
Химизм дыхания растений. Дыхания растения
Химизм дыхания растений
Химизм дыхания растений это выражение дыхательного процесса через ряд последовательных превращений и химических реакций.
 Химизм дыхания растений
Химизм дыхания растенийХимическая реакция окисления
Окислением называется химическая реакция, при которой происходит присоединение кислорода к окисляемому веществу (например, окисление водорода до воды: 2Н2 + О2 —> 2Н2О), или отнятие водорода от окисляемого вещества, или отнятие электрона от окисляемого вещества, вследствие чего повышается его валентность.
Вещество, принимающее электрон, — акцептор электрона — в данном случае является окислителем, например Fе++ — е + A —>Fе+++ + Aе. Акцептор электрона, обозначенный буквой А, приобретая электрон (е), восстанавливается, а железо, отдав электрон, окисляется. Вещество А может передать полученный электрон другому акцептору, восстановив его.
При окислении большую роль играет присоединение молекулы воды к окисляемому материалу с последующим отнятием водорода. К этому же типу окисления относится и окисление с предварительным присоединением к окисляемому веществу молекулы фосфорной кислоты с последующим отнятием водорода.
Представление о химизме дыхания
Представление о химизме дыхания создано на основе работ А. Н. Баха, В. И. Палладина, Д. Кейлина, О. Варбурга, В. А. Энгельгардта, Д. М. Михлина, А. И. Опарина и других ученых. В процессе дыхания различают 2 фазы: анаэробную и аэробную.
Начальная фаза превращения сахара — анаэробный распад — осуществляется одинаково как при дыхании, так и при брожениях. Через ряд последовательных превращений при распаде молекулы сахара образуется пировиноградная кислота, после чего дальнейшие превращения идут разными путями в зависимости от наличия в организме ферментативных систем и внешних условий.
В превращениях органических веществ в дыхательном процессе огромную роль играют фосфорные соединения. Во время анаэробной фазы к молекуле глюкозы при помощи специального фермента присоединяется один остаток фосфорной кислоты от АТФ.
Глюкоза +АТФ -> глюкоза-фосфат + АДФ
Далее глюкоза-фосфат претерпевает ряд сложных превращений, которые происходят под действием ферментов. В ходе этих превращений используется неорганический фосфор фосфорной кислоты и образуется дифосфоглицериновая кислота, которая имеет одну макроэргическую связь.
Фосфоглицериновын альдегид + Н2PO4 —> окисление дифосфоглицериновыи альдегид —> дифосфоглицериновая кислота
Образовавшаяся дифосфоглицериновая кислота реагирует с АДФ, перенося на нее макроэргическую связь, в результате чего и образуется АТФ и фосфоглицериновая кислота.
Дифосфоглицериновая кислота + АДФ —> АТФ + фосфоглицериновая кислота
В дальнейшем после сложных превращений фосфоглицериновая кислота образует пировиноградную кислоту, а остаток фосфорной кислоты вновь дает с молекулой АДФ молекулу АТФ.
Фосфоглицериновая кислота + АДФ —> АТФ + пировинограднан кислота
Следует подчеркнуть то, что на схемах даны только конечные результаты процессов, которые в действительности идут через ряд сложных промежуточных процессов. Более детально процесс изображен на рисунке.
 Схема анаэробного дыхания растений
Схема анаэробного дыхания растенийАнаэробная фаза дыхания заканчивается образованием двух молекул пировиноградной кислоты (СН3СОСООН), а весь цикл превращения сахара до пировиноградной кислоты называется гликолизом.
В результате превращений сахара в первой фазе дыхания образуется АТФ, в которой накапливается энергия. Эта энергия может быть мобилизована клеткой для любых процессов жизнедеятельности.
Вторая фаза дыхания является аэробной и начинается с превращения пировиноградной кислоты до конечных продуктов углекислого газа и воды. Это превращение связано с участием целого комплекса разных ферментативных систем и образованием ряда органических кислот (уксусной, щавелевоуксусной, лимонной, щавелево-янтарной, кетоглутаровой и др.).
В результате постепенного образования органических кислот весь углерод и водород пировиноградной кислоты окисляется. Этот цикл окисления пировиноградной кислоты был исследован Кребсом и получил название цикла Кребса.
В результате окисления пировиноградной кислоты получаются 3 молекулы углекислого газа, а так как из молекулы сахара получаются 2 молекулы пировиноградной кислоты, то общий выход углекислого газа будет равняться 6 молекулам, что и указывается в суммарном уравнении дыхания.
В процессе окисления принимают участие различные оксидазы. В зависимости от состояния растения, его вида и условий внешней среды может включаться та или иная ферментативная система.
В процессе окисления молекулы глюкозы до СО2 и Н2О образуется 38 молекул АТФ (2 в первую и 36 во вторую фазу дыхания). Благодаря этому в клетке сохраняется 380 000 кал, что составляет 50—55% химической энергии, заключенной в глюкозе. Остальная энергия рассеивается в виде тепла.
libtime.ru
Дыхание растений | Биология. Реферат, доклад, сообщение, краткое содержание, лекция, шпаргалка, конспект, ГДЗ, тест
Представляя собой одну из сторон обмена, а именно процесс диссимиляции, или распада, дыхание играет огромную роль в жизни растения, так как оно является источником энергии для всех протекающих в организме биохимических процессов.
Образованная в процессе дыхания химическая энергия претерпевает различные превращения, обусловливая протекание ряда процессов, связанных с превращением веществ, ростом и движением растения. Часть ее выделяется в виде тепловой энергии (рис. 198). Интересно, что в молодых растениях, когда особенно интенсивно идут процессы синтеза конституционных веществ плазмы, выделение тепловой энергии происходит в значительно меньшем количестве, чем во взрослых растениях. Последнее хорошо видно из приводимой ниже таблицы.
|
День прорастания семян пшеницы |
Энергия дыхания, вычисленная по выделившейся CO2 на 1 кг при 25° (в кал) |
Энергия, фактически выделенная в виде тепла на 1 кг при 25° (в кал) |
|
2-й |
2135 |
363 |
|
3-й |
3802 |
540 |
|
4-й |
6277 |
2938 |
|
5-й |
6886 |
3216 |
|
6-й |
8837 |
4341 |
 |
| Рис. 198. Выделение тепла при дыхании семян |
Если на 2-й день в виде тепла выделяется 12% энергии, то уже на 6-й день ее выделяется почти 50%.
Дыхание есть прежде всего физиологический, а не просто химический процесс. В этом процессе принимает активное участие целый комплекс окислительно-восстановительных ферментов. Дыхание нельзя рассматривать только как процесс распада, так как при дыхании образуется ряд веществ, богатых энергией. При дыхании химическая энергия окисления тратится на превращение веществ, рост и движение растения, и только часть ее выделяется в виде тепла, не используемого растением.
Основное уравнение (формула) дыхания обратно процессу фотосинтеза:
C6h22O6 + 6O2 = 6CO2 + 6h3O + 674 ккал.
 |
| Рис. 199. Респирометр Варбурга для изучения дыхания растений: слева — вид спереди; справа — вид сбоку; А — приемник для дышащих частей; Б — манометр; В — сосудик для КОН; Г — нажимной кран; Д — резиновый резервуар для манометрической жидкости; Е — расширение для помещения HCl |
Окисляясь кислородом воздуха, органическое вещество, в данном случае сахар, превращается в углекислый газ и воду. Калорийность процесса равняется 674 ккал. Однако эта суммарная формула говорит о начальных и конечных звеньях процесса превращения молекулы сахара при дыхании. Окисляясь, энергетический материал проходит ряд ступеней, которые лишь постепенно приводят его к таким простым веществам, как углекислота и вода. Материал с сайта http://worldofschool.ru
Обратимся к методам изучения дыхания. Так же как и методы изучения фотосинтеза, методы изучения дыхания могут быть направлены на изучение потери сухого веса или на количество выделенной углекислоты или поглощенного кислорода. В ряде случаев приходится учитывать и поглощенный кислород, и выделенную углекислоту, например при изучении дыхательного коэффициента, т. е. отношения CO2 / O2.
Потеря веса растением при дыхании измеряется по методу половинок. Для учета выделения углекислого газа при дыхании пользуются респирометром Варбурга (рис. 199). В приемник А помещают растение, а в сосудик В — раствор едкой щелочи для поглощения выделяемого углекислого газа. К сосуду А пришлифован манометр Б, соединенный с резиновым резервуаром. При помощи крана Г жидкость в манометре устанавливается на нужном уровне. По уменьшению объема газа в приборе вычисляют количество поглощенного кислорода. Вводя в конце опыта через кран Е соляную кислоту, определяют количество выделенного CO2.
 На этой странице материал по темам:
На этой странице материал по темам: Строение респирометр
Дыхание семян краткое содержание
Методы изучения дыхания растений
Дыхание растений 6 класс биология доклад
Дыхание растений доклад
worldofschool.ru
Дыхание растений
Залог нашего существования – дыхание. То же самое можно сказать и о других организмах. То есть растения умудряются одновременно и выделять кислород, и поглощать его. Это совершенно удивительная способность. При этом растения еще и ухитряются что-то оставить и нам с вами. На этом уроке мы узнаем, как дышат растения и как они делятся своим кислородом с окружающими их организмами.
Общие сведения
Процесс дыхания (см. Рис. 1, 2) – получение энергии при распаде сложных органических соединений.
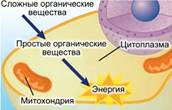
Рис. 1. Дыхание растений (первый этап)
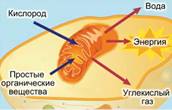
Рис. 2. Дыхание растений (второй этап)
Процесс дыхания
При сгорании дерева выделяется энергия, запасенная в ходе фотосинтеза, в виде тепла и света. Горение – химическая реакция, в ходе которой органические соединения взаимодействуют с кислородом и распадаются до воды и углекислого газа.
По сути, дыхание сходно с горением.
Дыхание растений (см. Рис. 3) – соединение сложных органических веществ с кислородом, в ходе которого происходит постепенное выделение энергии. Также в результате дыхания выделяются углекислый газ и вода.
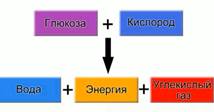
Рис. 3. Схема дыхания растений
В ходе дыхания энергия запасается в виде специальных энергетических веществ – соединений АТФ (см. Рис. 4).
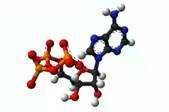
Рис. 4. АТФ
АТФ – универсальный носитель энергии, использующийся при любых процессах жизнедеятельности растения (рост, размножение).
Дыхательные органы растений
Водоросли поглощают кислород всей поверхностью тела, высшие растения – через устьица (см. Рис. 5).
Устьица – отверстия в коже листа растения, ограниченные замыкающими клетками.
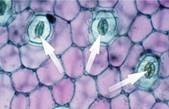
Рис. 5. Устьица
Кислород поступает к каждой клетки по сети межклетников (см. Рис. 6).
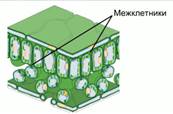
Рис. 6. Межклетники
Межклетники – пространства, возникающие в тканях растений при разъединении, разрушении или отмирании соседних клеток. Соединяясь друг с другом, межклетники образуют в растении систему полостей и ходов, сообщающихся с внешней средой (атмосферой) через устьица.
Опыт: взять 4 бутылки из прозрачного бесцветного стекла. В одну из них положить прорастающие набухшие семена гороха или фасоли. В другую положить такое же количество сухих семян. Несколько небольших корнеплодов моркови выдержать в теплой воде 2-3 дня, для активизации их клеток. Положить их в третью бутыль. В четвертую положить свежесрезанные стебли растений с листьями. Плотно закрыть бутылки и поставить в теплое темное место. На следующий день поднести в каждую бутылку зажженную лучинку. Во всех бутылках, кроме той, где были сухие семена, лучинка гаснет. Это говорит о том, что растения потребили кислород в ходе дыхания, выделив углекислый газ. А клетки сухих семян находятся состоянии покоя, поэтому не потребляют кислород.
Дыхание корней
Растения на тяжелых глинистых или заболоченных почвах сильно страдают от недостатка кислорода. Вода перекрывает доступ воздуха к корням. Поэтому у таких растений развиваются дыхательные корни.
При выращивании сельскохозяйственных растений необходимо следить за доступом кислорода к корням. В этих целях почву разрыхляют.
Взаимосвязь процессов дыхания и фотосинтеза
На свету у растений протекают 2 взаимосвязанных процесса – дыхание и фотосинтез (см. Рис. 7).
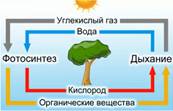
Рис. 7. Газообмен растений
При дыхании растение выделяет углекислый газ и поглощает кислород, а при фотосинтезе – поглощает углекислый газ и выделяет кислород.
При дыхании растение потребляет меньше кислорода, чем выделяет при фотосинтезе. При фотосинтезе растение потребляет меньше углекислого газа, чем может выделить при дыхании.
Дыхание происходит непрерывно во всех органах растения. Оно погибает без кислорода.
Рыхление почвы
Рыхление почвы необходимо для обеспечения достаточного поступления воздуха к корням и сохранения в почве влаги. При подсыхании почвы на ней образуется корка, которая испаряет имеющуюся влагу за счет капиллярного эффекта – свойства воды хорошо подниматься по тонким трубочкам. Также корка препятствует проникновению воды внутрь почвы.
Во время рыхления корка разрушается, и влага в поверхностном слое сохраняется дольше.
vipusk-2023-chehov7.edumsko.ru
Дыхание растений
Живая клетка представляет собой открытую энергетическую систему, она живёт и сохраняет свою индивидуальность за счет постоянного притока энергии. Как только этот приток прекращается, наступает дезорганизация и смерть организма. Энергия солнечного света, запасенная при фотосинтезе в органическом веществе, вновь высвобождается и используется на самые различные процессы жизнедеятельности. Энергетический цикл жизни схематично представлен рисунке.

Как видно, энергия квантов света, аккумулированная в углеводах, вновь высвобождается в процессе их распада (диссимиляции). В самой общей форме можно отметить, что все живые клетки получают энергию за счет ферментата в иных реакций, в ходе которых электроны переходят с более высокого энергетического уровня на более низкий. В природе существуют два основных процесса, в ходе которых энергия солнечного света, запасенная в органическом веществе, высвобождается,— это дыхание и брожение. Дыхание — это аэробный окислительный распад органических соединений на простые неорганические, сопровождаемый выделением энергии. Брожение — анаэробный процесс распада органических соединений на более простые, сопровождаемый выделением энергии. В случае дыхания акцептором электрона служит кислород, в случае брожения — органические соединения. Процессы, входящие в энергетический цикл, имеют настолько важное значение, что в XX в. возникла наука биоэнергетика, изучающая молекулярные и субмолекулярные основы трансформации энергии.
В этом разделе:
- Значение дыхания в жизни растения
- Аденозинтрифосфат. Структура и функции
- Окислительно-восстановительные процессы. работы А.Н. Баха и В.И. Палладина
- Субстраты дыхания
Пути дыхательного обмена:
- Гликолитический путь дыхательного обмена:
- Анаэробная фаза дыхания (гликолиз)
- Аэробная фаза дыхания
- Энергетический баланс процесса дыхания
- Взаимосвязь процессов дыхания и брожения
- Пентозофосфатный путь дыхательного обмена
Влияние внешних и внутренних факторов на интенсивность дыхания:
- Влияние внешних условий на процесс дыхания
- Влияние внутренних факторов на процесс дыхания
Пути регуляции дыхательного обмена:
- Локализация в клетке реакций дыхательного обмена
- Регуляция дыхательного обмена
- Взаимосвязь дыхания с другими процессами обмена
fizrast.ru
История учения о дыхании растений
Научные основы учения о роли кислорода в дыхании были заложены трудами А.Л.Лавуазье. В 1774 г. кислород независимо открыли Пристли и Шееле, а Лавуазье дал название этому элементу. Изучая одновременно процесс дыхания животных и горение, Лавувзье в 1773-1783 гг. пришел к выводу, что при дыхании, как и при горении, поглощается кислород и образуется углекислый газ, причем в том и другом случаях выделяется тепло. На основании своих опытов он заключил, что процесс горения состоит в присоединении кислорода к субстрату и что дыхание есть медленно текущее горение питательных веществ в живом организме.
 Я.Ингенхауз в 1778-1780 гг. показал, что зеленые растения в темноте, а незеленые части растений и в темноте, и на свету поглощают кислород и выделяют углекислый газ.
Я.Ингенхауз в 1778-1780 гг. показал, что зеленые растения в темноте, а незеленые части растений и в темноте, и на свету поглощают кислород и выделяют углекислый газ. В своей работе, опубликованной в 1779 г. он писал:
«Когда солнце, поднявшееся над горизонтом, разбудит своими лучами заснувшие за ночь растения, оно сделает их способными исполнять свою целительную функцию – исправлять воздух для животных; во мраке ночи эта деятельность совсем прекращается; днем же совершается с тем большим оживлением, чем светлее день и чем выгоднее расположено растение в отношении солнечных лучей. Затененные высокими зданиями или другими растениями, они не исправляют воздух, а, наоборот, выделяют вредный для дыхания животных воздух. К концу дня выработка очищенного воздуха ослабевает и при заходе солнца совершенно прекращается».
Первые точные исследования процесса дыхания у растений принадлежат Соссюру (1804). Он брал свежие листья и помещал их на ночь в сосуд, наполненный воздухом. При этом кислород воздуха поглощался и выделялся углекислый газ. Если на следующий день листья снова выставлялись на солнечный свет, то они выделяли почти такое же количество кислорода, какое поглотили ночью. Свои исследования Соссюр распространил и на незеленые части растений: стебли древесных растений, цветки, корни, плоды, и доказал, что дыхание наблюдается также в клетках этих органов. Он обнаружил, что при дыхании потеря в весе растения равна весу выделенного углерода.
Соссюр обратил внимание и на то, что молодые, растущие части растения, например новые побеги и распускающиеся цветки, дышат интенсивнее и потребляют кислорода больше, чем части растения, прекратившие рост.
Если, по Лавуазье, дыхание имеет сходство с процессом горения, то каким же образом органические вещества могут «гореть» при обычной температуре тела организма, да еще в водной среде, (ведь на 70 — 90% масса живых организмов состоит из воды)? Возникло предположение о том, что в живых клетках существуют механизмы, активирующие кислород. Швейцарский химик X. Ф. Шейнбайн, открывший озон, изучал причины быстрого потемнения пораненной поверхности растительных тканей, таких, как ткани яблок, картофеля, плодовых тел грибов. В 1845 г. он выступил со своей теорией окислительных процессов, согласно которой в живых клетках имеются соединения, способные легко окисляться в присутствии 02 и таким образом активировать молекулярный кислород. Если ткань прокипятить, то потемнения не происходит. Следовательно, потемнение тканей — каталитический окислительный процесс. Шейнбайн ошибочно полагал, что активация кислорода — это образование озона.
Исследования, начатые Шейнбайном, продолжил А. Н. Бах, который в 1897 г. разработал перекисную теорию биологического окисления, приложив ее к процессам дыхания. Несколько позже, в том же 1897 г., аналогичные взгляды высказал немецкий исследователь К. Энглер.
Суть перекисной теории биологического окисления Баха заключается в следующем. Молекулярный кислород имеет двойную связь и для того чтобы его активировать, необходимо эту двойную связь расщепить. Легко окисляющееся соединение А взаимодействует с кислородом и, разрывая двойную связь, образует пероксид А02 Таким образом, по мысли Баха, активация кислорода есть образование пероксида. В свою очередь пероксидное соединение, взаимодействуя с соединением В, окисляет его; затем эта реакция повторяется со вторым атомом кислорода и второй молекулой соединения В. Получается полностью восстановленное исходное соединение — акцептор кислорода А и полностью окисленное вещество В.
Много позднее, в 1955 г., две группы исследователей — О. Хаяиши с сотр. в Японии и Г. С. Мэзон с сотр. в США, используя современные методы, проанализировали возможность включения кислорода в органические соединения.
В настоящее время известно, что путь включения кислорода в органические соединения в соответствии с перекисной теорией биологического окисления Баха и Энглера не имеет отношения к дыханию, однако работы этих исследователей сыграли большую роль в изучении химизма дыхания, заложив основы современного понимания механизмов активации кислорода.
История современного учения о дыхании растений неразрывно связана с именем академика В.И. Палладина.
В годы первого петербургского периода работы Палладин исследовал ферментативную природу дыхательного процесса. Палладин показал, что и анаэробная, и аэробная фазы дыхания обеспечиваются специфическими ферментами, последовательно перерабатывающими продукты дыхания. Итоги работ этого периода изложены в монографии В.И. Палладина «Дыхание как сумма ферментативных процессов» (1907).
Одновременно с Палладиным проблемой дыхания занимались в целом ряде крупнейших научно-исследовательских институтов и лабораторий Западной Европы. Наибольшую популярность приобрели две новые школы – Виланда и Варбурга.
Т. Виланд развивал взгляды на роль дегидраз и водородных акцепторов, вполне аналогичные взглядам Палладина. Расхождение их теорий заключалось в том, что Виланд категорически отрицал какую бы то ни было роль оксидаз как специфических активаторов кислорода, считая молекулярный кислород способным самостоятельно отнимать водород от водородного акцептора. По мнению же Палладина, водородные акцепторы не могут самопроизвольно освобождаться от водорода, но требуют для этого участия оксидаз, которые поэтому являются обязательным фактором в реакции, выраженной во втором уравнении Палладина.
Противник Виланда, Варбург, считал, что молекулярный кислород не может вступить в организме в какой бы то ни было окислительный процесс, если в организме отсутствует система железоорганических соединений, типичным представителем которых он считал геминфермент. Варбург утверждал, что геминфермент активирует молекулярный кислород, т.е. как бы дает первый толчок к началу окислительных процессов, и без него никакой дыхательный процесс не может совершаться. Далее, по мнению Варбурга, окислительный импульс через промежуточные звенья (геминовые соединения) доходит до дыхательного субстрата и окисляет его. Резюмируя свои взгляды, Варбург утверждал, что дыхание осуществляется путем активации кислорода, а отнюдь не водорода. Но ведь Палладин как раз и говорил о той же необходимости активации молекулярного кислорода, защищая перед Виландом роль оксидаз в процессе дыхания.
Все различие в основных посылках Варбурга и Палладина заключается в том, что первый, работая по преимуществу с объектами животного происхождения, называл свой активатор молекулярного кислорода геминферментом, а Палладин, работавший с объектами растительного происхождения, сохранил за этим активатором ранее установившееся в науке название оксидазы. Но по существу оба говорили об одном и том же, протестуя против непримиримой позиции Виланда, отрицавшего необходимость энзиматической активации молекулярного кислорода.
Английский биохимик Д. Кейлин в 1925 г. доказал присутствие в клетках цитохромоксидазы, ускоряющей поглощение ими кислорода, и открыл другие цитохромы. Затем цитохромы были обнаружены у всех аэробов и было показано, что у этих организмов на завершающем этапе процесса дыхания осуществляется перенос на кислород электронов и протонов, в результате чего образуется Н2O (или Н2O2).
Значение дыхания в жизни растительного организма
Клеточное дыхание — это окислительный, с участием кислорода распад органических питательных веществ, сопровождающийся образованием химически активных метаболитов и освобождением энергии, которые используются клетками для процессов жизнедеятельности.
Суммарное уравнение процесса дыхания:
С6Н12О6 + 6O2 ► 6СO2 + 6Н2O + 2875 кДж/моль
Не вся энергия, высвобождаемая при дыхании, может быть использована в процессах жизнедеятельности. Используется организмом в основном та энергия, которая аккумулируется в АТФ. Синтезу АТФ во многих случаях предшествует образование разности электрических зарядов на мембране, что, в свою очередь, связано с разностью концентраций ионов водорода по разные стороны от мембраны. Согласно современным представлениям, е только АТФ, но и протонный градиент служат источником энергии для различных процессов жизнедеятельности клетки. Обе формы энергии могут быть использованы на процессы синтеза, процессы поступления, передвижения питательных веществ и воды, создание разности потенциалов между цитоплазмой и внешней средой. Энергия, не накопленная в протонном градиенте и АТФ, в основном рассеивается в виде тепла или света и является для растения бесполезной.
Значение дыхания в жизни растения.
Дыхание — один из центральных процессов обмена веществ растительного организма. Выделяющаяся при дыхании энергия тратится как на процессы роста, так и нa поддержание в активном состоянии уже закончивших рост органов растения. Вместе с тем значение дыхания не ограничивается тем, что это процесс, поставляющий энергию. Дыхание, подобно фотосинтезу, сложный окислительно_восстановительный процесc, идущий через ряд этапов. На его промежуточных стадиях образуются органические соединения, которые затем используются в различных метаболических реакциях. К промежуточным соединениям относят органические кислоты и пентозы образующиеся при разных путях дыхательного распада. Таким образом, процесс дыхания — источник многих метаболитов. Несмотря на то что процесс дыхания в суммарном виде противоположен фотосинтезу, в некоторых случаях они могут дополнять друг друга. Оба процесса являются поставщиками как энергетических эквивалентов (АТФ, НАДФ-Н), так и метаболитов. Как видно из суммарного уравнения, в процессе дыхания образуется также вода. Эта вода в крайних условиях обезвоживания может быть использована растением и предохранить его от гибели. В некоторых случаях, когда энергия дыхания выделяется в виде тепла, дыхание ведет к бесполезной потере сухого вещества. В этой связи при рассмотрении процесса дыхания надо помнить, что не всегда усиление процесса дыхания является полезным для растительного организма.
biofile.ru
Дыхание растений
Дыхание растенийДыхание растений— представляет процесс, соответствующий дыханию животных. Растение поглощает атмосферный кислород, а последний воздействует на органические соединения их тела таким образом, что в результате появляются вода и углекислота. Вода остается внутри растения, а углекислота выделяется в окружающую среду. При этом происходит уничтожение, трата органического вещества; следовательно, Д. прямо противоположно процессу ассимиляции углерода. До известной степени его можно уподобить окислению и горению вещества.Наиболее простой механизм обмена газами у водорослей, которые не имеют тканей и органов, а воздух непосредственно проникает в каждую клетку. У мхов, папоротников, голосеменных и покрытосеменных воздух проходит более сложный путь. Через устьица он поступает в межклетники, которые пронизывают все растения, а оттуда - в клетки.У наземных растений устьица, как правило, расположены на нижней стороне листа, а у живущих в воде - на верхней, так как нижней стороной он лежит на поверхности воды. Поступление воздуха в листья регулируется периодическим открыванием и закрыванием устьиц.
Внутрь стволов деревьев и кустарников, покрытых толстой пробкой или корой, воздух поступает через отверстия - чечевички. Хорошо видны чечевички у березы, они крупные (до 15 см) и имеют вид узких темных поперечных полосок.
У ряда болотных растений затруднено поступление воздуха в корни, так как в насыщенной влагой почве мало воздуха. У этих растений сформировались приспособления, обеспечивающие нормальный газообмен. Так, у некоторых растений образовались дыхательные корни, которые выступают над поверхностью воды, например у растений мангровых лесов.Процесс дыхания связан с непрерывным потреблением кислорода клетками и тканями растений и осуществляется при участии различных ферментов. Вначале сложные органические вещества (белки, жиры, углеводы) под действием ферментов распадаются на более простые, которые при участии кислорода расщепляются до конца, т.е. до образования углекислого газа и воды. При этом освобождается энергия, которая используется растением (а также любым живым организмом) на процессы жизнедеятельности: поглощение из почвы воды и минеральных веществ, их передвижение, рост, развитие, размножение.
В освобождении энергии, заключенной в органических веществах, состоит главное значение дыхания. По существу, при дыхании освобождается солнечная энергия, которую растение использовало в процессе фотосинтеза на образование органических веществ и таким путем запасло ее. В процессе дыхания окисление сложных органических веществ до углекислого газа и воды происходит постепенно и энергия освобождается небольшими порциями. Если бы энергия освобождалась вся сразу, тогда клетка сгорела бы.
Дыхание, подобно другим процессам жизнедеятельности, зависит от факторов среды: температуры, влажности, содержания кислорода, степени освещенности и др. Для протекания процессов дыхания требуются определенные температурные условия, причем они разные у каждого вида растений и его органов. У большинства растений для дыхания наиболее благоприятна температура 25 - 30°С. У некоторых видов растений дыхание происходит и при отрицательных температурах, хотя этот процесс протекает очень слабо. Например, почки лиственных и иглы хвойных деревьев дышат и при температуре - 20 - 25°С. У арктических растений даже при низких температурах интенсивность дыхания высокая.Наиболее интенсивно дышат молодые органы и ткани растений, находящиеся в состоянии активного роста. Цветение и плодоношение сопровождаются усилением дыхания развивающихся цветков и плодов, что связано с образованием новых органов и тканей, обладающих высоким уровнем обмена веществ.
Интенсивность дыхания растений зависит от содержания воды в клетках.
Чем меньше воды в клетках, тем слабее идет в них дыхание. Очень слабо дышат сухие семена. С увеличением влажности дыхание семян возрастает в сотни и тысячи раз. Это отрицательно сказывается на хранении семян, так как они сильно разогреваются и погибают. Повышение интенсивности дыхания имеет огромное биологические значение для прорастания семян, поскольку усиление дыхания сопровождается освобождением большого количества энергии, необходимой для роста и развития зародыша.
На дыхание растений влияет содержание кислорода в окружающей среде. Угнетение дыхания начинается при уменьшении содержания кислорода до 5%. Недостаток кислорода испытывают подземные органы (корни и корневища) растений, обитающих на заболоченных и глинистых почвах.
В растениеводстве применяются различные агротехнические приемы для улучшения дыхания корней. Так, проводят комплексную обработку посевов машинами, чтобы сократить число обработок и уменьшить уплотненность почвы. Специальными культиваторами почву рыхлят и таким путем улучшают доступ воздуха к корням, при этом срезают сорняки, подкармливают культурные растения. Сильно увлажненные земли осушают, создают дренаж.На дыхание растений влияет и свет, хотя дышат они днем и ночью, на свету и в темноте. Свет вызывает повышение температуры растения, отчего дыхание его усиливается. У светолюбивых растений дыхание более интенсивное, чем у теневыносливых.Изменения в окружающей среде, связанные с деятельностью человека, также воздействуют на дыхание растений. Отрицательно влияют на дыхание вредные примеси, пыль, выделяемые промышленными предприятиями.
Газообмен в легких и тканях
Содержание газов во вдыхаемом и выдыхаемом воздухе неодинаково. Во вдыхаемом воздухе содержится почти 21% кислорода, около 79% азота, примерно 0,03% углекислого газа, небольшое количество водяных паров и инертных газов. В выдыхаемом — 16% кислорода, 4% углекислого газа, увеличивается содержание паров, количество азота и инертных газов остается неизменным.

Газообмен в легких и тканях
Кровь, которая течет к легким от сердца (венозная), содержит мало кислорода и много углекислого газа; воздух в альвеолах, наоборот, содержит много кислорода и меньше углекислого газа. Вследствие этого через стенки альвеол и капилляров происходит двусторонняя диффузия —. кислород переходит в кровь, а углекислый газ поступает из крови в альвеолы. В крови кислород проникает в эритроциты и соединяется с гемоглобином. Кровь, насыщенная кислородом, становится артериальной и по легочным венам поступает в левое предсердие.
У человека обмен газами завершается в несколько секунд, пока кровь проходит через альвеолы легких. Это возможно благодаря огромной поверхности легких, сообщающейся с внешней средой. Общая поверхность альвеол составляет свыше 90 м3.
Обмен газов в тканях осуществляется в капиллярах. Через их тонкие стенки кислород поступает из крови в тканевую жидкость и затем в клетки, а углекислота из тканей переходит в кровь. Концентрация кислорода в крови больше, чем в клетках, поэтому он легко диффундирует в них.
Концентрация углекислого газа в тканях, где он собирается, выше, чем в крови. Поэтому он переходит в кровь, где связывается химическими соединениями плазмы и отчасти с гемоглобином, транспортируется кровью в легкие и выделяется в атмосферу.АТМОСФЕРА
газовая оболочка, окружающая небесное тело. Ее характеристики зависят от размера, массы, температуры, скорости вращения и химического состава данного небесного тела, а также определяются историей его формирования начиная с момента зарождения. Атмосфера Земли образована смесью газов, называемой воздухом. Ее основные составляющие - азот и кислород в соотношении приблизительно 4:1. На человека оказывает воздействие главным образом состояние нижних 15-25 км атмосферы, поскольку именно в этом нижнем слое сосредоточена основная масса воздуха. Наука, изучающая атмосферу, называется метеорологией, хотя предметом этой науки являются также погода и ее влияние на человека. Состояние верхних слоев атмосферы, расположенных на высотах от 60 до 300 и даже 1000 км от поверхности Земли, также изменяется. Здесь развиваются сильные ветры, штормы и проявляются такие удивительные электрические явления, как полярные сияния. Многие из перечисленных феноменов связаны с потоками солнечной радиации, космического излучения, а также магнитным полем Земли. Высокие слои атмосферы - это также и химическая лаборатория, поскольку там в условиях, близких к вакууму, некоторые атмосферные газы под влиянием мощного потока солнечной энергии вступают в химические реакции. Наука, изучающая эти взаимосвязанные явления и процессы, называется физикой высоких слоев атмосферы.
КЛИМАТ, многолетний режим погоды на данной территории. Погоду в любой момент времени характеризуют определенные комбинации температуры, влажности, направления и скорости ветра. В некоторых типах климата погода существенно меняется каждый день или по сезонам, в других – остается неизменной. Климатические описания основываются на статистическом анализе средних и экстремальных метеорологических характеристик. Как фактор природной среды климат влияет на географическое распределение растительности, почв и водных ресурсов и, следовательно, на землепользование и экономику. Климат также оказывает воздействие на условия жизни и здоровье человека.
Климатология – наука о климате, изучающая причины формирования разных типов климата, их географическое размещение и взаимосвязи климата и других природных явлений. Климатология тесно связана с метеорологией – разделом физики, изучающим краткосрочные состояния атмосферы, т.е. погоду.
Кислород – важнейший элемент на Земле.Без еды животные могут обходиться в течение нескольких недель, без воды — несколько дней. Но без кислорода ониумирают через несколько минут. Кислород — это химический элемент, причем один из самых распространенных на земле. Он находится повсюду вокруг нас, составляя примерно одну пятую воздуха (а почти все остальное — азот). Кислород соединяется практически со всеми другими элементами. В живых организмах он соединяется с водородом, углеродом и другими веществами, составляя в человеческом теле примерно две трети общего веса. При нормальной температуре кислород взаимодействует с другими элементами очень медленно, образуя новые вещества, называемые оксидами. Этот процесс называется реакцией окисления. Окисление постоянно происходит в живых организмах. Пища является горючим живых клеток. При окислении пищи высвобождается энергия, используемая телом для движения и для его собственного роста. Медленное окисление, происходящее в организмах живых существ, часто называют внутренним дыханием. Человек вдыхает кислород через легкие. Из легких он попадает в кровеносную систему и разносится ею по всему телу. Дыша воздухом, мы снабжаем клетки нашего тела кислородом для их внутреннего дыхания. Таким образом, кислород нам нужен для получения энергии, благодаря которой организм может функционировать. Людей с нарушениями дыхания часто помещают в кислородные камеры, где больной дышит воздухом, на сорок—шестьдесят процентов состоящим из кислорода, и ему не приходится затрачивать много энергии на получение необходимого ему количества кислорода. Хотя кислород из воздуха постоянно забирается живыми существами для дыхания, его запасы тем не менее, никогда не иссякают. Растения выделяют его в процессе своего питания, тем самым пополняя наши запасы кислорода.
Оксидами называются сложные вещества, в состав молекул которых входят атомы кислорода в степни окисления – 2 и какого-нибудь другого элемента.
 Оксиды могут быть получены при непосредственном взаимодействии кислорода с другим элементом, так и косвенным путём (например, при разложении солей, оснований, кислот). В обычных условиях оксиды бывают в твёрдом, жидком и газообразном состоянии, этот тип соединений весьма распространён в природе. Оксиды содержатся в Земной коре. Ржавчина, песок, вода, углекислый газ – это оксиды.
Оксиды могут быть получены при непосредственном взаимодействии кислорода с другим элементом, так и косвенным путём (например, при разложении солей, оснований, кислот). В обычных условиях оксиды бывают в твёрдом, жидком и газообразном состоянии, этот тип соединений весьма распространён в природе. Оксиды содержатся в Земной коре. Ржавчина, песок, вода, углекислый газ – это оксиды.
Они бывают солеобразующими и несолеобразующие.
Солеобразующие оксиды – это такие оксиды, которые в результате химических реакций образуют соли. Это оксиды металлов и неметаллов, которые при взаимодействии с водой образуют соответствующие кислоты, а при взаимодействии с основаниями – соответствующие кислые и нормальные соли. Например, оксид меди ((CuO) оксид меди) является оксидом солеобразующим, потому что, например, при взаимодействии её с соляной кислотой ((HCl) водород хлор) образуется соль:
CuO + 2HCl → CuCl2 + h3O.
В результате химических реакций можно получать и другие соли:
CuO + SO(оксид серы)3 → CuSO4.
Несолеобразующими оксидами называются такие оксиды, которые не образуют солей. Примером могут служить СО (оксид углерода), N2O(оксид азота)(I),NO (оксид азота)(II).
Солеобразующие оксиды в свою очередь бывают 3-х типов: основными (от слова «основание»), кислотными и амфотерными.
Основными оксидами называются такие оксиды металлов, которым соответствуют гидроксиды, относящиеся к классу оснований.
 Химические свойства основных оксидов
Химические свойства основных оксидов
1. Растворимые в воде основные оксиды вступают в реакцию с водой, образуя основания.
2. Взаимодействуют с кислотными оксидами, образуя соответствующие соли.
3. Реагируют с кислотами, образуя соль и воду.
4. Реагируют с амфотерными оксидами.
Если в составе оксидов в качестве второго элемента будет неметалл или металл, проявляющий высшую валентность (обычно проявляют от IV до VII), то такие оксиды будут кислотными. Кислотными оксидами (ангидридами кислот) называются такие оксиды, которым соответствуют гидроксиды, относящие к классу кислот. Это, например, CO2, SO3, P2O5, N2O3, Cl2O5, Mn2O7 и т.д. Кислотные оксиды растворяются в воде и щелочах, образуя при этом соль и воду.
Химические свойства кислотных оксидов
1. Взаимодействуют с водой, образуя кислоту.
Но не все кислотные оксиды непосредственно реагируют с водой.
2. Реагируют с основанными оксидами с образованием соли.
3. Взаимодействуют со щелочами, образуя соль и воду.
В состав амфотерного оксида входит элемент, который обладает амфотерными свойствами. Под амфотерностью понимают способность соединений проявлять в зависимости от условий кислотные и основные свойства. Например, оксид цинка ZnO может быть как основанием, так и кислотой (Zn(OH)2 и h3ZnO2). Амфотерность выражается в том, что в зависимости от условий амфотерные оксиды проявляют либо осно́вные, либо кислотные свойства.
Химические свойства амфотерных оксидов
1. Взаимодействуют с кислотами, образуя соль и воду.
2. Реагируют с твёрдыми щелочами (при сплавлении), образуя в результате реакции соль – цинкат натрия и воду.
При взаимодействии оксида цинка с раствором щелочи (того же NaOH) (гидроксид натрия) протекает другая реакция.
Координационное число – характеристика, которая определяет число ближайших частиц: атомов или инов в молекуле или кристалле. Для каждого амфотерного металла характерно свое координационное число. Для Be (берелий) и Zn – это 4; Для и Al (алюминий) – это 4 или 6; Для и Cr (хром) – это 6 или (очень редко) 4;
Амфотерные оксиды обычно не растворяются в воде и не реагируют с ней.
netnado.ru
Дыхание растений
ВведениеДыхание — процесс универсальный. Оно является неотъемлемым свойством всех организмов, населяющих нашу планету, и присуще любому органу, любой ткани, каждой клетке, которые дышат на протяжении всей своей жизнедеятельности. Дыхание всегда связано с жизнью, тогда как прекращение дыхания — с гибелью живого.Жизнь организма в целом, как и каждое проявление жизнедеятельности, необходимо связаны с расходованием энергии. Клеточное деление, рост, развитие и размножение, поглощение и передвижение воды и питательных веществ, разнообразные синтезы и все другие процессы и функции осуществимы лишь при постоянном удовлетворении обусловленных ими потребностей в энергии и пластических веществах, которые служат клетке строительным материалом.
Источником энергии для живой клетки служит химическая (свободная) энергия потребляемых ею питательных веществ. Распад этих веществ, происходящий в акте дыхания, сопровождается освобождением энергии, которая и обеспечивает удовлетворение жизненных потребностей организма.
Сам же процесс дыхания представляет собой сложную многозвенную систему сопряженных окислительно-восстановительных процессов, в ходе которых имеет место изменение химической природы органических соединений и использование содержащейся в них энергии. 1. Дыхание. Определение. Уравнение. Значение дыхания в жизни растительного организма. Специфика дыхания у растений
Клеточное дыхание — это окислительный, с участием кислорода распад органических питательных веществ, сопровождающийся образованием химически активных метаболитов и освобождением энергии, которые используются клетками для процессов жизнедеятельности.
Суммарное уравнение процесса дыхания: С6Н12О6 + 602 ► 6С02 + 6Н20 + 2875 кДж/мольНе вся энергия, высвобождаемая при дыхании, может быть использована в процессах жизнедеятельности. Используется организмом в основном та энергия, которая аккумулируется в АТФ. Синтезу АТФ во многих случаях предшествует образование разности электрических зарядов на мембране, что, в свою очередь, связано с разностью концентраций ионов водорода по разные стороны от мембраны. Согласно современным представлениям, е только АТФ, но и протонный градиент служат источником энергии для различных процессов жизнедеятельности клетки. Обе формы энергии могут быть использованы на процессы синтеза, процессы поступления, передвижения питательных веществ и воды, создание разности потенциалов между цитоплазмой и внешней средой. Энергия, не накопленная в протонном градиенте и АТФ, в основном рассеивается в виде тепла или света и является для растения бесполезной.
Значение дыхания в жизни растения.
Дыхание — один из центральных процессов обмена веществ растительного организма. Выделяющаяся при дыхании энергия тратится как на процессы роста, так и нa поддержание в активном состоянии уже закончивших рост органов растения. Вместе с тем значение дыхания не ограничивается тем, что это процесс, поставляющий энергию. Дыхание, подобно фотосинтезу, сложный окислительно_восстановительный процесc, идущий через ряд этапов. На его промежуточных стадиях образуются органические соединения, которые затем используются в различных метаболических реакциях. К промежуточным соединениям относят органические кислоты и пентозы образующиеся при разных путях дыхательного распада. Таким образом, процесс дыхания — источник многих метаболитов. Несмотря на то что процесс дыхания в суммарном виде противоположен фотосинтезу, в некоторых случаях они могут дополнять друг друга. Оба процесса являются поставщиками как энергетических эквивалентов (АТФ, НАДФ-Н), так и метаболитов. Как видно из суммарного уравнения, в процессе дыхания образуется также вода. Эта вода в крайних условиях обезвоживания может быть использована растением и предохранить его от гибели. В некоторых случаях, когда энергия дыхания выделяется в виде тепла, дыхание ведет к бесполезной потере сухого вещества. В этой связи при рассмотрении процесса дыхания надо помнить, что не всегда усиление процесса дыхания является полезным для растительного организма.
2. Основные этапы становления учения о дыхании растенийНаучные основы учения о роли кислорода в дыхании были заложены трудами А.Л.Лавуазье. В 1774 г. кислород независимо открыли Пристли и Шееле, а Лавуазье дал название этому элементу. Изучая одновременно процесс дыхания животных и горение, Лавувзье в 1773-1783 гг. пришел к выводу, что при дыхании, как и при горении, поглощается кислород и образуется углекислый газ, причем в том и другом случаях выделяется тепло. На основании своих опытов он заключил, что процесс горения состоит в присоединении кислорода к субстрату и что дыхание есть медленно текущее горение питательных веществ в живом организме.
Я.Ингенхауз в 1778-1780 гг. показал, что зеленые растения в темноте, а незеленые части растений и в темноте, и на свету поглощают кислород и выделяют углекислый газ. В своей работе, опубликованной в 1779 г. он писал:
«Когда солнце, поднявшееся над горизонтом, разбудит своими лучами заснувшие за ночь растения, оно сделает их способными исполнять свою целительную функцию – исправлять воздух для животных; во мраке ночи эта деятельность совсем прекращается; днем же совершается с тем большим оживлением, чем светлее день и чем выгоднее расположено растение в отношении солнечных лучей. Затененные высокими зданиями или другими растениями, они не исправляют воздух, а, наоборот, выделяют вредный для дыхания животных воздух. К концу дня выработка очищенного воздуха ослабевает и при заходе солнца совершенно прекращается».
Первые точные исследования процесса дыхания у растений принадлежат Соссюру (1804). Он брал свежие листья и помещал их на ночь в сосуд, наполненный воздухом. При этом кислород воздуха поглощался и выделялся углекислый газ. Если на следующий день листья снова выставлялись на солнечный свет, то они выделяли почти такое же количество кислорода, какое поглотили ночью. Свои исследования Соссюр распространил и на незеленые части растений: стебли древесных растений, цветки, корни, плоды, и доказал, что дыхание наблюдается также в клетках этих органов. Он обнаружил, что при дыхании потеря в весе растения равна весу выделенного углерода.
Соссюр обратил внимание и на то, что молодые, растущие части растения, например новые побеги и распускающиеся цветки, дышат интенсивнее и потребляют кислорода больше, чем части растения, прекратившие рост.
Если, по Лавуазье, дыхание имеет сходство с процессом горения, то каким же образом органические вещества могут «гореть» при обычной температуре тела организма, да еще в водной среде, (ведь на 70 — 90% масса живых организмов состоит из воды)? Возникло предположение о том, что в живых клетках существуют механизмы, активирующие кислород. Швейцарский химик X. Ф. Шейнбайн, открывший озон, изучал причины быстрого потемнения пораненной поверхности растительных тканей, таких, как ткани яблок, картофеля, плодовых тел грибов. В 1845 г. он выступил со своей теорией окислительных процессов, согласно которой в живых клетках имеются соединения, способные легко окисляться в присутствии 02 и таким образом активировать молекулярный кислород. Если ткань прокипятить, то потемнения не происходит. Следовательно, потемнение тканей — каталитический окислительный процесс. Шейнбайн ошибочно полагал, что активация кислорода — это образование озона.
Исследования, начатые Шейнбайном, продолжил А. Н. Бах, который в 1897 г. разработал перекисную теорию биологического окисления, приложив ее к процессам дыхания. Несколько позже, в том же 1897 г., аналогичные взгляды высказал немецкий исследователь К. Энглер.
Суть перекисной теории биологического окисления Баха заключается в следующем. Молекулярный кислород имеет двойную связь и для того чтобы его активировать, необходимо эту двойную связь расщепить. Легко окисляющееся соединение А взаимодействует с кислородом и, разрывая двойную связь, образует пероксид А02 Таким образом, по мысли Баха, активация кислорода есть образование пероксида. В свою очередь пероксидное соединение, взаимодействуя с соединением В, окисляет его; затем эта реакция повторяется со вторым атомом кислорода и второй молекулой соединения В. Получается полностью восстановленное исходное соединение — акцептор кислорода А и полностью окисленное вещество В.
Много позднее, в 1955 г., две группы исследователей — О. Хаяиши с сотр. в Японии и Г. С. Мэзон с сотр. в США, используя современные методы, проанализировали возможность включения кислорода в органические соединения.
В настоящее время известно, что путь включения кислорода в органические соединения в соответствии с перекисной теорией биологического окисления Баха и Энглера не имеет отношения к дыханию, однако работы этих исследователей сыграли большую роль в изучении химизма дыхания, заложив основы современного понимания механизмов активации кислорода.
История современного учения о дыхании растений неразрывно связана с именем академика В.И. Палладина.
В годы первого петербургского периода работы Палладин исследовал ферментативную природу дыхательного процесса. Палладин показал, что и анаэробная, и аэробная фазы дыхания обеспечиваются специфическими ферментами, последовательно перерабатывающими продукты дыхания. Итоги работ этого периода изложены в монографии В.И. Палладина «Дыхание как сумма ферментативных процессов» (1907).
Одновременно с Палладиным проблемой дыхания занимались в целом ряде крупнейших научно-исследовательских институтов и лабораторий Западной Европы. Наибольшую популярность приобрели две новые школы – Виланда и Варбурга.
Т. Виланд развивал взгляды на роль дегидраз и водородных акцепторов, вполне аналогичные взглядам Палладина. Расхождение их теорий заключалось в том, что Виланд категорически отрицал какую бы то ни было роль оксидаз как специфических активаторов кислорода, считая молекулярный кислород способным самостоятельно отнимать водород от водородного акцептора. По мнению же Палладина, водородные акцепторы не могут самопроизвольно освобождаться от водорода, но требуют для этого участия оксидаз, которые поэтому являются обязательным фактором в реакции, выраженной во втором уравнении Палладина.
Противник Виланда, Варбург, считал, что молекулярный кислород не может вступить в организме в какой бы то ни было окислительный процесс, если в организме отсутствует система железоорганических соединений, типичным представителем которых он считал геминфермент. Варбург утверждал, что геминфермент активирует молекулярный кислород, т.е. как бы дает первый толчок к началу окислительных процессов, и без него никакой дыхательный процесс не может совершаться. Далее, по мнению Варбурга, окислительный импульс через промежуточные звенья (геминовые соединения) доходит до дыхательного субстрата и окисляет его. Резюмируя свои взгляды, Варбург утверждал, что дыхание осуществляется путем активации кислорода, а отнюдь не водорода. Но ведь Палладин как раз и говорил о той же необходимости активации молекулярного кислорода, защищая перед Виландом роль оксидаз в процессе дыхания.
Все различие в основных посылках Варбурга и Палладина заключается в том, что первый, работая по преимуществу с объектами животного происхождения, называл свой активатор молекулярного кислорода геминферментом, а Палладин, работавший с объектами растительного происхождения, сохранил за этим активатором ранее установившееся в науке название оксидазы. Но по существу оба говорили об одном и том же, протестуя против непримиримой позиции Виланда, отрицавшего необходимость энзиматической активации молекулярного кислорода.
Английский биохимик Д. Кейлин в 1925 г. доказал присутствие в клетках цитохромоксидазы, ускоряющей поглощение ими кислорода, и открыл другие цитохромы. Затем цитохромы были обнаружены у всех аэробов и было показано, что у этих организмов на завершающем этапе процесса дыхания осуществляется перенос на кислород электронов и протонов, в результате чего образуется Н20 (или Н202). 3.Каталитические системы дыханияОкисление дыхательных субстратов в ходе дыхания осуществляется с участием ферментов. Ферменты как белковые катализаторы, помимо свойств, присущих неорганическим катализаторам, обладают рядом особенностей: высокой активностью, высокой специфичностью по отношению к субстратам и высокой лабильностью. Их пространственная организации зависящая от нее активность изменяются под действием внешних и внутренних факторов. Эти свойства обеспечивают возможность тонкой регуляции обмена веществ на уровне ферментов.
Типы окислительно-восстановительных реакций. Существуют четыре способа окисления, и все они связаны с отнятием электронов:
1) непосредственная отдача электронов, например:2) Отнятие водорода:
3) присоединение кислорода:
4) образование промежуточного гидратированного соединения с последующим отнятием двух электронов и протонов:
Оксидоредуктазы.
Поскольку окисление одного вещества (донора электронов и протонов) сопряжено с восстановлением другого соединения (их акцептора), ферменты, катализирующие эти реакции, называют оксидоредуктазами. Все они относятся к I классу ферментов:Донор (Д) отдает электроны и протоны, акцептор (А) принимает их, а энзим (Е) осуществляет реакцию переноса. Существуют три группы оксидоредуктаз:
а) анаэробные дегидрогеназы передают электроны различным промежуточным акцепторам, но не кислороду;
б) аэробные дегидрогеназы передают электроны различным акцепторам, в том числе кислороду;
в) оксидазы способны передавать электроны только кислороду.
Анаэробные дегидрогеназы. Это двухкомпонентные ферменты, коферментом которых может быть НАД+(никотинамидадениндинуклеотид):При окислении субстрата НАД+превращается в восстановленную форму НАДH, а второй протон субстрата диссоциирует в среду (НАДH+ Н+). К анаэробным НАД-зависимым дегидрогеназам относятся такие ферменты, как алкогольдегидрогеназа, лактатдегидрогеназа, малатдегидрогеназа и др. Коферментом анаэробных дегидрогеназ может быть также НАДФ+(никотинамидадениндинуклеотидфосфат), содержащий на одну фосфатную группировку больше, чем НАД + . НАДФ- зависимыми дегидрогеназами являются изоцитратдегидрогеназа, глюкозо-6-фосфатдегидрогеназа, 6-фосфоглюконатдегидрогеназа и др.
Субстратная специфичность фермента зависит от его белковой части. Многие НАД- и НАДФ-зависимые дегидрогеназы нуждаются в присутствии ионов двухвалентных металлов. Например, алкогольдегидрогеназа содержит ионы цинка.
Окисленные и восстановленные формы коферментов анаэробных дегидрогеназ могут взаимопревращаться в реакции, катализируемой ферментом НАД(Ф)-трансгидрогеназой:НАДФH + НАД+ = НАДФ+ + НАДHАнаэробные дегидрогеназы передают водород, т. е. электроны и протоны, различным промежуточным переносчикам и аэробным дегидрогеназам.
Аэробные дегидрогеназы. Это также двухкомнонентные ферменты, получившие название флавиновых (флавопротеины).
Помимо белков, в их состав входит прочно связанная с ними простетическая группа — рибофлавин (витамин В2).
Различают два кофермента этой группы: флавинмононуклеотид (ФМН), или желтый дыхательный фермент Варбурга, и флавинадениндинуклеотид (ФАД).
ФМН (рибофлавин-5-фосфат) содержит гетероциклическое азотистое основание — диметилизоаллоксазин, спирт рибит (производное рибозы) и фосфат: В ФАД кроме ФМН имеется еще один нуклеотид — аденозинмонофосфата:
Активной группой в реакции присоединения и отдачи электронов и протонов в ФМН и ФАД служит изоаллоксазин. Взаимодействие с восстановленным переносчиком, например НАДH, происходит следующим образом:
Примером дегидрогеназы, в состав которой входит ФАД, является сукцинатдегидрогеназа. Доноры электронов для аэробных дегидрогеназ — анаэробные дегидрогеназы, а акцепторы — хиноны, цитохромы, кислород.
Цитохромная система. Среди оксидаз очень важную роль играют железосодержащие ферменты и переносчики, относящиеся к цитохромной системе. В нее входят цитохромы " и цитохромоксидаза. Включаясь в определенной последовательности в процесс переноса электронов, они передают их от флавопротеинов на молекулярный кислород.
Все компоненты цитохромной системы содержат железопорфириновую простетическую группу.
При переносе электронов цитохромами железо обратимо окисляется и восстанавливается, отдавая или приобретая электрон и изменяя таким образом свою валентность. В дыхательной цепи направление транспорта электронов определяется величиной окислительно-восстановительного потенциала цитохромов.
В этой системе передавать электроны непосредственно на кислород способна только цитохромоксидаза (цит. а + а3). Из всех известных оксидаз она имеет наибольшее сродство к кислороду. Ингибиторами цитохромоксидазы являются СО, цианид, азид. Б растительных митохондриях кроме цитохромоксидазы функционирует оксидаза, не подавляемая цианидом и названная альтернативной оксидазой. Например, в митохондриях початков ароидных активность цианидустойчивой оксидазы в 10 раз превышает активность цитохромоксидазы.
Пероксидаза и каталаза. К пероксидазам относят целую группу ферментов, использующих в качестве окислителя пероксид водорода: классическую пероксидазу, НАД-пероксидазу, НАДФ-пероксидазу, пероксидазу жирных кислот, глутатионпероксидазу, цитохромпероксидазу и др. Все они работают по следующей схеме, где А — субстраты:В последние 2 — 3 десятилетия показана полифункциональность пероксидаз. Помимо пероксидазной, у них имеется оксидазная функция, т. е. способность переносить электроны в отсутствие пероксидного кислорода на молекулярный кислород. Пероксидаза может также функционировать как анаэробная дегидрогеназа, например НАДH-дегидрогеназа, передающая электроны от восстановленных пиридиновых нуклеотидов на разные акцепторы.
Пероксид водорода, помимо пероксидазы, расщепляется также каталазой, в результате чего образуется молекулярный кислород. В реакции участвуют две молекулы пероксида, одна из которых функционирует как донор, а другая — как акцептор электронов.
Простетической группой пероксидазы и каталазы служит гем, в состав которого входит атом железа.
Оксигеназы. Наряду с оксидазами, которые используют молекулярный кислород как акцептор электронов, в клетках широко представлены оксигеназы, активирующие кислород, в результате чего он может присоединяться к органическим соединениям. Ферменты, внедряющие в субстрат два атома кислорода, называют диоксигеназами, а присоединяющие один атом кислорода — монооксигеназами или гидроксилазами. В качестве доноров электронов оксигеназы используют НАД(Ф)H, ФАДh3 и др.
Оксигеназы присутствуют во всех типах клеток. Они участвуют в гидроксилировании многих эндогенных соединений в частности аминокислот, фенолов, стеринов и др., а также в детоксикации чужеродных токсических веществ (ксенобиотиков).
4.Основные пути диссимиляции углеродаСуществуют два основных пути окисления углеводов: 1) дихотомический (гликолитический) и 2) апотомический (пентозофосфатный). Белки, жиры и органические кислоты окисляются в глиоксилатном цикле.
Относительная роль этих путей дыхания может меняться в зависимости от типа растений, возраста, фазы развития, а также в зависимости от условий внешней среды. Процесс дыхания растений осуществляется во всех внешних условиях, в которых возможна жизнь. Растительный организм не имеет приспособлений к регуляции температуры, поэтому процесс дыхания осуществляется при температуре от – 50 до +50°С. Нет приспособлений у растений и к поддержанию равномерного распределения кислорода по всем тканям. Именно необходимость осуществления процесса дыхания в разнообразных условиях привела к выработке в процессе эволюции разнообразных путей дыхательного обмена и к ещё большему разнообразию ферментативных систем, осуществляющих отдельные этапы дыхания. При этом важно отметить взаимосвязь всех процессов обмена в организме. Изменение пути дыхательного обмена приводит к глубоким изменениям во всем метаболизме растительных организмов.4.1 Дихотомический путьЭто основной путь распада органических веществ для всех живых организмов. Выделяют 2 этапа дихотомического пути: гликолиз и цикл Кребса.
Рис. 1 Основные этапы дыхания
www.coolreferat.com







