
Двойное оплодотворение покрытосеменных растений. Двойное оплодотворение покрытосеменных растений
Половое размножение покрытосеменных растений – зародыш, двойное оплодотворение и особенности
Половое размножение покрытосеменных растений связано с цветком. В цветке, органе размножения, происходит созревание мужских и женских половых клеток (гамет), и их последующее слияние с образованием первой клетки дочернего организма.

Отличие полового размножения от вегетативного
Половое и вегетативное – два вида размножения покрытосеменных. При вегетативном размножении новые организмы возникают за счёт регенерации вегетативных органов (лист, корень, побег).
Цветок не относится к вегетативным, а является репродуктивным (лат. – reproductio – воспроизведение) органом. В нём при слиянии гамет образуется зигота, из которой впоследствии развивается зародыш нового растения.
Гаметы
Гаметы имеют существенное отличие от всех других клеток. Число хромосом в ядрах гамет в два раза меньше, чем в остальных клетках. Такой набор хромосом называется гаплоидным. Набор хромосом обычных клеток тела называется диплоидным.
Хромосомы содержат наследственную информацию о признаках организма. Дочерний организм имеет половину хромосом от мужской гаметы и столько же от женской.
Тычинки и пестики
На тычинках развивается пыльца. Пыльца содержит генеративную клетку, которая, делясь, образует две мужские гаметы, называемые спермиями.
Женская гамета, или яйцеклетка, вместе с сопутствующими ей клетками находится внутри завязи пестика, в полости зародышевого мешка.

Рис. 1. Зародышевый мешок.
Опыление
Опыление – это процесс переноса пыльцы на рыльце пестика, который осуществляется с помощью ветра, воды, насекомых и некоторых других животных. Человек может и сам, целенаправленно, вручную производить опыление растений.
Пыльца на пестик может попадать с других цветков, а может с тычинок этого же цветка.
С помощью ручного опыления можно повышать урожайность и выводить множество новых сортов растений.
Рис. 2. Ручное опыление.
Двойное оплодотворение
После опыления спермии продвигаются в зародышевый мешок. Это происходит с помощью пыльцевой трубки, которая является неполовой клеткой пыльцы. Пыльцевая трубка быстро растёт (35 мм/час) в направлении яйцеклетки, и с ней продвигаются спермии.
Спермии бывают различной формы и не имеют жгутиков. Когда пыльцевая трубка достигает яйцеклетки, один спермий соединяется с ней, а другой с центральной клеткой зародышевого мешка.
В итоге, при двойном оплодотворении у покрытосеменных растений происходит образование следующих клеток:
- первый спермий + яйцеклетка = зигота;
- второй спермий + центральная клетка = эндосперм.
Зигота впоследствии делится и превращается в зародыш. Эндосперм служит для зародыша источником питательных веществ. Вместе, зародыш и эндосперм, образуют семя.
Рис. 3. Схема двойного оплодотворения.
Зародыш
Зародыш покрытосеменных растений – это зачаточный дочерний организм, который находится в семени в состоянии покоя, пока семя не начало прорастать. Набор хромосом у зародыша, как и у зиготы, диплоидный.
Семя с зародышем созревает, в эндосперме накапливаются питательные вещества. Завязь пестика разрастается и превращается в плод.
Что мы узнали?
Изучая в 6 классе половое размножение покрытосеменных, мы должны понимать какие особенности характерны для этих растений. Главная особенность покрытосеменных – это наличие цветка. В цветке образуются и развиваются гаметы. Покрытосеменные размножаются семенами. Семя образуется в результате полового процесса, который у цветковых заканчивается двойным оплодотворением.
Тест по теме
obrazovaka.ru
MINIMUM_po_botanike
Водоросли.Общая характеристика. Водоросли – растения, живущие преимущественно в воде. Тело их не расчленено на органы и ткани. Органы размножения одноклеточные. Это одни из древнейших представителей растительного мира. Водоросли бывают одноклеточными (хламидоионада, хлорелла), колониальные (носток) и многоклеточные (спирогира, ламинария, ). Цитологические особенности:
Клетки покрыты клеточной стенкой. Характерно наличие хроматофоров, носителей окраски. Хроматофоры – органеллы, в которых происходит фотосинтез. Окраска зависит от глубины обитания водорослей: на большой глубине – буро-красные, ближе к поверхности – зеленые. В хроматофорах всех водорослей содержатся пиреноиды, они синтезируют крахмал.
Одноклеточные формы могут быть подвижными (со жгутиками) и неподвижными.
Также водоросли бывают прикрепленные (бентосные) и свободноплавающие (планктонные) Размножение: Вегетативное – новые водоросли образуются из обрывков нитей, кусков слоевищ и т.д. Бесполое – содержимое одной клетки (зооспорангия) делится многократно, образую новые подвижные клетки (зооспоры). Каждая из них дает начало новой особи. Половое размножение – широко распространено у водорослей. Формы полового процесса разнообразны: изогамия (♂подвижна, ♀ подвижна, одинаковые по размеру), гетерогамия (♂подвижна, ♀ подвижна, ♀ больше, чем ♂), оогамия (♂подвижна, ♀ неподвижна, ♀ больше, чем ♂), конъюгация (сливается содержимое двух вегетативных клеток). Образовавшаяся зигота покрывается толстой клеточной стенкой, накапливает запасные питательные вещества и в состоянии покоя способна легка переносить неблагоприятные условия.

1.Общая характеристика отдела Папоротниковые. За исключением нескольких родов все папоротники - равноспоровые. У них наблюдается смена поколений с преобладание спорофита над гаметофитом. Спорофит представлен многолетними корневищными травами с крупными, обычно перисто-рассеченными листьями , на нижней стороне которых расположены спорангии. В условия тропического климата встречаются древовидные папоротники. Листья папоротников нарастают верхушкой. То, что у папоротника напоминает лист — вовсе не лист, а по своей природе — целая система ветвей, да ещё расположенных в одной плоскости. Так это и называется — плосковетка, или вайя, или, ещё одно название, — предпобег. В спорангиях в результате мейоза образуются споры (n). Спорангии лопаются, споры высыпаются, прорастает заросток (гаметофит). Заростки представляют собой сердцевидную зеленую пластинку. Точка роста находится в выемке. От нижней стороны отходят ризоиды. На заростке образуются антеридии (в них образуются сперматозоиды) и архегонии ( в ней образуется яйцеклетка). Во время дождя или обильной росы сперматозоиды проникают в архегонии и оплодотворяют яйцеклетку. Из зиготы развивается зародыш, затем взрослый спорофит. Современные папоротники насчитывают около 300 родов и 12 000 видов. Основные представители: Щитовник мужской, Страусник обыкновенный, Сальвиния плавающая (разноспоровая)

Опыление у покрытосемянных растений. Приспособление цветков к различным видам опыления. Микро- и макроспорогенез у растений Опыление – перенос пыльцы с тычинок на рыльца пестика. Различают самоопыление и перекрестное опыление. Самоопыление: пыльца опыляет рыльце пестика того же цветка (пшеница, ячмень, овес, просо, горох, фасоль, бобы, хлопчатник, лен, томат и др.) . Преобладает у 10% растений. Самоопыление происходит как у раскрывшихся цветков: сельдерейные, так и у закрытых: арахис, фиалка. Однако для эволюционного развития этот вид самоопыления не совершенен, т.к.не несет новых генетических признаков. Поэтому многие растения имеют приспособления, препятствующие самоопылению: Двудомность ( ♂ и ♀ цветки формируются на разных растениях) Однодомность ( ♂ и ♀ цветки формируются на одном растении, но в разных цветках) Дихогамия – разновременное созревание пыльцы и рыльца Гетеростилия – рыльца и тычиночные нити различаются по длине Самонесовместимость. Перекрестное опыление : пыльца опыляет рыльца других цветков. Различают 2 типа: гейтоногамия – опыление в пределах одного растения ксеногамия – опыление в пределах разных цветков (наиболее оптимально для эволюционного процесса) Различают несколько видов: Абиотическое – при помощи неживых факторов внешней среды Анемофилия (ветром)
Гидрофилия (водой)
Биотическое – с помощью животных.
Энтомофилия – опыление насекомыми Орнитофилия – опыление мелкими птицами (колибри)

Строение семязачатка. Двойное оплодотворение у покрытосеменных растений.
Строение семязачатка. Семязачаток состоит из центральной части - нуцеллуса и окружающих его одного или двух покровов - интегументов , которые над верхушкой нуцеллуса образуют небольшой канал - микропиле. Семязачаток сообщается с плацентой с помощью семяножки, или фуникулуса . Нуцеллус является аналогом макроспорангия, в котором развивается одна макроспора. Двойное оплодотворение было открыто С.Г.Навашиным в 1898 г. Оплодотворение у покрытосемянных принято называть двойным, т.к. оба спермия сливаются с клетками зародышевого мешка. Один сливается с яйцеклеткой, в результате образуется зигота. Второй сливается с центральным ядром, в результате образуется триплоидная клетка (3n). Прочие клетки зародышевого мешка дегенерируют. После двойного оплодотворения из зиготы развивается зародыш, а из триплойдной клетки – эндосперм (питательная ткань), из нуцеллуса образуется перисперм (дополнительная питательная ткань), из интегумента – семенная кожура, из семязачатка – семя, из завязи – плод. Преимущества двойного оплодотворения заключаются в том, что одновременно с зиготой формируется триплоидная клетка (3n), которая делится быстрее, чем зигота. Соответственно, эндосперм образуется быстрее, чем растет зародыш. Поэтому не нужно запасать питательные вещества до оплодотворения, в отличие от голосеменных, у которых довольно мощный гаплоидный эндосперм развивается до оплодотворения. Этим достигается существенная экономии я энергетических ресурсов организма. Семязачатки покрытосеменных, не обремененные запасающей питательной тканью впрок, развиваются гораздо быстрее, чем у голосеменных.


1 - покровы семязачатка, или интегументы (а - наружный, б - внутренний), 2 - микропиле , 3 - халаза , 4 - фуникулус , 5 - нуцеллус , 6 -зародышевый мешок , 7 - яйцеклетка , 8 - синергиды , 9 - антиподы , 10 - вторичное ядро, 11 - плацента , 12 - проводящий пучок. После оплодотворения из зиготы развивается зародыш, из центрального ядра – эндосперм, из нуцеллуса – перисперм, из интигументов – семенная кожура, из семязачатка – семя, из завязи – плод.
Высшие растения. Общая характеристика и цикл развития.
Высшие растения, или Наземные растения,— тип зелёных растений, которым свойственна дифференциация тканей, в отличие от низших растений — водорослей. К высшим растениям относятся мхи и сосудистые растения (папоротникообразные, псилотовые,хвощевидные, плауновидные, голосеменные и покрытосеменные).
Развитие специализированных тканей было важным условием для выхода растений на сушу. Для комфортного существования в воздушной среде растениям было необходимо развить как минимум эпидермис с устьицами для защиты от высыхания и теплообмена и проводящие ткани для обмена минеральных и органических веществ. Результатом выхода растений на сушу также стало разделение организма растения на корень, стебель и лист.
В ЖЦ высших растений происходит чередование полового и бесполого способов размножения и связанное с этим чередование поколений. Бесполое поколение представлено спорофитом (2n), половое – гаметофитом (n). Спорофит – это растение, образующее споры. В многоклеточных спорангиях в результате мейотического деления формируются споры (n). Растения, у которых все споры одинаковые – равноспоровые, у более высокоорганизованных споры разной величины (микроспоры и мегаспоры) – это разноспоровые растения. Гаметофит – растение, образующее гаметы. Гаметы равиваются в многоклеточных органах полового размножения: яйцеклетки – в архегониях, сперматозойды – в антеридиях. Гаметофит вырастает из споры. У равноспоровых растений гаметофит обоеполый, у разноспоровых – однополый. В результате оплодотворения образуется зигота, из которой прорастает новый спорофит. Все высшие растения делятся на 2 группы по доминированию в ЖЦ гаметофита и спорофита: Растения с доминирующим гаметофитом – отдел Моховидные Растения с доминирующим спорофитом – все остальные
В целом для эволюции высших растений характерна тенденция к усложнению и усовершенствованию спорофита при одновременной редукции гаметофита.

Моховидные. Общая характеристика. Цикл развития мха Кукушкин лен. В отдел входит более 25 000 видов сравнительно просто организованных травянистых растений. В цикле развития преобладает гаметофит. У более примитивных форм он представлен талломом, а у остальных – расчленен не стебель и листья. Корней нет, из заменяют ризоиды. Спорофит самостоятельно не существует, развивается на гаметофите, получая от него воду и питательные вещества. Спорофит представляет собой коробочку, где развивается спорангий. Отдел делится на 3 класса: Антоцеротовые, Печеночные, Листостебельные мхи. Наиболее известен вид – Кукушкин лен (Класс Листостебельные мхи). Представляет собой прямостоячие стебли (15-20 см) густо покрытые жесткими острыми листьями. Прикрепляется к земле ризоидами. Гаметофиты раздельнополые. На верхушке мужских особей развиваются антеридии, окруженные красно-бурыми листьями (n), на верхушках женских – архегонии (n). Оплодотворение происходит подвижными двужгутиковыми сперматозоидами во влажную погоду. Из зиготы на верхушке женского гаметофита развивается спорофит (2n), имеющий вид коробочки на длинной ножке. Внутри коробочки – спорангий, где после мейоза образуются споры (n). После образования спор колпачок, а затем крышечка отделяются и споры высыпаются. Из споры сначала образуется протонема, на которой из особых почек формируются листостебельные побеги – гаплоидное поколение (n).



Общая хар-ка отдела Плауновидные. Цикл развития Плауна булавовидного. Плауновидные – очень древняя группа, возникшая в силуре, а в каменноугольном периоде достигла своего расцвета. Это были огромные деревья, образующие целые леса. В современной флоре они представлены вечнозелеными многолетними травами, реже полукустарниками. Около 1000 видов сохранилось. Плауновидные имеют мелкие листья с одной жилкой микрофильного типа. Выделяют 2 класса: равноспоровые Плауновые и разноспоровые Полушниковые. ^ Плаун булавовидный. В цикле развития преобладает спорофит (2n). Он представляет собой длинные стелющиеся ветвящиеся побеги густо усажены жесткими мелкими листьями. От стебля отходят тонкие придаточные корни. В середине лета на верхушках появляются спороносные колоски. Колосок состоит из оси и сидящих на ней листочков (спорофиллов). На верхней стороне спорофиллов находится спорангий на короткой ножке. В результате мейоза в нем образуются споры (n). Из спор развивается заросток (гаметофит) (n). Заросток развивается под землей. От нижней его части отходят ризоиды. Через них он врастает в грибы, образуя микоризу. Живет в симбиозе с грибом, питается от него. Растет очень долго (15-20 лет). На верхней стороне его образуется антеридий и архегонии. Двужгутиковый сперматозоид выходи из антеридия, проникает в архегоний и оплодотворяет яйцеклетку. В результате образуется зигота, из которой развивается новый спорофит.


Цикл развития голосеменных растений на примере Сосны обыкновенной.
Голосеменные – это древний отдел растений, которые характеризуются наличием семязачатков с последующим развитием семени, но не образуют цветков и плодов. К голосеменным относятся около 800 современных видов. Преобладающее большинство видов этого отдела (более 500) составляют класс хвойных растений.
Типичный представитель – СОСНА ОБЫКНОВЕННАЯ Сосна – разноспоровое однодомное растение. Микро- и мегаспорангии образуются на одном и том же дереве в отдельных шишках. Женские шишки состоят из оси и семенных чешуй, на верхней стороне которых развивается по 2 семязачатка. Семязачаток состоит из нуцеллуса, покрытого интегументом, в котором имеется отверстие – микропиле. Внутри нуцеллуса путем мейоза образуются 4 мегаспоры, 3 из них отмирают, а одна прорастает в женский гаметофит – зародышевый мешок.
Мужские шишки собраны в сошишие на верхушках удлиненных побегов. В микроспорангиях в результате мейоза из клеток спорогенной ткани возникают микроспоры (n). Микроспора покрыта 2 оболочками: интина (внутренняя) и экзима (внешняя). Экзима отстает от интины, образую 2 воздушных мешка. Увеличивая поверхность пыльцы, они способствуют переносу ее ветром, а при попадании пылинки в жидкость выполняют роль поплавков. Мужские шишки после производства пыльцы быстро засыхают и опадают с ветвей. Пыльцевое зерно содержит мужской гаметофит, состоящий из двух клеток – вегетативной и генеративной. Опыление происходит с помощью ветра в начале лета. Пыльца прилипает к капле клейкой жидкости, которая выделяется в области микропиле, вегетативная клетка начинает удлиняться в пыльцевую трубку, чешуи женских шишек смыкаются.
В начале весны происходит оплодотворение: генеративная клетка делится с образованием двух спермиев, которые движутся по пыльцевой трубке. Далее один из спермиев сливается с яйцеклеткой, а другой отмирает. Из оплодотворенной яйцеклетки (зиготы) формируется зародыш семени, а семязачаток превращается в семя.
Семена у сосны созревают на второй год, высыпаются из шишек и, подхваченные животными или ветром, переносятся на значительные расстояния.
Классификация покрытосеменных растений. Отдел Покрытосеменные делится на 2 класса: однодольный и двудольные. Эволюция среди этих классов шла разными путями, вследствие чего каждый из них имеет характерные отличия. Эти отличия настолько сильны, что двудольные не скрещиваются с однодольными и прививок между ними не получено. Основные различия
| Класс Двудольные | Класс Однодольные |
| Около 190 000 видов | Около 63 000 видов |
| Семя | |
| Зародыш с двумя семядолями | Зародыш с одной семядолей |
| Семядоли латеральные (боковые), обычно с тремя главными проводящими пучками | Семядоля терминальная (верхушечная), с двумя проводящими пучками |
| Корень | |
| Система главного корня , стержневая | Придаточная, мочковатая КС |
| Вторичное утолщение (камбий есть) | Отсутствие вторичного утолщения (камбия нет) |
| Стебель | |
| Сплошное расположение проводящих тканей или пучки, расположенные по кругу | Разбросанное расположение проводящих пучков |
| Вторичное утолщение, камбий есть | Отсутствие вторичного утолщения (камбия нет) |
| Лист | |
| Простой, сложный, более или менее четко разделен на черешок и пластинку | Простой, обычно не расчлененный на черешок и пластинку |
| Жилкование перистое или пальчатое | Жилкование параллельное или дуговое |
| Цветок | |
| Пяти-, много-, четырехчленный | Трехчленный |
Отдел Хвощевидные. Общая характеристика. Цикл развития Хвоща полевого. Отдел Хвощевидные в настоящее время представлены только одним родом Хвощ и 25 видами. В цикле развития преобладает спорофит (2n), который представлен в виде многолетней травы. От корневищ отходят придаточные корни и надземные побеги, обычно однолетние. Побеги имеют типичное членистое строение. От узлов отходят бурые чешуйчатые листья, сросшихся в трубчатое влагалище. Функцию фотосинтеза выполняют стебли. Споры (n) образуются в спороносных колосках, развивающихся в зависимости от вида хвоща на верхушках ассимилирующих или на специализированных спороносных бесхлорофилльных побегах. На оси колосков находятся спорангиофоры, имеющие вид щитка на ножке, к которому прикреплены 8-10 спорангиев. Образовавшиеся после мейоза споры покрыты толстой оболочкой, снабжены лентовидными пружинками – элатерами (при высыхании они раскручиваются, при увлажнении закручиваются, благодаря чему сцепляются между собой). В результате прорастания спор образуются мужские и обоеполые заростки. Заростки очень маленькие, имеют вид рассеченных пластинок, от нижней поверхности которых отходят ризоиды. Гаметофит созревает в течение 3-5 недель. На концах лопастей мужских заростков развиваются антеридии, в средней части архегонии. Архегонии развиваются раньше, чтобы увеличить вероятность оплодотворения чужими сперматозоидами. Сперматозоиды много жгутиковые. Из зиготы развивается новый спорофит.


Строение цветка, особенности строения околоцветника. Цветок – сложный репродуктивный орган покрытосеменных растений. Цветок имеет ось (цветоложе), несущее листочки околоцветника, тычинки и пестик (или пестики). Если цветоножка не выражена, цветок называют сидячим. Части цветка делятся на фертильные (тычинка, пестик) и стерильные (чашечка, венчик, околоцветник). Цветок, содержащий тычинки и пестики. называют обоеполым (у 70% цветков). Но бывают случаи, когда цветки однополые. Растения с однополыми цветками, находящимися на одном и том же экземпляре, называют однодомными (кукуруза, дуб, бук, огурец, тыква), если цветки расположены на разных экземплярах – двудомные (конопля, ива, тополь). В отличие от симметрии различают: правильные (актиноморфные) цветки (если можно провести несколько осей симметрии) – капустные, гвоздичные, розовые неправильные (зигоморфные) цветки (если можно провести только одну ось) – бобовые, яснотковые нессиметричные (ассиметричные) – если нельзя провести даже одну ось симметрии – валериана лекарственная. Околоцветник – это стерильная часть цветка, является покровом, защищающим более нежные тычинки и пестики. Он бывает двойной и простой. ^ Двойной околоцветник дифференцирован на чашечку и венчик. Чашечка состоит из чашелистиков, образует наружный круг околоцветника. Чашелистики обычно имеют небольшие размеры и зеленую окраску. Главная функция – защита цветка до раскрывания бутона. Венчик состоит из лепестков. Лепестки имеют более крупные размеры и различаются по окраске. Окраску определяют различные пигменты: антоциан (розовая, красная, синяя, фиолетовая), каротиноиды (желтая, оранжевая, красная), антохлор (лимонно-желтая), антофеин (коричневая) Аромат цветка создается эфирными маслами. Венчик может быть раздельнолепестным (лютик, земляника) и сростнолепестным (картофель, шалфей). ^ Простой околоцветник не дифференцирован на чашечку и венчик, состоит из совокупности однородных листков околоцветника. Может быть венчиковидным (тюльпан) и чашечковидным (манжетка) 
Простые и двойные околоцветники бывают раздельнолепестными (яблоня, тюльпан) и сростнолепестными (колокольчик, ландыш). Кроме чашечки и венчика с стерильным частям цветка относят еще нектарник. ФОРМУЛА И ДИАГРАММА ЦВЕТКА Строение цветка можно выразить в виде формулы. При ее составлении используют следующие обозначения:
 — цветок правильный (актиноморфный)
— цветок правильный (актиноморфный)  — цветок неправильный (зигоморфный) Ca— чашечка(calyx) Co— венчик(corolla) P — простой околоцветник(perigonium) A — тычинки(андроцей) (androceum) G — пестик, плодолистики (гинецей) (gynoeceum)
— цветок неправильный (зигоморфный) Ca— чашечка(calyx) Co— венчик(corolla) P — простой околоцветник(perigonium) A — тычинки(андроцей) (androceum) G — пестик, плодолистики (гинецей) (gynoeceum)
Рядом с буквенными выражениями частей цветка цифрами указывается количество элементов (пятилепестный венчик — Co5, шестичленный андроцей — A6), а в том случае если их число в цветках одного и того же вида непостоянно (обычно больше 12)— символом  . Если элементы цветка сросшиеся, то их число заключается в скобки: сросшийся пятичленный венчик — Co(5), двубратственный андроцей — A(9,1). Если элементы цветка расположены кругами, то между количеством элементов в каждом круге ставится знак «+» (P3+3). Положение завязи в цветке обозначается чёрточкой. При верхней завязи цветок является подпестечным, поэтому под цифрой числа плодолистиков ставят чёрточку G(
. Если элементы цветка сросшиеся, то их число заключается в скобки: сросшийся пятичленный венчик — Co(5), двубратственный андроцей — A(9,1). Если элементы цветка расположены кругами, то между количеством элементов в каждом круге ставится знак «+» (P3+3). Положение завязи в цветке обозначается чёрточкой. При верхней завязи цветок является подпестечным, поэтому под цифрой числа плодолистиков ставят чёрточку G( ), при обозначении нижней завязи — цветок надпестичный — чёрточку ставят над цифрой G(
), при обозначении нижней завязи — цветок надпестичный — чёрточку ставят над цифрой G( ). При простом околоцветнике знаки чашечки и венчика не применяются, и он обозначается буквой P (перигониум).. Например, формула цветка лилии и тюльпана:
). При простом околоцветнике знаки чашечки и венчика не применяются, и он обозначается буквой P (перигониум).. Например, формула цветка лилии и тюльпана:  P3+3A3+3G(
P3+3A3+3G( )
)
Диаграммой называют схематическую проекцию цветка на плоскости в следующей ориентации: ось соцветия вверху, кроющий лист – внизу. 
Классификации плодов. Морфогенетическая классификация основана на типе гинецея: Искусственная классификация плодов основана главным образом на признаках внешней морфологии: Простой плод развивается только из завязи одного пестика (монокарпный, ценокарпный гинецей). Сборный плод (сложный плод) формируется из завязей нескольких свободных пестиков (апокарпный гинецей). Соплодие – сросшиеся в единое целое несколько или много плодов, образовавшихся из цветков одного соцветия. В зависимости от консистенции околоплодника различают: сухие и сочные В зависимости от числа семян различают: многосемянные и односемянные. В зависимости от вскрывания околоплодника различают: невскрывающиеся и вскрывающиеся плоды.
Сочные плоды. В эволюции сочные плоды появились как производные сухих. Стенка СП может развиваться либо из стенки завязи (перикарпа), либо из стенки завязи, сросшейся с цветочной трубкой или с вогнутым цветоложем. Стенка незрелого СП имеет плотную структуру, по мере созревания плода она становится мягче. Созревание сопровождается как правило изменением окраски: за счет разрушения хлорофилла и накопления либо каротиноидов (желтый, оранжевый цвета), либо антоцианов (красные, синие). Созревание также сопровождается накоплением углеводов. Среди сочных плодов выделяют многосемянные сочные плоды Гранатина - развивается из нижней многогнездной завязи с двумя этажами. Мякоть образуется из сочного наружного слоя семенной кожуры. Околоплодник и ткани цветочной трубки подсыхают, образую твердую кожуру Геспередий (померанец) – цитрусовые, формируется из синкарпного гинецея. Экзокарп – желтая ткань, покрыта слоем кутикулы, воска и маслянистыми железками. Мезокарп – белая ткань. Эндокарп – пленчатый. Субэпидермальные клетки эндокарпа формируют соковые мешочки (мякоть плода). Тыквина – экзокарп жесткий, одревесневающий, кожистый; мякоть образована мезо- и эндокарпом (тыква, дыня) или разросшимися плацентами (арбуз). Образуется из паракарпного гинецея. Яблоко – формируется из апокарпного гинецея
Ягода – сочный мясистый эндо и мезокарпий, тонкий пленчатый экзокарпий Односемянные сочные плоды: Костянка – невскрывающийся монокарпий с мясистым сочным съедобным мезокарпом и склерифицированным эндокарпом (косточка). (слива, вишня ,черешня, абрикос, персик, алыча).
Простые сухие плоды:
| Многосемянные | Односемянные | |
| Вскрывающиеся | Распадающиеся | |
| Листовка (рогатый василек) | Двусемянка (морковь, укроп) | Зерновка (рожь, пшеница) |
| Боб (Бобовые) | Двукрылатка (клен) | Семянка (подсолнечник) |
| Стручок (капуста, сурепица) | Четырехорешек | Крылатка (ясень, вяз, береза) |
| Стручочек (пастушья сумка) | Дробная коробочка | Орех (фундук) |
| Коробочка (мак, белена, хлопчатник) | Членистый боб (копеечник, вязель) | Орешек (липа, гречиха) |
| Членистый стручок (редька дикая) | Желудь (дуб, бук) | |
Общая характеристика высших растений, чередование гаплоидной и диплоидной фаз у высших растений. Высшие растения – одно из подцарств царства Растений, объединяющее не менее 300 000 видов – от мхов до покрытосеменных. Их происхождение связано с выходом на сушу морских многоклеточных водорослей. В процессе приспособления к наземным условиям существования возникли новые типы растений с новыми признаками: 1. На смену таллому у высших растений формируется тело, расчлененное на органы: корень и побег (листья и стебель) 2. Усложняется внутреннее строение, появляются специализированные ткани: покровные (защита растений), проводящие, механические (опора) 3. Формируются многоклеточные органы размножения, т.к. многоклеточные стенки надежнее защищают гаметы или споры 4. Зигота развивается в типичный многоклеточный зародыш 5. Задержка зиготы внутри архегония – еще одна функция защиты в условиях суши 6. У высокоорганизованных семенных растений приспособление к наземному образу жизни выразилось в полной независимости полового размножения от капельно-жидкой воды 7. У низших растений встречаются различные варианты полового процесса: изогамия (мужская и женская половая клетка одного размера подвижные), гетерогамия (женская половая клетка больше, чем мужская, обе подвижные), оогамия (женская половая клетка неподвижная, больше, чем мужская). У высших растений встречается только оогамия. 8. У высших растений у цикле развития всегда происходит чередование поколений (спорофит – бесполое поколение, гаметофит – половое поколение). На спорофите (2n) за счет мейотического деления образуются гаплоидные споры (n). Из спор развивается гаплоидный гаметофит (n), производящий гаплоидные гаметы. При их слиянии в зиготе восстанавливается набор хромосом. Из зиготы вновь развивается диплоидный спорофит (2n) Высшие растения разделяются на отделы:
studfiles.net
Двойное оплодотворение покрытосеменных растений
Особенности двойного оплодотворения покрытосеменных растений
Оплодотворение - это сложный процесс, состоящий из двух основных фаз по Батыгиной (1987) - прогамной (от попадания пыльцы на рыльце до вхождения пыльцевой трубки в зародышевый мешок) и непосредственно двойного оплодотворения; из трех фаз по Dumac (2008) - опыление, прогамной фазы (от попадания пыльцы на рыльце до встречи спермия с яйцеклеткой) и сингамии или двойного оплодотворения.До сих пор не установлено, что побуждает зиготу к неравномерности первого раздела [Kranz, 2008]. Исследование оплодотворения in vitro может способствовать решению этого вопроса.Недавно была показана активность мужского генома кукурузы в изолированных зиготах и молодых проэмбрио, с указанием отличия у разных растений относительного вклада мужских гамет после оплодотворения. Это объясняет раннее появление высокого уровня гетерозиса у зародышей кукурузы, требующие активности обоих гибридных геномов [Meyer, 2007].У кукурузы была создана кДНК библиотека с использованием клеток спермиев [Engel, 2003]. Белок HAP2/GSC1 - является первым идентифицированным белком поверхности спермиев цветковых растений, что является существенным для двойного оплодотворения. LGC1, HAP2/GSC1 и AtGEX2 [Singh, 2008].Кукуруза считается удобной моделью для исследований многих процессов в культуре in vitro, в том числе явлений двойного оплодотворения покрытосеменных растений, эмбриогенеза, эндоспермиогенеза и проч. [Dumac, 2008; Singh, 2008]. Кукуруза является удачным объектом генетических исследований для изучения процессов, механизмов гаметогенеза и развития семян, которые связаны с пролиферацией, дифференциацией и выживанием клеток [Lorbiecke, 2005]. Клетки женского и мужского гаметофита кукурузы могут быть изолированы и использованы для изучения экспрессии генов на низком транскриптом уровне [Sauter, 1998].

Автор фото (продольный срез зерновки кукурузы, 6 дней после опыления): Ляпустина Е.В.
Автор статьи: Ляпустина Е.В.
bio-x.ru
Инструктивная карточка к уроку биологии "Двойное оплодотворение покрытосеменных растений"
Двойное оплодотворение у покрытосеменных растений
1. Строение цветка. Рассмотрите строение цветка, вставьте пропуски в предложения.
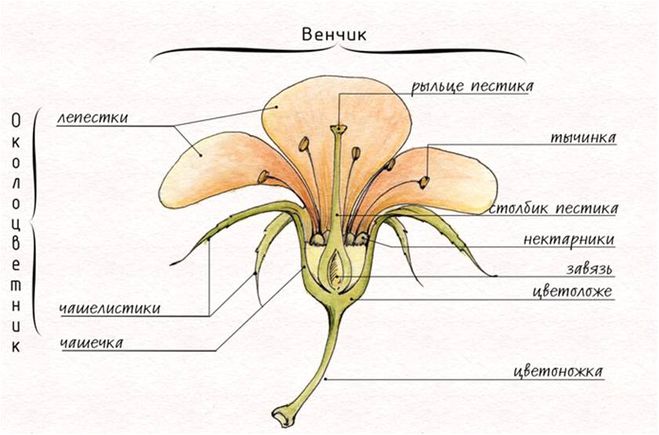
Цветок является органом ______ размножения. Он представляет собой видоизмененный _______. Главными (половыми) частями цветка являются мужские________ и женский_______. Пестик состоит из _______, _______ и _________. Тычинка состоит из ____________ и ____________. Вокруг тычинки и пестика расположен околоцветник. Он состоит из лепестков, объединенных в __________, а также __________ и _______.
2. Виды цветков.
Цветки
обоеполые раздельнополые
(?)
? ?
3. Двойное оплодотворение.
Процесс двойного оплодотворения был открыт русским ученым С.Г. Навашиным.
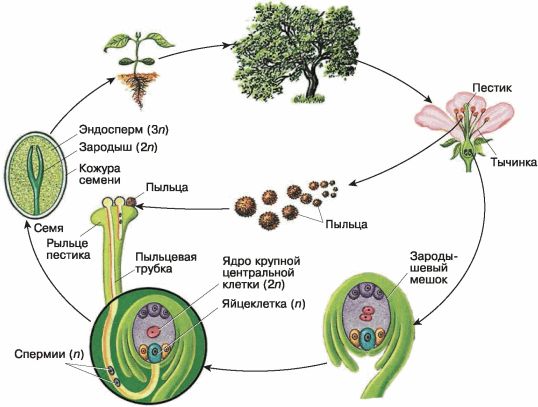
А) Рассмотрите рисунок. Что представляет собой спорофит покрытосеменных растений? Чем представлен мужской гаметофит? Чем представлен женский гаметофит?
Б) рассмотрите строение женского гаметофита. Сколько клеток в нем образуется? Какой набор хромосом они имеют? Какие функции выполняют?
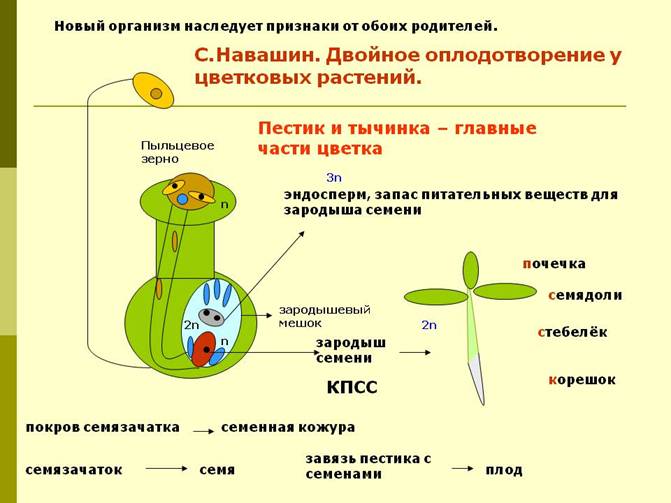
В) Рассмотрите схему двойного оплодотворения. Опишите, что происходит с двумя мужскими спермиями, попавшими на рыльце пестика?
Вставьте пропуски в предложения: Один из спермиев оплодотворяет ____________ и образуется _________ с набором хромосом (2n). Второй спермий сливается с ___________ и образуется __________ с набором хромосом (3n). Он выполняет функции ___________.
infourok.ru
Двойное оплодотворение
Пыльцевые зерна, попав на рыльце пестика, поглощают воду, сахара и другие питательные вещества и образуют трубку. Трубка буквально прорастает в пестик до зародышевого мешка. Пыльцевая трубка проникает в зародышевый мешок, где одна мужская гамета соединяется с яйцеклеткой, образуя зиготу. После митотического деления зигота становится зародышем будущего семени. Вторая мужская гамета сливается с двумя полярными ядрами и образует эндосперм. Этот процесс известен как двойное оплодотворение.[ ...]
Двойное оплодотворение представляет собой исторически сложившееся, качественно новое явление, свойственное только покрытосеменным растениям. Биологическое значение двойного оплодотворения заключается в том, что эндосперм, обогащенный двойной наследственностью, повышает жизнеспособность и приспособленность покрытосеменных, обеспечивает их преимущество перед другими растениями в современную геологическую эпоху.[ ...]
Двойное оплодотворение — тип оплодотворения, свойственный покрытосеменным растениям, при котором один из спермиев сливается с яйцеклеткой с образованием диплоидной зиготы, дающей начало зародышу семени, а второй спермпй сливается с диплоидным ядром зародышевого мешка с образованием клетки, дающей начало эндосперму семенп.[ ...]
После двойного оплодотворения первым обычно делится первичное ядро эндосперма (рис, 124). У пшеницы его деление начинается через 3—4 ч после опыления. В это время в цитоплазме зародышевого мешка и в первичном ядре эндосперма интенсивно накапливаются нуклеиновые кислоты (РНК и ДНК), белки, полисахариды, ферменты, витамины и другие физиологически активные вещества. С момента образования зиготы и до полного формирования зародыша эндосперм служит основным источником его питания.[ ...]
Впоследствии из оплодотворенной яйцеклетки (зиготы) образуется зародыш, а из оплодотворенного ядра центральной клетки — эндосперм. Двойное оплодотворение присуще всем систематическим группам покрытосеменных растений.[ ...]
Постмитотический тип двойного оплодотворения детально исследован Е. Н. Герасимовой-Навашиной у рябчика горного (Fritillaria tenella) (рис. Она же изучала промежуточный тип двойного оплодотворения у гальтонии. Премитотический процесс оплодотворения у пшеницы показан на рисунках 111, Л и 113.[ ...]
Как было отмечено выше, оплодотворение у цветковых (покрытосеменных) имеет существенную отличительную особенность в виде двойного оплодотворения (С. Г. Навашин, 1896), которое сводится к тому, что в зародышевом мешке гаплоидная яйцеклетка и диплоидная центральная клетка оплодотворяются спермиями, в результате чего образуется диплоидный зародыш и триплоидная клетка, развивающаяся в клетки эндосперма (см. гл. II).[ ...]
Герасимова-Навашнна Е. Н. Двойное оплодотворение покрытосеменных и некоторые теоретические аспекты//Проблемы эмбриологии.— Киев: Наукова думка, 1971.[ ...]
Правильным представлением об оплодотворении у покрытосеменных мы обязаны выдающемуся русскому цитологу-эмбриологу С. Г. Навашину, открывшему в 1898 г. двойное оплодотворение у лилейных и астровых. Он впервые показал, что у цветковых растений при оплодотворении один из спермиев сливается с ядром яйцеклетки, а другой — с ядром центральной клетки или полярными ядрами (рис. 109).[ ...]
У плодовых растений после завершения двойного оплодотворения из с.емяпочки формируется семя, а наружные покровы семяпочки (или интегументы) превращаются в кожуру семени. Клетки нуцеллуса используются растущим зародышем или реже превращаются в питательную ткань — перисперм. У болв-шинства плодовых и ягодных растений эндосперм и клетки нуцеллуса идут на формирование зародыша, а запасные пластические вещества локализуются в семядолях зародыша (рис. 28).[ ...]
При изучении структуры спермиев в процессе двойного оплодотворения С. Г. Навашин обнаружил, что их ядра находятся в состоянии поздней телофазы. Механизм движения спермиев в пыльцевой трубке и зародышевом мешке он рассматривал как активный процесс самостоятельного передвижения.[ ...]
Весь ;>тот процесс, как известно, принято намывать двойным оплодотворением. Ведь согласно общепринятому в литературе определению, оплодотворение (еипгамин) — это процесс слияния, мужской п женской половых клеток (гамет) с образованием зиготы, из которой и дальнейшем разнимается новый организм. Такое определение оплодотворения можно найти в любом учебнике биологии и в любом :нщнклопедичоском словаре (в том число в советских шщпклопедиях, в Большой и Малой). И даже в известном «Словаре русского языка» С. И. Ожегова (1973) мы читаем: «Оплодотворить. 1. Создать зародыш в ком-чем-н. слиянием мужской и женской половых клеток. 2. Послужить источником развития, совершенствования». Слияние одного из сиормпев с яйцеклеткой является, несомненно, оплодотворением, но тройное слияние по является, строго говоря, оплодотворением, так как 1) центральная клетка — не гамета и 2) в результате ¡»того слияния но образуется зигота, из которой и дальнейшем развивался бы новый организм. Очевидно, тройное слияние является оплодотворением лишь в указанном Ожеговым, втором, переносном смысле. Другими словами, в выражении «двойное оплодотворение» термин «оплодотворение» применяется в двух разных, смыслах — прямом и переносном. Тем не менее выражение «двойное онлодотиороние» настолько широко вошло в литературу, что было бы нецелесообразно его заменить (а попытки такого рода делались, в том числе известным, немецким ботаником. Достаточно, если мы будем помнить, что речь идет здесь о двух разных биологических процессах, условно объединяемых общим .названием.[ ...]
Открытие в 1896 г. выдающимся русским ученым Сергеем Гавриловичем Навашииым двойного оплодотворения у покрытоременных растений положило начало новой .эре исследований в биологической науке. С. Г. Навашин является также основателем науки о ядре — кариологии.[ ...]
Детально изучая поведение спермиев в зародышевом мешке, Е. Н. Герасимова-Навашина предложила различать два основных типа двойного оплодотворения: премитотический, когда объединение половых ядер происходит перед первым митозом зиготы, и постмитотический, когда объединение половых ядер наступает в начале первого митоза зиготы. Ею установлена также промежуточная форма двойного оплодотворения, характеризующаяся образованием собственной оболочки вокруг ядра спермия при медленном его погружении в ядро яйцеклетки. При этом объединение половых ядер происходит во время первого митоза зиготы (рис. 110).[ ...]
Рост семенного растения начинается с прорастапия семени. Семена у покрытосеменных растений образуются из семяпочки в результате двойного оплодотворения. В пыльце сосредоточено большое количество физиологически активных веществ — фитогормонов (ауксинов), ферментов, много аминокислоты пролина. Все ето приводит к тому, что уже сразу после опыления интенсивность обмена веществ в системе пыльца — пестик резко усиливается. Пыльцевая трубка несет два слермия. В результате слияния первого спермия с яйцеклеткой образуется зигота, дающая при последующем развитии зародыш семени. Анатомически ткани зародыша целиком состоят из первичной меристемы. Оно начинает делиться и вместе с цитоплазмой образует множество клеток, составляющих эндосперм. После опыления и особенно оплодотворения завязь начинает разрастаться. Вместе с пыльцой в зародышевый мешок привносятся как сами фитогормоны (ауксины), так и ферменты, катализирующие синтез гормонов. Дыхание развивающегося после оплодотворения семени идет очень интенсивно. Семя становится центром притяжения питательных веществ. В процессе развития и роста зародыша ткань вндосперма потребляется и в моменту созревания семян может частично или полностью исчезнуть. Некоторые семена лишены андосперма, питательные вещества сосредоточены у ттнт в тканях самого зародыша (в семядолях).[ ...]
У Crépis capillaris ядра спермиев при слиянии с женскими половыми ядрами находятся в периоде G прерванного митотического цикла (стадия телофазы), который они заканчивают в зиготе (рис. 112, 118). У астровых и злаковых в момент оплодотворения спермии находятся в стадии поздней телофазы митоза. На рисунке 114 показан конец двойного оплодотворения у кукурузы в момент слияния мужских ядрышек с женскими в ядрах зиготы и первичного ядра эндосперма; при этом второй сперматозоид находится в полярных ядрах; в яйцеклетке процесс слияния уже закончен, присутствует ядрышко спермия; слева виден контур второй пыльцевой трубки со спермиями.[ ...]
ru-ecology.info






