
96. Структура АТФ, ее синтез. Роль АТФ в обмене веществ. Атф в растениях
Синтез АТФ в растениях
Энергия, выделяемая при движении электронов от Р680 (Е = –0,8) до Р700 (Е = +0,4 В), используется для синтеза АТФ из АДФ и неорганического фосфата (фотофосфорилирование).
Существуют несколько теорий, разъясняющих механизм фосфорилирования АДФ, сопряженный с работой электрон-транспортной цепи. Наибольшее признание в настоящее время получила хемиосматическая теория английского биохимика П. Митчелла (1961), которая для объяснения процессов фотосинтетического фосфорилирования впервые была использована А. Ягендорфом (1967).
Опыты А. Ягендорфа состояли в следующем (рис. 2.16). Изолированные из хлоропластов граны с рН 8 помещали в буферный раствор с рН 4. После выдерживании в этом растворе в гранах восстанавливалась величина рН, равная 4. Затем граны с рН 4 переносились в буферный раствор с рН 8. Таким образом, на тилакоидных мембранах искусственно создавался градиент протонов, который обычно формируется при фотохимических процессах (внутри тилакоида – рН 4, снаружи – рН 8). В этих условиях после добавления в среду АДФ и Фн в темноте синтезировалось АТФ.

Рис. 2.16. Схема эксперимента Ягендорфа
Эти эксперименты позволили применить хемиосмотическую теорию Митчелла для объяснения процессов происходящих при фотофосфорилировании в хлоропластах.
В соответствии с этой теорией пластохинон, присоединивший два электрона, присоединяет еще 2Н+ протона со стороны стромы хлоропласта и переносит их через мембрану во внутритилакоидное пространство. Протоны накапливаются внутри тилакоида и в результате фотоокисления воды.
Благодаря неравномерному распределению протонов по обеим сторонам мембраны создается разность химических потенциалов ионов водорода и возникает электрохимический мембранный потенциал ионов Н+ включает две составляющие: концентрационную, возникающую в результате неравномерного размещения ионов Н+ по обеим сторонам мембраны, и электрическую (ΔΨ), которая обусловлена мембранным потенциалом (рис. 2.16).
Энергия ΔрН и ΔΨ используется для обратного транспорта протонов из внутритилакоидного пространства в строму хлоропласта по особым каналам. С обратным транспортом протонов и связано фосфорилирование АДФ. Реакцию катализирует Н+-АТФ-синтетаза, состоящая из двух частей: водорастворимой каталитической части, расположенной в строме хлоропласта, и мембранной части. Последняя представляет собой протонный канал, по которому протоны могут возвращаться в строму хлоропласта. Фермент Н+-АТФ-синтетаза может синтезировать АТФ пока двигаются протоны. Протоны двигаются, когда их концентрация во внутритилакоидном пространстве большая.

Рис. 2.17. Иллюстрация гипотезы хемиосмотического сопряжения как механизма фотофосфорилирования
На каждые два электрона, переданных по электрон-транспортной цепи, внутри тилакоида накапливается 4Н+. На каждые 3Н+, возвращающихся назад в строму хлоропласта, синтезируется 1 молекула АТФ.
Таким образом, конечными продуктами световых реакций фотосинтеза является НАДФН и АТФ. Эти соединения используются затем соответственно как восстановительная сила и как источник для превращения СО2 в сахар. Этапы, из которых складываются эти превращения, известны под общим названием «темновых реакций» фотосинтеза.
biofile.ru
96. Структура АТФ, ее синтез. Роль АТФ в обмене веществ. Предмет, задачи и методы физиологии растений
Похожие главы из других работ:
Анатомия и физиология центральной нервной системы
4. Обмен жиров, их биологическая роль, теплоемкость, участие в обмене веществ. Энергетическая стоимость жиров. Жировые отложения
Жиры - органические соединения, входящие в состав животных и растительных тканей и состоящие в основном из триглицеридов (сложных эфиров глицерина и различных жирных кислот). Помимо триглицеридов, в состав жиров входят вещества...
Влияние органических удобрений на микробиоту почвы
2. Роль микроорганизмов в круговороте веществ в природе
Химическая деятельность микроорганизмов проявляется в непрерывном круговороте азота, фосфора, серы, углерода и других веществ. При самом активном, широком участии микроорганизмов в природе, главным образом в почве и гидросфере...
Гормон окситоцин
1. Химическая структура и синтез окситоцина
Окситоцин не является собственным гормоном нейрогипофиза, а лишь накапливается в нем, перемещаясь по аксонам гипоталамо- гипофизарного пучка из ядер переднего гипоталамуса - супраоптического и паравентрикулярного...
Естествознание на молекулярном уровне
3. Реакционная способность веществ, анализ и синтез
...
Естествознание на молекулярном уровне
3. Реакционная способность веществ, анализ и синтез
...
Зависимость уровня тиреотропного и тиреоидных гормонов от заболеваний щитовидной железы
2.5 Влияние веществ на синтез тиреоидных гормонов
В настоящее время считается, что влияния на синтез различных веществ имеет смешанный характер. Этот тезис доказывается в статье Р.В. Кубасова, Е.Д...
Микроорганизмы в круговороте веществ в природе
Роль микроорганизмов в круговороте веществ в природе
С помощью микроорганизмов органические соединения растительного и животного происхождения минерализуются до углерода, азота, серы, фосфора, железа и др. Круговорот углерода. В круговороте углерода активное участие принимают растения...
Микроорганизмы, выделенные из различных природных жиров
1.1 Структура жировых веществ
Жиры являются веществами нелетучими и при нагревании до 250-300°С разлагаются с образованием летучих веществ, выделяющихся в виде паров, газов и дыма. Жиры плохие проводники тепла...
Обмен белков. Обмен жиров. Обмен углеводов. Печень, ее роль в обмене веществ
Глава 4. Печень, ее роль в обмене веществ
...
Обмен белков. Обмен жиров. Обмен углеводов. Печень, ее роль в обмене веществ
4.3 Роль печени в обмене веществ
Рассматривая обмен белков, жиров и углеводов мы не раз затрагивали печень. Печень является важнейшим органом, осуществляющим синтез белков. В ней образуется весь альбумин крови, основная масса факторов свертывания...
Основные принципы питания
7. Роль минеральных веществ в питании человека
В зависимости от количества минеральных веществ в организме человека и в пищевых продуктах их подразделяют на макро- и микроэлементы. К первым относятся кальций, калий, магний, натрий, фосфор, хлор, сера...
Роль микроорганизмов в круговороте химических элементов в природе
4. Роль микроорганизмов в круговороте серы в природе, их значение превращения веществ и практическое использование
Круговорот серы осуществляется в результате жизнедеятельности бактерий, окисляющих или восстанавливающих ее. Процессы восстановления серы происходят несколькими путями. Под влиянием гнилостных бактерий - клостридий...
Фотосинтез как основа энергетики биосферы
4.2 Каротиноиды. Их структура, функции и физиологическая роль
Каротиноиды -- жирорастворимые пигменты желтого, оранжевого, красного цвета -- присутствуют в хлоропластах всех растений. Они входят также в состав хромопластов в незеленых частях растений, например в корнеплодах моркови...
Фотосинтез как основа энергетики биосферы
4.3 Фикобилины. Их структура, функции и физиологическая роль
Синезеленые водоросли (цианобактерии), красные морские водоросли и некоторые морские криптомонады помимо хлорофилла а и каротиноидов содержат пигменты фикобилины...
Энергетический метаболизм микроорганизмов
1. Общие понятия об обмене веществ и энергии
Все живые организмы могут использовать только химически связанную энергию. Каждое вещество обладает определенным запасом потенциальной энергии. Главные материальные носители ее химические связи...
bio.bobrodobro.ru
Строение АТФ и биологическая роль. Функции АТФ
В любой клетке нашего организма протекают миллионы биохимических реакций. Они катализируются множеством ферментов, которые зачастую требуют затрат энергии. Где же клетка ее берет? На этот вопрос можно ответить, если рассмотреть строение молекулы АТФ – одного из основных источников энергии.
АТФ – универсальный источник энергии
АТФ расшифровывается как аденозинтрифосфат, или аденозинтрифосфорная кислота. Вещество является одним из двух наиболее важных источников энергии в любой клетке. Строение АТФ и биологическая роль тесно связаны. Большинство биохимических реакций может протекать только при участии молекул вещества, особенно это касается пластического обмена. Однако АТФ редко непосредственно участвует в реакции: для протекания любого процесса нужна энергия, заключенная именно в химических связях аденозинтрифосфата.
Строение молекул вещества таково, что образующиеся связи между фосфатными группами несут огромное количество энергии. Поэтому такие связи также называются макроэргическими, или макроэнергетическими (макро=много, большое количество). Термин макроэргические связи впервые ввел ученый Ф. Липман, и он же предложил использовать значок ̴ для их обозначения.
Очень важно для клетки поддерживать постоянный уровень содержания аденозинтрифосфата. Особенно это характерно для клеток мышечной ткани и нервных волокон, потому что они наиболее энергозависимы и для выполнения своих функций нуждаются в высоком содержании аденозинтрифосфата.
Строение молекулы АТФ
Аденозинтрифосфат состоит из трех элементов: рибозы, аденина и остатков фосфорной кислоты.
Рибоза – углевод, который относится к группе пентоз. Это значит, что в составе рибозы 5 атомов углерода, которые заключены в цикл. Рибоза соединяется с аденином β-N-гликозидной связь на 1-ом атоме углерода. Также к пентозе присоединяются остатки фосфорной кислоты на 5-ом атоме углерода.
Аденин – азотистое основание. В зависимости от того, какое азотистое основание присоединяется к рибозе, выделяют также ГТФ (гуанозинтрифосфат), ТТФ (тимидинтрифосфат), ЦТФ (цитидинтрифосфат) и УТФ (уридинтрифосфат). Все эти вещества схожи по строению с аденозинтрифосфатом и выполняют примерно такие же функции, однако они встречаются в клетке намного реже.
Остатки фосфорной кислоты. К рибозе может присоединиться максимально три остатка фосфорной кислоты. Если их два или только один, то соответственно вещество называется АДФ (дифосфат) или АМФ (монофосфат). Именно между фосфорными остатками заключены макроэнергетические связи, после разрыва которых высвобождается от 40 до 60 кДж энергии. Если разрываются две связи, выделяется 80, реже – 120 кДж энергии. При разрыве связи между рибозой и фосфорным остатком выделяется всего лишь 13,8 кДж, поэтому в молекуле трифосфата только две макроэргические связи (Р ̴ Р ̴ Р), а в молекуле АДФ - одна (Р ̴ Р).
Вот каковы особенности строения АТФ. По причине того, что между остатками фосфорной кислоты образуется макроэнергетическая связь, строение и функции АТФ связаны между собой.
Строение АТФ и биологическая роль молекулы. Дополнительные функции аденозинтрифосфата
Кроме энергетической, АТФ может выполнять множество других функций в клетке. Наряду с другими нуклеотидтрифосфатами трифосфат участвует в построении нуклеиновый кислот. В этом случае АТФ, ГТФ, ТТФ, ЦТФ и УТФ являются поставщиками азотистых оснований. Это свойство используется в процессах репликации ДНК и транскрипции.
Также АТФ необходим для работы ионных каналов. Например, Na-K канал выкачивает 3 молекулы натрия из клетки и вкачивает 2 молекулы калия в клетку. Такой ток ионов нужен для поддержания положительного заряда на наружной поверхности мембраны, и только с помощью аденозинтрифосфата канал может функционировать. То же касается протонных и кальциевых каналов.
АТФ является предшественником вторичного мессенжера цАМФ (циклический аденозинмонофосфат) - цАМФ не только передает сигнал, полученный рецепторами мембраны клетки, но и является аллостерическим эффектором. Аллостерические эффекторы – это вещества, которые ускоряют или замедляют ферментативные реакции. Так, циклический аденозинтрифосфат ингибирует синтез фермента, который катализирует расщепление лактозы в клетках бактерии.
Сама молекула аденозинтрифосфата также может быть аллостерическим эффектором. Причем в подобных процессах антагонистом АТФ выступает АДФ: если трифосфат ускоряет реакцию, то дифосфат затормаживает, и наоборот. Таковы функции и строение АТФ.
Как образуется АТФ в клетке
Функции и строение АТФ таковы, что молекулы вещества быстро используются и разрушаются. Поэтому синтез трифосфата – это важный процесс образования энергии в клетке.
Выделяют три наиболее важных способа синтеза аденозинтрифосфата:
1. Субстратное фосфорилирование.
2. Окислительное фосфорилирование.
3. Фотофосфорилирование.
Субстратное фосфорилирование основано на множественных реакциях, протекающих в цитоплазме клетки. Эти реакции получили название гликолиза – анаэробный этап аэробного дыхания. В результате 1 цикла гликолиза из 1 молекулы глюкозы синтезируется две молекулы пировиноградной кислоты, которые дальше используются для получения энергии, и также синтезируются два АТФ.
- С6Н12О6 + 2АДФ + 2Фн ––> 2С3Н4O3 + 2АТФ + 4Н.
Окислительное фосфорилирование. Дыхание клетки
Окислительное фосфорилирование – это образование аденозинтрифосфата путем передачи электронов по электронно-транспортной цепи мембраны. В результате такой передачи формируется градиент протонов на одной из сторон мембраны и с помощью белкового интегрального комплекта АТФ-синтазы идет построение молекул. Процесс протекает на мембране митохондрий.
Последовательность стадий гликолиза и окислительного фосфорилирования в митохондриях составляет общий процесс под названием дыхание. После полного цикла из 1 молекулы глюкозы в клетке образуется 36 молекул АТФ.
Фотофосфорилирование
Процесс фотофосфорилирования - это то же окислительное фосфорилирование лишь с одним отличием: реакции фотофосфорилирования протекают в хлоропластах клетки под действием света. АТФ образуется во время световой стадии фотосинтеза – основного процесса получения энергии у зеленых растений, водорослей и некоторых бактерий.
В процессе фотосинтеза все по той же электронно-транспортной цепи проходят электроны, в результате чего формируется протонный градиент. Концентрация протонов на одной из сторон мембраны является источником синтеза АТФ. Сборка молекул осуществляется посредством фермента АТФ-синтазы.
Интересные факты об АТФ
- В среднестатистической клетке содержится 0,04% аденозинтрифосфата от всей массы. Однако самое большое значение наблюдается в мышечных клетках: 0,2-0,5%.
- В клетке около 1 млрд молекул АТФ.
- Каждая молекула живет не больше 1 минуты.
- Одна молекула аденозинтрифосфата обновляется в день 2000-3000 раз.
- В сумме за сутки организм человека синтезирует 40 кг аденозинтрифосфата, и в каждый момент времени запас АТФ составляет 250 г.
Заключение
Строение АТФ и биологическая роль его молекул тесно связаны. Вещество играет ключевую роль в процессах жизнедеятельности, ведь в макроэргических связях между фосфатными остатками содержится огромное количество энергии. Аденозинтрифосфат выполняет множество функций в клетке, и поэтому важно поддерживать постоянную концентрацию вещества. Распад и синтез идут с большой скоростью, т. к. энергия связей постоянно используется в биохимических реакциях. Это незаменимое вещество любой клетки организма. Вот, пожалуй, и все, что можно сказать о том, какое строение имеет АТФ.
fb.ru
Синтез АТФ в клетке
Аденозинтрифосфорная кислота-АТФ - обязательный энергетический компонент любой живой клетки. АТФ также нуклеотид, состоящий из азотистого основания аденина, сахара рибозы и трех остатков молекулы фосфорной кислоты. Это неустойчивая структура. В обменных процессах от нее последовательно отщепляются остатки фосфорной кислоты путем разрыва богатой энергией, но непрочной связи между вторым и третьим остатками фосфорной кислоты. Отрыв одной молекулы фосфорной кислоты сопровождается выделением около 40 кДж энергии. В этом случае АТФ переходит в аденозиндифосфорную кислоту (АДФ), а при дальнейшем отщеплении остатка фосфорной кислоты от АДФ образуется аденозинмонофосфорная кислота (АМФ).
Схема строения АТФ и превращения ее в АДФ (Т.А. Козлова, В.С. Кучменко. Биология в таблицах. М.,2000)
АДФ
Следовательно, АТФ - своеобразный
аккумулятор энергии в клетке, который "разряжается" при ее расщеплении.
Распад АТФ происходит в процессе реакций синтеза белков, жиров,
углеводов и любых других жизненных функций клеток. Эти реакции идут с
поглощением энергии, которая извлекается в ходе расщепления веществ.
АТФ синтезируется в митохондриях в несколько этапов. Первый из них - подготовительный - протекает ступенчато, с вовлечением на каждой ступени специфических ферментов. При этом сложные органические соединения расщепляются до мономеров: белки - до аминокислот, углеводы - до глюкозы, нуклеиновые кислоты - до нуклеотидов и т. д. Разрыв связей в этих веществах сопровождается выделением небольшого количества энергии. Образовавшиеся мономеры под действием других ферментов могут претерпеть дальнейший распад с образованием более простых веществ вплоть до диоксида углерода и воды.
Схема Синтез АТФ в мвтохондрии клетки
ПОЯСНЕНИЯ К СХЕМЕ ПРЕВРАЩЕНИЕ ВЕЩЕСТВ И ЭНЕРГИИ В ПРОЦЕССЕ ДИССИМИЛЯЦИИ
I этап - подготовительный: сложные органические вещества под действием пищеварительных ферментов распадаются на простые, при этом выделяется только тепловая энергия. Белки ->аминокислоты Жиры-> глицерин и жирные кислоты Крахмал ->глюкоза
II этап-гликолиз (бескислородный):
осуществляется в гиалоплазме, с мембранами не связан; в нем участвуют
ферменты; расщеплению подвергается глюкоза:
У дрожжевых грибов молекула глюкозы без
участия кислорода превращается в этиловый спирт и диоксид углерода
(спиртовое брожение):
У других микроорганизмов гликолиз
может завершаться образованием ацетона, уксусной кислоты и т, д. Во
всех случаях распад одной молекулы глюкозы сопровождается образованием
двух молекул АТФ. В ходе бескислородного расщепления глюкозы в виде
химической связи в молекуле АТФ сохраняется 40% анергии, а остальная
рассеивается в виде теплоты.
III этап-гидролиз (кислородный): осуществляется в митохондриях, связан с матриксом митохондрий и внутренней мембраной, в нем участвуют ферменты, расщеплению подвергается молочная кислота: СзН6Оз+ЗН20 -->3СО2+ 12Н. С02 (диоксид углерода) выделяется из митохондрий в окружающую среду. Атом водорода включается в цепь реакций, конечный результат которых - синтез АТФ. Эти реакции идут в такой последовательности:
1. Атом водорода Н с помощью ферментов-переносчиков поступает во внутреннюю мембрану митохондрий, образующую кристы, где он окисляется: Н-е-->H+ 2. Протон водорода H+ (катион) выносится переносчиками на наружную поверхность мембраны крист. Для протонов эта мембрана непроницаема, поэтому они накапливаются в межмембранном пространстве, образуя протонный резервуар.
3. Электроны водорода e переносятся на внутреннюю поверхность мембраны крист и тут же присоединяются к кислороду с помощью фермента оксидазы, образуя отрицательно заряженный активный кислород (анион): O2 + е-->O2-
4. Катионы и анионы по обе стороны мембраны создают разноименно заряженное электрическое поле, и когда разность потенциалов достигнет 200 мВ, начинает действовать протонный канал. Он возникает в молекулах ферментов АТФ-синтетаз, которые встроены во внутреннюю мембрану, образующую кристы.
5. Через протонный канал протоны водорода H+ устремляются
внутрь митохондрий, создавая высокий уровень энергии, большая часть
которой идет на синтез АТФ из АДФ и Ф (АДФ+Ф-->АТФ), а протоны H+ взаимодействуют с активным кислородом, образуя воду и молекулярный 02:( 4Н++202- -->2Н20+02)
Таким образом, О2,
поступающий в митохондрии в процессе дыхания организма, необходим для
присоединения протонов водорода Н. При его отсутствии весь процесс в
митохондриях прекращается, так как электронно-транспортная цепь
перестает функционировать. Общая реакция III этапа:
(2СзНбОз + 6Oз + 36АДФ + 36Ф ---> 6С02 + 36АТФ + +42Н20)
В результате расщепления одной молекулы глюкозы образуются 38 молекул АТФ: на II этапе - 2 АТФ и на III этапе - 36 АТФ. Образовавшиеся молекулы АТФ выходят за пределы митохондрии и участвуют во всех процессах клетки, где необходима энергия. Расщепляясь, АТФ отдает энергию (одна фосфатная связь заключает 40 кДж) и в виде АДФ и Ф (фосфата) возвращается в митохондрии.
www.examen.ru
Синтез атф в хлоропластах
Синтез АТФ в хлоропластах – это биохимический процесс, проводимый в мембранах тилакоидов, где происходят также реакции фотосинтеза и перенос протонов для транспортировки энергии. Хлоропласты для воспроизводства процессов перекачивают через себя протоны, используемые для преобразования световой энергии в движущую силу для проведения синтеза в клетке растения.
Фотосинтез и производство кислорода
Для проведения фотосинтеза в водорослях и листьях растений имеются специальные структурные образования – хлоропласты с мелкими отсеками-тилакоидами, используемые для воспроизводства кислорода. Хлоропласт – это энергетическая станция, вырабатывающая энергию для проведения реакций, переносов протонов, формирования АТФ (аденозинтрифосфата) и фотосинтеза.
Основные процессы протекают в мембранах тилакоидов, которые выполняют следующие функции:
- Расщепление молекул воды для выделения кислорода – фотосинтез.
- Транспортировку протонов, переносящих энергию.
- Синтез атф в хлоропластах – универсального энергетического поставщика, участвующего в проведении биохимических реакций.
Тилакоиды располагаются не только в хлоропластах растений, но и присутствуют в цианобактериях, не имеющих ядра и внутреннего разделения на клетки. По мнению учёных, в появлении хлоропластов сыграли немаловажную роль цианобактерии. У них схожее строение, мембрана из двух оболочек, присутствует РНК, ДНК и возможен синтез белка.
Цианобактерии являются самыми древними представителями жизни на планете, и именно им принадлежит первенство в переработке углекислого газа в кислород и воспроизводство синтезируемого АТФ в хлоропластах, которыми они являются. Многие придерживаются мнения, что именно расщепление молекулы воды на кислород и водород с помощью бактерии дало начало зарождения жизни на Земле.
«Фармконтракт-Синтез» является инжиниринговым подразделением ГК «Фармконтракт», занимается разработкой и внедрением проектов для проведения биохимических исследований и лабораторных испытаний в максимально приближенных к естественным условиям. Мы осуществляем поставку комплексов и систем для наблюдения за фотосинтезом, расщеплением клеток или образованием молекул. Нами осуществляется доставка и установка оборудования, валидация методов исследования, квалификационные испытания, модернизацию действующих систем или внедрение автоматики.
phct-synthesis.ru
17. Механизмы образования атф в клетках животных и растений:
а) Фотосинтетическое фосфорилирование – синтез АТФ у растений за счет квантов солнечной энергии.
б) Окислительное фосфорилирование – синтез АТФ за счет энергии, выделяющейся при окислении водорода субстрата кислородом с участием дыхательной цепи. Основной способ синтеза АТФ для большинства клеток.
в) Субстратное фосфорилирование – синтез АТФ за счет энергии гидролиза макроэргической связи субстрата (пример: фосфоглицераткиназная и пируваткиназная реакции анаэробного гликолиза).
Для митохондрий характерно ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ.
18. Окислительное фосфорилирование – синтез АТФ за счет энергии, выделяющейся при окислении водорода органических субстратов кислородом с участием дыхательной цепи.
Основные положения хемиосмотической теории Митчелла:
а) Энергия, выделяющаяся при транспорте электронов I, III и IV комплексами дыхательной цепи, используется для перекачивания протонов в межмембранное пространство, генерируя градиент pH.
б) Обратный поток протонов по протонным каналам АТФ-синтазы в матрикс обеспечивает энергией головку АТФ-синтазы для синтеза АТФ.
19. Метаболический путь – последовательность химических превращений конкретного вещества в клетке:
а) Циклический метаболический путь – замкнутая последовательность химических превращений, приводящая в итоге к регенерации исходного вещества. Примеры: цикл лимонной кислоты, орнитиновый цикл мочеобразования Кребса.
б) Линейный метаболический путь – линейная последовательность химических реакций. Примеры: гликолиз, пентозофосфатный путь.
Регуляторные ферменты – ферменты, регулирующие метаболизм клеток, которые обычно располагаются либо в начале метаболических путей, либо в местах ключевых разветвлений, где сходятся два и большее число путей и которые катализируют в клетке либо практически необратимые реакции, либо реакции, протекающие наиболее медленно. Примером может служить фосфофруктокиназа-1, ацетил-КоА-карбоксилаза, пируваткарбоксилаза, протеинкиназа.
20. Цикл Кребса – центральный метаболический путь.
|
| Цикл Кребса является центральным метаболическим путем, т.к. 1) связывает в клетке процессы катаболизма и анаболизма (пример: образующийся оксалоацетат используется в процессах глюконеогенеза, синтеза заменымых АК, цитрат – в синтезе жирных кислот и т.д.) 2) объединяет белки, жиры, углеводы, в нем сходятся почти все метаболические пути 3) является общим путем окончательного окисления большей части органических молекул в форме ацетил-КоА, играющих роль клеточного «топлива» 4) является основным путем, обеспечивающим энергией большинство клеток. |

21.
Энергетический баланс окисления ацетил~КоА: 1 ГТФ в ходе ЦТК + 3*3 АТФ (за счет НАД-переносчика) + 2 АТФ (за счет ФАД-переносчика) = 12 АТФ
22. См. вопрос 21.
Витамины, принимающие участие в работе цикла:
| Витамин | Кофермент в составе витамина |
| Тиамин | ТПФ |
| Никотиновая кислота | НАД |
| Рибофлавин | ФАД |
| Пантотеновая кислота | КоА |
Также для нормальной работы цикла необходимо витаминоподобное вещество – липоевая кислота.
Недостаток хотя бы одного из витаминов, используемых в синтезе небелковых компонентов ферментов цикла, сопровождается лактатным ацидозом и нарушением энергопродукции в клетке.
studfiles.net
Синтез АТФ — SportWiki энциклопедия
Анаэробный синтез АТФ: субстратное фосфорилирование, синтез АТФ из фосфокреатина, аденилаткиназная реакция[править]
АТФ (аденозинтрифосфат): молекула, обеспечивающая энергией живые клетки[править]

 Рис. 10.1. Строение аденозинтрифосфата (АТФ)
Рис. 10.1. Строение аденозинтрифосфата (АТФ) Молекулы АТФ жизненно необходимы. АТФ служит источником энергии, необходимой для сокращения мышц, проведения нервного импульса, протекания многих биохимических реакций и т.п. В покое расходуется 28 г (1 унция) АТФ в минуту, что эквивалентно 1,4 кг (3 фунта) в час, а при физической нагрузке расход АТФ достигает 0,5 кг в минуту! АТФ состоит из аденина, рибозы и трех фосфатных групп, которые называются а-, β- и у-фосфатными группами (рис. 10.1). Гидролиз высокоэнергетических фосфоангидридных связей между β- и у-атомами фосфора или между а- и β-атомами фосфора высвобождает энергию, необходимую для проведения биохимических реакций, т.е. для поддержания жизни организма.
Самый энергетически выгодный метод синтеза АТФ — это аэробное окислительное фосфорилирование. Однако АТФ может также образоваться в анаэробных условиях, хотя и с меньшей эффективностью. Существует три способа анаэробного синтеза АТФ: субстратное фосфорилирование, образование АТФ из фосфокреатина и в результате аденилаткиназной реакции. Хотя анаэробный синтез АТФ и не так энергетически выгоден, способность синтезировать АТФ в бескислородной среде может иметь жизненно важное значение.
Синтез АТФ путем субстратного фосфорилирования[править]
На рис. 10.2 показано, что АТФ образуется в процессе гликолиза в ходе фосфоглицераткиназной и пируваткиназной реакций, а также в цикле Кребса в ходе реакции, катализируемой ферментом сукцинил-КоА-синтетазой с участием нуклеозидцифосфаткиназы (рис. 10.3). Примечание: для этих реакций кислород не требуется.
-

Рис. 10.2. Образование АТФ в ходе гликолиза путем субстратного фосфорилирования
-

Рис. 10.3. В цикле Кребса путем субстратного фосфорилирования образуется ГТФ, который затем под действием нуклеозиддифосфаткиназы превращается в АТФ
Получение АТФ из фосфокреатина[править]
Фосфокреатин — это «аварийный запас» высокоэнергетического фосфата, который можно использовать для быстрого образования АТФ, необходимого для сокращения мышц в анаэробных условиях. Этот интенсивный механизм синтеза АТФ может в экстренных ситуациях спасти жизнь; однако запасы фосфокреатина расходуются очень быстро, всего за несколько секунд.
В периоды покоя, когда молекул АТФ много, креатинкиназа фосфорилирует креатин с образованием фосфокреатина. Особенно важна роль этой реакции в мышцах. Если вдруг требуется сделать резкий рывок, фосфокреатин фосфорилирует АДФ до АТФ, необходимого для сокращения мышц (рис. 10.4) По этой причине фосфокреатин еще называют «фосфаген».
 Рис. 10.4. Образование АТФ из фосфокреатина при физической нагрузке и синтез фосфокреатина из креатина в период покоя
Рис. 10.4. Образование АТФ из фосфокреатина при физической нагрузке и синтез фосфокреатина из креатина в период покоя Креатин выводится в форме креатинина[править]
Креатин — это аминокислота, не входящая в состав белков. Креатин синтезируется из аргинина и выводится с мочой в форме креатинина. Уровень креатинина в крови и клиренс креатинина используются для оценки скорости клубочковой фильтрации при нарушении работы почек. Примечание: не путайте креатин, креатинин и карнитин.
Прием креатина повышает работоспособность[править]
Эргогенные средства — это вещества, которые повышают скорость, силу или выносливость спортсмена. Многие из них опасны и запрещены к использованию. Мнения противоречивы, тем не менее многие ученые сходятся на том, что креатин — единственное эргогенное средство, для которого научно доказано его свойство повышать работоспособность как при спринтерских, так и при продолжительных нагрузках.
Образование АТФ из АДФ под действием аденилаткиназы[править]
После того как АТФ гидролизуется для высвобождения энергии, необходимой для сокращения мышц, в клетках образуется и накапливается АДФ. Но АДФ тоже содержит энергоемкую а-фосфоангидридную связь (рис. 10.1). Природа изобретательна: эта энергия становится доступной после того, как в анаэробных условиях две молекулы АДФ под действием аденилаткиназы образуют АТФ (рис. 10.5) (раньше фермент аденилаткиназу называли миокиназой).
 Рис. 10.5. Образование АТФ из двух молекул АДФ в процессе аденилаткиназной реакции
Рис. 10.5. Образование АТФ из двух молекул АДФ в процессе аденилаткиназной реакции Аэробный синтез АТФ[править]
Синтез АТФ в дыхательной цепи путем окислительного фосфорилирования[править]
Митохондрия[править]
 Рис. 11.1. Схема строения митохондрии
Рис. 11.1. Схема строения митохондрии Митохондрия — это органелла, по размеру сопоставимая с бактериальной клеткой. Примечательно, что у митохондрии есть две мембраны. Наружная мембрана пронизана молекулами порина. Порины образуют каналы, по которым через мембрану могут проходить молекулы с массой менее 10 кДа. Внутренняя мембрана ПРАКТИЧЕСКИ непроницаема; она образует впячивания — кристы. Через внутреннюю мембрану свободно проходят только небольшие молекулы — вроде Н20 и Nh4. Лишь немногие другие молекулы с помощью белков-переносчиков и челночных систем способны преодолеть этот барьер.
Считается, что митохондрия — пример эндосимбиоза. Внутренняя мембрана митохондрии с заключенным в ней содержимым когда-то была древней анаэробной бактерией, которая проникла в примитивную клетку на ранних этапах эволюции. Сохранились и следы прошлого: так, митохондрия имеет свою собственную ДНК (мтДНК), кодирующую 37 генов. 24 из них участвуют в трансляции мтДНК, остальные кодируют белки дыхательной цепи. Примечательно, что только 13 из всех белков митохондриальной дыхательной цепи (а всего их более 85) закодированы в мтДНК. Остальные кодирует ядерная ДНК, и они транспортируются в митохондрию из цитоплазмы.
Дыхательная цепь[править]
 Рис. 11.2. Транспорт электронов в дыхательной цепи. На схеме показан поток электронов от промежуточных метаболитов цикла Кребса (малата и сукцината) к кислороду по цепи переноса электронов, через комплексы I, II, III, IV
Рис. 11.2. Транспорт электронов в дыхательной цепи. На схеме показан поток электронов от промежуточных метаболитов цикла Кребса (малата и сукцината) к кислороду по цепи переноса электронов, через комплексы I, II, III, IV Дыхательная цепь — эффективный путь получения АТФ с использованием НАДН и ФАДН2, которые образуются в процессе окисления метаболического «топлива» [прежде всего углеводов и жирных кислот]. Дыхательная цепь состоит из пяти комплексов — I, II, III, IV и сложного комплекса грибовидной формы (комплекс V). Грибовидный «мультикомплекс» состоит из субъединиц F1 (субъединица «один») и F0 (субъединица «О», связывает олигомицин). Некоторые из комплексов дыхательной цепи содержат цитохромы, которые транспортируют электроны по цепи: комплекс III содержит цитохром b, а комплекс IV — цитохром а/аЗ. Кроме того, в транспорте электронов принимают участие убихинон (кофермент Q10) и цитохром С. Все комплексы дыхательной цепи расположены во внутренней мембране митохондрий. Комплексы I, III и IV не только переносят электроны, но также выполняют функцию молекулярных протонных насосов: они «выкачивают» протоны из матрикса в межмембранное пространство. Внутренняя мембрана непроницаема, в частности, она непроницаема для протонов, поэтому они возвращаются в матрикс только одним путем — через протонный канал комплекса F1/F0, который в этот момент синтезирует АТФ.
Поток электронов упрощенно показан на рис.
Патологии митохондрий[править]
Существуют различные нарушения дыхательной цепи. Многие из них наследственно передаются по материнской линии, поскольку все митохондрии зиготы происходят из митохондрий яйцеклетки. При делении клетки тысячи молекул мтДНК случайным образом распределяются между дочерними клетками, поэтому разные ткани могут содержать как нормальные, так и мутантные молекулы мтДНК (это состояние называется гетероплазмией). Вследствие этого клиническая картина при таких патологиях очень изменчива. Мутации в ядерных генах, кодирующих белки дыхательной цепи, передаются по аутосомному типу и обычно вызывают более тяжелые нарушения.
Атрофия зрительного нерва Лебера[править]
Атрофия зрительного нерва Лебера вызывается мутацией участка митохондриальной ДНК, который кодирует одну из субъединиц комплекса I. От этого нарушения дыхательной цепи митохондрий сильнее всего страдает, по-видимому, зрительный нерв. Болезнь проявляется во взрослом возрасте и приводит к потере зрения.
Митохондриальная энцефаломиопатия, лактацидоз и инсультоподобные эпизоды (синдром MELAS)[править]
Причина синдрома MELAS — мутация гена мтДНК, кодирующего лейциновую транспортную РНК митохондрий. Эта мутация влияет на трансляцию мтДНК, и поэтому при синдроме MELAS нарушена структура всех комплексов дыхательной цепи, кроме комплекса II, который полностью кодируется ядерным геномом.
Болезнь Ли[править]
Болезнь Ли — дегенеративное заболевание центральной нервной системы с характерными патологическими изменениями. Обычно развивается в раннем возрасте. Заболевание генетически гетерогенно: чаще всего причиной болезни являются мутации участков ядерных геномов, кодирующих компоненты дыхательной цепи, однако в некоторых случаях болезнь Ли развивается из-за мутаций митохондриальных генов. При болезни Ли может быть нарушена активность АТФ-синтетазы (комплекса V) или комплексов I, II, III, IV. При некоторых формах болезни Ли имеют место нарушения активности пируватдегидрогеназного комплекса.
Недостаточность пируватдегидрогеназного комплекса приводит к повышению в крови концентраций пирувата, лактата и аланина. У некоторых больных наблюдается улучшение состояния при приеме липоевой кислоты или тиамина (коферментов пируватдегидрогеназного комплекса). С ограниченным успехом применяется лечение кетогенной низкоуглеводной диетой. (Кетоновые тела легко проходят гематоэнцефалический барьер, и при их катаболизме образуется ацетил-КоА независимо от пируватдегирогеназного комплекса.)
Биосинтез АТФ путем окислительного фосфорилирования (часть I)[править]
Биосинтез АТФ в дыхательной цепи, в котором принимает участие как поток электронов (е-), так и поток протонов (Н+), происходит путем окислительного фосфорилирования. Дыхательная цепь состоит из четырех комплексов (I, II, III, IV) и структуры грибовидной формы — АТФ-синтаза с субъединицами F0/F1 или комплекса V, который синтезирует АТФ из АДФ и неорганического фосфата (Фн). Ниже будет рассмотрено, как создается поток электронов и протонов в дыхательной цепи: первый начинается от комплекса I, второй — от комплекса II. 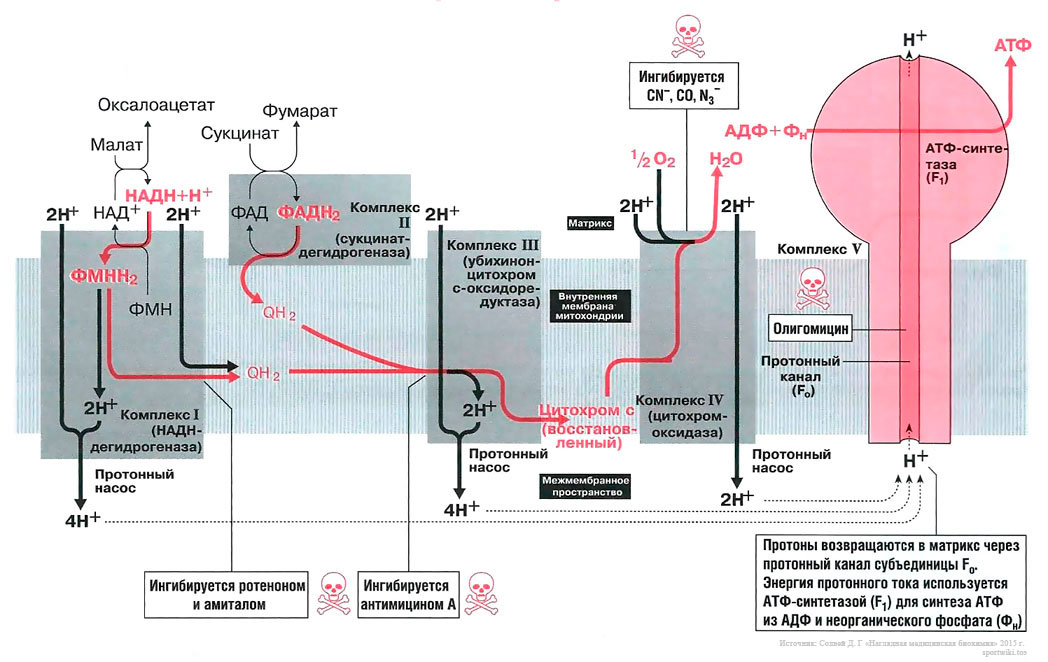
Комплекс I[править]
Поток электронов начинается от НАДН. Энергия для синтеза АТФ возникает благодаря разделению зарядов в комплексе I, которое вызывает движение электронов (электрический ток) и протонов (протонный ток). Молекулярный насос комплекса I выкачивает четыре протона в межмембранное пространство. Другие комплексы (III и VI) тоже выкачивают протоны в межмембранное пространство, и образуется электрохимический градиент протонов; в конечном итоге все эти протоны возвращаются обратно через протонный канал АТФ-синтазы (см. ниже). В комплексе I электроны переносятся с НАДН на убихинон (кофермент Q). Два электрона и два протона восстанавливают убихинон до убихинола (Qh3), и убихинол движется в толще мембраны к комплексу III.
Комплекс II[править]
Поток протонов начинается от ФАДН2. Комплекс II содержит ФАД в виде простетических групп нескольких дегидрогеназ (например, сукцинатдегидрогеназы). Этот ФАД восстанавливается до ФАДН2. Далее комплекс II передает электроны убихинону (Q) для последующего переноса электронов к комплексу III. Примечание, комплекс II не выкачивает протоны.
Комплекс III[править]
В роли донора электронов выступает убихинол Qh3, который при этом окисляется до убихинона Q и в этом виде может возвращаться обратно и принимать следующую пару электронов и протонов. Убихинол передает электроны на цитохром, который транспортирует их в комплекс IV. Протонный насос комплекса III выбрасывает 4 протона в межмембранное пространство.
Комплекс IV[править]
Электроны, поступившие от цитохрома с, передаются кислороду, и кислород полностью восстанавливается с образованием воды. Комплекс IV выбрасывает в межмембранное пространство только 2 протона.
АТФ-синтаза (комплекс V)[править]
Этот комплекс состоит из «ножки гриба» — субъединицы F0, содержащей протонный канал, и из «выпуклой части» — АТФ-синтазы (или F1). Субъединица F0 получила свое название благодаря тому, что она ингибируется олигомицином, а субъединица F1 была первой открытой и выделенной «фракцией» среди всех элементов дыхательной цепи. Поток протонов проходит через протонный канал и запускает молекулярный двигатель, который заставляет АДФ и Фн реагировать друг с другом, образуя молекулы АТФ.
Чтобы синтезировать одну молекулу АТФ и транспортировать ее в цитозоль, требуется 4 протона. При окислении 1 молекулы НАДН+ высвобождается 10 протонов, энергия которых достаточно для образования 2,5 молекул АТФ. При окислении 1 молекулы ФАДН2 высвобождается 6 протонов, энергии которых достаточно для синтеза 1,5 молекул АТФ[1].
Утечка электронов приводит к образованию активных форм кислорода[править]
Примерно 2% электронов высвобождаются из дыхательной цепи и связываются непосредственно с кислородом, образуя активные формы кислорода (АФК). Если работа дыхательной цепи нарушена, АФК образуются в большем количестве. Эти вещества повреждают митохондрии, вызывая все большие нарушения дыхательной цепи. Возникает порочный круг, и в результате из-за накопления различных повреждений под действием АФК происходит старение клетки.
Дыхательные яды[править]
Вещества, которые ингибируют образование АТФ, потенциально токсичны для организма.
Амитал и ротенон блокируют транспорт электронов в комплексе I. Ротенон выделяют из корней растения деррис (Derris scandens) и нередко используют в качестве природного пестицида. Он малотоксичен для человека, поскольку плохо всасывается в желудочно-кишечном тракте. Однако ротенон ядовит для рыб, так как быстро всасывается через жабры. К тому же при долговременном воздействии ротенон опасен и для человека, так как вызывает развитие болезни Паркинсона.
Антимицин блокирует транспорт электронов в комплексе III.
Цианиды(CN-), угарный газ (СО) и азиды (N3-) ингибируют комплекс IV. Поэтому при отравлении цианидом блокируются аэробные метаболические процессы, несмотря на то что кровь достаточно насыщена кислородом. Из-за остановки аэробного метаболизма венозная кровь принимает цвет артериальной крови. Кроме того, наблюдается гипервентиляция, поскольку из-за накопления молочной кислоты стимулируется дыхательный центр.
Олигомицин блокирует протонный канал (F0 в комплексе V) и не дает протонам возвращаться в матрикс. Поэтому АТФ-синтаза (F1) теряет способность синтезировать АТФ.
Биосинтез АТФ путем окислительного фосфорилирования (часть II)[править]
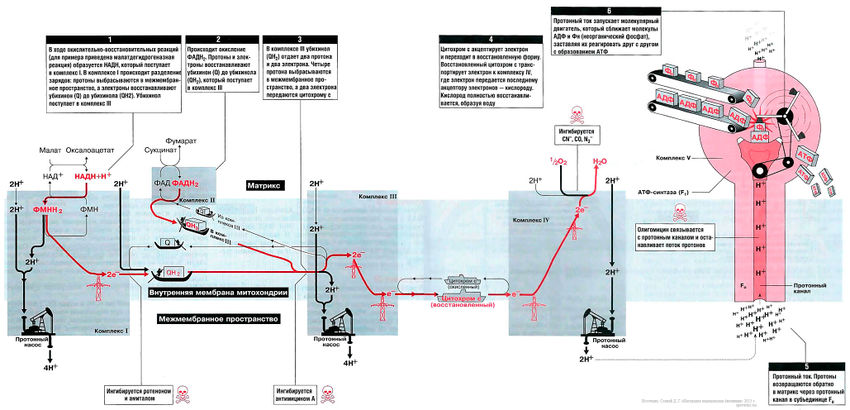
На рис. 13.1 показан поток электронов и протонов в дыхательной цепи. Электроны и протоны от НАДН+ через комплекс I и от ФАДН2 через комплекс II передаются комплексу III. Затем электроны транспортируются в комплекс IV, где они присоединяются к кислороду. В это время протоны выкачиваются протонными насосами из матрикса в межмембранное пространство и возвращаются обратно в матрикс через протонный канал F0-субъединицы АТФ-синтазы (комплекс V). Поток протонов (протонный ток) включает молекулярным двигатель — F1-субъединицу АТФ-синтазного комплекса, и она располагает молекулы АДФ и Фн таким образом, что они объединяются в молекулы АТФ.
- ↑ В отечественных учебниках биохимии принято считать эти значения равными «3» и «2» соответственно, т.е. при окислении 1 молекулы НАДН образуется 3 молекулы АТФ, а при окислении ФАДН2 — 2 молекулы АТФ, что отражает теоретический максимум синтеза АТФ.
sportwiki.to










