Ростовые движения растений. Движения растений
Движения растений. Способы, предпосылки возникновения и значение
Способность растений к движениям давно привлекала внимание ученых.
Особенно оно усилилось после публикаций работ Ч. Дарвина «Движения и повадки лазающих растений» (1865), «Насекомоядные растения» (1875) и «Способность к движениям у растений» (1880), в которых он рассмотрел широкий круг явлений из жизни растений под углом зрения их приспособительного значения. Он также поставил проблему движений растений на экспериментальную основу, показал возможность их строгого анализа. Тем самым дарвинизм оказал плодотворное влияние на развитие исследований в этой области (Н. Г. Холодный, 1941).
Под термином движение растений (фитодинамика) объединяют разнородные по механизмам и значимости явления: движение цитоплазмы и органоидов клетки, передвижение с помощью жгутиков (монады, хлорококковые), рост растяжением, изменение положения прорастающих семян, круговые нутации, тропизмы, ростовые и тургорные настии и т. п. (Э. Бюннинг, 1960; А. И. Меркис, 1973; Э. Либберт, 1976; А. Гэлстон, и др., 1983; Ф. Уоринг, Ф. Филлипс, 1984). Их рассмотрение занимает большое место в учебнике «Физиология растений» (В: В. Полевой, 1989), где они описаны не только с точки зрения проявления и природы, но и происхождения и эволюции.
Некоторые из названных форм движений характерны и для животных (движение цитоплазмы и жгутиками), остальные встречаются почти у всех растений. Однако в зависимости от систематического, филогенетического и ценотического положения видов доминируют или круговые нутации, или тропизмы, или настии, причем меняются способ и степень осуществления каждой из форм и в онтогенезе. При доминировании одной из них другие не исчезают, а сохраняются в виде дополнительных или в сочетаниях. Универсальное значение для растений, особенно высших наземных, имеют обратимые тургорные движения и настии. Сейсмонастии по механизму близки к мышечным сокращениям.
Не останавливаясь на анализе механизмов разных форм движений (В. В. Полевой, Т. С. Саламатова, 1985; В. В. Полевой, 1989), подчеркнем, что даже на клеточном уровне они далеко не совпадают, не говоря уже об уровне органов и целостного растения. В адаптивном плане разные способы движений также неравноценны. Некоторые из них (движение цитоплазмы и органоидов, круговые нутации) имеют более общее значение, другие более специфичны (лазающие, вьющиеся растения, сейсмо- и никто- настии) в экологическом отношении.
Внимание Ч. Дарвина к движениям было вызвано и в связи с возражением Ст. Д. Майварта о том, что принцип естественного отбора не объясняет «начальные стадии полезных черт», в том числе и у вьющихся растений. Эту способность растений Ч. Дарвин объяснил как результат усиления под действием естественного отбора «общей способности к движению, приобретенной ради совершенно иных и полезных целей», которая не представляла первоначально для них особой пользы. Далее он объясняет, что видоизменением круговой нутации были «приобретены многие весьма полезные и необходимые движения» растениями. Наблюдения над усиками Bignonia capreolata, Echinocystis lobata, Passiflora gracilis, Sicyos angulatus убедили Ч. Дарвина не только в высокой их чувствительности, но и в специфике реакции на раздражитель.
Возможность возникновения специальных форм движений на основе простой двигательной реакции под действием естественного отбора не является невероятной в случае их выгоды в борьбе за существование. В сомкнутых фитоценозах (лес) при недостатке освещения, в Поиске других ресурсов (воды, минеральных элементов и т. п.) усилением зачаточного ростового движения в разных целях могут сложиться вьющиеся и лазающие формы, различные способы ориентации в пространстве (тропизмы и настии) или другие обратимые движения у растений. В борьбе за существование путем параллелизмов и конвергенции возникают сходные формы движений у представителей различных семейств высших растений. Экологически значимы нутации, настии, тропизмы, насекомоядность (В. Е. Tuniper et al., 1989) для растений независимо от их систематического положения. Без них сложно представить стабильность растений. Несколько труднее объяснить роль сейсмонастий у мимозы и ряда других растений, хотя рассматривают их как предотвращение повреждений от воздействия сильных ветров и ливней.
Так или иначе способность к движениям — основа ориентации в пространстве. Она важна и при прикрепленном к субстрату состоянии, когда у растения нет возможности активно изменить положение по отношению к односторонне действующему фактору. Поэтому почти все формы движения растений представляют целесообразную реакцию на среду (Н. Г. Холодный). Они повышают экологическую стабильность растений, обеспечивая максимальное использование среды путем движения органов и структур — «поисковая» активность индивидуума. В связи с этим происходит усиление контроля целостности организма над способностью к движениям растений через механизмы гормональной регуляции и активности Н+-помпы. Отбор ведет к возникновению подобных механизмов. Заслуга Ч. Дарвина состоит в объяснении экологической роли движений растений в связи с действием естественного отбора.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Движение растений
Растения в отличие от животных прикреплены к месту своего обитания и не могут перемещаться. Однако и для них характерно движение. Движение растений — это изменение положения органов растений в пространстве, обусловленное разными факторами внешней среды: светом, температурой, силой тяжести, химическими элементами и др.
Движения растений относятся к быстротекущим процессам. В основе движений лежит раздражимость растений. Движения могут иметь и эндогенный характер.
Для цветкового растения свойственно главным образом движение органов путем изгиба, скручивания, откидывания и т. д., т. е. движение органов, неподвижно прикрепленных к субстрату. Среди этих движений есть движения, связанные с изменением тургора в отдельных клетках и тканях, а также с изменением скорости роста различных сторон, частей и органов растений. К первым относятся «сон» растений, обусловленный движением листьев или лепестков, сейсмонастия и другие явления. Ко вторым относятся тропизмы и настии.
Тропизмы — это ростовые движения, вызываемые односторонним действующим раздражителем (свет, сила тяжести, химические элементы и др.). Изгибание растений в сторону действующего фактора (раздражителя) называют положительным тропизмом, а в противоположную от действующего фактора сторону — отрицательным.
В зависимости от природы раздражителя, вызывающего изгиб, каждый конкретный тропизм получил соответствующее название. Если изгиб вызван действием света — фототропизм, силой тяжести — геотропизм, неравномерным распределением воды в почве — гидротропизм, химических соединений (удобрений) — хемотропизм, кислорода — аэротропизм и т. д.
Фототропизм
Многолетними исследованиями выяснено, что к органам растений, отклоняющимся в сторону источника света (положительный фототропизм), относятся молодые побеги и листья. Свет оказывает влияния на направление роста корней. Отрицательно фототропны зародышевые корешки растений, усики, гипокотили и другие части растений.
Во время развития растений фототропизм его органов может изменяться. Например, до оплодотворения цветоножки некоторых растений проявляют положительный фототропизм, а после оплодотворения — отрицательный. Изменение положительное фототропизма на отрицательный у одних и тех же органов может быть вызвано и интенсивностью освещенности. При относительно слабой освещенности отмечается положительный, а при сильной — отрицательный фототропизм.
Благодаря изгибаньям, обусловленным фототропизмом, органы растений занимают наиболее выгодное положение. В результате изгибания стеблей возникает листовая мозаика, которая способствует оптимальному использованию света при фотосинтезе (рис. ).
Способность растении располагать пластинки листьев перпендикулярно падающим лучам называют диафототропизмом, а расположение листьев под острым или тупым углом — плагиотропизмом. Листовая мозаика особенно отчетливо видна у тенелюбивых растении.
Как же осуществляются фототропные движения?
Родоначальником учения о фототропизме считают Ч. Дарвина. Он выяснил, что световое раздражение воспринимается верхушкой, ответная реакция (изгиб) возникает в зоне растяжения, лежащей ниже. Следовательно, клетки, воспринимающие световое раздражение (рецептор), и клетки, отвечающие на это раздражение, пространственно разобщены. Это дало ему возможность предположить существование веществ, которые синтезируются в одном месте, а передвигаются в другое (типа гормонов у животных).
В 1928 г. физиологи Н. Г. Холодный и Ф. В. Вент независимо друг от друга сформулировали гормональную теорию фототропизма, которая пользуется всеобщим признанием. В ее основе лежит представление о фототропизме как о процессе, стимулирующем перераспределение гормонов в верхушке побега. Одностороннее освещение вызывает электрическую поляризацию тканей верхушки: освещенная сторона получает отрицательный заряд, а затененная — положительный. Под влиянием этой поляризации ток гормона смещается на затененную сторону, клетки этой стороны растягиваются сильнее, чем освещенной, в результате получается изгиб в сторону света.
Выяснено, что гормонами, вызывающими рост клеток при фототропической реакции, являются ауксины или -индолилуксусная кислота. На рисунке 133 показана схема движения ауксина на колеоптиле овса при действии света. При одностороннем действии света на колеоптиль (Б) происходит поперечное перераспределение ауксинов в неосвещенную сторону (б), что и приводит к изгибу колеоптиля (5). В случае, когда поперечный поток ауксина прерывается (Г, в) при действии одностороннего света изгиб не наблюдается (Д). Неравномерное распределение ауксина оказывается очень важным, но не единственным следствием поперечной поляризации.
Возникают также физико-химические различия, связанные с физиологией обмена веществ, в частности с изменением содержания сахара и концентрации ионов водорода в клетках освещенной стороны. Химическая природа других веществ пока еще неизвестна. В развитие теории Холодного—Вента по данным последних исследований следует признать и роль ингибиторов, их соотношение и баланс с ауксином, что принято во всех процессах, в основе которых лежит рост растений (М. X. Чайлахян, О. Н. Кулаева, 1967).
Исследования показали, что спектр действия фототропизма определяется и качеством света. Фототропизм лучше проявляется в УФ области и в синей части спектра. У семенных растений фототропическая реакция вызывается коротковолновыми лучами. Фототропическая реакция зависит и от интенсивности освещения: чем слабее свет, тем дольше нужно освещать растения для получения фототропического эффекта. Произведение силы света на продолжительность его воздействия является величиной постоянной. Эта зависимость получила название закона количества раздражения.
Фототропическая чувствительность может не проявиться, если освещать растения сразу сильным светом после их проращивания в темноте. Такое же действие оказывают хлороформ и эфир.
В фототропической реакции свет выступает как раздражитель, необходимый лишь для запуска различных физиологических процессов; при этом расходуется малое количество энергии. Для ростовых процессов требуется много энергии.
studfiles.net
Движения у растений | Мир Знаний
Некоторые растения способны совершать активные движения, заметные для человеческого глаза. У одних видов листья могут ловить насекомых, у других — складываться от прикосновений.
На востоке США, на болотах между Южной и Северной Каролиной, встречается удивительное растение — венерина мухоловка. Его листья состоят из двух створок с длинными зубцами по краям. Поверхность створок унизана множеством тонких чувствительных волосков. Если насекомое касается этих волосков, створки листа быстро закрываются, а зубцы смыкаются, образуя решетку, которая не дает жертве вырваться на свободу.

Венерина мухоловка
Ловчие листья помогают венериной мухоловке жить на болотах. Бедные кислые почвы, на которых она растет, не обеспечивают ее достаточным количеством питательных веществ. Чтобы восполнить этот недостаток, растение научилось ловить насекомых.
Листья венериной мухоловки унизаны красноватыми железками, выполняющими различные функции. В центре листа расположены пищеварительные железы, по краям — нектарники. Ярко окрашенный лист со сладким нектаром — лакомая приманка для насекомых. Но стоит усевшейся на него жертве дотронуться до чувствительных волосков в центральной части, как она оказывается в ловушке, где ее ждет неминуемая гибель.
Дней через десять, когда насекомое будет переварено, створки листа снова раскроются. Порыв ветра унесет остатки покровов добычи, освободив в ловушке место для новой жертвы.
Пассивные и активные движения
Растения постоянно совершают движения под влиянием ветра и других внешних факторов. Такие движения можно назвать пассивными, потому что они не требуют никаких затрат энергии. Пассивные движения играют важную роль в размножении растений, способствуя опылению или расселению семян. Когда весной у некоторых растений раскрываются пыльники, поднявшиеся в воздух клубы пыльцы вызывают у многих людей аллергическую реакцию (сенную лихорадку). Активные движения, требующие затрат энергии, встречаются в мире растений довольно редко.
Многие одноклеточные водоросли снабжены жгутиками, биения которых помогают им передвигаться в толще воды. Сложно устроенные многоклеточные растения способность к движению утратили. Приспосабливаясь к жизни на суше, они обзавелись корнями, которые «намертво» закрепили их в почве. Но под влиянием внешних факторов у ряда растений, зачастую не состоящих в близком родстве, в процессе эволюции возникла способность к активным движениям.
Стыдливое растение
Европейцы, впервые увидевшие мимозу в Бразилии в XVI в., усмотрели в ее поведении признаки стыдливости: от малейшего прикосновения она складывает и опускает листья. За эту особенность растение и получило свое научное название — мимоза стыдливая (Mimosa pudica).
Мелкие розовые цветки мимозы собраны в соцветия-головки. Она высоко ценится садоводами и широко культивируется в районах с мягким климатом. Листья у нее сложные, то есть состоят из множества листочков. Если прикоснуться к одному из них, он складывается вдоль жилки, а затем такое же движение совершают и остальные листочки. От сильной встряски листья могут сложиться на всем растении. При этом оно выглядит увядшим — будто все его листья внезапно погибли. Эта реакция может передаваться даже от одного растения мимозы другому — соседнему.
По-видимому, чувствительность к прикосновениям выработалась у мимозы как приспособление для защиты от травоядных животных. Со сложенными листьями она выглядит засохшей, и млекопитающее, собравшееся их пощипать, предпочтет мимозе соседнее, более свежее на вид растение.
Способность к обучению
Помимо способности к быстрым активным движениям, у мимозы есть и другие особенности, свойственные животным. Например, ей можно сделать анестезию эфиром или хлороформом, и на несколько часов она совсем потеряет чувствительность к прикосновениям и прекратит складывать листья. Более того, это удивительное растение способно к своего рода обучению!
Если мимозу регулярно подвергать воздействию одного и того же раздражителя, она постепенно начнет реагировать на него все слабее, а через несколько часов и вовсе перестанет складывать листья. Если на время раздражение растения прекратить, а затем возобновить его, мимоза вновь будет реагировать на раздражитель, правда не так сильно, как вначале. Словно она помнит о полученном опыте!
Японские ученые установили, что способностью к движениям мимоза обязана белку актину, благодаря которому сокращаются и наши мышцы. А воздействие раздражителей мимоза воспринимает с помощью особых осязательных волосков на листьях, у основания каждого из которых расположена группа тонкостенных чувствительных клеток.
340mir-znaniy.com
|
Поиск Лекций
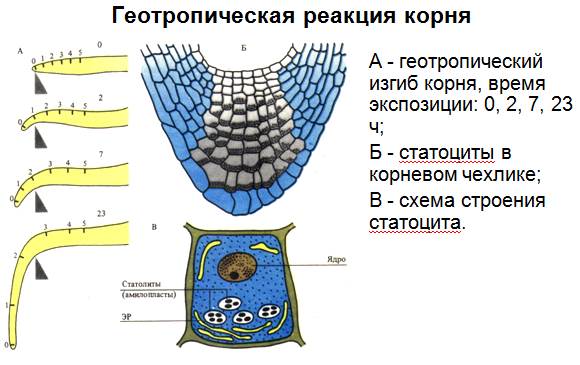
Сравнивая растительный и животный организмы, обычно говорят, что в отличие от животных растения не передвигаются, прикреплены в течение жизни к определенному месту. Однако движение как одно из свойств живой материи характерно и для растений. Движениями растений называют изменения расположения их органов в пространстве, обусловленные разными внешними факторами. Органы прикрепленных растений изменяют свое положение в пространстве благодаря росту, точнее, растяжению клеток и колебаниям тургорного давления. Ткани, непосредственно участвующие в движениях, имеют растяжимые и относительно тонкие клеточные стенки. К таким клеткам относятся, например, тонкостенные клетки основной паренхимы; клетки, находящиеся в основании черешков (подушечках) листьев бобовых растений; моторные клетки в эпидерме некоторых злаков, замыкающие клетки устьиц. Изменения тургорного давления происходят в результате транспирации, а также выхода воды из вакуоли в свободные пространства клеточных стенок и в межклетники. Однако нередко вместе с водой выходят и растворенные вещества. Следовательно, существует тесная связь между движениями и водным обменом растения и содержанием в его клетках минеральных веществ. В основе движений лежит универсальное свойство живой материи – раздражимость. В зависимости от типа раздражителя движения высших растений делят на две группы: тропизмы и настии. Многие раздражители действуют на растение направленно, с одной стороны. Растение способно различать направление действия света или вещества, силы тяжести. В ответ на одностороннее действие внешних факторов побеги или корни изгибаются. В результате изменяется расположение органов в пространстве. Эти изменения положения органов, вызываемые односторонне действующим внешним раздражителем, получили название тропизмов. В зависимости от природы раздражителя (свет, сила тяжести, химические вещества, прикосновение, электрический ток, поранение) тропизмы называют фото-, гео-, хемо-, тигмо-, электро-, травмотропизмами. Характер ответной реакции может быть разным. Те органы, которые поворачиваются к раздражителю, называют положительно тропными, а те, которые отворачиваются от раздражителя, – отрицательно тропными. Тропизмы чаще связаны с более быстрым ростом клеток на одной стороне стебля, корня или черешка, реже – с изменением тургорного давления. Геотропизмом называют движения растений в результате воздействия силы земного притяжения. Корень растения обладает положительным геотропизмом: – он растет вниз, а стебель – отрицательным, ибо растет в противоположном направлении. Весьма характерны в этом отношении проростки, помещенные в горизонтальном положении во влажную среду, или деревья, растущие на склонах. У первых корень вскоре будет изгибаться вниз, а стебель – вверх. Вторые будут ориентированы в направлении радиуса Земли, а не перпендикулярно поверхности склона. Ориентация к горизонтальной плоскости сразу же проявляется у прорастающего семени и у укореняющегося черенка. Корень углубляется в почву, где добывают воду и минеральные вещества, стебель же поднимается вверх, вынося листья навстречу падающим солнечным лучам.
Объяснения тому факту, что при геотропизме определяющим воздействием является сила земного притяжения, были получены еще в начале XIX в. с помощью простого, но остроумного приема. Проростки растений помещались на вращающееся колесо и увлажнялись водой. Растения росли таким образом, что стебли и корни их располагались по радиусам в плоскости вращения колеса: корни по направлению к периферии, а стебли – к центру, Т.е. первые росли по направлению центробежной силы, а стебли – в обратном направлении. Растения не знали ни верха, ни низа. Рост их определялся только направлением центробежной силы, заменившей силу земного притяжения. Наиболее чувствительными к силе тяжести являются кончик корня и зона роста стебля, а у злаков – верхушка колеоптиля. Первым, кто открыл место восприятия силы земного притяжения, был Ч. Дарвин. В его опытах корень с удаленной точкой роста утрачивал способность реагировать на эту силу и продолжал расти в том же положении, в котором он находился. Голландский ученый Ф. Вент и советский академик Н.Г. Холодный установили, что если к поверхности среза прикрепить отрезанный кончик корня или верхушку колеоптиля злаков, то тот же корень снова приобретает способность к проявлению геотропической чувствительности. Было высказано предположение, что дело здесь заключается в наличии в кончике корня или верхушке колеоптиля злаков фитогормонов. При вертикальном положении растений ауксины из верхушек перетекают в зону растяжения равномерно по всем сторонам стебля и корня. При горизонтальном положении растения ауксин накапливается на нижней стороне стебля и корня. Объяснить это можно тем, что нижняя сторона органа приобретает положительный заряд, а верхняя сторона - отрицательный. Ауксин обычно перемещается в виде аниона и поэтому концентрируется на нижней стороне. Стебель имеет гораздо более высокий оптимум концентрации ауксина (примерно 10-5моль/ л), чем корень (10-10-10-11 моль/л). Обладая различной чувствительностью к концентрации ауксина, корень и стебель, поэтому совершенно по-разному относятся к повышенной концентрации ауксина на нижней стороне горизонтально ориентированного органа. При накоплении ростового вещества на нижней стороне стебля рост ее ускоряется и вызывает изгиб кверху, а та же концентрация в нижней стороне корня приводит к противоположному эффекту: рост клеток ингибируется, и корень изгибается вниз. Изложенная теория Вента - Холодного объяснила вопрос, над решением которого трудились ученые в течение более ста лет. Очевидно, что в процессе длительной эволюции сохранялись лишь те особи, у которых под действием силы тяжести наблюдалась указанная реакция на неравномерное распределение ауксина. Ростовые движения растений, вызванные односторонним действием света, получили название фототропизма.Если изгиб направлен в сторону света, то говорят о положительном, а в обратную сторону – отрицательном фототропизме. Положительный фототропизм стеблей и колеоптилей объясняется тем, что сторона органа, обращенная к свету, растет медленнее, чем противоположная. Механизм фототропической реакции также связан с ауксинами: на затененной стороне ауксина больше, чем на освещенной. Поэтому растяжение клеток на затененной стороне происходит быстрее, чем на освещенной, и растение поворачивается в сторону большей освещенности.
Воспринимают одностороннее действие света верхушки надземных частей растений. Корни и корневища редко реагируют на этот фактор. Но у некоторых растений, например из семейства крестоцветных, корни проявляют иногда и отрицательный фототропизм. Если в специальную фототропическую камеру с небольшимотверстием в боковой стенке поместить проростки, например злаков, то уже через несколько часов верхушка колеоптиля повернется в сторону луча света, а изгиб произойдет в зоне растяжения. Если же, как показал впервые Ч. Дарвин, надеть на верхушку колеоптиля непроницаемый для света колпачок, то фототропическая реакция не проявляется. Фототропизм имеет колоссальное значение в жизни растений. Именно благодаря нему листья принимают положение, наиболее благоприятное для осуществления процесса фотосинтеза. Листья многих растений занимают более освещенные места, располагаясь так, чтобы не затенять друг друга. Это явление, которое называется листовой мозаикой, отчетливо наблюдается у клена, вяза, орешника, плюща и многих других растений. Фототропические движения листьев особенно важны в затененных условиях. В значительной мере нормальный рост растений в комнатах, цехах заводов и фабрик и других закрытых помещениях обязан фототропизму. Некоторые растения с такой силой реагируют на изменение направления света, что следуют за движением солнца на небосводе (гелиотропизм). К таким растениям относятся хорошо известные подсолнечник, череда и некоторые другие. Не все лучи спектра одинаковы в проявлении фотопериодических изгибов. Наиболее активными являются сине-фиолетовые лучи. Желтая, красная, ультрафиолетовая части спектра в этом отношении неактивны. Хемотропическиеростовые изгибы связаны с односторонним действием тех или иных химических веществ. Наиболее ярким примером хемотропизма является рост корней в сторону больших концентраций питательных веществ в почве. Большое значение хемотпропизм имеет и для нормальной жизнедеятельности грибов и высших растений, относящихся к сапрофитам и паразитам. Их гифы и присоски растут в сторону источника органических веществ. Явление хемотропизма наблюдается и при прорастании пыльцы и росте пыльцевой трубки. В данном случае, очевидно, основное влияние оказывает наличие выделяемых созревающими яйцеклетками веществ, которые управляют движением пыльцевых трубок. В числе этих веществ находятся некоторые специфические органические соединения, а также бор и кальций. Отрицательный хемотропизм корней наблюдается в случаях воздействия на них различных ядов. Движения корней растений в сторону кислорода воздуха получило название аэротропизма,а в сторону более оптимальной влажности почвы – гидротропизма.Первый можно наблюдать в избыточно увлажненных или сильно уплотненных почвах, второй при недостатке воды в почве. Эти виды тропизмов имеют важное экологическое значение, обеспечивая существование растений в стрессовых условиях, вызываемых недостатком кислорода или воды. Под тигмотропизмом подразумевают ростовую реакцию растений на прикосновение. Особенно сильно выражен тигмотропизм стеблевых и листовых усиков цепляющихся растений. Сторона усика при соприкосновении с опорой растет медленнее противоположной, что и приводит к обвиванию усика вокруг опоры. Эти движения происходят с затратой энергии. Предполагают участие в этих процессах сократительных белков и фитогормонов. Различают также травмотропизмы,позволяющие избегать соприкосновения с острыми предметами или химическими веществами, вызывающими травмы, радиотропизмы – ростовые изгибы и движения относительно источника радиоактивного излучения и некоторые другие. Настия – это движение органов растения, вызываемое раздражителем, не имеющим строгого направления, а действующим равномерно на все растение. Факторами, вызывающими настические движения, являются изменения температуры, освещенности, влажности воздуха в течение суток, перед дождем или после него. Если тропизмы – это движения органов в ответ на изменения напряженности внешнего фактора в пространстве, то настии – это движения органов, возникающие под действием смены условий во времени. Названия настии, как и тропизмов, зависят от природы раздражителей, которые их вызывают. Различают термо-, фото-, хемо-, гидро-, тигмо-, сейсмо-, электро- и травмонастии. К настическим движениям способны лишь двусторонне-симметричные органы (листья, лепестки). Примером термонастииможет служить раскрывание цветков тюльпана при высокой температуре и закрывание при низкой. К фотонастическим движениям относятся раскрывание цветков кувшинки и соцветий одуванчика при хорошем освещении и закрывание при уменьшении количества света, поднимание листовых пластинок кислицы на свету и опускание в темноте. Наиболее распространенными являются никтинастические движения, т.е. вызываемые сменой дня и ночи, когда одновременно меняются условия температуры и освещения. Очень многие цветки открываются утром и закрываются на ночь или наоборот. Раздражителем, вызывающим никтинастическую реакцию, может быть изменение температуры или только интенсивности света, или одновременно температуры и интенсивности света. Никтинастические движения облегчают опыление цветков в благоприятную погоду и защищают внутренние органы цветка при неблагоприятных условиях. Разновидностью настических изгибов являются сеисмонастии, вызываемые прикосновением или сотрясением, например листьев стыдливой мимозы: клетки одной стороны сочленения теряют клеточный сок с последующим мгновенным падением тургора, сжиманием клеток и опусканием черешков и листочков сложных листьев. Передается возбуждение при помощи электрического импульса или особых гормонов.
Механизм настий до конца не изучен. Предполагают, что настии происходят благодаря неравномерному росту разных сторон лепестков или листочков околоцветника. Если быстрее растет верхняя сторона, то происходит эпинастия, в результате которой раскрываются цветки, почки, опускаются листья. Если быстрее растет нижняя сторона, то происходит гипонастия, в результате которой органы закрываются. Открывания и закрывания цветков могут повторяться в определенном ритме. При каждом новом открывании и закрывании происходит рост клеток соответственно верхней или нижней стороны. Если процессы повторяются много раз, то лепестки или листочки околоцветника значительно удлиняются. Неравномерный рост, возможно, обусловлен гормонами. Это доказывается с помощью опыта. Если срезать верхушку молодого растения томата и на поверхность среза нанести пасту с ИУК, то через некоторое время все оставшиеся листья опустятся (эпинастия). В основе сейсмонастий, а также изменения положения листьев при переходе от дня к ночи лежит быстрое изменение тургорного давления в клетках на различных сторонах органа. Изменения тургора в моторных клетках, расположенных на разных сторонах подушечек, вызываются увеличением или уменьшением концентрации в них ионов калия. Калий перемещается из сокращающихся клеток в растягивающиеся благодаря работе калиевых насосов. За ним поступает вода, и клетки набухают. Центральная вакуоль при этом распадается на множество мелких вакуолей. Тургорные движения обратимы. Лучше изучен механизм опускания листьев у стыдливой мимозы, у которой различают два механизма проведения возбуждения: химический и электрический. Химический механизм состоит в том, что раздраженные клетки выделяют вещество, передвигающееся по флоэме и паренхиме. Когда это вещество дойдет до подушечки, происходит ответная реакция (листочек складывается). Реакцию вызывает очень низкая концентрация этого вещества 10-8 г/мл. Предполагают, что этими веществами являются аминокислоты, например, глутамат и аланин или малат. При электрическом механизме проведения возбуждения сразу после прикосновения в ее клетках возникает потенциал действия, равный -140 мВ (потенциал покоя равен -160 мВ). Потенциал действия распространяется от клетки к клетке по проводящим пучкам со скоростью 2-5 см/с. Он передается в подушечки у основания листочков и всего листа, в результате в моторных клетках уменьшается тургорное давление (вода выходит в межклетники), и лист складывается. Изменение тургорного давления зависит от ионов калия и хлора, концентрация которых в клетке регулируется ионными насосами. Возбуждение моторных клеток приводит к увеличению проницаемости плазмалеммы, ионы хлора и калия выходят, одновременно резко падает тургорное давление. Возможно, что в выделении воды участвуют и сократительные белки. На это указывает высокое содержание АТФ в моторных клетках, сильно уменьшающееся во время движения. |
|