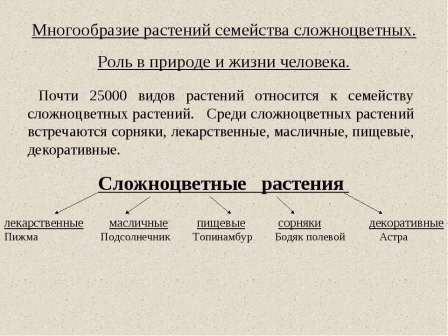
Что такое луковица, клубень и отпрыск? Клубнеобразующие растения это
Борец двухцветковый

Семейство Лютиковые — Ranunculaceae Борец двухцветковый Aconitum biflorum Fisch. ex DC. Категория и статус: 3 д — редкий вид. Краткая характеристика. Корневищное и клубнеобразующее растение 15-40 см выс., поликарпик. Почка возобновления закладывается на клубне в основании надземного побега в период цветения растений и формируется около двух лет. Цветет в августе, плодоносит в конце августа - сентябре. Размножение семенное и вегетативное. Распространение. В России встречается в основном в Западном Саяне (хр. Алаш, Саянский, Сабинский, Куртушибинский, Хемчикский, Шаман), в Кузнецком Алатау и Юго-Восточном Алтае известно по одному местонахождению (1-19). В Красноярском крае встречается в Ермаковском и Шушенском р-нах, расположенных на юге края (3, 5,9,10,15-19), самое восточное местонахождение отмечено на горе Ашпан в левобережье р. Енисей (9). В Республике Алтай известен из хр. Чихачёва (8), Шапшальского и Абаканского (11). В Республике Хакасия известно одно местонахождение на г. Шаман (5,12, 16,17), в Кемеровской обл. вид найден на г. Белый Камень в дол. р. Кондома (5,7,13). Большая часть находок сделана на западе Республики Тыва (3,4, 6,8,14,16,18). Вид описан с Зап. Саяна. За пределами России произрастает в Монголии (20). Особенности экологии и фитоценологии. Психрофит. Растет в субальпийском поясе и в нижней части альпийского в пределах высот 1530-2650 м н. ур. м. в моховых и кустарниковых тундрах, на альпийских лугах, изредка встречается среди каменных россыпей. Численность. Зарослей не образует, в фитоценозах отмечаются единичные особи. Состояние локальных популяций. По всему ареалу численность локальных популяций не высокая. Лимитирующие факторы. Интенсивный выпас скота. Принятые меры охраны. Был занесен в Красную книгу РСФСР (1988). Вид включен в Красные книги Кемеровской обл. (2004)), Республик Алтай (1996), Тыва (2002), Хакасия (2002). Охраняется в заповедниках: Алтайский, «Кузнецкий Алатау», «Хакасский», «Саяно-Шушенский» и «Убсунурская котловина». Необходимые меры охраны. Контроль за состоянием популяций. Возможности культивирования. Выращивается в ботаническом саду Самары. Следует расширить работы по культивированию. Источники информации. 1. Аильчева и др., 2003; 2. Анкипович, 1999; 3. Красноборов, 1976; 4. Красноборов и др., 1989; 5. Флора Западной Сибири, 1958; 6. Ломоносова и др., 1984; 7. Малышев, 1968; 8. Красная книга Республики Алтай, 1986; 9. Мартьянов, 1923; 10. По ложий, Ревердатто, 1976; 11. Ревушкин, 1988; 12. Редкие ... Хакасии, 1999; 13. Седельников, 1979; 14. Соболевская, 1953; 15. Сонникова, 1992; 16. Фризен, 1993; 17. Черепнин, 1961; 18. Шишкин, 1914; 19. Штейнберг, 1937; 20. Губанов, 1996. Составитель: Д.Н. Шауло.
AOF | 17.12.2014 08:21:29
cicon.ru
Жизненные формы комнатных растений
При озеленении внутренних объектов, где живет, отдыхает и работает человек, используют разнообразные жизненные формы растений. Согласно И.Г. Серебрякову, жизненная форма – это своеобразная внешняя форма организмов, обусловленная биологией развития и внутренней структурой их органов, возникающая в определённых почвенно-климатических и ценотических условиях как приспособление к жизни в этих условиях, т.е. они представляют собой формы приспособленности видов к жизни в той или иной среде, исторически возникшие под длительным влиянием экологических факторов.
Ввиду многообразия комплексов условий на Земле, существует большое количество жизненных форм организмов. Жизненные формы покрытосеменных растений И.Г. Серебряков в 1964 г разделил на 4 отдела: древесные, полудревесные, наземные травянистые и водные травянистые растения. Каждый из них, в свою очередь, представлен многими более мелкими группами жизненных форм. Детальное изучение их строения, формирования, экологической приуроченности и географического распространения было проведено И.Г.Серебряковым. Описание основных жизненных форм я привожу, основываясь на его публикациях.
Среди комнатных растений из типа Деревья можно выделить:
Кронообразующие: араукария, гранат, драцена, камелия, кипарисовик, кодиеум, кофейное дерево, лавр, ливистона, фатсия, фикус, цитрус, юкка.
Безлистные суккуленты: агава, алое, апорокактус, гавортия, гастерия, пахиподиум, селеницериус, цереус, эониум, эпифиллум.
К жизненной форме Деревья относятся многолетние растения с одним одревесневшим стволом, сохраняющимся на протяжении всей жизни растения. Распространены от экваториального до умеренно-холодных поясов. Среди них можно выделить формы “наземных кронообразующих”, где имеются деревья: с прямостоячими стволами, “кустовидные” (немногоствольные) и “одноствольные с низкими стволами”. Их ствол всегда растет прямо вверх (ортотропно). Среди наземных кронообразующих деревьев имеются жизненные формы с лежачими стволами – стланцы. Они формируются в районах, мало благоприятных для жизни древесных растений, – там, где длинная зима, прохладное лето, где часто дуют холодные ветры. У всех у них на верхушках обычно имеются розетки крупных жестких или небольших мясистых листьев (драцена, юкка, панданус, алоэ и др.). У многих вместо листьев появляются колючки, мясистые стволы (разные виды молочаев, кактусов).
К кустарникам из комнатных растений относятся: абутилон, агапетес, азалия, акалифа, белопероне, бересклет, гардения, гибискус, гортензия, катарантус, олеандр, переския, плектрантус, полисциас, пуансеттия, роза, тунбергия, шеффлера, эрантетум.
К числу древесных растений относится большая группа жизненных форм – кустарники. Для них характерно, что одновременно у растения бывает развито много равных по размеру стволов. Главный ствол, имеющийся в начале жизни, живет недолго, и когда он есть, то почти не выделяется по длине среди боковых. Жизненная форма кустарника сформировалась в процессе эволюции под воздействием условий среды с зимними дождями, с сухим и жарким летом, наиболее значительную роль в сложении фитоценозов они играют в районах аридного климата тропических областей и теплоумеренных зон северного и южного полушарий. В этих районах кустарники обычно являются важными эдификаторами (средообразователями) растительных сообществ.
Много кустарников в саванновых и колючих лесах тропической зоны, в областях с жестколистной древесной растительностью теплоумеренных зон обоих полушарий, в ксерофильных редколесьях и горно-степных районах Передней и Средней Азии, севернее предела лесной растительности в холодно-умеренной зоне и в ряде мест в зоне тундр. Много их в субальпийских районах гор и других, т. е. всегда в условиях, неблагоприятных для развития высокоствольных растений. У одних видов кустарниковая форма образуется за счет лишь боковых побегов, в большом количестве отходящих от нижней, но надземной части главного ствола (олеандр). Новые стволы у многих из них могут возникать лишь как корнеотпрысковые.
К кустарничкам среди комнатных растений относятся: вереск, розмарин.
Кустарнички – третий тип жизненных форм древесных растений. Для всех них характерен низкий рост стеблей. Главный стебель, имеющийся лишь в начале жизни, существует недолго (3–7 лет). На смену ему развиваются укореняющиеся боковые подземные одревесневающие стебли. Они, как правило, развиваются из спящих почек. У большинства кустарничков каждый вновь появляющийся куст, вырастая из спящей почки, первоначально растет подобно столону – параллельно поверхности почвы (т. е. плагиотропно), затем переходит к вертикальному (ортотропному) росту главной оси, а, выйдя на поверхность, начинает ветвиться. Такой разветвленный укоренившийся побег (представляющий собой надземный “кустик”) называется парциальным кустом. От каждого парциального куста, в свою очередь могут развиваться новые–отделившись от материнского, они дают начало новым растениям.
Формирование данной жизненной формы в историческом развитии происходило под влиянием условий умеренно-холодных, холодных зон и высокогорных областей, она является индикатором (показателем) этих условий.
Ко второму отделу Полудревесные растения, к типу полукустарники можно отнести аспарагус, антеннария, диморфотека, зигокактус, колеус, колумнея, кордилина, паслён сладко-горький, пеларгония, псевдерантемум, рехстейнерия, ривина, хризантема, якобиния.
Среди жизненных форм следует выделить полудревесные растения, к которым относятся полукустарники. Они свидетельствуют об аридности местообитании.
Характерный признак для полукустарников – регулярное отмирание верхних частей надземных побегов. Оставшиеся, неопавшие, части стеблей одревесневают и в таком виде сохраняются на протяжении нескольких лет. На этих одревесневших надземных частях стебля всегда имеются почки возобновления, из которых на следующий год развиваются многочисленные новые травянистые стебли. Этим полукустарники всегда отличаются настоящих травянистых растений.
Очень большой и разнообразной группой жизненных форм являются наземные травянистые растения. Их II.Г.Серебряков разделяет на две части: плодоносящие много раз в своей жизни (травянистые поликарпики) и плодоносящие лишь однажды (травянистые монокарпики):
Травянистые поликарпикиявляются формами, широко представленными от экваториальных до арктических и субантарктических стран. Это обычные наземные травы, многообразные по своему облику, биологии и экологии, но для всех них характерно отмирание их надземных ортотропных (растущих вверх) побегов ежегодно в конце вегетационного периода. В течение ряда лет и на зиму сохраняются из надземных побегов лишь плагиотропные (растущие параллельно поверхности почвы). Подземные части побегов функционируют как органы возобновления или же, как запасающие органы. В то же время имеется целый ряд видов, подземные побеги которых сохраняются живыми много лет (ландыш, ирис и др.).
Кистекорневые: примула.
Среди травянистых поликарпиков имеется жизненная форма – стержнекорневые растения. К ним относятся многолетние травы, у которых на протяжении всей жизни растения сохраняется стержневой главный корень. Стеблевое укоренение у них обычно отсутствует или выражено чрезвычайно слабо. Вегетативное размножение возможно лишь благодаря корнеотпрысковости. Эта жизненная форма в природе представлена рядом длинностержнекорневых растений и короткостержнекорневых. Первые сформировались в местообитаниях с глубоким залеганием грунтовых вод, вторые, там, где почвенная влага лежит сравнительно близко, но почва очень сильно задернована).
Кистекорневые и короткокорневищныерастения широко распространены от тундровой до степной зоны, в местообитаниях с достаточным увлажнением воздуха и почв.
Наземные столонообразующие: дюшенея, живучка, камнеломка.
Столонообразующиерастения весьма характерны для влажных и избыточно увлажненных местообитаний с рыхлой, мало плодородной (иногда заторфованной) почвой, например в широколистных и темнохвойных лесах.
Размножение осуществляется в основном вегетативным путем. Столонообразующие во многом сходны с длиннокорневищными, но в столонах нет отложения запасных питательных веществ, и они легко разрываются на части. К столонообразующим поликарпикам относятся: майник двулистный, адокса мускусная, седмичник европейский, фиалка удивительная, развивающие подземные столоны, и земляника, клубника, камнеломка пучковатая, живучка ползучая, имеющие надземные столоны (“усы”).
Клубнеобразующие: аморфофаллюс, ахименес, бегония, глоксиния, каладиум, калла, канна, колерия, хлорофитум, цикламен.
Клубнеобразующиеполикарпики обладают специализированными органами запаса питательных веществ – клубнями корневого, стеблевого и листового происхождения. Все эти поликарпики характерны для мест, в которых четко выражена смена периодов покоя и вегетации (в связи с зимой или засухой). Среди этих растений имеется много видов, у которых с помощью клубней осуществляется возобновление и расселение растений. К данной группе жизненных форм принадлежат:
Корнеклубневые, с клубнями стеблевого происхождения (цикламен, хохлатки, шафраны, гладиолусы, монтбреция, безвременник и др.), с клубнями на концах столонов (стрелолист, иксиолирион и др.).
Луковичные: агапантус, амариллис, валотта, гемантус, зефирантес, кринум, панкрациум, птицемлечник, эухарис.
Жизненная форма луковичные поликарпики характерна для засушливых районов, где жаркое сухое лето надолго прерывает вегетацию их побегов. Одни из них являются эфемероидами, другие способны вегетировать и в сухое время, например лук голубой, туркестанский, Королькова и др.
Корнеотпрысковые:алоказия, аспидистра, калатея, кислица, кливия, молочай, родея, сальвия, сансевиерия, смитианта, стангопея.
Лиановидные травянистые поликарпики:жасмин, партеноцисс, пассифлора, плющ, сингониум, тетрастигма, филодендрон, хойя, циссус, эпипремнум.
Лианы– особая жизненная форма растений, выработавшаяся в борьбе за свет и пространство в густых растительных сообществах. Для лиан характерны длинные тонкие стебли и способность к очень быстрому росту, позволяющая им выносить листья на поверхность крон деревьев. Большинство лиан – светолюбивые растения. Возникновение лиан в природе происходило независимо в разных классах и семействах растительного царства под влиянием сходных условий существования: есть лианы среди папоротников, голосеменных, однодольных и двудольных покрытосеменных растений.
Способы прикрепления к опорным растениям у лиан различны. Вьющиеся лианы обвивают опору молодыми неодревесневающими верхушками стеблей, обладающими способностью совершать круговые движения. Таковы восковое дерево, алламанда слабительная, клеродендрон госпожи Томсон, тунбергия. Лазящие лианы взбираются на опору при помощи загнутых назад крючков и щетинок. Многие виды семейства ароидных (монстера, филодендрон, сингониум), различные представители семейства перечных и фикусы прикрепляются к опоре придаточными корнями. Наиболее совершенными приспособлениями для прикрепления к опоре являются усики различной морфологической природы. В усик может превращаться то лист, то черешок, то целое соцветие. Встречаются лианы и в странах с субтропическим и умеренным климатом, но там их немного и они не играют большой роли в растительном покрове.
Ползучие травянистые поликарпики (луговой чай, вероника лекарственная и др.) имеют лежачие хорошо олиственные многолетние стебли, легко укореняющиеся. Данная жизненная форма характеризует местообитание с достаточно влажной почвой и умеренным освещением.
Травянистые монокарпики широко распространены в засушливых областях умеренной зоны северного полушария.
Среди таких монокарпиков есть многолетние и двулетние растения (ряд видов из сем. зонтичных, крестоцветных). У большинства из них корень утолщен и содержит запасные питательные вещества.
Однолетние травянистые монокарпики могут быть длительно вегетирующими, эфемеры, лиановидные, полупаразитные и паразитные (повилика).
Однолетние в большинстве случаев являются представителями областей с засушливым климатом и спутниками полевых культур в искусственных биогеоценозах.
Среди всех типов жизненных форм нередко встречаются подушковидные растения. Это, как правили, многолетние травянистые, реже – древесные растения, иногда вечнозеленые. Для всех них характерны чрезвычайно маленький годичный прирост главной оси и очень сильное ветвление боковых побегов, которые, располагаясь радиально или этажами, создают компактную форму “подушки”.
Подушковидные растения приурочены обычно к местообитаниям или с влажной, холодной и малоплодородной почвой, или в сухих жарких областях в условиях очень сухих глинистых и каменистых почв, среди скал, но развиваются в условиях сильного освещения.
Отмирая, листья не опадают, а остаются на растении в течение 15–16 лет.
Этому способствуют условия высокой сухости почвы и воздуха, низкая температура в течение всего года, сильные ветры и очень яркое освещение.
studfiles.net
Открытие роли листьев в актиноритмизме
Открытие роли листьев в актиноритмической реакции растений признано главнейшим этапом в теоретическом аспекте познания этого важнейшего явления.
Истоками изучения актиноритмической реакции листьев растений послужили работы, начатые автором в связи с сомнениями, возникшими по поводу «фотопериодической локализации», описанной в публикациях Гарнера и Алларда, а также Нотта (Knott, 1925).
Двум первым авторам принадлежит открытие «фотопериодической локализации», а с именем последнего связана попытка доказать значение вершин побегов и вообще точек роста в этом явлении.
Актиноритмическая локализация была обнаружена Гарнером и Аллардом в опытах 1923—1924 гг. с короткодневными растениями космос — Cosmos sulphureus и Cosmos bipinnatus. Помещая в противоположные актиноритмические условия различные зоны и части растения, они пришли к выводу, что фотопериоды воспринимаются как всем растением в целом, так и любой отдельной его зоной и ветвью вне зависимости друг от друга. В тех случаях, когда одна зона растения находилась в условиях короткого, а другая — длинного дня, цветение наблюдалось только в короткодневной зоне. Это явление получило название «фотопериодической локализации».
Проводя свои опыты и обсуждая их результаты, Гарнер и Аллард не затронули тему об органах растений, воспринимающих фотопериоды и реагирующих на них. Такая первая попытка была сделана Ноттом в 1925 г.
Ставя опыты с растениями космос, Нотт исходил из предварительных наблюдений за активностью фермента каталазы в листьях и вершинах побегов шпината, находящегося в условиях длинного и короткого дня (в первом случае шпитан цветет, во втором нет). Активность каталазы в зависимости от длины дня оказалась различной именно в вершинах побегов, а не в иных органах. Поэтому Нотт и сделал заключение, что и само восприятие фотопериодических условий может быть локализовано там же.
Опыт с космосом был им поставлен, чтобы экспериментально доказать значение для восприятия актиноритмических условий вершины) побега или, точнее, вершинной точки роста. Применив неправильную методику и не учтя, что восприятие внешнего воздействия и конечная реакция на последнее — вещи разные, Нотт в разбираемой работе пришел к ошибочному выводу о локализации фотопериодической реакции в вершинах побегов. Дело в том, что, по письменному свидетельству самого автора (Нотт, 1926), в закрываемой зоне растения находилась не только точка роста, но одновременно и несколько ближайших к ней листьев и, хотя последние и были причиной цветения космоса при коротком дне, автор не принял их во внимание.
Вслед за Ноттом в 1931 г. выступил В. И. Разумов, повторивший с некоторыми дополнениями опыты Гарнера, Алларда и Нотта. Объектом его исследований были клубнеобразующие растения Ullucus, Oxalis и некоторые виды картофеля. Показателями актиноритмической реакции и ее локализации в опытах Разумова служили появление столонов и клубней и их израстание, а не переход от роста к цветению, как в работах его предшественников. Выращивая опытные растения при длинном дне и создавая условия короткого дня отдельным их зонам и стеблям, Разумов наблюдал образование клубней на подземных столонах даже в том случае, если короткий день получали только верхние части растений. В этом случае воздействие короткодневной зоны передавалось вниз к корням через зону, находившуюся при длинном дне. Однако и в этом случае нельзя видеть прямого доказательства нарушения актиноритмической локализации, так как, находясь на коротком дне, вершина растения оканчивала рост, а это приводит к клубнеобразованию. Не случайно Разумов, обсуждая вопрос об органе восприятия актиноритмических воздействий у растений, высказался так же, как и Нотт, в пользу точки роста, не учтя того, что, помимо точек роста, в закрываемой зоне находилось несколько вполне развитых листьев.
Таким образом, в 1931 г. господствовало представление о растущей зоне, или точке роста, как об основном органе, воспринимающем актиноритмические условия.
В тот период объектом наших опытов были только древесные виды. Это обстоятельство и послужило одной из причин постановки исследований, цель которых — выяснить роль листьев при локальных актиноритмических воздействиях. У травянистых однолетних растений, особенно у однодольных, различие между вершиной стебля и прилегающими к ней листьями разграничено не так резко, как у древесных видов. После окончания роста ствол и ветви последних быстро древеснеют и покрываются пробкой. Поэтому трудно предположить, что они как-то активно могут реагировать на актиноритмический фактор. Иное дело — листья. Прежде всего с их фотосинтетической деятельностью связано накопление углеводов, и в частности сахаров, способствующих, по мнению многих авторов, повышению морозоустойчивости растений (Н. А. Максимов, Н. И. Туманов и др.).
В нашем первом актиноритмическом опыте, проводимом в Ленинграде, было обнаружено, что воздействие коротким днем на акацию белую (Robinia pseudoacacia) приводит к весьма значительному накоплению ею крахмала. Поэтому, ставя в 1931 г. опыты по воздействию коротким днем на отдельные зоны саженцев акации белой, мы включили в них варианты, выясняющие роль листьев в реагировании растений на актиноритмические локальные условия.
Весной 1931 г. сеянцы акации белой, зимовавшие в прикопанном виде в подвале, были высажены на гряды. После того как они достигли высоты 40—50 см, их подвергли локальному актиноритмическому воздействию, которое началось в первых числах июля и продолжалось в течение 30 сут. Чтобы создать короткий 12-часовой день для отдельных частей растений, последние закрывали на 12 ч в сутки картонными и матерчатыми муфтами и колпаками, черными внутри и белыми снаружи. Повторность каждого варианта — десятикратная. Контролем служили саженцы, находящиеся в условиях естественного дня. Они полностью вымерзли зимой 1931/32 гг.
В первой опытной группе коротким днем действовали на вершины саженцев длиной 15 см со всеми расположенными на них листьями. Рост таких вершин скоро прекратился, весной следующего года все 10 растений этого варианта оказались почти не поврежденными морозами. У них сохранились даже верхушечные почки. Только у отдельных экземпляров отмерзли концы нижних ветвей.
Вариант 2-й отличался от 1-го лишь тем, что в закрываемой зоне растений, находящейся при коротком дне, обрывали все листья, кроме еще не распустившихся. Все 10 акаций этой группы вымерзли.
В 3-м варианте коротким днем действовали на нижние половины стеблей высотой 25 см со всеми находящимися на них листьями. Верхние части саженцев в данном случае находились в условиях длинного дня и росли так же, как контрольные.
Рост саженцев не был закончен до наступления холодов. Весной установили, что перезимовали только нижние части растений, испытавшие воздействие короткого дня. Все, что было выше закрываемой зоны, отмёрзло.
Наконец, в 4-м варианте коротким днем воздействовали, как и в 3-м, на нижние части стволов акации, но у саженцев этой группы удалили все листья. Следствием этого было вымерзание всех 10 растений данного варианта.
На основании результатов опыта, поставленного с целью выяснить роль листьев в актиноритмической реакции растений, тогда же были сделаны следующие выводы:
1) физиологические процессы, обусловливающие морозоустойчивость многолетних видов, связаны с актиноритмической реакцией листьев;
2) основным, а у древесных пород — единственным органом, воспринимающим актиноритмические условия, являются листья, потому что воздействие коротким днем на любую зону без листьев не дает положительных результатов.
В 1932 г. подобные опыты были поставлены уже не только с акацией белой, но и с другими вымерзающими в Ленинграде видами: айвой, абрикосом и некоторыми сортами малины. Здесь, как и ранее, обрыв листьев в зоне, находящейся в условиях короткого дня, всегда приводил к отсутствию актиноритмического эффекта, выражающегося в изменении зимостойкости растений.
Повторение результатов 1931 г. служило полным доказательством значения листьев в актиноритмической реакции растений, и мы имели все основания отметить данное обстоятельство в работе, посвященной этим исследованиям.
Таким образом, мы можем утверждать, что первые опыты, доказавшие роль листьев в актиноритмической реакции растений, были проведены нами в работе 1931 г., посвященной локальным актиноритмическим воздействиям на акацию белую. Результаты ее были подготовлены к печати в 1933 г. и опубликованы в 1935 г.
В то же время был проведен повторный опыт с акацией белой. В опыте было три основных группы растений. В первой группе растения все время находились на длинном дне (не короче 17 ч), во второй группе — полностью на коротком 10-часовом дне. Растения третьей группы были в тех же световых условиях, что и растения первой группы, т. е. находились на длинном дне, но на все листья по мере их распускания воздействовали коротким (10-часовым) днем. Для этого каждый лист заключали в светонепроницаемый пакет на 14 ч в сутки. Повторность в каждой группе — 10—12 растений. В течение всего опыта проводили биометрические наблюдения 2 раза в декаду.
При длинном дне рост акации белой не оканчивается и она дает значительные приросты. В описываемом опыте не было ни одного сеянца, который бы в условиях короткого дня превысил 5 см, тогда как при длинном дне в 60-дневном возрасте акация белая достигала в отдельных случаях 50 см.
Высота сеянцев третьей группы, где короткий день был создан только для листьев, а сами они находились на длинном дне, была такой же незначительной, как и сеянцев, росших при коротком дне.
Уже этого факта достаточно, чтобы считать доказанным, что и актиноритмическая реакция, выражающаяся ростом, является полностью листовой. Точки роста и в данном случае были совершенно пассивны.
Различие в приростах акации белой при длинном и коротких днях (причем только за счет надсемядольного стебля) наблюдались уже с первых дней опыта.
Вытягивание подсемядольного колена проходило одинаково во всех вариантах опыта, что, видимо, не было связано с актиноритмической реакцией листьев. Это и понятно, так как к моменту распускания первого листа прирост подсемядольных колен уже был окончен. Очевидно, ростовые процессы гипокотилей обусловливаются прежде всего запасами семян. Поэтому при более точном учете приростов главного стебля в высоту в зависимости от актиноритмов не следует принимать во внимание длину подсемядольного колена. Сделав это в разбираемом примере, мы получим следующие средние высоты двухмесячных растений акации; при длинном дне 47,4 см, или 100%, при коротком дне 2,3 см, или около 5%, при коротком дне только для листьев — 2,8 см, или около 6 %.
Как мы и ожидали, укорачивание длины дней для одних только листьев акации привело ее к тому же физиологическому состоянию, как если бы все растение находилось при коротком дне. В условиях непрерывного освещения или длинного дня (не короче 17—18 ч) рост акации белой продолжается и она становится вечновегетирующей и вечнозеленой. При коротком дне, наоборот, рост ее заканчивается очень быстро, в данном эксперименте при 10-часовом дне он завершается за 12—15 сут. За этот же срок окончился рост и сеянцев, у которых при 10-часовом дне были одни листья. Следовательно, продолжительность сезонного периода роста растений — это актиноритмическая реакция листьев.
Вслед за окончанием роста в условиях коротких дней у акации белой сравнительно быстро (через 15—30 сут) начинается листопад. Так было и на этот раз, причем наблюдалась полная одновременность сбрасывания листьев как сеянцами, находившимися на коротком дне, так и теми, у которых на коротком дне были только листья.
Листья акаций, росших при длинном дне, имели значительно большие размеры, чем листья у растений, на которые воздействовали коротким днем. Так же сильно они отличались и по форме. Сложный непарноперистый лист акации белой при длинном дне состоял из восьми пар листочков, а у сеянцев, получавших короткий день, всего из одной пары, что характерно для первого листа всхода. Точно такие же однопарные листочки имели и сеянцы акации, выращенные при длинном дне, но у которых листья были при 10-часовом дне.
Следовательно, форма и размеры листьев зависят от актиноритмических условий, в которых они находились во время формирования.
Ранее говорилось, что впервые роль листьев в актиноритмической реакции растений была выявлена на примере изменения морозоустойчивости древесных видов. Однако тогда актиноритмическое воздействие проводили не на отдельные листья, а на целые зоны растения, на которых в одних вариантах листья оставляли, а в других обрывали. Желая устранить малейшие сомнения в правильности наших первоначальных выводов, мы использовали с этой целью только что приведенный опыт с акацией белой. Для этого сеянцы акации, выращенные при длинном дне, при коротком 10-часовом дне и при длинном дне, но с листьями, для которых был создан короткий день, подвергли замораживанию в холодильном шкафу при температуре —18 °С. Перед замораживанием растения прошли температурную закалку, для чего находились в течение 10 сут при 2—5°С. После замораживания сеянцы акации выдерживали в течение 3 сут снова при температуре 2—5°С, а затем их помещали в теплую оранжерею для отрастания.
В результате замораживания все растения акации, росшие при длинном дне, полностью вымерзли вместе с корнями. Растения, выращенные при коротком дне, совершенно не пострадали от низкой температуры и начали рост из верхушечных почек.
Растения, вегетировавшие при длинном дне, но с листьями, для которых был создан короткий 10-часовой день, оказались такими же морозоустойчивыми, как и сеянцы, росшие при коротком дне.
Таким образом, было получено еще одно доказательство значения актиноритмической реакции листьев для прохождения процессов, определяющих максимальную морозоустойчивость растений.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
| Все цветоводы-любители знают, что некоторые цветы размножаются луковицами, клубнями, корневищем, клубнелуковицами, корнеклубнями. А вот чем различаются между собой луковица и клубнелуковица, клубень и корнеклубень?  Корневище — это горизонтально растущий подземный многолетний побег с остатками отмерших листьев, почками в их пазухах и на конце побега, и придаточными корнями. В корневище обычно откладываются запасные питательные вещества, и оно служит для вегетативного размножения растения. При искусственном размножении его обычно делят после цветения. В это время корневище находится в состоянии готовности к дальнейшему росту и формированию новых корней. Корневища могут расти двумя способами. У ириса германского верхушечная почка развивается в цветонос, а рост корневища в горизонтальной плоскости осуществляется за счет боковой почки. В следующий сезон у этого образовавшегося бокового побега формируется собственная верхушечная почка, образующая цветонос, а растение продолжает горизонтально расти, закладывая новые боковые почки. В другом случае, как, например, у мяты или всеми нелюбимого пырея, рост корневища осуществляется за счет продолжительного функционирования верхушечной и иногда боковых почек, которые обычно дают цветоносные побеги. Корневища разных растений отличаются и скоростью роста: например, корневище спаржи дает очень незначительный ежегодный прирост, а корневища мяты и пырея быстро и долго растут, благодаря чему за короткое время побеги занимают большую площадь. Корневищами размножаются: бегония королевская, белокрыльник, ирис, купена, ландыш майский, мята, пион, ревень, спаржа. Клубень — это однолетний подземный побег растения с утолщенным стеблем, часто имеющим сферическую форму, и зачаточными листьями, из пазушных почек которых на следующий год вырастают новые побеги. Настоящий клубень образуют не очень много растений. Наиболее известное — картофель. Всем известные «глазки» — это собрание почек в пазухе листового рубца. Они соответствуют узлам обычного стебля и располагаются по спирали последовательно или один против другого, начиная с верхушечной почки, которая расположена в той части клубня, которая противоположна месту его прикрепления к материнскому растению. Растения, образующие клубни: артишок, земляная груша, календула, картофель, кувшинка, клубненосная настурция, стахис, ямс. Клубнелуковица — это подземный утолщенный стебель, который служит запасным хранилищем питательных веществ. На ней размещаются превратившиеся в тонкие сухие пленки листья, закрывающие ее и предохраняющие от повреждений и подсыхания. В пазухе каждого листа находится почка, верхушка стебля развивается в цветоносный побег, корни вырастают из основания клубнелуковицы, часто имеющей выгнутую форму. У некоторых клубнелуковичных растений в верхней части клубнелуковицы формируется несколько цветочных почек. Ежегодно у основания каждого стебля образуется новая дочерняя клубнелуковица, а между новой клубнелуковицей и старой формируются вторичные побеги — мелкие дочерние клубнелуковички (клубнепочки). Число их зависит от вида.  Клубнелуковицы гладиолусов. Наиболее известные растения, образующие клубнелуковицы: ацидантера, гладиолус, иксия, крокосмия, шафран (крокус). Луковица — это видоизмененный, обычно подземный побег растений с коротким плоским стеблем (донцем) и мясистыми бесцветными листьями (чешуями), приспособленными для накопления питательных веществ. Существует два типа луковиц: пленчатые и черепитчатые, которые отличаются по способу образования чешуй. 1. Пленчатые луковицы (у нарцисса, тюльпана) имеют сочные утолщенные чешуи, которые представляют собой пленчатые влагалища листьев, прикрывающие друг друга и формирующие вокруг точки роста почти сомкнутые концентрические круги. У каждого чешуевидного листа образуется пазушная почка. Внешние чешуи сухие, благодаря чему они предохраняют луковицу от повреждений и высыхания. Корни у них отрастают в самом начале, сразу после посадки, они появляются на донце у основания нижней чешуи.  Луковицы тюльпанов после выкапывания. 2. Черепитчатые луковицы (у рябчика, лилии) имеют сочные, но часто слишком узкие чешуи, не охватывающие друг друга и не образующие общего покрова, из-за него такие луковицы подсыхают намного быстрее, чем пленчатые. Корни образуются в середине лета или позже и сохраняются до следующего года. У некоторых видов лилий (Тигровая, Красная, Саржента) в пазухах листьев на стебле формируются мелкие, так называемые воздушные луковички. Корневище, луковица, клубень, клубнелуковица — это измененные стебли, и к ним же относятся усы и отпрыски. Отпрыск — боковой побег, развивающийся из пазухи листа, расположенного в районе корневой шейки растения. Обычно побег имеет только одну почку, развивающуюся в молоденькое растение с собственной корневой системой и точкой роста (очиток). Ус также вырастает из пазухи листа на корневой шейке. Растет он горизонтально и в некоторых его узлах из боковых почек образуются молодые растения. После укоренения они формируют цветочные почки и новые побеги. Усы образуются у земляники, костяники, хлорофитума, камнеломки, лапчатки. Корнеклубень — это разросшийся в толщину корень, на котором расположены почки возобновления. Сами по себе корнеклубни служат для накопления запасов питательных веществ, служащих базой для роста почек. Есть два основных типа корнеклубней: одногодичные, развивающиеся каждый год заново, как у георгин, и нарастающие ежегодно, как у бегоний. Одногодичные корнеклубни образуются при разрастании боковых корней у корневой шейки. Во время вегетационного периода некоторые из них развиваются как вместилища запасных питательных веществ. Каждый год за счет этих запасов у растений развивается новая надземная часть, корни после этого отмирают. Развитие многолетних корнеклубней протекает проще. Обычно молодой корень растения уже с самого начала превращается в запасающий орган, увеличиваясь в размерах по мере поступления питательных веществ. |
gardenmix.ru
Маркаров, Александр Макарович - Морфофизиология клубнеобразующих растений [Текст]
Поиск по определенным полям
Чтобы сузить результаты поисковой выдачи, можно уточнить запрос, указав поля, по которым производить поиск. Список полей представлен выше. Например:author:иванов
Можно искать по нескольким полям одновременно:author:иванов title:исследование
Логически операторы
По умолчанию используется оператор AND. Оператор AND означает, что документ должен соответствовать всем элементам в группе:исследование разработка
author:иванов title:разработка
оператор OR означает, что документ должен соответствовать одному из значений в группе:исследование OR разработка
author:иванов OR title:разработка
оператор NOT исключает документы, содержащие данный элемент:исследование NOT разработка
author:иванов NOT title:разработка
Тип поиска
При написании запроса можно указывать способ, по которому фраза будет искаться. Поддерживается четыре метода: поиск с учетом морфологии, без морфологии, поиск префикса, поиск фразы. По-умолчанию, поиск производится с учетом морфологии. Для поиска без морфологии, перед словами в фразе достаточно поставить знак "доллар":$исследование $развития
Для поиска префикса нужно поставить звездочку после запроса:исследование*
Для поиска фразы нужно заключить запрос в двойные кавычки:"исследование и разработка"
Поиск по синонимам
Для включения в результаты поиска синонимов слова нужно поставить решётку "#" перед словом или перед выражением в скобках. В применении к одному слову для него будет найдено до трёх синонимов. В применении к выражению в скобках к каждому слову будет добавлен синоним, если он был найден. Не сочетается с поиском без морфологии, поиском по префиксу или поиском по фразе.#исследование
Группировка
Для того, чтобы сгруппировать поисковые фразы нужно использовать скобки. Это позволяет управлять булевой логикой запроса. Например, нужно составить запрос: найти документы у которых автор Иванов или Петров, и заглавие содержит слова исследование или разработка:author:(иванов OR петров) title:(исследование OR разработка)
Приблизительный поиск слова
Для приблизительного поиска нужно поставить тильду "~" в конце слова из фразы. Например:бром~
При поиске будут найдены такие слова, как "бром", "ром", "пром" и т.д. Можно дополнительно указать максимальное количество возможных правок: 0, 1 или 2. Например:бром~1
По умолчанию допускается 2 правки.Критерий близости
Для поиска по критерию близости, нужно поставить тильду "~" в конце фразы. Например, для того, чтобы найти документы со словами исследование и разработка в пределах 2 слов, используйте следующий запрос:"исследование разработка"~2
Релевантность выражений
Для изменения релевантности отдельных выражений в поиске используйте знак "^" в конце выражения, после чего укажите уровень релевантности этого выражения по отношению к остальным. Чем выше уровень, тем более релевантно данное выражение. Например, в данном выражении слово "исследование" в четыре раза релевантнее слова "разработка":исследование^4 разработка
По умолчанию, уровень равен 1. Допустимые значения - положительное вещественное число.Поиск в интервале
Для указания интервала, в котором должно находиться значение какого-то поля, следует указать в скобках граничные значения, разделенные оператором TO. Будет произведена лексикографическая сортировка.author:[Иванов TO Петров]
Будут возвращены результаты с автором, начиная от Иванова и заканчивая Петровым, Иванов и Петров будут включены в результат.author:{Иванов TO Петров}
Такой запрос вернёт результаты с автором, начиная от Иванова и заканчивая Петровым, но Иванов и Петров не будут включены в результат. Для того, чтобы включить значение в интервал, используйте квадратные скобки. Для исключения значения используйте фигурные скобки.search.rsl.ru