов видов.[ ...]
При прохождении жизненного цикла животные в различные периоды жизни концентрируются в различных видах ПТК. Животные осваивают не отдельные фации, типы леса, а их сочетания. В одних они кормятся, жируют, в других - укрываются от нападения, в третьих - занимаются брачными играми, в четвертых - спасаются от гнуса, пятые используются для миграции и переходов. Стации животных - места их обитания можно подразделить на кормовые, гнездовые, токовые и миграционные, а по длительности и времени пребывания - на годичные, сезонные, суточные. Можно говорить о стациальной структуре местообитаний ландшафта. Она будет характеризоваться сочетанием ландшафтных фаций в урочищах, сочетанием урочищ в местностях, видов местностей в ландшафтах. В целом - это ландшафтные и экологические ареалы местообитания видов.[ ...]
Многим известна водоросль каулерпа (Саи-lerpa)—типичный представитель семейства каулерповых (Caulerpaceae). Этот обширный род (около 60 видой) весьма характерен для флоры тропических морей. Лишь немногие виды заходят в субтропические широты, и среди них распространенная в Средиземном море каулерпа прорастающая (С. proliféra). Крупные стелющиеся растения каулерпы часто покрывают большие пространства морского дна. Поселяясь обычно на различных твердых грунтах — камнях, скалистых рифах, мертвых кораллах, каулерпа благодаря своему стелющемуся слоевищу и наличию многих ризоидов неплохо чувствует себя также на песчаных и илистых грунтах. Излюбленным местообитанием видов каулерпы является мелководье с более или менее спокойной водой, в удалении от постоянного прибоя, например лагуны, защищенные от действия прибоя коралловыми рифами.[ ...]
БИОТ0П [био... +гр. topos местность] — 1) относительно однородное по абиотическим факторам среды пространство, занятое биоценозом-, 2) синоним местообитания вида.[ ...]
В географически протяженных местообитаниях видов должно быть больше, чем в небольших периферических местообитаниях; поскольку в глубоководных морях абиссальные области занимают обширные площади, необычайно высокое разнообразие морских беспозвоночных, описанное Сандерсом [230], возможно, просто отражает эту закономерность и не связано с гипотезой стабильность — время. В самом деле, 99 % изменчивости видового богатства, обнаруженной в этом исследовании, можно отнести за счет зависимости число видов — площадь [232].[ ...]
И генетический, и демографический анализ заставляют предположить, что отдаленное будущее существующей популяции мангобеев довольно мрачно. Из-за ограниченности территории и местообитаний вида и растущем народонаселении в этой области, увеличение размера популяции до 5-8 тыс. особей кажется нереальным. Увеличить вероятность выживания маншбея сможет менеджмент-план, в котором будут предусмотрены увеличение площади охраняемых лесов, высаживание в них растений, служащих для мангобеев источником корма, и создание коридоров, облегчающих их передвижение между фрагментами леса.[ ...]
Второе обобщение дополняет первое и носит название теории биоценотической регуляции численности популяции К. Фри-дерикса (1927): регуляция численности популяции есть результат комплекса воздействий абиотической и биотической среды в местообитании вида.[ ...]
Состав травянистого яруса изменяется под влиянием факторов внешней среды, таких как нарушение минерального питания, механические повреждения корней, надземной части, изменение фитоценотической обстановки (Бельков, Берг, Стародубова, 1993), что проявляется в снижении видового разнообразия и численности лесных видов и увеличении численности луговых и сорных растений. В ходе геоботанических описаний, проведенных в широколиственных насаждениях Москвы в 2003 г., было выявлено, что 44 % встречающихся в данных местообитаниях видов травянистых растений являются сорными или луговыми. Такая смена вызвана экологическими причинами, в ряду которых не последнее место занимает социальный фактор - рекреация. В результате вытаптывания и обрывания наиболее красивых и ценных особей в букеты, растения лишаются возможности семенного возобновления. В результате продолжение рода осуществляют более слабые и наименее декоративные растения, что в дальнейшем ведет к их полному исчезновению.[ ...]
Биотоп (био... и от греч. — место)— относительно однородное по абиотическим факторам жизненное пространство, занятое одним биоценозом. Он включает в себя минеральные и органические вещества, климатические факторы (свет, температура, влажность, pH среды и др.), физико-химические свойства различных субстратов (почва, дно водоема). Близок к понятиям «эко-топ» и «местообитание вида». Между биоценозом и биотопом, вместе образующих биогеоценоз, существует тесное взаимодействие, основанное на постоянном обмене веществом и энергией.[ ...]
Программа ООН по окружающей среде (ЮНЕП) выделила четыре важные причины потери биоразнообразия, связанные с хозяйственной деятельностью человека. Это, во-первых, нынешняя неэффективная структура землепользования: примерно 1-2% поверхности суши подвержено урбанизации и изъято из природной активности, около 25% - занято сельскохозяйственной деятельностью. В результате примерно 40% первичной продукции биоты Земли изъято деятельностью человека. Эти экономические процессы связаны как с ростом народонаселения (и нагрузки на природу), так и с формами собственности на землю и другие природные ресурсы. Вырубка тропических лесов в 80-е годы XX веке в Южной Америке, Африке и Юго-Восточной Азии способствовала снижению биоразнообразия. Это приводит к ускоренному исчезновению биологических видов темпами, которые уже сейчас в 50-100 раз превышают природные фоновые процессы и могут в ближайшее время достигнуть 10 000 раз. Вызывает озабоченность наблюдаемая в США тенденция к малым земельным участкам, что расчленяет и фрагментирует ландшафт, вызывает проблемы с местообитанием видов флоры и фауны. Такая же фрагментация происходит в Южной Америке.[ ...]
ru-ecology.info
Местообитание и экологическая ниша — МегаЛекции
Именно в процессе накопления изменений в генетическом аппарате, а также адаптации к определенному месту в окружающей среде происходит видообразование. То есть каждый вид или его части (популяции, группировки различного ранга) занимают определенное место в окружающей их среде. Например, определенный вид животного не может произвольно менять пищевой рацион или время питания, место размножения, убежища и т. п. Для растений подобная обусловленность условий выражается, например, через светолюбие или тенелюбие, место в вертикальном расчленении сообщества (приуроченность к определенному ярусу), время наиболее активной вегетации. Например, под пологом леса одни растения успевают закончить основной жизненный цикл, завершающийся созреванием семян, до распускания листьев древесного полога (весенние эфемеры). В более позднее время их место занимают другие, более теневыносливые растения. Особая группа растений способна на быстрый захват свободного пространства (растения-пионеры), но отличается низкой конкурентной способностью и поэтому быстро уступает свое место другим (более конкурентоспособным) видам.
Приведенные примеры иллюстрируют экологическую нишу или отдельные ее элементы. Под экологической нишей понимают обычно место организма в природе и весь образ его жизнедеятельности, или, как говорят, жизненный статус, включающий отношение к факторам среды,видам пищи, времени и способам питания, местам размножения, укрытий и т. п. Это понятие значительно объемнее и содержательнее понятия «местообитание». Американский эколог Одум образно назвал местообитание «адресом» организма (вида), а экологическую нишу — его «профессией». На одном местообитании живет, как правило, большое количество организмов разных видов. Например, смешанный лес - это местообитание для сотен видов растений и животных, но у каждого из них своя и только одна «профессия» — экологическая ниша. Так, сходное местообитание, как отмечалось выше, в лесу занимают лось и белка. Но ниши их совершенно разные: белка живет в основном в кронах деревьев, питается семенами и плодами, там же размножается и т. п. Весь жизненный цикл лося связан с подпологовым пространством: питание зелеными растениями или их частями, размножение и укрытие в зарослях и т. п.
Биологический вид — это совокупность особей, обладающих наследственным сходством морфологических, физиологических и биохимических особенностей, способных к скрещиванию с образованием плодовитого потомства, приспособленных к определенным условиям жизни и занимающих в природе определенную область (ареал).
В зависимости от величины ареала и характера распространения различают космополитов, убиквистов, эндемиков. Космополиты — виды растений и животных, представители которых встречаются на большей части обитаемых областей Земли (например, комнатная муха, серая крыса). Убиквисты — виды растений и животных с широкой экологической валентностью, способны существовать в разнообразных условиях среды, имеют обширные ареалы (например, тростник обыкновенный, волк). Эндемики— виды растений и животных, которые имеют небольшие ограниченные ареалы (часто встречаются на островах океанического происхождения, в горных районах и изолированных водоемах).
Для животных также различают трофическийи репродуктивный ареалы, между которыми существует связь в виде путей пролета для птиц или путей миграции для некоторых млекопитающих и рыб.
7.3 Принцип конкурентного исключения (принцип Гаузе)
Если организмы занимают разные экологические ниши, они не вступают обычно в конкурентные отношения, сферы их деятельности и влияния разделены. В таком случае отношения рассматриваются как нейтральные.
Вместе с тем, в каждой экосистеме имеются виды, которые претендуют на одну и ту же нишу или ее элементы (пищу, укрытия и пр.). В таком случае неизбежна конкуренция, борьба за обладание нишей. Эволюционно взаимоотношения сложились так, что виды со сходными требованиями к среде не могут длительно существовать совместно. Эта закономерность не без исключений, но она настолько объективна, что сформулирована в виде положения, которое получило название «правило конкурентного исключения». Автор этого правила эколог Г. Ф. Гаузе. Звучит оно так: если два вида со сходными требованиями к среде (питанию, поведению, местам размножения и т. п.) вступают в конкурентные отношения, то один из них должен погибнуть либо изменить свой образ жизни и занять новую экологическую нишу. Иногда, например, чтобы снять острые конкурентные отношения, одному организму (животному) достаточно изменить время питания, не меняя самого вида пищи (если конкуренция возникает на почве пищевых отношений), или найти новое местообитание (если конкуренция имеет место на почве данного фактора) и т. п.
Из других свойств экологических ниш отметим, что вид может их менять на протяжении своего жизненного цикла. Наиболее яркий пример в этом отношении - насекомые. Так, экологическая ниша личинок майского жука связана с почвой, питанием корневыми системами растений. В то же время экологическая ниша жуков связана с наземной средой, питанием зелеными частями растений.
Сообщества (биоценозы, экосистемы) формируются по принципу заполнения экологических ниш. В природном сформировавшемся сообществе обычно все ниши заняты. Именно в такие сообщества, например, в долгосуществующие (коренные) леса, вероятность внедрения новых видов очень мала. В то же время следует иметь в виду, что занятость экологических ниш в определенной мере понятие относительное. Все ниши обычно освоены теми организмами, которые характерны для данного региона. Но, если организм приходит извне (например, заносятся семена или другие зачатки) случайно или преднамеренно, например, в результате внедрения человеком новых видов (интродукция, акклиматизация), то он может найти для себя свободную нишу в связи с тем, что на нее не было претендентов из набора существующих видов. В таком случае обычно неизбежно быстрое увеличение численности (вспышка) вида-пришельца, поскольку он находит крайне благоприятные условия (свободную нишу) и, в частности, не имеет врагов (хищников, паразитов или других организмов, которые им питаются). Такие явления не единичны. Например, размножение кроликов, завезенных в Австралию; перемещение ондатры из Азии в европейскую часть; интенсивное продвижение колорадского жука в новые районы.
С экологическими нишами в значительной мере связаны жизненные формы организмов. К последним относят группы видов, часто систематически далеко отстоящие, но выработавшие одинаковые морфологические адаптации в результате существования в сходных условиях. Например, сходством жизненных форм характеризуются дельфины (млекопитающие) и интенсивно передвигающиеся в водной среде хищные рыбы. В условиях степей сходными жизненными формами представлены тушканчики и кенгуру (прыгуны). В растительном мире отдельными жизненными формами представлены многочисленные виды деревьев, занимающие в качестве ниши верхний ярус, кустарники, существующие под пологом леса, и травы — в напочвенном покрове.
Любой вид занимает определенное местообитание и определенную экологическую нишу.
Местообитание — это территория или акватория, занимаемая видом, с комплексом присущих ей экологических факторов. Местообитание вида является компонентом его экологической ниши. Применительно к наземным животным местообитание вида называется стация, местообитание сообщества — биотоп.
Экологическая ниша — совокупность всех факторов среды, в пределах которых возможно существование вида в природе. То есть экологическая ниша — это место вида в природе, включающее не только его положение в пространстве и отношение к абиотическим горизонтом, пологом, слоем, ярусом биогеоценоза. Например, в сосновом лесу можно выделить синузию сосны, синузию брусники, синузию зеленых мхов и т.д факторам, но и его функциональную роль в сообществе (прежде всего трофический статус). Местообитание — это как бы «адрес» вида, а экологическая ниша — это его «профессия».
Для характеристики экологической ниши вида обычно используют два важных показателя: ширина ниши и степень перекрывания ее с соседними. Экологические ниши разных видов могут быть разной ширины и перекрываться в различной степени.
Разделение экологических ниш между видами происходит за счет приуроченности разных видов к разным местообитаниям, разной пищи и разному времени использования одного и того же местообитания. Принцип конкурентного исключения (принцип Гаузе) гласит: «Два вида не могут сосуществовать в одной и той же местности, если их экологические потребности идентичны. Такие виды обязательно должны быть разобщены в пространстве или во времени».
Группы видов в сообществе, обладающие сходными функциями и нишами одинакового размера, т.е. роль которых в сообществе одинакова или сравнима, называются гильдиями. Например, лианы тропического леса представлены многими видами растений. Между видами внутри гильдии наблюдается особенно острая конкуренция.
Виды, занимающие одинаковые ниши в разных географических областях, называются экологическими эквивалентами. Например, крупные кенгуру Австралии, бизоны Северной Америки, зебры и антилопы Африки и т.д. являются экологическими эквивалентами. В настоящее время они значительно замещены коровами и овцами.
megalektsii.ru
Фитоценоз - это... Что такое Фитоценоз?

 Лесной фитоценоз
Лесной фитоценоз Фитоцено́з (от греч. φυτóν — «растение» и κοινός — «общий») — растительное сообщество, существующее в пределах одного биотопа. Характеризуется относительной однородностью видового состава, определённой структурой и системой взаимоотношений растений друг с другом и со внешней средой. По Н. Баркману[1] фитоценоз — суть конкретный сегмент растительности, в котором внутренние флористические различия меньше, чем различия с окружающей растительностью. Термин предложен польским ботаником И. К. Пачоским в 1915 году. Фитоценозы является объектом изучения науки фитоценологии (геоботаники).
Фитоценоз входит в состав биоценоза наряду с зооценозом и микробиоценозом.
Биоценоз, в свою очередь, в сочетании с условиями абиотический среды (экотопом) образует биогеоценоз. Фитоценоз является центральным, ведущим элементом биогеоценоза, так как трансформирует первичный экотоп в биотоп, создавая среду обитания для других организмов, а также является первым звеном в круговороте веществ и энергии. От растительности зависят свойства почв, микроклимат, состав животного мира, такие характеристики биогеоценоза, как биомасса, биопродуктивность и т. д. В свою очередь, элементами фитоценоза являются ценопопуляции растений — совокупности особей одного вида в границах фитоценозов.Развитие взглядов на природу фитоценоза
На заре развития геоботаники оформилось представление о фитоценозе как реально существующей дискретной единице растительного покрова, что на тот момент представлялось вполне целесообразным, так как выделение отдельных фитоценозов существенно облегчало задачу изучения растительности в целом. Однако в начале XX века была высказана[2][3][4][5] диаметрально противоположная точка зрения, согласно которой растительный покров представлялся непрерывным, а его подразделение на отдельные элементы — фитоценозы — искусственным. Отсутствие резких границ между растительными сообществами и наличие переходных зон между ними способствовало возникновению учения о непрерывности (континуальности) растительного покрова, основанного на индивидуалистической концепции:
- Каждый вид растений индивидуален по своим требованиям к условиям внешней среды и имеет характерные экологические амплитуды по каждому экологическому фактору
- Факторы среды изменяются постепенно, как в пространстве, так и во времени
- Переход от одной комбинации ценопопуляций к другой осуществляется непрерывно: одни виды постепенно уменьшают своё обилие и исчезают, другие — появляются и увеличивают.
Крайними сторонниками концепции континуальности растительного покрова в качестве объекта изучения геоботаники рассматривался не фитоценоз, в силу его искусственности, а отдельное растение. Крайние сторонники идеи дискретности же постулировали чёткое различие и разграничение отдельных фитоценозов.
На основе синтеза обеих концепций была выдвинута[6][7] идея о сочетании в природе растительного покрова и дискретности, и непрерывности. Это представлялось как одно из проявлений противоречивости, свойственной для материального мира в целом. Согласно этой идеи, растительный покров обладает свойством непрерывности, но она не абсолютна, а относительна. В то же время он обладает и свойством дискретности, но и она не абсолютна, а относительна. Эти свойства органично сочетаются, не исключая, а дополняя друг друга.
Формирование фитоценоза
 Первично свободный участок земной поверхности после извержения вулкана
Первично свободный участок земной поверхности после извержения вулкана Формирование фитоценозов можно рассматривать как в динамическом аспекте (смена сообществ), так и в плане образования их на свободных участках земной поверхности.
Различают первично свободные участки, которые в прошлом не заселялись растениями и не содержат их зачатков. Фитоценозы на них могут образоваться только при внесении диаспор извне. К таким участкам относятся скальные обнажения, свежие речные и морские наносы, обнажившееся дно водоёмов, участки, освободившиеся от ледников, лавовые поля и др. В целом на Земле они занимают незначительные площади.
Вторично свободные участки образуются в местах, где раньше растительность существовала, но была уничтожена вследствие влияния какого-либо неблагоприятного фактора. Примером могут служить гари, осыпи, незасеянные пашни, участки выеденных вредителями или скотом фитоценозов. На них в большинстве случаев сохраняется почва и диаспоры, а формирование фитоценозов происходит значительно быстрее, чем на первично свободных участках.
Формирование фитоценоза является непрерывным процессом, но условно может быть разделено на стадии:
- Отсутствие фитоценоза — случайный состав видов; отсутствие взаимодействия между растениями; очень слабое влияние на среду; невыраженность структуры.
- Открытый фитоценоз — неустойчивый состав, преимущественно из однолетников; структура с отдельными не взаимодействующими друг с другом ценопопуляциями.
 Сгоревший лес — вторично свободный участок
Сгоревший лес — вторично свободный участок - Закрытый невыработавшийся фитоценоз — выпадение значительной части видов-первопоселенцев; пятнистая структура с проникновением отдельных растений в скопления других видов; намечается ярусность.
- Закрытый выработавшийся фитоценоз — относительно постоянный видовой состав; затруднение проникновения новых видов; взаимодействие всех ценопопуляций; выраженная ярусность.
- Пионерная группировка — ценопопуляции малочисленны, взаимоотношения между ними отсутствуют
- Группово-зарослевое сообщество — ценопопуляции распределены куртинами, в которых происходит взаимодействие между растениями
- Диффузное сообщество — ценопопуляции смешиваются, вырабатывается система межвидовых взаимодействий
- Миграция — занос диаспор
- Эцезис — закрепление первопоселенцев
- Агрегация — образование групп потомства вокруг материнских растений
- Инвазия — перемешивание ценопопуляций
- Конкуренция — выработка конкурентных отношений вследствие резкого увеличения сомкнутости
- Стабилизация — формирование устойчивого замкнутого сообщества
Е. П. Прокопьев, суммируя различные схемы членения процесса формирования фитоценоза, предлагает выделять в нём три этапа[11]:
- Поступление зачатков на свободный участок. Видовой состав формирующегося фитоценоза будет зависеть от видового состава растений окружающей территории и характера распространения их диаспор, причём основную роль будут играть зачатки аллохорических видов, главным образом анемохоры.
- Экотопический (абиотический) отбор. Не все попавшие на свободный участок диаспоры на нём приживутся: часть не прорастёт, а часть проросших погибнет в молодом состоянии вследствие неблагоприятного сочетания абиотических факторов. Закрепившиеся растения будут являться пионерными для данной территории.
- Фитоценотический отбор. Вследствие размножения и расселения пионерных видов по участку, они начнут влиять друг на друга и изменять экотоп, формируя биотоп (местообитание). Первичная абиотическая среда экотопа превращается во вторичную биотическую — фитосреду. Под влиянием фитосреды и взаимовлияний растений некоторые пионерные виды, не приспособленные к ней, выпадают. Это может происходить, например, вследствие затенения или аллелопатии. Параллельно происходит закрепление на участке новых видов, уже приспособленных к данной фитосреде.
Факторы организации фитоценоза
Факторы организации растительного сообщества можно условно разделить на четыре группы: характеристики среды (экотопа), взаимоотношение между растениями, влияние на растительность гетеротрофных компонентов (животных, грибов, бактерий) и нарушения. Эти группы факторов определяют сочетание и характеристики ценопопуляций видов в фитоценозе.
Экотоп является главным фактором организации фитоценоза, хотя он может быть в значительной степени трансформирован биотическими влияниями растений или нарушениями. К абиотическим факторам, влияющим на организацию сообщества можно отнести:
Взаимоотношения растений подразделяются на контактные и опосредованные: трансабиотические — через абиотические факторы среды обитания и трансбиотические — через третьи организмы.
Влияние на организацию фитоценозов гетеротрофных компонентов биогеоценозов исключительно разнообразно. Влияние животных проявляется в опылении, поедании, распространении семян, изменении стволов и крон деревьев и связанных с ними характеристик, разрыхлении почвы, вытаптывании и др. Микоризные грибы улучшают снабжение растений элементами минерального питания и водой, повышают устойчивость к патогенам. Бактерии-азотфиксаторы повышают снабжение растений азотом. Другие грибы и бактерии, а также вирусы могут являться патогенами.
Нарушения, как антропогенного, так и природного генезиса могут полностью трансформировать фитоценоз. Это происходит при пожарах, вырубках, выпасе скота, рекреационной нагрузке и т. п. В этих случаях формируются производные фитоценозы, которые постепенно изменяются в сторону восстановления коренного, если воздействие нарушающего агента прекратилось. Если воздействие долговременно (например, при рекреации) формируются сообщества, приспособленные к существованию при данном уровне нагрузки. Деятельность человека привела к образованию фитоценозов, ранее не существовавших в природе (например, сообществ на токсичных отвалах промышленных производств).
Взаимовлияния организмов в фитоценозах
Наличие системы взаимоотношений между растениями является одним из главных признаков сложившегося фитоценоза. Изучение их, в силу большой перекрываемости и сильного влияния абиотических факторов, представляет собой трудную задачу и может быть реализовано либо в виде эксперимента, в ходе которого изучаются взаимоотношения двух конкретных видов, либо вычленением таких отношений из комплекса других с применением методов математического анализа.
Прямые (контактные) взаимовлияния

Возникают при соприкосновении или проникновении организмов друг в друга. Подразделяются на физиологические (паразитизм и симбиоз), когда между организмами осуществляется активный обмен веществом и энергией, и механические (взаимоотношения эпифитов с форофитами и лиан с опорными растениями) — когда таковой отсутствует.
При паразитизме один организм (паразит) использует другой (хозяина) для получения необходимых ему веществ и энергии, при этом его угнетая. Среди высших растений паразитизм встречается только у покрытосеменных (см. растения-паразиты). Также на всех растениях способны поселяться паразитические грибы и бактерии. Степень поражения растения паразитами зависит от особенностей растения-хозяина (разные виды поражаются неодинаково), от условий местообитания (в условиях засоления растения практически не поражаются), от наличия эволюционной сопряжённости паразита и хозяина (если её нет, у хозяина обычно отсутствуют механизмы защиты от паразита).
Против паразитических грибов и бактерий у растений в ходе эволюции сформировался комплекс защитных механизмов:
- выделение растением фунгицидных и бактерицидных веществ, предотвращающих заражение паразитами или подавляющих их развитие
- наличие мощных покровных тканей, препятствующих проникновению паразитов
- особенности биохимического состава и метаболизма в клетках растения, препятствующие росту паразитов
Несмотря на наличие подобных механизмов защиты, фитопаразиты способны вызывать многочисленные болезни растений, ведущие к их ослаблению и гибели. Растения-паразиты распространены не столь широко, но также могут значительно угнетать ценопопуляции и отдельные растения.

Симбиотические отношения проявляются в сосуществовании растений с грибами и бактериями (в том числе цианобактериями). Соответственно различают микосимбиотрофию и бактериосимбиотрофию.
Микосимбиотрофия реализуется в виде микоризы — взаимодействия гиф грибов и корней растений. Предполагается[11], что на ранних этапах эволюции растительного мира грибы выступали по отношению к растениям только как паразиты и лишь в процессе длительной коэволюции сформировались взаимовыгодные отношения. К настоящему времени микориза обнаружена более чем у 80 % видов сосудистых растений: её образуют все виды голосеменных, 77—78 % видов покрытосеменных и около 60 % видов сосудистых споровых растений[12]. Различают эктомикоризу, когда мицелий опутывает корни растения и эндомикоризу, когда проникает в них.
Экологический смысл образования микоризы состоит в том, что гриб получает от растения углеводы и некоторые витамины, а растение следующие выгоды:
- мицелий гриба увеличивает всасывающую поверхность корней растения, улучшая снабжение его водой и минеральными веществами
- гриб может выделять некоторые витамины и ростовые вещества
- разлагая недоступные растениям органические вещества, грибы переводят их в доступную усвояемую форму
- заражение корней микоризными грибами предохраняет растение от воздействия патогенных микроорганизмов, в том числе паразитических грибов
- при эндомикоризе часть грибных клеток в корнях растений разлагается и используется в качестве источника питательных веществ
Растения, образующие микоризу, по требовательности к наличию микосимбионта можно разделить на две группы:
- облигатные микосимбиотрофы — не способные к развитию без микосибионта (сем. Орхидные)
- факультативные микосимбиотрофы — способные к существованию без микосимбионта, но лучше развивающиеся при его наличии
Бактериосимбиотрофия — симбиоз растений с клубеньковыми бактериями (Rhizobium sp.). Распространена не столь широко, как микосимбиотрофия — в симбиоз с бактериями вступает порядка 3 % растений мировой флоры (в основном семейства Бобовые (около 86 % видов семейства), а также некоторые виды семейств Мятликовые, Берёзовые, Лоховые, Крушиновые)[12][13]. Клубеньковые бактерии играют роль азотфиксаторов, переводя атмосферный азот в доступные растениям формы. Выделяют корневую и листовую формы взаимодействия. При корневой форме бактерии заражают корни растения, вызывая интенсивные локальные деления клеток и образование клубеньков. Листовая бактериосимбиотрофия встречается у некоторых тропических растений и ещё слабо изучена.
На способности растетий семейства Бобовых вступать в симбиоз с клубеньковыми бактериями основан один из методов трёхпольной культуры.
 Тропический эпифитный папоротник Олений рог (Platycerium bifurcatum)
Тропический эпифитный папоротник Олений рог (Platycerium bifurcatum) Эпифиты, поселяющиеся на растениях-форофитах используют последние только в качестве субстрата, не вступая с ними в физиологические взаимодействия. Эпифитные формы встречаются в группах покрытосеменных, папоротников, мхов, водорослей и лишайников. Наибольшего разнообразия эпифиты достигают во влажных тропических лесах.
В экологическом плане взаимоотношения эпифитов и форофитов обычно представлены комменсализмом, но могут проявляться и элементы конкуренции:
- эпифиты частично перехватывают свет и влагу у форофитов
- задерживая влагу, способствуют загниванию форофита
- затеняя форофит, эпифиты уменьшают его эффективную фотосинтетическую поверхность
- обильно разрастаясь, могут быть причиной деформации или слома форофитов
Лианы — растения, использующие другие растения или иные объекты для сохранения вертикального положения. Лианы появились в ходе эволюции как одно из проявлений конкуренции за свет. Подавляющее число видов лиан относится к покрытосеменным растениям. Большинство лиан укореняется в почве, однако существуют эпифитные и паразитные формы, например, растения рода Повилика (Cuscuta) сем. Вьюнковые (Convolvulaceae).
Лианы, вынося в ходе роста свои листья ближе к свету, получают от сожительства с опорным растениием пользу, тогда как последнее — преимущественно вред, как прямой — вследствие механического воздействия лианы и слома / гибели опорного растения[14], так и косвенный — вследствие перехвата лианой света, влаги и питательных веществ.
Наибольшего разнообразия лианы также достигают во влажных тропических лесах.
Трансабиотические взаимовлияния
Влияния растений друг на друга, опосредованные абиотическими факторами внешней среды. Возникают благодаря перекрыванию фитогенных полей расположенных по соседству растений. Подразделяются на конкуренцию и аллелопатию.
Конкуренция развивается либо из-за исходной ограниченности ресурсов местообитания, либо в результате уменьшения их доли, приходящейся на каждое растение, вследствие перенаселённости. Конкуренция ведёт к снижению потребления ресурсов растением и, как следствие, уменьшению скорости роста и запасания веществ, а это, в свою очередь, приводит к снижению количества и качества диаспор. Различают внутри- и межвидовую конкуренцию.
Внутривидовая конкуренция влияет на показатели рождаемости и смертности в ценопопуляции, определяя тенденцию к поддержанию её численности на определённом уровне, когда обе величины уравновешивают друг друга. Такая численность называется предельной плотностью и зависит от количества ресурсов местообитания. Внутривидовая конкуренция асимметрична — неодинаково влияет на разные особи. Суммарная фитомасса ценопопуляции остаётся постоянной в достаточно большом диапазоне значений плотности, тогда как средняя масса одного растения при загущении начинает неуклонно снижаться — закон постоянства урожая (C=dw, где C — урожай, d — плотность ценопопуляции и w — средняя масса одного растения).
Межвидовая конкуренция также широко распространена в природе, поскольку абсолютное большинство фитоценозов (кроме некоторых агроценозов) являются многовидовыми. Многовидовой состав обеспечивается тем, что каждый вид имеет характерную только для него экологическую нишу, которую и занимает в сообществе. При этом ниша, которую вид мог бы занять при отсутствии межвидовой конкуренции — фундаментальная, сужается до размеров реализованной. В фитоценозе дифференциация экологических ниш происходит за счёт:
- разной высоты растений
- разной глубины проникновения корневой системы
- контагиозного распределения особей популяции (отдельными группами / пятнами)
- разных сроков вегетации, цветения и плодоношения
- неравной эффективности использования растениями ресурсов местообитания
При слабом перекрывании экологических ниш может наблюдаться сосуществование двух ценопопуляций, при сильном же более конкурентноспособный вид вытесняет из местообитания менее конкурентноспособный. Сосуществование двух сильно конкурирующих видов также возможно за счёт динамичности среды, когда тот или иной вид получает временное преимущество.
Аллелопатия — влияние растений друг на друга и на другие организмы путём выделения в среду активных метаболитов как при жизни растения, так и при разложении его остатков. Аллелопатическая активность того или иного вида обуславливается определённым набором химических веществ различной природы, качественный и количественный состав которого существенно зависит от внешних условий. Аллелопатически активные вещества выделяются как надземными органами (главным образом листьями), так и подземными в основном тремя способами:
- активное выделение через железы или гидатоды
- вымывание атмосферными осадками
- выделение посредством разложения опада микроорганизмами
Сумма выделений различных растений в фитоценозе — его биохимическая среда. Так как состав выделений не постоянен, можно говорить о существовании аллелопатического режима фитоценоза, наряду с водным, воздушным и др.
Трансбиотические взаимовлияния
Влияние фитоценоза на среду
Структура фитоценоза
В зависимости от специфики исследований в понятии «структуре биоценоза» В. В. Мазингом[15] выделяются три направления, разработанные им для фитоценозов.
1. Структура, как синоним состава (видовая, конституционная). В этом смысле говорят о видовой, популяционной, биоморфологической (состав жизненных форм) и других структурах ценоза, имея ввиду лишь одну сторону ценоза — состав в широком смысле. В каждом случае проводится качественный и количественный анализ состава.
2. Структура, как синоним строения (пространственная, или морфоструктура). В любом фитоценозе растения характеризуются определенной приуроченностью к экологическим нишам и занимают определенное пространство. Это относится и к остальным компонентам биогеоценоза. Между частями пространственного расчленения (ярусы, синузии, микрогруппировки и др.) можно достаточно легко и точно провести границы, можно нанести их на план, вычислить площадь, а затем, например, рассчитать ресурсы полезных растений или кормовые ресурсы животных. Только на основе данных по морфоструктуре можно объективно определить точки постановки тех или иных экспериментов. При описании и диагностики сообществ всегда проводится изучение пространственной неоднородности ценозов.
3. Структура, как синоним совокупностей связей между элементами (функциональная). В основе понимания структуры в таком смысле лежит изучение взаимоотношений между видами, в первую очередь изучение прямых связей — биотический коннекс. Это изучение цепей и циклов питания, обеспечивающих круговорот веществ и раскрывающих механизм связей трофических (между животными и растениями) или топических (между растениями — конкуренция за питательные вещества в почве, за свет в надземной сфере, взаимопомощь).
Все три аспекта структуры биологических систем тесно взаимосвязаны на ценотическом уровне: видовой состав, конфигурация и размещение структурных элементов в пространстве являются условием для их функционирования, то есть жизнедеятельности и продуцирования растительной массы, а последнее, в свою очередь, в значительной степени определяет морфологию ценозов. И все указанные аспекты отражает условия среды, в которых формируется биогеоценоз.
Фитоценоз состоит из ряда структурных элементов. Различают горизонтальную и вертикальную структуру фитоценоза. Вертикальная структура представлена ярусами, выделяемыми по визуально определяемым горизонтам концентрации фитомассы. Ярусы состоят из «разновысотных» растений. Примерами ярусов являются 1-й древесный ярус, 2-й древесный ярус, напочвенный покров, мохово-лишайниковый ярус, ярус подлеска и др. Число ярусов может различаться. Эволюция фитоценозов идёт в направлении увеличения числа ярусов, так как это ведёт к ослаблению конкуренции между видами. Поэтому в более древних лесах умеренного пояса Северной Америки число ярусов (8—12) больше, чем в аналогичных более молодых лесах Евразии (4—8).
Горизонтальная структура фитоценоза формируется благодаря наличию пологов деревьев (под которыми формируется среда, несколько отличная от среды в межпологовом пространстве), неоднородностям рельефа (которые обуславливают изменение уровня грунтовых вод, различную экспозицию), видовым особенностям некоторых растений (размножающихся вегетативно и образующих моновидовые «пятна», изменение среды одним видов и реагирование на это другими видами, аллелопатические воздействия на окружающие растения), деятельности животных (например, формирование пятен рудеральной растительности на пороях грызунов).
Закономерно повторяющиеся пятна (мозаики) в фитоценозе, различающиеся составом видов или их количественным соотношением, называются микрогруппировками[16], а такой фитоценоз — мозаичным.
Неоднородность может также носить случайный характер. В этом случае она называется пестротностью.
Динамика фитоценозов
динамические процессы: обратимые(суточные, сезонные, флуктуации) и необратимые(сукцессии, эволюция сообществ, нарушения сообществ). Флуктуации-это разногодичные изменения, связанные с неодинаковыми условиями существования растений в разные годы. состав не изменяется, может менятся численность и возрастной состав популяции. Сукцессии-это постепенные изменения фитоценозов необратимы и направлены. вызванные внутренними или внешними причинами, по отношению к фитоценозам, причинам. выделяются первичные и вторичные сукцессии. Первичные сукцессии начинаются на безжизненных субстратах, вторичные же начинаются на субстратах на которых растительность была но нарушена.
Классификация фитоценозов
При классификации фитоценозов сходные сообщества объединяют в группы — классификационные единицы.
Низшая единица классификации — ассоциация (совокупность однородных фитоценозов, имеющих более или менее одинаковый внешний облик, сходный флористический состав и одни и те же доминирующие виды по ярусам). Названия ассоциаций даются перечислением русских названий доминирующих растений каждого яруса фитоценоза начиная с самого верхнего яруса (сосна обыкновенная + ель европейская — брусника + черника — мох плеуроциум) или латинских родовых и видовых названий доминантов (Pinus sylvestris + Picea abies — Vaccinium vitis-idaea + Vaccinium myrtillus — Pleurozium schreberi) с прибавлением к основе суффиксов лат. -etum, -osum, -estosum: Piceetum oxalidosum (от Picea и Oxalis) — ельник кисличный.
Формация — это совокупность ассоциаций, у которых в верхнем ярусе доминирует один и тот же вид растения (например, сосняки, дубравы и пр.)
Ординацией называют построение рядов фитоценозов по постепенному изменению какого-либо фактора окружающей среды в определенном направлении. Так, можно провести ординацию по фактору увлажнения почвы. В этом случае получится ряд сообществ, где каждое займет соответствующее место в зависимости от условий увлажнения, в которых оно развивается, причём крайнее из них будет соответствовать наиболее влажным почвам, а противоположное — наиболее сухим.
Эдафо-фитоценотические ряды типов еловых лесов (по В. Н. Сукачеву, 1972):
1) Ельники-зеленомошники. Встречаются в условиях расчленённого рельефа, на более или менее богатых суглинках, а также глинах и супесях. К этой группе относят ельники кисличные, черничные и брусничные.
2) Ельники-долгомошники (в моховом ярусе господствует кукушкин лён). Встречаются в условиях более или менее равнинного рельефа на слабо дренированных почвах с признаками заболачивания (состав почв тот же, что и в ельниках первой группы).
3) Ельники сфагновые. Встречаются в условиях равнинного рельефа, нередко в котловинных понижениях, на недренированных заболоченных почвах. Моховой покров состоит в основном из сфагновых мхов.
4) Ельники травяные. Располагаются по дну долин небольших речек и ручьёв в виде узких полос на богатых сырых почвах с проточным увлажнением. Хорошо развит травяной покров.
5) Ельники сложные. Приурочены к богатым, хорошо дренированным почвам, часто с близким залеганием известняков. В этих лесах хорошо развиты подлесок и травяной покров, присутствуют широколиственные породы — дуб, клен, липа.
Эти пять групп ассоциаций образуют экологические ряды, отражающие изменения еловых лесов в зависимости от увлажнения и богатства почвы. Ряд А (от кисличного к брусничному ельнику) — увеличение сухости почвы и некоторое обеднение почвы. Ряд В (от кисличного к сфагновому ельнику) — увеличение увлажнения и ухудшения аэрации почвы. Ряд С (от кисличного к сложному ельнику) — увеличение богатства и аэрации почвы. Ряд Д (от кисличного к приручьевому ельнику) — увеличение проточного увлажнения. Ряд Е (от кисличного к сфагновому ельнику) — увеличение застойного увлажнения и уменьшение аэрации почвы.
Примечания
- ↑ Баркман Н. Современные представления о непрерывности и дискретности растительного покрова и природе растительных сообществ в фитосоциологической школе Браун-Бланке. — Ботанический журнал, 1989, т. 74 № 11.
- ↑ Раменский Л. Г. О сравнительном методе экологического изучения растительных сообществ // Дневник съезда русских естествоиспытателей и врачей — СПб., 1910.
- ↑ Greason H. A. The structure and development of the plant assotiation // Bul. Torrey Bot. Club — 1917.
- ↑ Negri G. Le unita ecologiche fundamentali in fitogeografia — Roy. Acad Sc. Torino, 1914.
- ↑ Lenoble F. A propos des associations vegetales // Bull. Soc. Bot. France — 1926.
- ↑ Александрова В. Д. О единстве непрерывности и дискретности в растительном покрове // Философские проблемы современной биологии. — М.-Л.: Наука, 1966.
- ↑ Александрова В. Д. Классификация растительности. — Л.: Наука, 1969.
- ↑ Сукачёв В. Н. Дендрология с основами лесной геоботаники. — Л.: Гослестехиздат, 1938.
- ↑ Шенников А. П. Введение в геоботанику. — Л.: Изд-во ЛГУ, 1964.
- ↑ Weaver J. E., Clements F. E. Plant ecology. — N.-Y.-London, 1938.
- ↑ 1 2 Прокопьев Е. П. Введение в геоботанику. Учебное пособие. — Томск: Изд-во ТГУ, 1997. — 284 с.
- ↑ 1 2 Работнов Т. А. Фитоценология. — М.: Изд-во МГУ, 1978.
- ↑ Работнов Т. А. История фитоценологии. — М.: Аргус, 1995.
- ↑ Тихнова Г. Н. Фикусы-удушители с острова Борнео
- ↑ Мазинг В. В. Что такое структура биогеоценоза // Проблемы биогеоценологии. — М.: Наука, 1973. — С. 148–156.
- ↑ Ярошенко П. Д. Геоботаника: Пособие для студентов педвузов. — М: Просвещение, 1969. — 200 с.
Ссылки
dic.academic.ru
Местообитание как экологическое понятие
Геолого-исторические факторы за миллионы лет образовали материк с крупным расчленением его поверхности (хребты, низменности и пр.).
Параллельно развились флора и фауна каждого района, сложились его климатические и гидрологические условия.
Процессы эрозии детально расчленили рельеф местности, покрыли площадь пестрым плащом чередующихся материнских рыхлых пород, создали мозаику различных местоположений. В условиях определенного климата каждое местоположение стало типом местообитания. По местообитаниям распределились организмы и уклады почвообразовательных процессов — сложился характерный ландшафт местности. Большое значение в выработке ландшафта имело и имеет вековое влияние человека (истребление лесов, распашка, выпас скота и пр.). В зависимости от наличного подбора зачатков возникают процессы взаимодействия организмов: это приводит к формированию определенной растительности, отвечающей условиям данной конкретной площади, а также к выработке всех современных особенностей почвы (включая ее подвижные изменчивые признаки).
Современные растительные группировки, а вместе с ними и почва во всех своих признаках (относительно устойчивых и изменчивых) являются непосредственным продуктом жизнедеятельности населяющих данную площадь организмов в их взаимодействии с внешними факторами среды. Состав и все особенности растительных группировок определены экологическими режимами занимаемой ими площади, их местообитанием, а также режимом воздействия на них человека. Однако обусловленность растительной группировки местообитанием (экологией) не прямая, а опосредствованная взаимодействием организмов.
В связи с этим обстоятельством к каждому типу местообитания приурочена целая группа экологически равноценных растительных группировок. Одни из растительных группировок при этом считаются первичными, чисто природными или коренными (девственная степь, слабо измененный выпасом луг, еловый лес — рамень, дубрава и т. п.), другие производными (степные залежи, сбитые пастбища, послелесные луга и пр.). Виды местообитаний обобщаются в более крупные единицы — типы и классы местообитаний (луга и болота разных градаций увлажнения и химизма почвы и пр.). Виды местообитаний систематизируются, располагаясь по градациям рядов увлажнения и его сезонной переменности, активного богатства почвы и пр.
Условия местообитания природной степи не такие, какие в степи, сильно измененной интенсивным выпасом;
определенные условия свойственны молодой, мягкой залежи, старой залежи. Но в ряде случаев воздействие растительности и биоценоза на условия среды настолько качественно своеобразно, глубоко и необратимо, что следует говорить уже о возникновении нового местообитания. Это наблюдается, например, в степи при заселении ее лесом или, наоборот, на послелесных лугах в условиях лесной зоны при сменах растительной формации.
Следует иметь в виду, что термины «местообитание» и «местоположение» мы не считаем синонимами.
Местообитание характеризуется определенными экологическими условиями, а местоположение представляет собой лишь пункт расположения на местности. В условиях одной зоны местоположение по рельефу во многих случаях будет определять собой и местообитание, но в разных зонах однохарактерное по рельефу местоположение будет иметь экологически разное местообитание. Наоборот, различные по рельефу местоположения могут создавать очень близкие местообитания и иметь сходную растительность; некоторые склоны в лесостепной зоне заняты в природных условиях группировками степных растений, южнее приуроченными к равнинам; в степной зоне в притеррасной части долин, равномерно увлажняемых близкими к поверхности грунтовыми водами, встречаются лужки со щучкой и красной овсяницей, растительность которых характерна для равнинных суходолов лесной зоны.
Пестрый ковер различных растительных группировок, связанных с определенными местообитаниями, не является случайным нагромождением их; они распределены строго закономерно, образуя ландшафт местности — результат ее долгого исторического развития. Определенный набор, а также расположение местообитаний и растительных группировок свойственен пойме, надлуговой террасе, водораздельным площадям, зандровым полям, конечным моренам и т. д. Выяснение истории развития страны, ее фауны, флоры, воздействия на нее культуры завершает всестороннее исследование географии страны, ее геоморфологии, почв, растительности — в целом ее ландшафта. Именно такое всестороннее и законченное исследование обеспечивает наибольшую полноту теоретических и практических выводов.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
тип местообитания
Тип охотничьего угодья равнозначен таким научным терминам, как тип местообитания, биотоп, стация вида и т. п. Его целесообразно использовать лишь применительно к каждому виду охотничьих зверей и птиц отдельно. Дело в том, что примирить несходные требования разных видов к среде обитания, да еще в разные сезоны года, крайне сложно, часто невозможно, а главное не вызывается практической необходимостью. Любое мероприятие в конкретном охотничьем хозяйстве решается всегда для определенного вида, а не вообще. Вместе с тем тип лесного охотничьего угодья—это очевидная, осязаемая реальность, только значение его часто разное и несходное.[ ...]
Местообитания можно классифицировать по влияниям «размера» на остаточную репродуктивную ценность, хотя такой способ дает только сравнительные результаты. — Различные типы местообитаний могут возникать по многим причинам.[ ...]
Из других типов местообитаний наибольшую площадь занимают леса - 32% (28 тыс. га).[ ...]
Из других типов местообитаний наибольшую площадь занимают леса - 32% (28 тыс. га).[ ...]
Хороший пример такого типа местообитания — пахотные земли, поскольку смертность взрослых особей на них неизбирательна. При ежегодной вспашке прижившиеся растения уничтожаются независимо от их размера. Характерная сорная флора состоит из скороспелых и моноцикличных однолетников (вместе с некоторыми полицикличными многолетниками, для которых вспашка не смертельна).[ ...]
Поэтому при интерпретации этих типов разнообразия необходимы осторожность и здравый смысл. Большое внимание следует уделять применяемым классификациям типов местообитаний и ресурсов, которые должны быть одинаковы в разных случаях, соблюдаясь во всех деталях. Очень важно, чтобы размеры выборок были постоянны и достаточно велики для адекватного выявления разнообразия.[ ...]
Сопоставление выделенных серий типов леса на интенсивно осушенных торфах с единицами типологических классификаций других авторов показало следующее: для таежной и подтаежной зоны Европы выделяется обычно от 2 до 5 типов леса (типов местообитаний, типов лесорастительных условий).[ ...]
Распространение трутовых грибов по типам местообитаний, таким образом, оказывается в прямой зависимости от наличия определенных пород деревьев (субстрата), а также от соответствующих условий влажности, температуры и освещенности в экологической нише. Дополнительным фактором этого является деятельность человека, которая накладывает отпечаток на состав флоры эксплуатируемых лесов. Немаловажное, хотя обычно недостаточно отмечаемое, влияние на состав флоры имеют взаимоотношения грибов, определяющие сукцессию видов.[ ...]
| Демографическая классификация местообитаний. А. Местообитания прижившихся особей (в противоположность недавно появившемуся потомству) могут быть либо сравнительно благоприятными в отношении размера (остаточная репродуктивная ценность при увеличении «размера» резко возрастает), либо сравнительно нейтральными, безразличными или неблагоприятными для размера. Б. Местообитания недавно появившегося потомства могут быть либо сравнительно благоприятными в отношении его размера, либо относительно нейтральными, безразличными или неблагоприятными для размера. В. Комбинируя эти пары, можно получить четыре основных типа местообитаний для всего жизненного цикла организма (I—IV) | 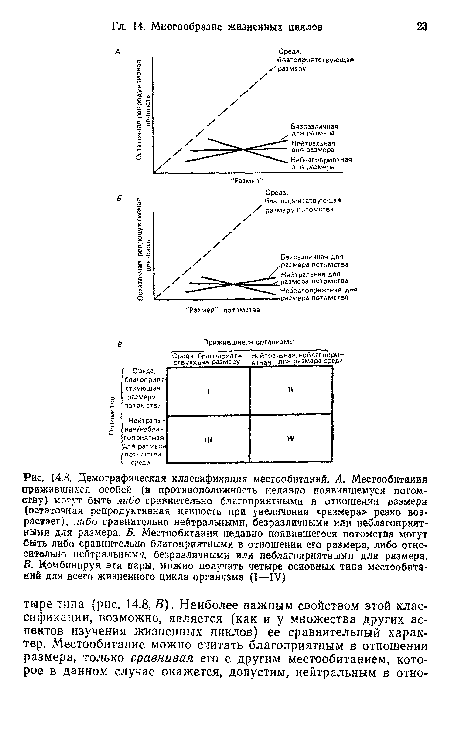 |
Последняя категория — разнообразие микробиотопов в пределах местообитания. Бугры и мелкие понижения на постоянных пастбищах связаны с характерными для них видами и значительно различаются видовым богатством. Своеобразие механического состава почвы, дренажа, экспозиции и бесконечного множества других экологических факторов может влиять па таксономическую принадлежность и обилие видов в конкретном типе местообитания.[ ...]
В фауне птиц отмечено 102 вида, относящихся к комплексам трех типов местообитаний: лесных, степных и луговых, водных и околовод-ных. Лесной орнитокомплекс сильно обеднен и вблизи озер представлен, в основном, охотящимися хищниками, в их числе: обыкновенная пустельга (Falco tinnunculus), чеглок (Falco subbuteo), кобчик (Falco vespertinus), канюк (Buteo buteo), черный коршун (Milvus migrans). Среди фоновых обитателей степных участков наиболее многочисленны полевой жаворонок (Alauda arvensis) и полевой конек (Anthus campestris), обычны перепел (Coturnix cot и г nix), луговой лунь (Суreus pygargus). На лугах многочисленны желтые трясогузки (Motacilla flava).[ ...]
На основе этих пространственно-временных характеристик может быть выделено 12 типов местообитаний. Десять из них пригодны для жизни. Два типа, очевидно, в этом плане весьма проблематичны — непрерывные и изолированные эфемерные местообитания.[ ...]
Экологические популяции. По Н.П. Наумову, экологическая популяция — это население одного типа местообитания (биотопа), характеризующееся общим ритмом биологических циклов и характером образа жизни. Это наиболее мелкие территориальные группировки, обладающие свойством целостности —по крайней мере, в процессе воспроизведения (репродукции). Они возникают в силу наличия в пределах зон сходного благоприятствования мелких, но существенных отличий в особенностях мезо- и микроклимата, рельефа, гидрологического режима и т. п. В порядке приспособления к этим особенностям у всех особей, занимающих данное место обитания, формируется единый тип физиологических реакций, биоритмов и общего образа жизни. Эти признаки объединяют представителей данной экологической популяции и отличают их от других группировок того же ранга.[ ...]
Наличие подходящего субстрата обусловливает в первую очередь распространение трутовых грибов по типам леса и другим типам местообитаний. В остальном, кроме фактора состояния древесного субстрата (жизнеспособное дерево, угнетенное, сухостой, валеж и т. д.), наибольшее значение имеют основные экологические факторы: температура, влажность, в меньшей степени освещенность.[ ...]
Крайне важно также понимание множества разных причин, обусловливающих возникновение того или иного типа местообитаний. Вероятно, благоприятными в отношении размера они будут по двум основным причинам. Во-первых, крупный размер даст преимущество при наличии интенсивной конкуренции, когда выживают и размножаются только «самые крупные» конкуренты. Хороший пример этого — самцы благородного оленя, среди которых только самые конкурентоспособные имеют гаремы. С другой стороны, крупный размер благоприятен и тогда, когда мелкие взрослые особи особенно подвержены гибели из-за хищничества или влияния абиотических факторов. Не исключено, например, что именно величина крупных двустворчатых моллюсков предохраняет их от выедания крабами и гагами.[ ...]
| Зависимость между остаточной репродуктивной ценностью и размером литторин на валунах и в ¿целях. Типы местообитаний характеризуются градиентом этой зависимости, как показано в тексте (размножение в обоих случаях начинается при одном и том же градиенте). Показаны черты жизнен-ных циклов, выгодные в том или ином местообитании и соответствующие полевым наблюдениям (табл. 14.3) |  |
В процессе исследования территории было выявлено 190 видов высших сосудистых растений.[ ...]
Фитотопологическое направление заключается в том, что классификация растительных ассоциаций основана на различии типов местообитания растений. Характеристика лугов в этом случае дается на основании изучения местообитаний (рельеф, увлажнение, почва, материнская порода и т. д.), в результате чего устанавливают различные типы лугов, которые объединяют в группы по сходности местообитания (луга суходольные, горные, низинные, пойменные и др.).[ ...]
Меры видового разнообразия можно использовать и для других целей. Две обычные сферы их применения — изучение разнообразия местообитаний и ширины ниш (т. е. разнообразия ресурсов, используемых организмом или видом). Подобно видовому разнообразию, эти формы экологического разнообразия можно измерять с помощью простых индексов богатства и более сложных мер. Как и меры видового разнообразия, эти оценки также не свободны от недостатков типа отклонения при малых выборках. Предварительным этаном является разработка системы классификации типов местообитаний или ресурсов, причем как бы с точки зрения изучаемых организмов.[ ...]
Тейлор (Taylor, 1986) применял параметр а лог-ряда для мониторинга разнообразия бабочек во многих пунктах Британии, различающихся типом местообитания, широтой и характером землепользования. Эти данные будут нужны для прогнозирования изменений окружающей среды.[ ...]
Зависимость между общим числом размножающихся видов наземных птиц на каждый остров и числом видов в выборках для двух типов местообитаний в 17 участках на 12 Больших и Малых Антильских островах [215].[ ...]
Ряд предположений, выдвинутых в предыдущем разделе, можно объединить в концепцию, имеющую особое значение при изучении разных типов жизненных циклов. Это концепция г- и /С-отбора, выдвинутая первоначально Мак-Артуром и Уилсоном (MacArthur, Wilson, 1967) и развитая Пианкой (Pianka, 1970). Подобно всем обобщениям, такое противопоставление слишком упрощено, однако достаточно обосновано, чтобы тщательно его проанализировать.[ ...]
Водно-болотные угодья занимают в заповеднике, в зависимости от состояния обводненности, от 33 до 42% территории. Выделяются следующие типы местообитаний: пресные и солоноватые озера с обширными тростниковыми зарослями и обильной погруженной растительностью - 20-21 тыс. га (отмечен 61 вид, гнездится 43), соры и соленые озера - около 9 тыс. га (розовый пеликан, пеганка, огарь, морской зуек, ходулочник, шилоклювка, черноголовый хохотун, серебристая и сизая чайки, чайконосая крачка), открытые берега озер (47 видов, гнездится 8-9), лиманы (мелководья поросшие осокой и пыреем) и заливные луга - 7-8 тыс га.[ ...]
Расширим пример и предположим, что решено создать несколько лесных заповедников. Для этого можно либо выбрать наиболее богатые видами участки независимо от типа леса, либо сначала провести классификацию сообществ, объединив их в группы [используя, например, прекрасную схему, разработанную Петеркеном (Ре1егкеп, 1974,1981)], н лишь затем определить наиболее разнообразные при прочих равных условиях участки в пределах каждой Группы. Результатом первого подхода были бы несколько богатых видами территорий, ограниченных одним или двумя типами леса (например, смешанными лесами на южных известняках, рис. 6.7), и, напротив, второй подход обеспечил бы заповедный режим гораздо большему числу лесных типов и, поскольку многие виды приурочены к специфическим типам местообитания, охрану в целом большего количества видов. Развивая эту концепцию, Маргу-лес с соавторами (Ма и1еБ е1 а1м 1982), считает, что генетическое разнообразие можно максимизировать, опираясь не на единственный критерий видового богатства, а на охрану «полных видовых свит».[ ...]
В изучении и оценке пространственного размещения и подвижности отдельных популяций видов следует иметь в виду, что потребность в пространственной протяженности отдельных типов местообитаний, типов угодий очень различна. Если для кормовых площадей она обратно пропорциональна степени обилия и концентрации на них доступного корма, то в других целях она может быть для одной особи пространственно очень ограниченна. Например, для вывода молодых и дневного отдыха лесная куница заинтересована в наличии дуплистых деревьев или пней определенного диаметра и высоты. Но отдельной кунице вполне достаточно нескольких деревьев на участке ее обитания в несколько сот гектаров и участки с таким фаутным древостоем могут в сумме составлять не более нескольких гектаров. У бурого медведя участок леса для устройства берлоги территориально может быть удален от участка обитания в период активности, а его площадь имеет очень относительное значение. Можно сказать, что он имеет определенные (защитные) свойства, но пространственной протяженности не имеет. Не случайно, что там, где бывал дефицит «берло-жистых» мест, наблюдались высокие концентрации занятых медведями берлог на очень ограниченной площади, что летом было бы совершенно невероятно (так называемые «медвежьи деревни»).[ ...]
В изучении и оценке пространственного размещения и подвижности отдельных популяций видов следует иметь в виду, что потребность в пространственной протяженности отдельных типов местообитаний, типов угодий очень различна. Если для кормовых площадей она обратно пропорциональна степени обилия и концентрации на них доступного корма, то в других целях она может быть для одной особи пространственно очень ограниченна. Например, для вывода молодых и дневного отдыха лесная куница заинтересована в наличии дуплистых деревьев или пней определенного диаметра и высоты. Но отдельной кунице вполне достаточно нескольких деревьев на участке ее обитания в несколько сот гектаров и участки с таким фаутным древостоем могут в сумме составлять не более нескольких гектаров. У бурого медведя участок леса для устройства берлоги территориально может быть удален от участка обитания в период активности, а его площадь имеет очень относительное значение. Можно сказать, что он имеет определенные (защитные) свойства, но пространственной протяженности не имеет. Не случайно, что там, где бывал дефицит «берло-жистых» мест, наблюдались высокие концентрации занятых медведями берлог на очень ограниченной площади, что летом было бы совершенно невероятно (так называемые «медвежьи деревни»).[ ...]
Наверное, самое очевидное объяснение высокого видового богатства обширных территорий заключается в том, что на большем пространстве, как правило, встречаются более разнообразные типы местообитаний. Применительно к островной биогеографии наиболее активным сторонником этого взгляда был Лак (Lack, 1969b, 1976; обсуждение — см. Williamson, 1981). Он утверждал, что число встречающихся на острове видов просто отражает его «тип», который определяется климатом и набором имеющихся местообитаний. При этом крупные острова богаче видами, поскольку условия на них более разнообразны. Очевидно, что до какой-то степени это верно, хотя такое объяснение и вызвало ряд критических отзывов (они обсуждаются в разд. 20.3.1).[ ...]
Мы можем выбрать три разных аспекта зависимости вида от окружающей среды. Ареал вида — его географический ранг, его распространение в пространстве, которое можно показать на карте. Местообитание вида — тот тип среды, где вид встречается; эта среда может характеризоваться физическими и химическими показателями, высотой над уровнем моря, положением в рельефе и т. д. или типом сообщества. Вид в различных частях ареала может занимать ряд различных местообитаний, то есть более чем один специфический тип местообитания. В пределах каждого местообитания можно описать позицию вида в пространстве, времени и функциональные связи с природным сообществом, занимающим это местообитание. Позиция вида в сообществе в зависимости от других видов является его нишей.[ ...]
Ширина ниши — это мера разнообразия ресурсов, используемых особью или видом. Для ее расчета обычно применяют индексы Шеннона (уравнение 2.17, с. 40) или Симпсона (уравнение 2.27, с. 45). Вместо числа видов в формулу подставляется число категорий ресурсов (например, типов нищи, местообитаний, форм поведения). Естественно, для каждого типа ресурсов должна быть рассчитана отдельная величина. Меры обилия будут зависеть от того, каким образом используется индекс. Например, если рассматривается ширима ниши какого-то вида, обилие можно оценить числом особей, поедающих каждый вид пищи, живущих в каждом типе местообитания или применяющих ту или иную форму поведения. Если же речь идет о ширине ниши особи, под обилием удобно понимать количество каждого типа поедаемой пищи, время, проведенное в каждом типе местообитания, или частоту использования того или иного типа поведения.[ ...]
Метод избирательного обогащения позволяет выявить метаболические процессы, которые могут протекать в среДе. Например, на симпозиуме, посвященном проблемам обогащения сред и выделения микроорганизмов, Шлегель (1964) сообщил, что у микроорганизмов существуют совершенно неизвестные типы метаболизма и что изучение влияния обогащения на избирательный рост позволяет лучше представить себе, в каких типах местообитаний такие организмы встречаются. Впервые описанные формы фотосинтезирующих бактерий иллюстрируют экологическое значение поглощения света и пигментного состава (Пфенниг, 1967; Трапер и Яннаш, 1968).[ ...]
В природе закономерности, лежащие в основе правила минимума, определяют многие важные момента географического распространения, морфологии, экологии и физиологии животных и растений. Именно лимитирующие экологические факторы в раде случаев ограничивают проникновение вида в те или иные типы местообитаний. Во многих случаях «экологические барьеры» формировали в истории видов их современные ареалы. Выше уже упоминалось лимитирование распространения ряда ввдов на север глубиной снежного покрова. Недостаток источников влаги жестко ограничивает возможность заселения аридных зон малоподвижными животными. Не менее эффективно минеральный состав почвы (например, засоление) может лимитировать набор видов, формирующих растительные сообщества. Специфику их, связанную с определенным составом почвы, широко используют в геологии для поисков ряда видов полезных ископаемых и запасов воды.[ ...]
Говоря конкретнее, жизненный цикл организма почти наверняка меняется в зависимости от обеспеченности ресурсами. Однако еще многие работы при сравнении циклов этого не учитывают. Следовательно, существует опасность связать наблюдаемые особенности не с различиями в уровне потребления внутри одного типа местообитаний, а с разницей между самими местообитаниями.[ ...]
Эммонс (Emmons, 1980), изучая биологию девяти сосуществующих видов африканских древесных белок в равнинном вечнозеленом дождевом лесу, обнаружил, что расхождение ниш происходит по четырем измерениям. Результаты его работы в обобщенном виде представлены в табл. 18.1. Каждый из двух видов рода Funisciurus занимал особый тип местообитаний: F. isabel-la — густой ,подлесок, часто связанный с бывшими вырубками, а F. anerythrus —сезонно или постоянно затопленные леса. Остальные семь видов, населяющие господствующую лесную формацию можно разделить на две группы в зависимости от того, где проходит основная часть их жизнедеятельности: в пологе или же на уровне почвы. Все виды питаются главным образом плодами; исключение составляет Myosciurus pumilio, поедающая в большом количестве кору. Однако размеры потребляемых плодов различны. Два наиболее крупных вида — один древесный, второй наземный — способны справиться даже с самыми твердыми орехами. Наконец, количество потребляемого животного корма (всевозможные членистоногие) находится в обратной зависимости от размеров тела белки, причем по одному виду среди кроновых и наземных форм поедают такую пищу в заметных количествах.[ ...]
Всего в обследованных водоемах нами обнаружено 211 видов (включая разновидности и формы), относящихся к 62 родам 37 семействам 5 отделам. Наиболее разнообразен отдел Chlorophyta - 102 вида, на втором месте Cyanophyta - 98 видов, затем Xanthophyta - 6, Rhodophyta - 3, Chrysophyta - 2 вида. По типам местообитаний преобладают планктонные и план-ктонно-бентосные формы. По отношению к солености и кислотности среды во всех водоемах доминируют индифференты. Индекс сапробности для всех водоемов находился в пределах 1,33-1,75, что является критерием чистых вод и вод удовлетворительной чистоты. На основе альгоиндикации можно предварительно оценить современное состояние исследованных водоемов национального парка как чистое (I и II класс качества вод), что свидетельствует об отсутствии или незначительном антропогенном воздействии на водные экосистемы. Полученные результаты будут использованы как фоновые показатели в системе экологического мониторинга и при прогнозировании изменений состояния водоемов под влиянием планируемых разработок полезных ископаемых на территории парка.[ ...]
Лесная типология, дисциплина вполне сложившаяся исторически, является наиболее надежной основой в разработке теоретических положений и методов классификации всех наземных экосистем в СССР, США, Канаде, Австралии и большинстве европейских стран (см. обзоры). Именно подходы лесотипологов оказались наиболее комплексными и приемлемыми, а результаты систематического изучения типов леса и типов местообитаний — наиболее содержательными и общезначимыми для широкого круга природопользователей. Лесная типология сегодня является специализированной отраслью экологии, составной частью классификации и прогнозирования поведения наземных экосистем. Она входит и в ’’географию экосистем”, представляющую собой "направление современной биогеографии, развиваемое на стыке теоретических достижений и методических подходов географических и биологических наук с привлечением методологии системного анализа”, основная задача которой ’’создание теории организации экосистем на локальном, региональном и глобальном уровнях, как основы для управления состоянием окружающей среды и ее возобновляемых ресурсов” .[ ...]
Наиболее защищены от проникновения людей и домашних животных широкие кулисы и «острова» сплавин, няши (участки водоема с поднявшейся на поверхность иловой массой), заросли телореза и плавающей водной растительности (кубышки, кувшинки), маленькие островки с заросшими кустарником берегами, заболоченные луга в условиях дождливого лета, свалки в береговой зоне. Редко посещаются также индустриальные зоны (заросшие сорняками и нередко имеющие сточные водоемы пустыри) вокруг предприятий. Последний тип местообитаний дает возможность гнездиться малому зуйку. Эти участки имеют максимальную плотность гнезд изученной группы птиц, что обязательно должно учитываться в градостроительных планах.[ ...]
Анализ 1000 пар больших синиц из окрестностей Оксфорда показал, что инбредные пары составили в популяции всего 1,6 %; общий коэффициент инбридинга —0,0035. Выяснилось, что на степень инбридинга влияет подвижность самцов: у инбредных пар «мать — сын» среднее расстояние удаления самца от места рождения было на 85 % меньше среднего ожидаемого (P. Greenwood, P. Harvey, 1978). Вообще, выраженная дисперсия характерна главным образом для самцов, что связывают с конкуренцией за самок. У видов, у которых самцы более конкурируют за ресурсы, дальнее расселение характерно для самок: например, у пищухи Ochoíona princeps репродуктивный успех определяется исключительно типом местообитания; у этого вида самцы защищают участки, а самки расселяются в поисках лучших мест для поселения.[ ...]
ru-ecology.info
| АрхеологияАрхитектураАстрономияАудитБиологияБотаникаБухгалтерский учётВойное делоГенетикаГеографияГеологияДизайнИскусствоИсторияКиноКулинарияКультураЛитератураМатематикаМедицинаМеталлургияМифологияМузыкаПсихологияРелигияСпортСтроительствоТехникаТранспортТуризмУсадьбаФизикаФотографияХимияЭкологияЭлектричествоЭлектроникаЭнергетика | Плауновидные - самая древняя группа современных высших растений, ископаемые остатки растений рода плаун (Lycopodium) известны с конца силура - начала верхнего девона палеозойской эры (примерно 380 млн лет назад). В палеозое Плауновидные процветали, достигали огромных размеров (до 40 м), занимали господствующее положение в растительном покрове Земли. В настоящее время это угасающая группа растений. Все современные плауновидные - травянистые растения численностью примерно 1 тыс. видов. Это самые первые сосудистые растения: кроме хорошо развитых олиственных побегов у них есть настоящие корни. Для плауновидных характерна микрофилия, т. е. относительно мелкие размеры листьев. Листья плауновидных возникли как поверхностные выросты на осевых органах, что делает их уникальной группой. Стебель хорошо развит, листорасположение спиральное, супротивное и мутовчатое; ветвление дихотомическое. Молодые побеги нарастают за счет верхушечной меристемы, деятельность которой со временем угасает и поэтому плауновидные ограничены в росте. Плауновидные не имеют главного корня. От подземного корневища отходят придаточные корни. В жизненном цикле спорофит (взрослое растение) преобладает над гаметофитом. Спорангии у плауновидных формируются на специализированных спороносных побегах - стробилах в виде колосков. Гаметофит небольших размеров, называется заростком, который отмирает сразу же, как только из него образуется новый спорофит (рис. 175). Для оплодотворения яйцеклетки необходима водная среда. Внутри отдела два класса: Плауновидные (Lycopodiopsida) и Полушниковые (Isoetopsida). Отдел Хвощевидные - Equisetophyta Ныне живущие хвощевидные, или членистые, представлены многолетними корневищными травами с мутовками бурых редуцированных бесцветных листьев (рис. 178). В прошлом, будучи гигантских размеров, вместе с древовидными папоротниками и лепидодендронами они составляли каменноугольные леса. В конце палеозоя в связи с изменением климата древесные формы вымерли, и в настоящее время в этой группе растений остался только один класс Хвощевые (Equisetopsida), один порыядок (Equisetales), одно семейство (Equisetaceae) с одним родом хвощ (Equisetum), включающим 25 видов. 12 видов встречаются в России. Проводящая система представлена различными трахеидами и иногда сосудами. Во флоэме есть паренхима и ситовидные элементы. Листья чешуевидные, бурые или желтоватые. Функцию фотосинтеза выполняет зеленый стебель, на верхушке которого вырастает спороносный колосок. Стебли у хвощей умеренных широт не превышают 1 м, у представителей тропиков они достигают 10-12 м при толщине стебля 6 - 8 см. В жизненном цикле спорофит преобладает над гаметофитом (рис. 178, Б). Хвощ - равноспоровое растение. Для репродуктивных органов бесполого размножения характерны стробилы в виде колосков на верхушке стеблей. К оси стробил мутовками прикреплены спорангиофоры, которые представляют собой шестиугольные щитки. На внутренней стороне щитков находятся спорангии (рис. 178, 1, 2, 3). При созревании спор щитки подсыхают, разрываются и споры разлетаются. Созревшая спора одета двумя оболочками, а снаружи спирально обернута своеобразной лентой - элатерой, способной отгибаться и совершать гигроскопические движения. При подсыхании элатеры раскручиваются и сцепляются друг с другом, образуя своеобразные клубочки спор. Это способствует распространению спор и прорастанию заростков недалеко друг от друга. Заростки обоеполые, представляют собой пластинку, прикрепленную к почве ризоидами. Наверху пластинки формируются антеридии и архегонии. Многие виды хвощей проявляют физиологическую разноспоровость, иногда в связи с условиями произрастания, иногда она генетически заложена. После оплодотворения яйцеклетки многожгутиковым сперматозоидом в условиях капельно-водной среды из зиготы формируется зародыш, который имеет все зачаточные вегетативные органы. Корешком зародыш внедряется в почву и начинает самостоятельное существование. Хвощи - сильно разветвленные корневищные растения, в узлах которых иногда развиваются богатые крахмалом клубеньки. От узлов корневища отходят многочисленные придаточные корни. Хвощи - представители разных растительных сообществ земного шара, но произрастающие в местах достаточного или избыточного увлажнения. Самыми распространенными видами для средней полосы европейской части России являются хвощ полевой (Е. arvense), хвощ лесной (Е. sylvaticum, рис. 179), хвощ болотный (Е. palustre) и др. Хвощ полевой - особо злостный сорняк, однако молодые побеги его используют в медицине как мочегонное средство. Отмершие остатки древних каменноугольных гигантов хвощевидных дали начало залежам каменного угля.
|
studopedya.ru










