
Экологические группы растений по отношению к свету и их адаптивные особенности. Адаптивные признаки растений
Глава 6. АДАПТИВНАЯ МОРФОЛОГИЯ ОРГАНИЗМОВ
Н. М. Чернова, А. М. Былова. «Общая экология»
6.1. Жизненные формы растений
Понятие жизненной формы зародилось при изучении растительного покрова. Сам термин вошел в науку только в конце XIX в., но большое разнообразие форм растений в природе, определяющих ландшафты разных территорий, привлекало ботаников еще в древности.
Древнегреческий ученый и философ, ученик и друг Аристотеля, Теофраст (Феофраст) более чем за три столетия до нашей эры в произведении «Исследования о растениях» систематизировал накопленные знания по морфологии растений, выделил деревья, кустарники, полукустарники, травы и описал их. Деревья он характеризовал как растения со стволом,кустарники – со множеством веток, отходящих прямо от корня,полукустарники – как растения, которые дают от корня много стеблей и множество веточек.Травы он объединял в группы по длительности жизни, характеру побегов, листьев, корневых систем, наличию луковиц и клубней. Он отмечал зависимость формы роста от климата, почвы, способов возделывания. Группы жизненных форм служили Теофрасту, как и многим ботаникам нового времени, основой для систематики растений.
Основоположник ботанической географии немецкий ученый А. Гумбольдт (1769– 1859) в результате знакомства с растительностью разных континентов в начале XIX в. выделил сначала 16, а затем 19 «основных форм», различающихся физиономически: пальмы,
бананы, мальвовые и баобабовые, вересковые, кактусовые, орхидеи, казуариновые, аро-
идные, лианы, алоэ, злаки, папоротники, лилейные, ивовые, миртовые, меластомовые,
хвойные, мимозовые, лотосовые. Это не систематические группы. В основе выделения «основных форм» лежит сходство во внешнем облике. Форма злаков, например, включает осоки и представителей других семейств с узкими листьями. В форму алоэ включены агавы, ананас, панданусовые и др. Сходство внешнего облика разных растений Гумбольдт объяснял влиянием климатических, почвенных условий, высотой над уровнем моря и проч.
Термин «жизненная форма» предложен в 1884 г. датским ботаником Е. Вармингом. Под жизненной формой Варминг понимал «форму, в которой вегетативное тело растения (индивида) находится в гармонии с внешней средой в течение всей его жизни, от колыбели до гроба, от семени до отмирания». Из определения следует, что жизненная форма отражает приспособленность растения ко всему комплексу факторов внешней среды во все периоды его жизни.
Датский ботаник К. Раункиер рассматривал жизненные формы как результат приспособления растений к внешней среде. Решающее значение он придавал климату. В основу выделения жизненных форм Раункиер положил различия в приспособлении растений к переживанию неблагоприятного времени года. Эта приспособленность отражена в размещении почек или верхушек побегов по отношению к поверхности почвы (рис. 69).
Большой вклад в разработку учения о жизненных формах внесли и другие зарубежные и российские ученые. Как синонимы и близкие понятия в экологии растений употребляются термины «растительные формы», «формы роста», «биологический тип», «экобиоморфа», «эпиморфа».
studfiles.net
Экологические группы растений по отношению к свету и их адаптивные особенности
Световой режим любого местообитания определяется интенсивностью прямого и рассеянного света, количеством света (годовой суммарной радиацией), его спектральным составом, а также альбедо – отражательной способностью поверхности, на которую падает свет.
Перечисленные элементы светового режима очень переменчивы и зависят от географического положения, высоты над уровнем моря, от рельефа, состояния атмосферы, характера земной поверхности, растительности, от времени суток, сезона года, солнечной активности и глобальных изменений в атмосфере.
У растений возникают различные морфологические и физиологические адаптации к световому режиму местообитаний.
По требованию к условиям освещения принято делить растения на следующие экологические группы:
1) светолюбивые (световые), или гелиофиты, – растения открытых, постоянно хорошо освещаемых местообитаний;
2) тенелюбивые (теневые), или сциофиты, – растения нижних ярусов тенистых лесов, пещер и глубоководные растения; они плохо переносят сильное освещение прямыми солнечными лучами;
3) теневыносливые, или факультативные гелиофиты, – могут переносить большее или меньшее затенение, но хорошо растут и на свету; они легче других растений перестраиваются под влиянием изменяющихся условий освещения.
Можно отметить некоторые общие приспособительные особенности, свойственные растениям каждой экологической группы.
Световые адаптации гелиофитов и сциофитов.Гелиофиты часто имеют побеги с укороченными междоузлиями, сильно ветвящиеся, нередко розеточные. Листья гелиофитов обычно мелкие или с рассеченной листовой пластинкой, с толстой наружной стенкой клеток эпидермы, нередко с восковым налетом или густым опушением, с большим числом устьиц на единицу площади, часто погруженных, с густой сетью жилок, с хорошо развитыми механическими тканями. У ряда растений листья фотометричные, т. е. повернуты ребром к полуденным лучам или могут менять положение своих частей в зависимости от высоты стояния Солнца. Так, у степного растения софоры листочки непарноперистосложного листа в жаркий день подняты вверх и сложены, у василька русского так же ведут себя сегменты перисторассеченного листа.
Оптический аппарат гелиофитов развит лучше, чем у сциофитов, имеет большую фотоактивную поверхность и приспособлен к более полному поглощению света. Обычно у них лист толще, клетки эпидермы и мезофилла мельче, палисадная паренхима двухслойная или многослойная (у некоторых саван-ных растений Западной Африки – до 10 слоев), нередко развита под верхней и нижней эпидермой. Мелкие хлоропласты с хорошо развитой гранальной структурой в большом числе (до 200 и более) расположены вдоль продольных стенок.
Хлорофилла на сухую массу в листьях гелиофитов приходится меньше, но зато в них содержится больше пигментов I пигментной системы и хлорофилла П700. Отношение хлорофилла а к хлорофиллу b равно примерно 5: 1. Отсюда высокая фотосинтетическая способность гелиофитов. Компенсационная точка лежит в области более высокой освещенности. Интенсивность фотосинтеза достигает максимума при полном солнечном освещении. У особой группы растений – гелиофитов, у которых фиксация СО2 идет путем С4-дикарбоновых кислот, световое насыщение фотосинтеза не достигается даже при самой сильной освещенности. Это растения из засушливых областей (пустынь, саванн). Особенно много С4-растений среди семейств мятликовых, осоковых, аизовых, портулаковых, амарантовых, маревых, гвоздичных, молочайных. Они способны к вторичной фиксации и реутилизации СО2, освобождающегося при световом дыхании, и могут фотосинтезировать при высоких температурах и при закрытых устьицах, что часто наблюдается в жаркие часы дня.
Обычно С4-растения, особенно сахарный тростник и кукуруза, отличаются высокой продуктивностью.
Сциофиты– это растения, постоянно находящиеся в условиях сильного затенения. При освещенности 0,1–0,2 % могут расти только мхи и селягинеллы. Плауны довольствуются 0,25-0,5 % полного дневного света, а цветковые растения встречаются обычно там, где освещенность в пасмурные дни достигает не менее 0,5–1% (бегонии, недотрога, травы из семейств имбирные, мареновые, коммелиновые).
В северных широколиственных и темнохвойных лесах полог сомкнутого древостоя может пропускать всего 1–2% ФАР, изменяя ее спектральный состав. Сильнее всего поглощаются синие и красные лучи, и пропускается относительно больше желто-зеленых лучей, дальних красных и инфракрасных. Слабая освещенность сочетается с повышенной влажностью воздуха и повышенным содержанием в нем СО2, особенно у поверхности почвы. Сциофиты этих лесов – зеленые мхи, плауны, кислица обыкновенная, грушанки, майник двулистный и др.
Листья у сциофитов располагаются горизонтально, нередко хорошо выражена листовая мозаика. Листья темно-зеленые, более крупные и тонкие. Клетки эпидермы крупнее, но с более тонкими наружными стенками и тонкой кутикулой, часто содержат хлоропласты. Клетки мезофилла крупнее, палисадная паренхима однослойная или имеет нетипичное строение и состоит не из цилиндрических, а из трапециевидных клеток. Площадь жилок вдвое меньше, чем у листьев гелиофитов, число устьиц на единицу площади меньше. Хлоропласты крупные, но число их в клетках невелико.
У сциофитов по сравнению с гелиофитами меньше хлорофилла П700. Отношение хлорофилла а к хлорофиллу b равно примерно 3: 2. С меньшей интенсивностью протекают у них такие физиологические процессы, как транспирация, дыхание. Интенсивность фотосинтеза, быстро достигнув максимума, перестает возрастать при усилении освещенности, а на очень ярком свету может даже понизиться.
У лиственных теневыносливых древесных пород и кустарников (дуба черешчатого, липы сердцевидной, сирени обыкновенной и др.) листья, расположенные по периферии кроны, имеют структуру, сходную со структурой листьев гелиофитов, и называются световыми, а в глубине кроны – теневые листья с теневой структурой, сходной со структурой листьев сциофитов (рис. 24).
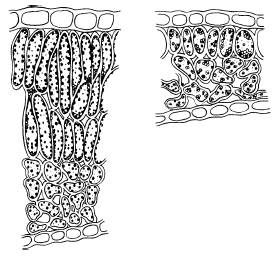
Рис. 24. Поперечный срез светового (слева) и теневого (справа) листа сирени (по И. С. Михайловской, 1977)
Факультативные гелиофиты, или теневыносливые растения, в зависимости от степени теневыносливости имеют приспособительные особенности, сближающие их то с гелиофитами, то со сциофитами. К этой группе можно отнести некоторые луговые растения, лесные травы и кустарники, растущие и в затененных участках леса, и на лесных полянах, опушках, вырубках. На осветленных местах они разрастаются часто сильнее, однако оптимальное использование ФАР у них происходит не при полном солнечном освещении.
У деревьев и кустарников теневая или световая структура листа часто определяется условиями освещения предыдущего года, когда закладываются почки: если закладка почек идет на свету, то формируется световая структура, и наоборот.
Если в одном и том же местообитании закономерно периодически изменяется световой режим, растения в разные сезоны могут проявлять себя то как светолюбивые, то как теневыносливые.
Весной в дубравах под полог леса проникает 50–60 % солнечной радиации. Листья розеточных побегов сныти обыкновенной имеют световую структуру и отличаются высокой интенсивностью фотосинтеза. В это время они создают основную часть органического вещества годичной продукции. Листья сныти летней генерации, появляющиеся при развитом древесном пологе, под который проникает в среднем 3,5 % солнечной радиации, имеют типичную теневую структуру, и интенсивность фотосинтеза их значительно ниже, в 10–20 раз. Подобную двойственность по отношению к свету проявляет и осока волосистая, светолюбивая весной и теневыносливая летом. По-видимому, это свойственно и другим растениям дубравного широкотравья.
Отношение к световому режиму меняется у растений и в онтогенезе. Проростки и ювенильные растения многих луговых видов и древесных пород более теневыносливы, чем взрослые особи.
Иногда у растений меняются требования к световому режиму, когда они оказываются в иных климатических и эдафических условиях. Так, обычные теневыносливые растения хвойного леса – черника, седмичник европейский и некоторые другие – в тундре приобретают особенности гелиофитов.
Наиболее общая адаптация растений к максимальному использованию ФАР – пространственная ориентация листьев. При вертикальном расположении листьев, как, например, у многих злаков и осок, солнечный свет полнее поглощается в утренние и вечерние часы – при более низком стоянии солнца. При горизонтальной ориентации листьев полнее используются лучи полуденного солнца. При диффузном расположении листьев в разных плоскостях солнечная радиация в течение дня утилизируется наиболее полно. Обычно при этом листья нижнего яруса на побеге отклонены горизонтально, среднего направлены косо вверх, а верхнего располагаются почти вертикально.
Считают, что кукуруза является одной из самых высокопродуктивных сельскохозяйственных культур потому, что наряду с высоким КПД фотосинтеза у нее наблюдается диффузное расположение листьев, при котором полнее поглощается ФАР.
На севере, где высота стояния солнца меньше, встречается больше растений с вертикальным расположением листьев, а на юге – с горизонтальным. Для получения большей биомассы выгодны также посевы и насаждения, в которых сочетаются растения с разной пространственной ориентацией листьев, причем в верхнем ярусе лучше иметь растения с вертикальным расположением листьев, которые полнее используют свет при низком стоянии солнца, не препятствуют прохождению полуденных лучей к расположенным в нижнем ярусе листьям с горизонтальной ориентацией.
lektsia.com
адаптивный признак
Указанные примеры адаптивных признаков строения, физиологических процессов и поведения насекомых, связанных с паразитизмом и хищничеством, характеризуют разнообразие взаимных приспособлений паразитов, хищников и насекомых, на которых они нападают.[ ...]
| Варьирование адаптивного признака при изменении величины воздействующего фактора (по И А. Шилову, 1985) |  |
Обитание в почве повело к образованию у насекомых адаптивных признаков строения и поведения.[ ...]
У людей увеличиваются затраты энергии, а потому питание является важным адаптивным признаком. Для оценки калорийности пищи в районах Крайнего Севера используется шкала, предложенная Комитетом по потребностям в калориях организма и вопросам пищевых продуктов и сельского хозяйства при ООН. Комитет рекомендует с понижением среднемесячной температуры на каждые 10° С увеличивать калорийность питания на 5%, считая за исходную температуру +10 С.[ ...]
Естественно, что евтропные насекомые имеют для растений наибольшее значение в качестве опылителей. Адаптивные признаки у растений могут все же возникать и по отношению к аллотропным насекомым — такие цветки имеют нектар и пыльцу в верхних их частях, например Euphorbia. А у некоторых растений (Arum) создались цветки-ловушки, задерживающие аллотропных насекомых (мух) до тех пор, пока опыление не будет обеспечено.[ ...]
Сходный тип передвижения и внешний облик имели и некоторые вымершие динозавры мезозоя. Вполне закономерно, что кроме общих адаптивных признаков, каждый вид отличается деталями внешнего строения в зависимости от экологической специфики.[ ...]
По вопросу об адаптивных признаках строения насекомых имеется к настоящему времени довольно обширная литература.[ ...]
Биосфера пережила, по крайней мере, пять катастроф, когда, казалось, живое вещество вот-вот вымрет. Однако всякий раз жизнь «воскрешалась» за счет тех видов, которые имели новые адаптивные признаки. А потому и говорят, что жизнь не располагает иным способом сохраняться в режиме динамического равновесия, кроме приспособления к среде, в чем и проявляется зависимость жизни от среды.[ ...]
Все животные имеют тенденцию к расселению на более широкую территорию, но не при всяких новых условиях среды они могут выживать. Учитывая это, некоторые авторы вопрос о жизненных формах по существу сводили или к избирательной способности животных к комплексу внешних условий (Эйгенманн, 1908, например, подчеркивал, что различные признаки организмов становятся адаптивными только благодаря выбору ими соответствующего места обитания), или к пассивному расселению и дальнейшему естественному отбору, выживанию наиболее приспособленных форм ‘(например, Девенпорт, 1903). Такие взгляды не только односторонни, но и принципиально ошибочны, так как в основе их лежит отрицание роли среды в формировании жизненных форм. Возникновение тех или иных адаптивных признаков под влиянием взаимосвязей со средой теперь может считаться доказанным и это весьма существенно для понимания жизненных форм. Наиболее прочные элементы в цепях питания и экологических нишах — жизненные формы, сформировавшиеся в данном биотопе.[ ...]
Отмеченные особенности непосредственно отражаются на гидробионтах. Растения, произрастающие в условиях переувлажнения или непосредственно в воде, обладают рядом морфоанатомических, а также физиологических и биохимических особенностей, обеспечивающих их нормальное существование в данных условиях. Степенью связи с водной, воздушной средами, а также способом снабжения растений минеральными веществами обусловлено разделение макрофитов на группы, характеризующиеся определенными адаптивными признаками. Иными словами, комплекс водных макрофитов образует целый ряд жизненных форм.[ ...]
Заманчиво говорить о К, г и ¿ видах, но такой прием мог бы ввести в заблуждение. У всех видов отбор должен действовать так, чтобы продуцировать достаточно высокое г, а большинство видов будет испытывать влияние ¿-отбора, по крайней мере в течение некоторого времени и в некоторых условиях среды. Более того, отбор не просто направлен на получение максимальных значений г (или К, или ¿).Для всех видов, включая и те, которые мы назвали г-отобран-ными, величина г будет результатом выбора между потенциальным преимуществом высоких значений г (продуцирование большего количества потомства) и издержками, или «штрафами», для особи (или ее потомства) за усилия увеличить размножение. Значения г очень широко меняются в каждой из трех групп, и некоторые возникшие в результате К- и ¿-отбора виды имеют высокие значения г. Эти три типа нельзя просто охарактеризовать значениями г или долей энергии, затрачиваемой отдельными организмами на размножение. Лучше обратить внимание на различные обстоятельства отбора, влияющие на эти популяции, и использовать термины К-, г- и ¿-отборы только как «этикетки» для обозначения этих обстоятельств. Мы различаем следующие обстоятельства: 1) отбор насыщения, или взаимодействие отборов в условиях среды, которые преимущественно благоприятны, относительно стабильны и насыщенны, то есть полностью заняты другими организмами; 2) отбор эксплуатации в среде, в которой чередуются благоприятные и неблагоприятные условия, или в среде, которая только временно благоприятна в измененных местоположениях, таких, как гари, площади, занятые иван-чаем, и которая соответственно в меньшей степени занята другими видами, и 3) отбор в неблагоприятной обстановке, в условиях преимущественно неблагоприятной и ограничивающей среды, которая только временами позволяет популяции расти. Виды не образуют трех четко очерченных групп в согласии с описанными ситуациями, но различаются по относительному влиянию этих ситуаций на их адаптивные признаки. Виды могут формировать континуум адаптаций, которые вызваны соответственно относительным преобладанием отбора насыщения, эксплуатации и отбора в неблагоприятной обстановке, и характеризоваться соотношением генов, отобранных для реализации взаимодействия видов в занятой и стабильной среде, для распространения и роста в нестабильной среде и для выживания в периоды неблагоприятной среды.[ ...]
ru-ecology.info
АДАПТИВНЫЕ ПРИЗНАКИ СТРУКТУРЫ ЛИСТА РАСТЕНИЙ РАЗНЫХ ЭКОЛОГИЧЕСКИХ ГРУПП
Транскрипт
1 ЭКОЛОГИЯ, 214, 2, с УДК (924.82) АДАПТИВНЫЕ ПРИЗНАКИ СТРУКТУРЫ ЛИСТА РАСТЕНИЙ РАЗНЫХ ЭКОЛОГИЧЕСКИХ ГРУПП 214 г. Л. А. Иванова Ботанический сад УрО РАН Екатеринбург, ул. 8 Марта, 22а e-mil: Поступила в редакцию г. Адаптивное значение структуры листа растений хорошо известно, однако точные структурные критерии для индикации экологических свойств вида до сих пор отсутствуют. Нами изучены параметры листьев у 1 видов травянистых растений южной тайги разных экологических групп по факторам освещения и увлажнения. Площадь, толщина и тип анатомии листа были в меньшей степени связаны с экологическими свойствами видов, чем структурные показатели фотосинтетических тканей. В ряду тенелюбивые мезофиты (М) > светолюбивые гигро-м > М > ксеро-м > мезоксерофиты постепенно до 7 1 крат возрастала концентрация клеток и хлоропластов в листе, в 2 3 раза уменьшались их размеры и в 4 5 раз увеличивалось отношение поверхности мезофилла к поверхности листа. Сделан вывод об адаптивном характере показателей мезофилла и предложена система признаков для идентификации экологических групп растений бореальной зоны. Ключевые слова: бореальная зона, экологические группы растений, затенение, засухоустойчивость, структура листа, мезофилл, число клеток, размеры клетки. DOI: /S Адаптация растений к экологическим условиям тесно связана с перестройкой фотосинтетического аппарата. Процесс фотосинтеза требует постоянного поглощения света, углекислого газа и поддержания транспирационного потока с поверхности листа, для чего необходимо полное соответствие структуры листа световому и гидротермическому режиму местообитания. В связи с этим именно структурные параметры листа многие авторы считают наиболее информативными при сравнительном исследовании растений разных эколого-географических групп (Василевская, 1965; Мокроносов, 1978; Гамалей, 1988, 24). Однако не все листовые признаки одинаково значимы для индикации экологических свойств вида. Известная консервативность структуры покровных тканей и проводящей системы, успешно используемых для глобальных систематических и филогенетических исследований (Василевская, 1954; Эсау, 1969; Гамалей, 24), делает невозможным их применение при изучении адаптации к локальным экологическим условиям. Наиболее пластична в экологическом плане структура мезофилла листа. Так, тип строения мезофилла отражает условия произрастания вида: гомогенный тип чаще встречается у тенелюбивых растений, в то время как дорзовентральный характерен для светолюбивых мезофитов, а изопалисадный для ксерофитов (Эсау, 1969; Гамалей, 1988; Василевская, Бутник, 1982). Начиная с работ В.Р. Заленского (194), F. Turrel (1936), Т.Н. Годнева и С.В. Калишевич (1938) известно, что листья растений разных местообитаний существенно различаются по количественным показателям мезофилла, связанным с фотосинтетической функцией. В х годах были определены общие направления структурной перестройки фотосинтетических тканей у растений, обитающих в разных экологических условиях (Мокроносов, 1978; Цельникер, 1978; Гамалей, 1988; Горышина, 1989; Noel, 1977). Показано, что адаптация листа к затенению связана со снижением концентрации хлоропластов и увеличением их размеров (Горышина, 1989; Пьянков, 1993; Tershim et l., 21), а приспособление к высокой инсоляции и водному дефициту сопровождается увеличением численной плотности клеток и хлоропластов (Мокроносов, 1978; Горышина, 1989; Зверева, 1986, 1988; Tershim et l., 21; Ivnov et l., 24). Результаты этих исследований свидетельствуют о том, что регулирование количественных показателей мезофилла является более тонкой настройкой листа на условия среды, чем тип морфологии или анатомии листа. Однако имеющаяся в настоящее время информация о количественных показателях фотосинтетических тканей далеко не полная и пока понятны лишь самые общие тенденции их изменения в зависимости от основных экологических факторов (Пьянков, Мокроносов, 1993). Доступные в литературе данные разрозненны и немногочисленны. На начальном этапе осмысления роли структуры фотосинтетического аппарата в адаптации растений сравнивались только очень кон- 19
2 11 ИВАНОВА трастные группы например, мезофиты умеренных широт и склероксерофиты пустынь (Мокроносов, Шмакова, 1978). Точные количественные данные об экологических группах в пределах одной климатической зоны отсутствуют, что до сих пор делает невозможным использование количественных параметров мезофилла для объективной идентификации экологических свойств видов. Цель данной работы изучить параметры листьев и количественные показатели мезофилла у растений разных экологических групп в пределах бореальной зоны, определить наиболее значимые показатели и выявить точные количественные критерии для идентификации экологических свойств видов. МАТЕРИАЛ И МЕТОДЫ Исследования проводили в окрестностях г. Екатеринбурга, а также на биологической станции УрГУ, находящейся в 5 км к юго-востоку от г. Екатеринбурга. Район исследований находится в пределах южной тайги, вблизи границы с предлесостепной подзоной, что обусловило достаточно большое разнообразие условий произрастания. Растения собирали в экотопах, наиболее характерных для каждого вида, среди них: разные типы соснового леса (разнотравно-черничный, брусничноосоковый, бруснично-орляковый), опушки лесов, злаково-разнотравный луг, пойменный луг, суходольный луг, остепненные склоны. Климат умеренно-континентальный, среднемноголетняя температура воздуха.9 C, средняя температура июля 17.6 C. Период интенсивного вегетативного роста растений длится с мая до конца августа. Растительный материал собирали преимущественно в июне июле, в период цветения большинства изученных видов. Было исследовано 1 видов растений естественной флоры Среднего Урала. Сорные и интродуцированные виды не изучали. Учитывая существенные различия в структуре листа у растений разных жизненных форм (Гамалей, 24; Иванова, 212), для исследований отбирали растения только одной жизненной формы травянистые многолетники. Абсолютное большинство изученных видов относились к классу Двудольных. Экологические группы растений были выделены согласно литературным источникам (Куликов, 25; Флора Сибири, ) по уровню освещенности сциофиты и гелиофиты и степени увлажнения среды гигромезофиты, мезофиты, ксеромезофиты, мезоксерофиты. Отбирали листья среднего яруса с 1 15 растений одного вида, находившихся в фазе бутонизации цветения. Измерения проводили с помощью цифрового анализатора Simgis Mesoplnt (ООО СИАМС, Россия, Екатеринбург) и светового микроскопа Zeiss Axiostr (Zeiss, Германия). Площадь листа и удельную поверхностную плотность листа (УППЛ, или сухой вес единицы площади листа) измеряли с помощью фотографирования свежих листьев, компьютерного анализа изображений и взвешивания сухих листьев. Объемную плотность листа (сухой вес единицы объема листа, D) рассчитывали через отношение УППЛ и толщины листа. Тип строения мезофилла определяли, основываясь на общепринятой классификации (Эсау, 1969; Василевская, Бутник, 1982). Толщину листа и размеры хлоропластов измеряли на поперечных срезах листьев. Показатели мезофилла получены согласно методике мезоструктуры листа (Мокроносов, 1978). Число клеток и хлоропластов определяли на фиксированном в 7%-ном этаноле материале. Объем и площадь поверхности клеток мезофилла определяли с помощью проекционного метода (Иванова, Пьянков, 22). Используя число клеток или хлоропластов в единице площади листа и среднюю площадь поверхности клетки или хлоропласта, рассчитывали общую площадь поверхности наружных мембран клеток (индекс мембран клеток, ИМК) и хлоропластов (индекс мембран хлоропластов, ИМХ) в единице площади листа (см 2 см 2 ). Статистический анализ проводили с использованием критерия Манна-Уитни для попарного сравнения групп, а также однофакторного дисперсионного анализа (ANOVA) и F-критерия для выявления наиболее значимых параметров. РЕЗУЛЬТАТЫ Параметры листовой пластинки. Размеры листа были наиболее изменчивым листовым показателем, по которому экологические группы значительно перекрывались и не обнаруживали достоверных различий (см. таблицу). По толщине листа различались лишь крайние группы в экологическом ряду. Наиболее тонкими листьями отличались тенелюбивые мезофиты и светолюбивые гигромезофиты более половины видов имели толщину листа мкм на поперечном срезе, а максимальной толщиной листа (2 25 мкм) характеризовались ксеромезо- и мезоксерофиты. УППЛ в большей степени, чем толщина и площадь листовой пластинки, изменялась в ряду экологических групп и увеличивалась в 2 раза от сциофитов к светолюбивым мезоксерофитам. Объемная плотность листа была на 25% ниже у тенелюбивых, чем у светолюбивых мезофитов, а у ксеромезофитов она возрастала в 1.5 раза по сравнению с мезофитами. Типы строения мезофилла. Экологические группы различались по встречаемости типов строения мезофилла (рис. 1), однако полного соответствия типа анатомии и экологических свойств вида не обнаружено. Среди одной экологической группы встречались растения с разными ти- ЭКОЛОГИЯ 2 214
3 АДАПТИВНЫЕ ПРИЗНАКИ СТРУКТУРЫ ЛИСТА РАСТЕНИЙ 111 Параметры листьев растений разных экологических групп Экологическая группа Число видов Площадь листа, см 2 Толщина листа, мкм УППЛ, мг/дм 2 D, г/см 3 Сциофиты Мезофиты и гигромезофиты ± ± ± ±.6 Гелиофиты Гигромезофиты 8 23 ± ± ± ±.12 Мезофиты ± ± ± ±.8 Ксеромезофиты ± ± ± ±.14 Мезоксерофиты 5 6 ± ± ± ±.11 F p Примечание. Представлены средние арифметические значения и стандартные отклонения. Латинскими буквами обозначена достоверность различий при попарном сравнении экологических групп по критерию Манна-Уитни на уровне p <.5 одинаковые буквы обозначают отсутствие достоверных различий; УППЛ удельная поверхностная плотность листа; D объемная плотность листа; F значение F-критерия для данного показателя при однофакторном дисперсионном анализе; p уровень достоверности F-критерия. пами мезофилла, а одинаковый тип мезофилла был у представителей нескольких экологических групп. Из тенелюбивых растений лишь половина видов имели гомогенный тип мезофилла (см. рис. 1), остальные виды дорзовентральный тип. Основная масса гелиофитов около 9% обладала дорзовентральным мезофиллом. Изолатеральный мезофилл обнаружен у видов с признаками ксерофитизации ксеромезофитов и мезоксерофитов. Параметры клеток и хлоропластов. Обнаружены постепенные направленные изменения параметров в экологическом ряду сциофиты гигромезофиты мезофиты ксеромезофиты мезоксерофиты. Наиболее значимыми были изменения количества клеток и хлоропластов, о чем свидетельствуют максимальные значения F-критерия (рис. 2). Так, число клеток хлоренхимы различалось между крайними группами в 7 1 раз, в то время как варьирование размеров фотосинтетических клеток среди экологических групп было 2 3-кратным. Концентрация клеток была минимальной у сциофитов 1 2 тыс. в 1 см 2 листа и постепенно увеличивалась в экологическом ряду до 8 12 тыс/см 2 у мезоксерофитов. При этом последние отличались небольшими размерами клеток 5 6 тыс. мкм 3, а наиболее крупными клетками обладали тенелюбивые мезофиты и светолюбивые гигромезофиты 1 2 тыс. мкм 3. Число хлоропластов в клетке практически не зависело от экологической группы растения (см. рис. 2). Большинство изученных видов Среднего Урала содержали 2 4 хлоропластов в клетке мезофилла, и лишь у сциофитов в среднем было больше пластид 4 5 на клетку мезофилла. Объем клетки, приходящийся на один хлоропласт (КОХ, клеточный объем хлоропласта), снижался от 5 мкм 3 в среднем у сциофитов до 2 мкм 3 у ксеромезофитов. При этом средний объем хлоропласта также уменьшался от 6 мкм 3 у сциофитов до 4 мкм 3 у мезофитов и 3 мкм 3 у мезоксерофитов. Интегральные количественные параметры фотосинтетических тканей. В направлении от тенелюбивых мезофитов к светолюбивым мезоксерофитам происходило постепенное увеличение количества хлоропластов в единице площади листа от 6 млн хлоропластов в 1 см 2 листа у сциофитов, 13 млн у светолюбивых мезофитов и до млн у ксеромезо- и мезоксерофитов. В этом же направлении равномерно возрастали такие интегральные показатели структуры мезофилла, как Встречаемость, % С 7 1 ГМ М КМ МК Экологическая группа 2 3 ИЛ ДВ Гом Рис. 1. Встречаемость разных типов мезофилла среди изученных видов растений разных экологических групп. Типы мезофилла: Гом гомогенный, ДВ дорзовентральный, ИЛ изолатеральный; экологические группы: С сциофиты, ГМ гигромезофиты, М мезофиты, КМ ксеромезофиты, МК мезоксерофиты; цифры количество изученных видов. ЭКОЛОГИЯ 2 214
4 112 ИВАНОВА Объем клетки, 1 3 мкм F = 5.8* Число хлоропластов в клетке 1 F = Объем хлоропласта, мкм F = 11.3* Число клеток в ед. площади листа, 1 3 см 2 2 F = 28.6* d Число хлоропластов в ед. площади листа, 1 6 см 2 4 F = 19.8* КОХ, мкм F = 6.5* ИМК, см 2 см 2 3 F = 15.6* 2 с 1 ИМХ, см 2 см 2 2 F = 7.2* C ГМ М КМ МК C ГМ М КМ МК Экологическая группа Рис. 2. Параметры фотосинтетических тканей у растений разных экологических групп. Обозначения экологических групп см. на рис. 1. Точка на диаграмме обозначает медиану; заштрихованный прямоугольник 25 75% выборки; концевые отметки минимальное и максимальное значения; F значение F-критерия для данного показателя при однофакторном дисперсионном анализе, * достоверность F-критерия p <.1. Латинскими буквами обозначена достоверность различий при попарном сравнении экологических групп по критерию Манна-Уитни на уровне p <.5 одинаковые буквы обозначают отсутствие достоверных различий. ЭКОЛОГИЯ 2 214
5 АДАПТИВНЫЕ ПРИЗНАКИ СТРУКТУРЫ ЛИСТА РАСТЕНИЙ 113 общая поверхность клеток и хлоропластов в расчете на единицу площади листа (ИМК и ИМХ) (см. рис. 2). Так, сциофиты характеризовались минимальными значениями внутренней и внешней поверхности листа в среднем 6 см 2 поверхности клеток и 5 см 2 хлоропластов на 1 см 2 площади листа, тогда как у ксеромезофитов общая площадь поверхности клеток в 2 раз, а поверхность хлоропластов в 1 раз превышали площадь листа. Изменения значений ИМК в ряду экологических групп были более значительными, чем ИМХ. Соотношение параметров у видов одной экологической группы, но с разными типами строения мезофилла. Сравнительный анализ растений с разными типами мезофилла внутри одной экологической группы показал отсутствие различий по большинству параметров (рис. 3). У сциофитов с гомогенным и дорзовентральным типами были сходные показатели листьев и тканей небольшие толщина и плотность листа, крупные размеры клеток и хлоропластов и низкое их количество в единице площади листа. Ксеромезофиты с изолатеральным типом не отличались от ксеромезофитов с дорзовентральными листьями абсолютно по всем показателям, за исключением толщины листа. Число и размеры клеток и хлоропластов были сходными у всех ксеромезофитов независимо от типа строения листа. При этом растения с одинаковым дорзовентральным типом мезофилла, но принадлежавшие к разным экологическим группам тенелюбивые мезофиты и светолюбивые ксеромезофиты, существенно отличались друг от друга по всем показателям структуры мезофилла (см. рис. 3). ОБСУЖДЕНИЕ Проведенные исследования показали, что в условиях бореальной зоны внешние признаки листьев размеры и толщина листовой пластинки мало соответствовали экологическим свойствам видов. В то же время эти параметры часто указывают в качестве критериев экологической приуроченности вида. Так, принято считать, что тенелюбивые растения характеризуются тонкими листьями, а ксерофиты имеют толстые, плотные, кожистые листья (Варминг, 191; Shimper, 193; Максимов, 1926; Василевская, 1954, 1965; Гамалей, 1988; Горышина, 1989). Однако не все тенелюбивые растения обладают тонкими листьями. Например, типичные теневые листья лесных видов растений Северной Каролины (Северная Америка Slton, Smith, 22) имели листья толщиной до 265 мкм, что превышает средние значения, обнаруженные нами для мезоксерофитов Среднего Урала (см. таблицу). По данным Arus et l. (1986), толщина листа облигатно теневого вида Phylodendron sndens составляла 296 мкм, в то время как у относительно светоустойчивого растения Cissus rhomifoli лист был в 2.5 раза тоньше. Также указываемая для ксерофитов большая плотность листа свойственна, кроме того, деревьям, медленно растущим растениям и растениям с вечнозелеными листьями (Pynkov et l. 1998; Poorter et l. 29; Иванова, 212). В наших исследованиях средние значения толщины и плотности листа отчетливо различались лишь у наиболее контрастных групп тенелюбивых мезофитов и светолюбивых мезоксерофитов (см. таблицу). Кроме того, в качестве отличительного признака ксероморфизма часто указывают мелкие размеры листьев (Гамалей, 1988; Зверева, 1986; Василевская, 1954; Буинова, Бадмаева, 1989). Уменьшение площади листа приводит к увеличению теплового рассеяния и снижению транспирационных потерь важным факторам для существования растений в ксеридных условиях. Однако мы не обнаружили достоверных различий по площади листа между растениями разных экологических групп. Видимо, в условиях бореальной зоны сокращение листовой площади не выгодно из-за опасности снижения конкурентоспособности, и ведущую роль в адаптации фотосинтеза к условиям среды играет анатомическое строение листа. При этом тип анатомии листа в наших исследованиях также не являлся однозначной характеристикой экологических свойств вида. Экологические группы различались по степени встречаемости типов мезофилла с преобладанием гомогенного среди сциофитов и присутствием изолатерального среди наиболее ксерофитных видов (см. рис. 1). Однако абсолютное большинство изученных видов имели дорзовентральный тип и при этом принадлежали к разным экологическим группам от сциофитов до светолюбивых мезоксерофитов. Итак, основные различия между экологическими группами состояли не в размерах листьев и типе анатомии листа, а в количественных параметрах мезофилла (см. рис. 2). Нами показано, что в ряду экологических групп растений бореальной зоны: тенелюбивые мезофиты светолюбивые гигромезофиты мезофиты ксеромезофиты мезоксерофиты, происходит постепенное увеличение концентрации клеток и хлоропластов в листе, что обусловливает возрастание внутренней поверхности листа, выраженной индексами ИМК и ИМХ (см. рис. 2). Похожее соотношение количественных параметров мезофилла минимальная концентрация клеток и хлоропластов у тенелюбов и большее их число у светолюбивых видов и ксерофитов было показано ранее несколькими авторами (Мокроносов, Шмакова, 1978; Дьяченко, 1978; Горышина, 1989; Зверева, 1986; Пьянков, 1993; Ivnov et l., 24). Указанные работы наглядно продемонстрировали существенные различия между тенелюбивыми и светолюбивыми, мезофитными 3 ЭКОЛОГИЯ 2 214
6 114 ИВАНОВА Толщина листа, мкм 3 2 УППЛ, мг дм Объем клетки, 1 3 мкм Число клеток, 1 3 см Объем хлоропласта, мкм 3 Число хлоропластов, 1 6 см С-Гом С-ДВ КМ-ДВ КМ-ИЛ С-Гом С-ДВ КМ-ДВ КМ-ИЛ Рис. 3. Параметры структуры листа сциофитов и ксеромезофитов с разными типами строения мезофилла (среднее арифметическое значение и ошибка среднего). Латинскими буквами обозначена достоверность различий (см. рис. 2). Названия экологических групп и типов мезофилла см. на рис. 1. и ксерофитными видами. В то же время, несмотря на достаточно длительную историю развития данного подхода, точные данные по количеству клеток и хлоропластов в листьях растений с разными экологическими свойствами представлены лишь в единичных работах. Выявленные закономерности получены для разнородных контрастных групп или на небольшом числе видов, часто без учета жизненной формы и систематического положения вида. Имеющиеся литературные данные разрозненны, часто противоречивы и в основном дают представления о различиях между экологическими группами на уровне больше меньше. Так, в работе А.Т. Мокроносова и Т.В. Шмаковой (1978) для 5 видов мезофитов указано число клеток 2 4 тыс/см 2, в то время как у Т.К. Горышиной (1989) для 5 видов луговых мезофитов от 3 до 12 тыс. клеток в 1 см 2 листа. Среди 39 изученных нами светолюбивых мезофитов абсолютное большинство имели объем клетки мезофилла не более 1 12 тыс. мкм 3, что не совпадает с модальными значениями мезофитов умеренных широт от 1 до 25 тыс. мкм 3 (Мокроносов, Шмакова, 1978) и в 2 раза ниже значений, указанных для мезофитов в работе А.П. Дьяченко (1978). Таким образом, информация о параметрах мезофилла листа растений разных экологических групп является далеко не полной, а такие промежуточные экологические группы, как ксеромезофиты и мезоксерофиты, вообще не изучены. Вместе с тем важность точной количественной оценки структурных параметров мезофилла при ЭКОЛОГИЯ 2 214
7 АДАПТИВНЫЕ ПРИЗНАКИ СТРУКТУРЫ ЛИСТА РАСТЕНИЙ 115 изучении адаптации растений к условиям среды не вызывает сомнений. Структурные признаки мезофилла имеют адаптивный характер, поскольку оказывают прямое влияние на приспособление фотосинтеза к экологическим условиям. Способность достигать необходимого уровня фотосинтеза позволяет растению поддерживать положительный углеродный баланс, т.е. напрямую определяет выживаемость особей в конкретных экологических условиях. Соотношение размеров и количества фотосинтетических клеток и хлоропластов определяет интегральные параметры мезофилла суммарную поверхность клеток и хлоропластов, которые прямо влияют на эффективность поглощения света и скорость газообмена между внешней средой и листом (Noel, 1977; Pynkov et l., 1998; Tershim et l., 21). Так, у тенелюбивых растений низкая концентрация фотосинтетических элементов соответствует слабому развитию внутрилистовой ассимиляционной поверхности, что, с одной стороны, способствует увеличению коэффициента пропускания света внутри листа, а с другой обусловливает низкую скорость газообмена листа. По нашим расчетам, используя формулу А. Лайска с соавт. (197), у сциофитов при ИМК 4 8 скорость газообмена через поверхность клеток является минимальной и составляет всего.3.6 см/сек, в то время как у светолюбивых мезофитов при ИМК 9 14 скорость поступления СО 2 внутрь клеток составляет.7 1. см/сек. Низкие значения уровня газообмена обычны для тенелюбивых растений, поскольку при явном дефиците света скорость газообмена через поверхность клеток не является лимитирующим фактором фотосинтеза (Мокроносов, 1978; Arus et l., 1986; Пьянков, 1993; Tershim et l., 21). Для гелиофитов, фотосинтезирующих при насыщающем свете, напротив, величина внутрилистовой ассимиляционной поверхности играет определяющую роль в ограничении уровня фотосинтеза. С увеличением светолюбия концентрация клеток и хлоропластов в листе возрастает, что увеличивает их общую поверхность (ИМК, ИМХ) и способствует ускорению поглощения углекислого газа мезофиллом. С другой стороны, возрастание концентрации клеток приводит к увеличению плотности листовых тканей (см. таблицу) и уменьшению относительного объема межклетников, что способствует снижению транспирационных потерь (Гамалей, 1988; Шереметьев, 25). Так, показано (Wright et l., 21), что растения более сухих местообитаний при том же уровне фотосинтеза, что и растения влажных мест, имели более низкую скорость транспирации. В условиях дефицита влаги устьица часто находятся в закрытом состоянии, и фотосинтез лимитируется недостатком поступления СО 2 внутрь листа. Поэтому ЭКОЛОГИЯ усиление ксероморфности листа связано с развитием внутрилистовой поверхности для облегчения диффузии углекислого газа. Действительно, изученные нами мезоксерофиты характеризовались максимальными значениями ИМК 22 28, при которых расчетная скорость продвижения молекул СО 2 из межклетников к хлоропластам была максимальной и достигала см/сек. Увеличение скорости поступления СО 2 в клетки должно соответствовать повышению уровня фотосинтеза. Например, у ксерофитов пустыни Каракумы отмечена более высокая интенсивность фотосинтеза единицы площади листа, чем у мезофитов умеренных широт, и основная причина этого была связана со значительно большим количеством клеток и хлоропластов и их поверхности у ксерофитов (Мокроносов, Шмакова, 1978). Таким образом, постепенное увеличение концентрации клеток и хлоропластов и их общей поверхности в экологическом ряду является закономерным адаптивным изменением, направленным на оптимизацию фотосинтетической функции растений. В наших исследованиях размеры клеток мезофилла меньше различались у растений разных экологических групп, чем количество клеток. При этом одной из линий адаптации растений к затенению часто называют крупные размеры клеток, а ксерофитизацию связывают с уменьшением размеров клеток листа (Мокроносов, Шмакова, 1978; Горышина, 1989; Зверева, 1986). Однако имеющиеся данные по размерам клеток в разных экологических условиях противоречивы. По данным некоторых авторов (Arus et l., 1986), тенелюбивые растения имели разные размеры клеток и хлоропластов, но сходные значения численной плотности хлоропластов и ИМК. Другие авторы также отмечали отсутствие различий между мезофитами и ксерофитами по средним размерам клеток хлоренхимы, но существенные различия в их численной плотности (Дьяченко, 1978; Ivnov et l., 24). Показано, что размеры клеток могут определяться размером генома и жизненной формой растения (Hodgson et l., 21; Иванова, 212), а изменение размеров клеток может служить одним из механизмов регуляции их концентрации в листе (Noel, 1977; Tershim et l., 21). Интересно также то, что число пластид в клетке у растений разных экологических групп было сходным (см. рис. 3). У пустынных и бореальных растений количество хлоропластов в клетках также существенно не различалось, несмотря на разницу в размерах клеток (Мокроносов, Шмакова, 1978). В целом полученные нами и литературные данные позволяют предположить, что изменение размеров клеток и числа хлоропластов в них не играет большой роли в адаптации растений к экологическим условиям, в то время как увеличение количества клеток и хлоропластов в единице площа- 3*
8 116 ИВАНОВА 25 Сциофиты Мезофиты и гигромезофиты Гелиофиты Объем клетки, 1 3 мкм Гигромезофиты Мезофиты 3 45 Ксеромезофиты Мезоксерофиты Ксерофиты степей и пустынь* Число клеток в единице площади листа, 1 3 см Рис. 4. Схематичное расположение экологических групп растений в поле параметров мезофилла листа. Положение на графике соответствует средним значениям параметров для данной группы; цифрами рядом со схемой поперечного среза указана удельная поверхностная плотность листа (УППЛ) в мг дм 2 ; * параметры ксерофитов степей и пустынь приведены согласно литературным данным (Мокроносов, Шмакова, 1978; Ivnov et l., 24). 16 ди листа вдоль градиента освещенности и водного дефицита имеет большое адаптивное значение. Выявленные закономерности адаптивных изменений мезофилла позволяют свести все структурное разнообразие экологических групп в единую схему в поле основных параметров количество клеток мезофилла в единице площади листа и их размеры (рис. 4). Учитывая, что отнесение вида к определенной экологической группе часто затруднено и носит субъективный характер, систему количественных показателей мезофилла, представленную на рис. 4, можно использовать для определения положения вида в экологическом ряду. При этом следует отметить отсутствие резких границ между экологическими группами растений, которое может быть связано с тем, что сами экологические группы растений не являются дискретными единицами (Миркин, Наумова, 1998). Предложенная нами схема демонстрирует непрерывность экологического ряда растений. На наш взгляд, для идентификации экологических свойств вида необходима оценка комплекса количественных показателей мезофилла (а не отдельно взятого признака), которая может объективно охарактеризовать степень приспособления данного вида к уровню освещения или водного дефицита. Использование данного комплекса показателей может также идентифицировать смещение свойств популяций при изучении экологической дифференциации внутри вида, поскольку направление внутривидовых изменений параметров мезофилла при воздействии экологических факторов совпадает с полученными закономерностями для растений разных экологических групп (Горышина, 1989; Иванова, Пьянков, 22; Иванова и др., 28). Следует также учитывать, что предложенная схема разработана для двудольных многолетних травянистых растений с летнезелеными листьями, в то время как древесные растения, однолетние формы, однодольные растения (злаки, осоки, лилейные), а также растения с вечнозелеными листьями могут иметь свои особенности адаптации структуры листа к затенению и увлажнению. ЗАКЛЮЧЕНИЕ Растения разных экологических групп бореальной зоны существенно различаются по параметрам структуры листа, имеющим адаптивное значение. Различия обнаружены в количественных параметрах мезофилла и не были связаны с морфологическими признаками листа. В ряду тенелюбивые мезофиты светолюбивые гигромезофиты мезофиты ксеромезофиты мезоксерофиты выявлено постепенное увеличение коли- ЭКОЛОГИЯ 2 214
9 АДАПТИВНЫЕ ПРИЗНАКИ СТРУКТУРЫ ЛИСТА РАСТЕНИЙ 117 чества клеток и хлоропластов в единице площади листа. В этом же ряду уменьшались размеры клеток и хлоропластов, но значительно в меньшей степени. Увеличение концентрации фотосинтетических элементов в листьях растений на градиенте освещения и увлажнения среды приводило к повышению плотности сложения листовых тканей, возрастанию площади внутрилистовой поверхности и усилению фотосинтетического потенциала единицы площади листа. Градуированный характер изменения параметров фотосинтетических тканей при адаптивной дивергенции видов с разными экологическими свойствами позволяет использовать систему количественных показателей мезофилла листа для объективной оценки положения вида в экологическом ряду травянистых растений бореальной зоны. Исследование выполнено при финансовой поддержке РФФИ (проект а). СПИСОК ЛИТЕРАТУРЫ Буинова М.Г., Бадмаева Н.К. Структурно-функциональные особенности листа растений житняково-ковыльных степей Западного Забайкалья // Эколого-биологические особенности растений и фитоценозов Забайкалья. Улан- Удэ: БНЦ СО АН СССР, С Варминг Е. Ойкологическая география растений. Введение в изучение растительных сообществ. М.: Типография И.А. Баландина, с. Василевская В.К. Формирование листа засухоустойчивых растений. Ашхабад: АН Туркм. ССР, с. Василевская В.К. Структурные приспособления растений жарких и холодных пустынь Средней Азии и Казахстана // Проблемы современной ботаники. М.; Л.: Наука, Т. 2. С Василевская В.К., Бутник А.А. Типы анатомического строения листьев двудольных (к методике анатомического описания) // Ботан. журн Т С Гамалей Ю.В. Структура растений Заалтайской Гоби // Пустыни Заалтайской Гоби / Под ред. Гамалея Ю.В., Гунина П.Д., Камелина Р.В., Слемнева Н.Н. Л., С Гамалей Ю.В. Транспортная система сосудистых растений. СПб.: Изд-во С.-Петерб. ун-та, с. Годнев Т.Н., Калишевич С.В. Наблюдения над увеличением числа и размеров хлоропластов и накоплением в них хлорофилла во время роста листьев // Сборник памяти акад. В.Н. Любименко. Киев: Изд-во АН УССР, С Горышина Т.К. Фотосинтетический аппарат растений и условия среды. Л.: Изд-во ЛГУ, с. Дьяченко А.П. Сравнительный анализ структурных и функциональных особенностей фотосинтетического аппарата различных экологических групп высших растений // Мезоструктура и функциональная активность фотосинтетического аппарата /Под ред. Мокроносова А.Т. Свердловск: Уральский ун-т, С Заленский В.Р. Материалы по количественной анатомии различных листьев одних и тех же растений // Изв. Киев. политех. ин-та Т С Зверева Г.К. Экологические особенности ассимиляционного аппарата степных растений Центральной Тувы // Экология С Зверева Г.К. Некоторые особенности структуры листа и его фотосинтетического аппарата у многолетних сеяных трав при разных уровнях увлажнения почвы // Сибирский вестник с.-х. науки С Иванова Л.А. Структурная перестройка мезофилла листа в ряду жизненных форм растений // Докл. РАН С Иванова Л.А., Пьянков В.И. Структурная адаптация мезофилла листа к затенению // Физиол. растений С Иванова Л.А., Иванов Л.А., Ронжина Д.А., Пьянков В.И. Структурные параметры мезофилла листа при затенении растений разных функциональных типов // Физиология растений. 28. T С Куликов П.В. Конспект флоры Челябинской области (сосудистые растения). Екатеринбург Миасс: Геотур, с. Лайск А., Оя В., Рахи М. Диффузионные сопротивления листьев в связи с их анатомией // Физиол. раст Т С Максимов Н.А. Физиологические основы засухоустойчивости растений. Приложение 26 к Трудам по прикладной ботанике и селекции. Л., с. Миркин Б.М., Наумова Л.Г. Наука о растительности (история и современное состояние основных концепций). Уфа: Гилем, с. Мокроносов А.Т. Мезоструктура и функциональная активность фотосинтетического аппарата // Мезоструктура и функциональная активность фотосинтетического аппарата / Под ред. Мокроносова А.Т. Свердловск: Уральский ун-т, С Мокроносов А.Т., Шмакова Т.В. Сравнительный анализ мезоструктуры фотосинтетического аппарата у мезофитных и ксерофитных растений // Мезоструктура и функциональная активность фотосинтетического аппарата / Под ред. Мокроносова А.Т. Свердловск: Уральский ун-т, С Пьянков B.И. Роль фотосинтетической функции в адаптации растений к условиям среды: Автореф. дис. докт. биол.наук. М.: ИФР РАН, с. Пьянков B.И., Мокроносов А.Т. Основные тенденции изменения растительности Земли при глобальном повышении температуры // Физиол. растений Т С Флора Сибири (в 14 томах). Новосибирск: Наука, Цельникер Ю.Л. Физиологические основы теневыносливости древесных растений. М.: Наука, с. Шереметьев С.Н. Травы на градиенте влажности почвы. М.: Тов. науч. изд. КМК, с. Эсау К. Анатомия растений. М.: Мир, с. Arus J.L., Alegre L., Tpi L. et l. Reltionships etween photosyntheti pity nd lef struture in severl shde plnts // Amer. J. Bot V P Hodgson J. G., Shrfi M., Jlili A. et l. Stomtl vs. genome size in ngiosperms: the somti til wgging the genomi dog? // Annls of Botny. 21. V. 15. P ЭКОЛОГИЯ 2 214
10 118 ИВАНОВА Ivnov L.A., Ronzhin D.A., Ivnov L.A. et l. Struturl nd funtionl sis of dpttion of Goi plnts to desertifition // Arid eosystems. 24. V P Noel P.S. Internl lef re nd ellulr CO 2 resistne: photosyntheti implitions of vritions with growth onditions nd plnt speies // Physiol. Plnt V. 4. P Poorter H., Niinemets U., Poorter L. et l. Cuses nd onsequenes of vrition in lef mss per re (LMA): metnlysis // New Phytologist. 29. V P Pynkov V.I., Ivnov L.A., Lmers H. Quntittive ntomy of photosyntheti tissues of plnts speies of different funtionl types in orel vegettion // Inherent vrition in plnt grouwth. Physiologil mehnisms nd eologil onseqenes / H. Lmers, H. Porter, M.M.I. Vn Vuuren Eds. The Netherlnd, Leiden: Bkhuys Pulishers, P Shimper A.F.W. Plnt-geogrphy upon physiologil sis. Trnsl. W.R. Fisher. Oxford, UK: Clrendon Press, р. Slton M.R., Smith W.K. Mesophyll rhiteture nd ell exposure to interellulr ir spe in lpine, desert, nd forest speies // Interntionl J. of Plnt Si. 22. V P Tershim I., Miyzw S., Hn Y.T. Why re sun leves thiker thn shde leves? Considertion sed on nlyses of CO2 diffusion in the lef // J. of Plnt Reserh. 21. V P Turrell F.M. The re of the internl exposed surfe of diotyledon leves // Am. J. Bot V. 23. P Wright I.J., Reih P.B., Westoy M. Strtegy-shifts in lef physiology, struture nd nutrient ontent etween speies of high nd low rinfll, nd high nd low nutrient hitts // Funtionl Eology. 21. V. 15. P ЭКОЛОГИЯ 2 214
docplayer.ru
Экологические группы растений по отношению к свету и их адаптивные особенности
Световой режим любого местообитания определяется интенсивностью прямого и рассеянного света, количеством света (годовой суммарной радиацией), его спектральным составом, а также альбедо – отражательной способностью поверхности, на которую падает свет.
Перечисленные элементы светового режима очень переменчивы и зависят от географического положения, высоты над уровнем моря, от рельефа, состояния атмосферы, характера земной поверхности, растительности, от времени суток, сезона года, солнечной активности и глобальных изменений в атмосфере.
У растений возникают различные морфологические и физиологические адаптации к световому режиму местообитаний.
По требованию к условиям освещения принято делить растения на следующие экологические группы:
1) светолюбивые (световые), или гелиофиты, – растения открытых, постоянно хорошо освещаемых местообитаний;
2) тенелюбивые (теневые), или сциофиты, – растения нижних ярусов тенистых лесов, пещер и глубоководные растения; они плохо переносят сильное освещение прямыми солнечными лучами;
3) теневыносливые, или факультативные гелиофиты, – могут переносить большее или меньшее затенение, но хорошо растут и на свету; они легче других растений перестраиваются под влиянием изменяющихся условий освещения.
Можно отметить некоторые общие приспособительные особенности, свойственные растениям каждой экологической группы.
Световые адаптации гелиофитов и сциофитов.Гелиофиты часто имеют побеги с укороченными междоузлиями, сильно ветвящиеся, нередко розеточные. Листья гелиофитов обычно мелкие или с рассеченной листовой пластинкой, с толстой наружной стенкой клеток эпидермы, нередко с восковым налетом или густым опушением, с большим числом устьиц на единицу площади, часто погруженных, с густой сетью жилок, с хорошо развитыми механическими тканями. У ряда растений листья фотометричные, т. е. повернуты ребром к полуденным лучам или могут менять положение своих частей в зависимости от высоты стояния Солнца. Так, у степного растения софоры листочки непарноперистосложного листа в жаркий день подняты вверх и сложены, у василька русского так же ведут себя сегменты перисторассеченного листа.
Оптический аппарат гелиофитов развит лучше, чем у сциофитов, имеет большую фотоактивную поверхность и приспособлен к более полному поглощению света. Обычно у них лист толще, клетки эпидермы и мезофилла мельче, палисадная паренхима двухслойная или многослойная (у некоторых саван-ных растений Западной Африки – до 10 слоев), нередко развита под верхней и нижней эпидермой. Мелкие хлоропласты с хорошо развитой гранальной структурой в большом числе (до 200 и более) расположены вдоль продольных стенок.
Хлорофилла на сухую массу в листьях гелиофитов приходится меньше, но зато в них содержится больше пигментов I пигментной системы и хлорофилла П700. Отношение хлорофилла а к хлорофиллу b равно примерно 5: 1. Отсюда высокая фотосинтетическая способность гелиофитов. Компенсационная точка лежит в области более высокой освещенности. Интенсивность фотосинтеза достигает максимума при полном солнечном освещении. У особой группы растений – гелиофитов, у которых фиксация СО2 идет путем С4-дикарбоновых кислот, световое насыщение фотосинтеза не достигается даже при самой сильной освещенности. Это растения из засушливых областей (пустынь, саванн). Особенно много С4-растений среди семейств мятликовых, осоковых, аизовых, портулаковых, амарантовых, маревых, гвоздичных, молочайных. Они способны к вторичной фиксации и реутилизации СО2, освобождающегося при световом дыхании, и могут фотосинтезировать при высоких температурах и при закрытых устьицах, что часто наблюдается в жаркие часы дня.
Обычно С4-растения, особенно сахарный тростник и кукуруза, отличаются высокой продуктивностью.
Сциофиты– это растения, постоянно находящиеся в условиях сильного затенения. При освещенности 0,1–0,2 % могут расти только мхи и селягинеллы. Плауны довольствуются 0,25-0,5 % полного дневного света, а цветковые растения встречаются обычно там, где освещенность в пасмурные дни достигает не менее 0,5–1% (бегонии, недотрога, травы из семейств имбирные, мареновые, коммелиновые).
В северных широколиственных и темнохвойных лесах полог сомкнутого древостоя может пропускать всего 1–2% ФАР, изменяя ее спектральный состав. Сильнее всего поглощаются синие и красные лучи, и пропускается относительно больше желто-зеленых лучей, дальних красных и инфракрасных. Слабая освещенность сочетается с повышенной влажностью воздуха и повышенным содержанием в нем СО2, особенно у поверхности почвы. Сциофиты этих лесов – зеленые мхи, плауны, кислица обыкновенная, грушанки, майник двулистный и др.
Листья у сциофитов располагаются горизонтально, нередко хорошо выражена листовая мозаика. Листья темно-зеленые, более крупные и тонкие. Клетки эпидермы крупнее, но с более тонкими наружными стенками и тонкой кутикулой, часто содержат хлоропласты. Клетки мезофилла крупнее, палисадная паренхима однослойная или имеет нетипичное строение и состоит не из цилиндрических, а из трапециевидных клеток. Площадь жилок вдвое меньше, чем у листьев гелиофитов, число устьиц на единицу площади меньше. Хлоропласты крупные, но число их в клетках невелико.
У сциофитов по сравнению с гелиофитами меньше хлорофилла П700. Отношение хлорофилла а к хлорофиллу b равно примерно 3: 2. С меньшей интенсивностью протекают у них такие физиологические процессы, как транспирация, дыхание. Интенсивность фотосинтеза, быстро достигнув максимума, перестает возрастать при усилении освещенности, а на очень ярком свету может даже понизиться.
У лиственных теневыносливых древесных пород и кустарников (дуба черешчатого, липы сердцевидной, сирени обыкновенной и др.) листья, расположенные по периферии кроны, имеют структуру, сходную со структурой листьев гелиофитов, и называются световыми, а в глубине кроны – теневые листья с теневой структурой, сходной со структурой листьев сциофитов (рис. 24).

Рис. 24. Поперечный срез светового (слева) и теневого (справа) листа сирени (по И. С. Михайловской, 1977)
Факультативные гелиофиты, или теневыносливые растения, в зависимости от степени теневыносливости имеют приспособительные особенности, сближающие их то с гелиофитами, то со сциофитами. К этой группе можно отнести некоторые луговые растения, лесные травы и кустарники, растущие и в затененных участках леса, и на лесных полянах, опушках, вырубках. На осветленных местах они разрастаются часто сильнее, однако оптимальное использование ФАР у них происходит не при полном солнечном освещении.
У деревьев и кустарников теневая или световая структура листа часто определяется условиями освещения предыдущего года, когда закладываются почки: если закладка почек идет на свету, то формируется световая структура, и наоборот.
Если в одном и том же местообитании закономерно периодически изменяется световой режим, растения в разные сезоны могут проявлять себя то как светолюбивые, то как теневыносливые.
Весной в дубравах под полог леса проникает 50–60 % солнечной радиации. Листья розеточных побегов сныти обыкновенной имеют световую структуру и отличаются высокой интенсивностью фотосинтеза. В это время они создают основную часть органического вещества годичной продукции. Листья сныти летней генерации, появляющиеся при развитом древесном пологе, под который проникает в среднем 3,5 % солнечной радиации, имеют типичную теневую структуру, и интенсивность фотосинтеза их значительно ниже, в 10–20 раз. Подобную двойственность по отношению к свету проявляет и осока волосистая, светолюбивая весной и теневыносливая летом. По-видимому, это свойственно и другим растениям дубравного широкотравья.
Отношение к световому режиму меняется у растений и в онтогенезе. Проростки и ювенильные растения многих луговых видов и древесных пород более теневыносливы, чем взрослые особи.
Иногда у растений меняются требования к световому режиму, когда они оказываются в иных климатических и эдафических условиях. Так, обычные теневыносливые растения хвойного леса – черника, седмичник европейский и некоторые другие – в тундре приобретают особенности гелиофитов.
Наиболее общая адаптация растений к максимальному использованию ФАР – пространственная ориентация листьев. При вертикальном расположении листьев, как, например, у многих злаков и осок, солнечный свет полнее поглощается в утренние и вечерние часы – при более низком стоянии солнца. При горизонтальной ориентации листьев полнее используются лучи полуденного солнца. При диффузном расположении листьев в разных плоскостях солнечная радиация в течение дня утилизируется наиболее полно. Обычно при этом листья нижнего яруса на побеге отклонены горизонтально, среднего направлены косо вверх, а верхнего располагаются почти вертикально.
Считают, что кукуруза является одной из самых высокопродуктивных сельскохозяйственных культур потому, что наряду с высоким КПД фотосинтеза у нее наблюдается диффузное расположение листьев, при котором полнее поглощается ФАР.
На севере, где высота стояния солнца меньше, встречается больше растений с вертикальным расположением листьев, а на юге – с горизонтальным. Для получения большей биомассы выгодны также посевы и насаждения, в которых сочетаются растения с разной пространственной ориентацией листьев, причем в верхнем ярусе лучше иметь растения с вертикальным расположением листьев, которые полнее используют свет при низком стоянии солнца, не препятствуют прохождению полуденных лучей к расположенным в нижнем ярусе листьям с горизонтальной ориентацией.
lektsia.info
Анатомическая характеристика листа, как основного фотосинтезирующего органа растения. Адаптивные признаки строения мезоструктуры фотосинтетического аппарата, возникшие у высших растений в определенных условиях среды
ДИСЦИПЛИНА СПЕЦИАЛИЗАЦИИ«МЕЗОСТРУКТУРА ФОТОСИНТЕТИЧЕСКОГО АППАРАТА РАСТЕНИЙ»
ЛЕКЦИЯ
Введение. Анатомическая характеристика листа, как основного фотосинтезирующего органа растения. Адаптивные признаки строения мезоструктуры фотосинтетического аппарата, возникшие у высших растений в определенных условиях среды.
Познание структуры и функции фотосинтетического аппарата, механизмов его регуляции имеет не только теоретическое, но и большое практическое значение.
С одной стороны, это открывает большие возможности эффективного управления формированием урожая за счет повышения интенсивности и качества образующихся продуктов фотосинтеза.
С другой стороны, это позволит использовать в промышленности отдельные реакции и структуры, которые были «изобретены» растениями в ходе эволюции процесса использования солнечного света для осуществления различных видов работы в клетках.
Важным является и использование знаний об изменениях в структуре фотосинтетического аппарата растений в экологии, характеризующих связь с окружающей средой. Изменение мезоструктуры листа можно рассматривать как существенное проявление регуляции фотосинтеза на морфологическом уровне, обеспечивающее оптимизацию и адаптацию фотосинтетического аппарата при различных экологических условиях.
Все это определило большой интерес исследователей к изучению мезоструктуры фотосинтетического аппарата растений. На кафедре ботаники КБГУ данное направление стало развиваться с 1986 года по предложению Слонова Л.Х. как «Мезоструктура и функциональная активность фотосинтетического аппарата». Было выделено два направления исследований:
- Изучение структурных и функциональных характеристик фотосинтетического аппарата различных систематических групп растений и наиболее распространенных культурных растений, выращиваемых в КБР.
- Формирование фотосинтетического аппарата растений под влиянием различных условий произрастания.
Для усвоения этого курса необходимо познакомиться со следующей литературой ( см. Список по рабочей программе).
^
У высших однодольных и двудольных растений фотосинтезирующие клетки обнаруживаются в тканях нескольких типов, локализованных, как правило, в листьях – специальных органах, приспособленных для осуществления фотосинтетической функции. Форма и анатомическое строение листьев обеспечивают, с одной стороны, рациональное пространственное размещение хлорофилла в надземной части растительного сообщества и поэтому достаточно эффективное поглощение световой энергии и, с другой – поглощение углекислого газа из окружающего воздуха и отведение образовавшихся ассимилятов в нефотосинтезирующие или слабо фотосинтезирующие органы.
- Кутикула. Воск и входящий в состав кутикулы кутин гидрофобны, что затрудняет прохождение воды через покровную ткань листа.
- Строение устьичного аппарата у разных видов растений.
- Основная масса хлоропластов сосредоточена в клетках двух типов паренхимы листьев: губчатой и столбчатой.
- Проводящий пучок.
- Хлоропласты
Лекция
Особенности формирования мезоструктуры листа и фотосинтетической активности в зависимости от экологических условий.
Мезоструктура листа большинства высших растений имеет целый ряд специфических особенностей. Если рассматривать поперечный срез листовой пластинки, то можно увидеть кутикулу.
КУТИКУЛА (от лат. сuticula-кожица) – слой жирового вещества (кутина), покрывающего пленкой поверхность листьев. Кутикула малопроницаема для водных растворов, газов, болезнетворных организмов. Кутин синтезируется клетками эпидермиса и выделяется наружу, формируя кутикулу и частично пропитывая наружную стенку клеточной оболочки.
Комплекс кутикулы и кутинизированной оболочки включает целлюлозу, пектин, кутин, воск и другие инкрустирующие вещества и имеет слоистую структуру. Толщину кутикулы пронизывает сеть гидрофильных пектиновых капилляров. Степень развития кутикулы, характер распределения в ней воска и гидрофильных капилляров в значительной мере определяют защитные свойства эпидермы и зависят от условий обитания и возраста растения.
Мощная, плотно покрытая кристаллами воска кутикула характерна для ксерофитов. Рельеф кутикулы повторяет и часто усиливает рельеф наружных клеток эпидермальных клеток, в результате чего создается структура поверхности, специфичная для видов. Таким образом, наличие и состояние кутикулы является диагностическим признаком.
Важная роль кутикулы – регуляция испарения воды. При кутикулярной транспирации молекулы воды диффундируют через кутинизированные слои наружных стенок эпидермиса и через кутикулу. Поэтому можно рассматривать кутикулярную транспирацию как диффузию через гидрофобную среду. Кутикулярное диффузное сопротивление в большинстве случаев очень велико, причем у разных видов оно различно в зависимости от расположения, плотности и числа прослоек кутина и воска, а также от толщины кутикулы.
Кроме того, на формировании изолирующих структур на поверхности листьев сильно сказываются условия произрастания: при атмосферной и почвенной засухе взрослые листья обладают более толстой кутикулой и более плотным восковым налетом, чем у побегов, которые развивались при большей влажности воздуха.
У тонких листьев кутикулярное сопротивление диффузии составляет 20-100 с/см, у листьев и хвои с массивной защитой от транспирации оно может достигнуть величин около 400 с/см. При отбухании и подсыхании наружных стенок эпидермиса гидрофобные слои плотнее придвигаются друг к другу, поэтому кутикулярное сопротивление может удвоиться.
При низких температурах сопротивление диффузии также возрастает. Куткулярная защита от транспирации весьма эффективна. Даже у растений затененных и влажных местообитаний кутикулярная транспирация составляет менее 10% от свободного испарения, у жестколиственных и вечнозеленых хвойных растений она сокращается до 0,5%, а у кактусов даже до 0,05% испаряющей способности. Весь диапазон между этими крайними случаями заполняют светолюбивые травы, деревья и кустарнички.
ЭПИДЕРМИС или эпидерма (epi- греч.-на, derma- кожа, кожица) – первичная покровная ткань, образующаяся из протодермы конуса нарастания на всех молодых листьях. Клетки эпидермы таблитчатые, однослойные, без межклетников. Через поры и пектиновые тяжи в наружных стенках клеточных оболочек эпидермы, проникают вода и питательные вещества.
В протодерме листа некоторых растений происходят параллельные переклинальные деления, в результате чего образуется многослойная эпидерма. Она обнаружена, например, в листьях таких известных комнатных растений как фикус, пеперомия. Считается, что такая эпидерма служит водозапасающей тканью.
В основной своей массе клетки эпидермы плотно примыкают друг к другу, это живые клетки, плоские, центральное место в них занимает вакуоль, ядро, а лейкопласты смещены к периферии, хлоропласты отсутствуют. Исключения составляют некоторые водные растения, папоротники, тенелюбивые покрытосеменные.
Эпидермис выполняет барьерную роль: защищает внутренние ткани от повреждения и высыхания, обеспечивает газообмен, транспирацию, всасывание и секрецию различных веществ (гормонов, ферментов). Газообмен и обмен парами воды между окружающим воздухом и межклеточным пространством листьев обеспечивается с помощью устьиц.
УСТЬИЦЕ – (stoma), высокоспециализированное образование эпидермиса растений, состоящее из двух замыкающих клеток и межклетника (устьичной щели) между ними.
Строение устьичного аппарата у разных видов растений может сильно отличаться. В большинстве случаев устьичные щели ограничены двумя замыкающими клетками бобовидной формы, имеющими утолщенные стенки, обращенные к устьичной щели, и тонкие- противоположные. При повышении тургорного давления происходит искривление клеточных стенок: тонкая становится выпуклой, а толстая - вогнутой, за счет чего и происходит открывание устьичной щели. У злаков строение замыкающих клеток устьиц иное. На концах замыкающих клеток, там, где они соприкасаются друг с другом, имеется по одному тонкостенному отделению. Остальная часть клетки имеет толстые стенки. При повышении тургорного давления объем тонкостенных отделений увеличивается, что приводит к расхождению друг о друга ранее соприкасавшихся толстыми стенками замыкающих клеток.
У злаков есть виды, имеющие устьица с двумя замыкающими клетками и двумя вспомогательными клетками, с двумя замыкающими и четырьмя вспомогательными клетками.
Лекция
^


 Формирование листа начинается с генетически детерминированного числа актов пролиферации, упорядоченных в пространстве, чем определяется общее число клеток в листе и форма листа.
Формирование листа начинается с генетически детерминированного числа актов пролиферации, упорядоченных в пространстве, чем определяется общее число клеток в листе и форма листа.
Последовательная дифференцировка ведет к формированию специализированных тканей и клеток листа, в том числе фотосинтезирующих клеток (палисадных, губчатых, обкладочных, мезофилльных), системы устьиц, анатомических структур ближнего и дальнего транспорта.
Развитие листа включает:
1. формирование системы межклетников;
2. оптимизацию численности и распределения пластид в листовой пластинке;
3. изменяется соотношение процессов синтеза и распада структур разной сложности;
4. соотношение анаболических и катаболических процессов.
^ соответствует наиболее сбалансированному соотношению этих процессов, а период старения — преобла-нию процессов деструкции. Обычно максимум функциональной активности листа, в том числе и по фотосинтезу, наступает раньше, чем лист достигает максимальной площади.
Существенным фактором возрастной физиологии листа является преобладание импорта ассимилятов растущим ювенильным листом из более зрелых листьев или запасающих органов и абсолютное доминирование экспорта ассимилятов зрелым листом.
Стареющие листья, особенно многолетние листья вечнозеленых растений, все более утрачивают фотосинтетическую функцию и обретают вторичные функции (депонирование и распределение ассимилятов, синтез веществ вторичного происхождения и др.).

 Каждому уровню структурно-функциональной организации присущи свои специфические проявления онтогенетических изменений фотосинтеза. Только выделяя одну за другой последовательные иерархические ступени фотосинтетических систем и прослеживая развитие каждой в отдельности, можно приблизиться к истинной сложности поставленной в этой книге задачи.
Каждому уровню структурно-функциональной организации присущи свои специфические проявления онтогенетических изменений фотосинтеза. Только выделяя одну за другой последовательные иерархические ступени фотосинтетических систем и прослеживая развитие каждой в отдельности, можно приблизиться к истинной сложности поставленной в этой книге задачи.
Важную роль в мезоструктуре листа отводится хлоропластам.
^ включает согласованную последовательность синтеза пигментов, компонентов электротранспортной цепи, структурных и функциональных белков, в том числе ферментов фотосинтетического цикла. Синтез этих компонентов сопряжен с формированием мембранной системы хлоропласта.
Формируется сначала ламеллярная, затем гранальная системы пластидных мембран. Соответственно такой последовательности развития мембранной системы фс 1 формируется раньше, чем фс 2.
Старение хлоропласта проявляется в потере рибосом, накоплении малоактивных структур и соединений (пластоглобулы), в изменении липидного комплекса мембран и постепенной деструкции мембранной системы.
Развитие фотосинтезирующей клетки осуществляется как результат координированного взаимодействия двух генетических систем — генома_ и пластома.
В процессе роста и дифференцировки клетки образуется генетически детерминированное число хлоропластов. Число пластид в клетке может отклоняться от нормы и определяется главным образом количеством и фенотипическим варьированием ядерной ДНК.
Система целого растения обеспечивает управление в онтогенезе фотосинтетической функцией на всех более низких уровнях организации. Кроме того, существует организменный уровень регуляции фотосинтеза, частично рассмотренный. Реализация генетической программы развития и формирование фенотипа на ее основе нуждаются в фотосинтетическом обеспечении функции роста и морфогенеза.
В онтогенезе растения происходит смена ярусов или поколений листьев, при этом каждый новый лист (source) оказывается в новой функциональной связи с акцептором.
Иногда лист более или менее строго специализируется на обеспечении ассимилятами ближайшего по расположению плода, колоса или вегетативного запасающего органа,
Это обстоятельство чрезвычайно важно для практического растениеводства.
Из этого положения следует несколько выводов:
- В современном земледелии продукционный процесс часто лимитируется фотосинтетическим обеспечением ростового процесса. Однако по мере оптимизации условий фотосинтеза, особенно в условиях промышленной фитотроники, все чаще ограничивающим фотосинтез фактором будет выступать не внешняя среда, а общая напряженность и направленность процессов эпигенеза. Тогда единственным путем интенсификации фотосинтеза станет эндогенная регуляция этой функции через эпигенетические процессы;
- Направленная интенсификация фотосинтеза селекционным путем должна быть сбалансирована с генетической программой развития;
- Констатация онтогенетических изменений фотосинтеза составляет лишь одну, наиболее доступную для исследователя грань проблемы.
^ феногенез фотосинтетической системы в связи с иерархией регуляторных систем механизмов на всех уровнях от молекулярного до организменного. Именно это направление может быть основой радикальных воздействий на фотосинтез в исследовательском эксперименте и в растениеводческой практике.
Можно выделить три типа регуляции
1 -Стабилизирующее регулирование — поддержание постоянства регу
лируемой величины. В биологических системах выражается в поддержании гомеостаза системы и относительной стабилизации скорости процессов. Уже давно было отмечено, что в фотосинтезе отчетливо выражены две противоположные тенденции: сохранять относительно устойчивое состояние при изменении внешних условий и, наоборот, изменяться при смене среды и физиологического состояния растений. Первая обеспечивает высокую стабильность работы фотосинтетического аппарата в широком диапазоне, вторая — постоянное изменение интенсивности и качественной направленности процесса в соответствии с общей тенденцией изменений среды и физиологического состояния растения.
^ _величиныпо заданной программе. По-существу, этот тип управления составляетоснову развития фотосинтетического аппарата в онтогенезе, которое,в свою очередь, является составным элементом общей генетическойпрограммы развития. Регуляция в онтогенезе обеспечивается изменениемдифференциальной активности генов. Для этого вида управления характерно определенное соотношение двух типов обратной связи — отрицательной и положительной. Например, на ранних этапах онтогенеза(экспоненциальный участок кривых формирования системы) преобладаетмеханизм положительной обратной связи, при котором увеличение выходных величин (ферментов, гормонов, хлоропластов, площади листьев)выполняет функцию усилителя регулируемого процесса. Наоборот,на поздних этапах развития фотосинтетической системы доминируютмеханизмы отрицательной обратной связи, различные формы автоингибирования, характерные для стареющих систем.3. ^ — изменение регулируемой величиныв соответствии с изменением внешней среды. Этот тип управления обеспечивает адаптацию фотосинтеза к условиям среды на уровне быстрых,кратковременных изменений длительностью секунды—десятки минут(быстрое, динамическое регулирование на уровне изменения активностиферментов или условий протекания ферментных реакций) или медленных изменений длительностью часы—сутки (медленное, оперонное регулирование на уровне индукции или репрессии генов). Фенотипическаяизменчивость фотосинтетического аппарата в рамках одного генотипаявляется следствием регуляторных процессов этого типа.
Если каждый из этих трех вариантов регуляции фотосинтеза проследить на всех уровнях от молекулярного до ценотического, то получится удивительная по совершенству и композиции система саморегуляции фотосинтеза.
Лекция
Характеристика света. Оптические свойства пигментов.
Лучистая энергия, необходимая для растения, представляет собой энергию электромагнитных колебаний, возникающих в результате колебательных движений электронов возбужденных атомов и молекул. Лучистая энергия распространяется в виде отдельных порций - квантов, или фотонов, величина которых определяется длиной волны или частотой.
Согласно закону Эйнштейна, интенсивность фотохимических процессов зависит не от абсолютного поглощения энергии безотносительно к ее квантовой природе, а от числа поглощенных квантов определенной величины. При одном и.том же поглощении энергии количество поглощенных квантов возрастает вместе с длиной волны.
Спектр солнечного света по отношению к физиологической характеристике ограничивается длинами волн от 6000 нм. Фотосинтетически активная радиация (ФАР), поглощаемая пигментами листа и обеспечивающая нормальный рост и развитие растений, находится в пределах 300—-750 нм.
Короткие ультрафиолетовые лучи с длиной волны
Сине-фиолетовые лучи с длиной волны 400—500 нм, поглощаемые хлорофиллом, каротиноидами, ферментами и другими компонентами клетки, в большинстве случаев задерживают переход растения к цветению, благоприятствуют синтезу белков и влияют на химический состав растений.
Зеленые лучи с длиной волны 500—600 нм, поглощаемые на 75%, обладают минимальной активностью, физиолого-биохимические процессы сильно заторможены, и выращивание растений нерационально из-за большого расхода лучистой энергии.
Оранжево-красные лучи с длиной волны 600—700 нм обладают максимальной активностью и поглощаются только хлорофиллом на 95%. В этих лучах с максимальной скоростью совершается фотосинтез, образование листьев, формирование корнеплодов, луковиц, стеблевых утолщений, переход растения к цветению и др. При этом наблюдается максимальный урожай.
Инфракрасные лучи с длиной волны 700—1000 нм не поглощаются или поглощаются очень мало (15%) пигментами и тканями растений. Физиологического значения они практически не имеют, и поэтому их излучение называют абиотической радиацией.
Далекие инфракрасные лучи с длиной волны 1000— 6000 нм, поглощаемые на 90% в основном водой, пигментами, цитоплазмой, определяют тепловой режим тканей растения. Они влияют на скорость и направление физиолого-биохимических процессов в соответствии с температурой окружающей среды. При низких температурах воздуха (30°С) они отрицательно влияют на обмен веществ.
Видимую солнечную радиацию с длиной волны 400— 720 нм растения поглощают неодинаково: мезофиты на 78%, ксерофиты на 86%, суккуленты на 88%, а растения, окрашенные антоцианами, на 92%.
В общем можно сказать, что поглощение видимой радиации растения возрастает с уменьшением широты местности, с увеличением прозрачности атмосферы, с высотой над уровнем моря, с онтогенезом и преимущественно у древесных.
Для нормального фотосинтеза, кроме воды и световой энергии, необходим углекислый газ, которого в атмосфере в среднем 0,03%. Максимальное количество его в почвенном воздухе до 0,5%, он поступает туда в результате разложения органических остатков. За счет градиента активности молекул углекислый газ накапливается на поверхности почвы, откуда конвекционными воздушными потоками поднимается вверх в атмосферу. В связи с тем что углекислый газ имеет большую плотность, чем воздух, он опускается в пониженные места на поверхности Земли. Поэтому минимальное его количество (0,006%) отмечено высоко в горах.
Для поглощения углекислого газа растения имеют огромную поверхность клеток паренхимы листьев, исчисляемую гектарами. Пропуская через листья воздух, растение буквально вылавливает молекулы углекислого газа.
Чтобы синтезировать 4 кг глюкозы, растение поглощает 3 м3 углекислого газа из 10000 м3 воздуха.
Хлорофиллы а и б различаются спектрами поглощения: у б полоса поглощения несколько смещена в сторону коротковолновых лучей, а в сине-фиолетовой области максимум поглощения смещен в сторону длинноволновых, т.е. красных лучей.
^
Онтогенез листа и возрастная динамика фотосинтеза
Развитие листа начинается с образования в периферической зоне апикальной меристемы листового бугорка — листового примордия. Серия периклинальных делений клеток в листовом бугорке приводит к закладке листа. Гистогенез листа и его развитие во времени сильно отличаются у растений разных жизненных форм. Например, у древесных растений значительная часть гистогенеза листа, совершается при формировании почек, которые после перезимовки возобновляют рост и днфференцировку в следующем году. У большинства травянистых двудольных растений гистогенез, как правило, не имеет такого перерыва.
Следом за формированием листового примордия развивается листовая пластинка. Пластинка листа формируется из так называемых маргинальных меристем, которые тянутся вдоль оси листа. Маргинальные меристемы начинают функционировать очень рано, когда примордии имеют длину меньше I мм. Эти меристемы дифференцируются на два тяжа — поверхностный и внутренний, которые называют маргинальными и субмаргинальными инициалями. Благодаря организованному сочетанию переклинальных, антиклинальных и косых делений клеток из маргинальных инициалей формируется верхний- и нижний эпидермис, а из субмаргннальных — палисадная и губчатая ткани мезофилла и проводящие пучки. Анатомы отмечают, что строгая идентификация маргинальной и субмаргинальной инициален оказывается затруднительной [Эсау, 1969].
Неравномерность деления и растяжения клеток в разных слоях пластинки приводит к дифференцировке разных тканей. Деление и растяжение 1 клеток губчатой паренхимы завершается раньше, чем в палисадной ткани. В эпидермисе деление клеток заканчивается раньше, чем в мезофилле. После завершения делений клетки эпидермиса очень интенсивно и многократно увеличиваются в горизонтальной плоскости, в то время как палисадные клетки растут вертикально, что является одной из причин обособления ранее плотно сомкнутых палисадных клеток и вызывает образование межклетников. Аналогично формируются межклетники в губчатой ткани. Таким образом, разрастание эпидермальных клеток в горизонтальной плоскости в период интенсивного роста листа растаскивает клетки мезофилла и способствует формированию структуры листа, в которой уменьшается сопротивление диффузии С02 к центрам карбоксилирования. Кроме того, это явление имеет и другое последствие — изменяется (обычно уменьшается) число хлоропластов в единице площади листа.
Лекция
^
Функции листа, как и всякого другого органа в растительном организме, многообразны. Мы уже говорили о листе как об органе транспирации, однако главной его функцией является фотосинтез. Выполнение листом этой функции связано с содержащимися в хлоропластах фотосинтетическими пигментами. Рассмотрим прежде всего особенности строения листа как фотосинтетического аппарата.
Нижняя и верхняя эпидерма листовой пластинки, если не считать замыкающих клеток устьиц, состоит из клеток с большими вакуолями, лишенных хлоропластов. Такие клетки хорошо пропускают свет в мезофилл и, следовательно, непосредственного участия в фотосинтезе не принимают. Эпидермальные покрытые кутикулой и воском, уменьшая транспирацию,
помогают поддерживать водный гомеостаз листа. Последнее очень важно, так как скорость фотосинтеза зависит от количества воды в тканях (о чем речь пойдет дальше). С другой стороны, через кутикулу проходит в 20—30 раз меньше СО2, чем через устьица. Создается противоречие между водным и газовым обменами, о чем мы уже упоминали, и только наличие устьиц попеременно то открывающихся, то закрывающихся и регулирующих таким образом скорость транспирации и скорость поступления углекислого газа из атмосферы, может снять указанное противоречие. Прогрессивная эволюция наземных растений была бы в сущности невозможна, если бы не было «изобретено» такое регулирующее устройство, как устьичный аппарат. Таким образом, эпидерма задерживает воду и пропускает свет.
Устьица — основные ворота для СО2. Правда, есть наблюдения, что у некоторых растений, например у яблони, углекислый газ может поступать в лист через временные отверстия в кутикуле.
Устьица работают по принципу обратной связи: СО2 используется на фотосинтез, его концентрация в межклетниках уменьшается, устьице открывается; СОг поступает в лист, его концентрация в межклетниках увеличивается, устьица закрываются.
Мезофилл у большинства растений состоит из столбчатой и губчатой паренхимы. В клетках мезофилла содержатся хлоропласты, здесь и происходит вся фотосинтетическая деятельность зеленого растения, поэтому важно рассмотреть особенности мезофилла и то, как он приспособлен к поглощению углекислого газа из атмосферы. Столбчатая паренхима, расположенная под верхней эпидермой, поглощает больше света, чем губчатая. Она является главной тканью, где идет фотосинтез. Для губчатой паренхимы характерно наличие межклетников. Последние, во-первых, помогают газообмену листа. Объем их составляет до 15—20% общего объема листовой пластинки. Благодаря верхней и нижней эпидерме, а также межклетникам, в листе создается внутренняя газовая среда, которая, хотя и сообщается с внешней средой при помощи устьичных щелей, практически всегда отличается от нее по своему составу. Во-вторых, межклетники значительно увеличивают внутреннюю поверхность листа: она в 7—10 раз больше, чем наружная поверхность. Следовательно, если на 1 га пашни листья всех растений имеют площадь 5 га, то поверхность, поглощающая СО2, равна 35—50 га. Это важно в связи с небольшим содержанием углекислого газа в атмосфере (0,045%). Однако одновременно увеличивается опасность обезвоживания тканей листа. Так как фотосинтез идет главным образом в столбчатой паренхиме, ее называют ассимиляционной. Доказательством того, что столбчатая и губчатая паренхимы выполняют разные функции, являются и подсчеты числа хлоропластов в их клетках.
Так, у клещевины в одной клетке столбчатой паренхимы содержится 36, а губчатой—20 хлоропластов. Вообще число хлоропластов в клетке сильно варьирует: от 20 до нескольких сотен. В пересчете на 1 мм2 поверхности листа число хлоропластов может достигать почти 1 млн. Усиленное азотное питание и хорошее водоснабжение растений вызывают увеличение размеров мезофильных клеток и число хлоропластов в них. Суммарная поверхность всех хлоропластов может превышать в десятки раз поверхность листовой пластинки, что также способствует лучшему поглощению СОа.
Таким образом, внутренняя поверхность листа, поглощающая углекислый газ и свет (о последнем речь еще впереди), во много раз больше поверхности листовой пластинки, что помогает более интенсивному протеканию фотосинтеза. Поэтому лист лучше, чем другие органы, приспособлен к выполнению фотосинтетической функции, хотя фотосинтез идет и в зеленых клетках стеблей, цветков, плодов.
Мезофилл пронизан сетью проводящих пучков, в состав которых входят сосуды, доставляющие хлоропластам воду и минеральные соли, и ситовидные трубки, отводящие от хлоропластов продукты фотосинтеза. Прямого контакта проводящих пучков с каждой клеткой нет. Поэтому в листе, как и в корне, транспорт веществ для фотосинтеза и отток продуктов фотосинтеза идет не только по специальным проводящим тканям — .ксилеме и флоэме, но и по симпласту и апопласту.
В зависимости от внешних условий анатомическое строение листа может значительно изменяться. Так, соотношение между столбчатой и губчатой паренхимами в мезофилле в зависимости от освещения резко меняется. Например, при слабом освещении сохраняется только одна губчатая паренхима.

Рис. 1. Строение листовой пластинки у кукурузы — С4-растение
Еще более интересные отклонения от типичного строения листа, влияющие на фотосинтез, связаны с генетическими различиями. Для примера приведем строение листовой пластинки кукурузы или сахарного тростника. Для этих растений характерно большое количество проводящих пучков. Каждый проводящий пучок окружен одним слоем крупных клеток хлорофил-лоносной паренхимы, образующих обкладку проводящего пучка (рис. 1). Паренхимная обкладка, в свою очередь, окружена одним или несколькими слоями мелких клеток палисадной паренхимы, между которыми имеются маленькие межклетники. Между клетками обкладки межклетников нет. Поскольку мезофилл и обкладка образуют концентрические слои воруг проводящего пучка, хорошо видимые на поперечных срезах, такое строение получило название «кранц-анатомии». Фотосинтез идет и в клетках мезофилла, и в клетках обкладки проводящего пучка.
300>300>medznate.ru
15.4.2. Адаптивные экологические типы человека
расовые признаки составляют только 8%, в то время как основная доля генетического разнообразия определяется многообразием отдельных индивидуумов. Иными словами, немец может быть генетически гораздо ближе к полинезийцу, чем
кдругому немцу, живущему в соседней квартире. Изучение геногеографии популяций человека показало, что географическое распределение частот генов групп крови системы АВ0, MN, Lutheran, Duny, Diego и др., а также различных форм ферментов и иммуноглобулинов не соответствует ареалам расселения ни одной из рас. Так, по группам крови АВ0 и MN жители Европы оказываются ближе
кафриканцам, в то время как по системе иммуноглобулинов они ближе к монголоидам Азии. Сходные результаты получены и в отношении распределения в популяциях вариантов митохондриальной ДНК.
Эти данные свидетельствуют о том, что биохимический полиморфизм человека
эволюционно возник раньше и развивался дольше по сравнению с возникновением комплексов расовых признаков. Из этого следует, что расы не представляют собой особых изолированных групп людей, характеризующихся наборами специфических генов. Расовые же характеристики являются не более чем отдельными проявлениями общего генетического полиморфизма, выражающегося в первую очередь в сложных морфологических признаках. Некоторые из них адаптивны, другие сформировались на основе коррелятивной изменчивости, но все они касаются лишь ряда второстепенных особенностей (цвета кожи, волос, глаз и т.д.) и не затрагивают таких общечеловеческих признаков, как морфология головного мозга, а также строение и функции руки как органа труда.
На основании определения числа аллелей, свойственных той или иной группе организмов, возможно определение генетического расстояния между ними. Эта величина для больших рас человека составляет 0,03. Она гораздо ниже цифр, характерных для истинных подвидов(0,17—0,22),и еще более мала по сравнению с межвидовым расстоянием(0,5—0,6и более). В животном мире генетическое расстояние, равное 0,03, соответствует обычно генетическим отличиям местных популяций друг от друга. Все эти данные свидетельствуют о том, что понятие расы условно, второстепенно и не позволяет подводить под иерархическую классификацию рас глубокую биологическую, а значит, и социальную базу.
Человечество, заселившее уже около 15 тыс. лет назад все более или менее благоприятные для жизни природно-географическиезоны, встретилось с необходимостью адаптироваться к самым разнообразным условиям существования. Адаптации человека к среде, как уже указывалось, проявляются в основном на социальном уровне, однако человечество на ранних этапах эволюции подвергалось непосредственному действию биотических и абиотических экологических факторов в значительно большей степени по сравнению с современной эрой научнотехнического прогресса. Комплексы таких факторов имели разнонаправленное действие на человеческие популяции. В результате в разных
165
климатогеографических зонах сформировались разнообразные адаптивные типы людей.
Адаптивный тип представляет собой норму биологической реакции на комплекс условий окружающей среды и проявляется в развитии морфофункциональных, биохимических и иммунологических признаков, обеспечивающих оптимальную приспособленность к данным условиям обитания.
Вкомплексы признаков адаптивных типов из разных географических зон входят общие испецифические элементы. К первым относят, например, показателикостно-мускульноймассы тела, количество иммунных белков сыворотки крови человека. Такие элементы повышают общую сопротивляемость организма к неблагоприятным условиям среды. Специфические элементы отличаются разнообразием и тесно связаны с преобладающими условиями в данном месте обитания — гипоксией, жарким или холодным климатом. Именно их сочетание служит основанием к выделению адаптивных типов: арктического, тропического, зоны умеренного климата, высокогорного, пустынь и др.
Разберем особенности условий жизни человеческих популяций в различных климатогеографических зонах и адаптивные типы людей, сформировавшиеся в них.
Условия обитания в Арктике характеризуются постоянно низкими температурами воздуха, скудной растительностью, богатством животного мира и сезонной периодичностью поступления продуктов питания: растительная пища доступна только в короткие летние месяцы, а животная — в периоды нереста рыбы, гнездования птиц, размножения оленей и морского зверя. Продукты питания богаты витаминами, белками, жирами и микроэлементами, но бедны углеводами растительного происхождения. Воздух и почва в Заполярье содержат очень мало микроорганизмов, низкие температуры препятствуют сохранению цист патогенных простейших, яиц и личинок гельминтов.
Из перечисленных факторов наибольшее влияние на формирование комплекса признаков арктического адаптивного типа оказали,по-видимому,холодный климат и преимущественно животная пища. Арктическому комплексу признаков свойственны относительно сильное развитиекостно-мускульногокомпонента тела, большие размеры грудной клетки, высокий уровень гемоглобина, относительно большое пространство, занимаемое костным мозгом, повышенное содержание минеральных веществ в костях, высокое содержание в крови белков, холестерина, повышенная способность окислять жиры. Среди аборигенов Арктики почти не встречаются лица с астеническим телосложением.
Вцелом арктический тип характеризуется усиленным энергетическим обменом, который отличается стабильностью показателей в условиях переохлаждения. Имеют свои особенности и механизмы терморегуляции. Так, при одинаковой степени охлаждения у канадских индейцев резко падает температура кожи, но уровень обмена веществ меняется незначительно, а у пришлого белого населения наблюдается меньшая степень снижения кожной температуры, но появляется сильная дрожь, т.е. интенсифицируется обмен.
Втропиках и субтропиках располагается чуть ли не большая часть Ойкумены.
166
Этот регион отличается в целом большим количеством тепла и влаги и сглаженностью сезонных колебаний условий обитания. Вместе с тем благодаря особенностям рельефа наблюдается значительная контрастность распределения тепла и влаги — массивы влажных лесов нередко соседствуют с засушливыми плато, обширными равнинами и редколесьем. В экваториальной и субэкваториальной областях сосредоточены огромные количества растительной биомассы. Влажные тропические леса относительно бедны животными, тогда как в саваннах животный мир разнообразен и включает крупных стадных животных, издавна используемых человеком как объект охоты. Почва, воздух и вода содержат большое количество микроорганизмов, яиц гельминтов и цист патогенных простейших. Богатство и разнообразие животного мира обеспечивает существование огромного количества промежуточных и окончательных хозяев биогельминтов и переносчиков возбудителей трансмиссивных заболеваний. К преобладающим экологическим факторам, под влиянием которых формировался комплекс признаков тропического адаптивного типа, относят жаркий влажный климат и рацион с относительно низким содержанием животного белка.
В тропической области наблюдается исключительно широкая вариабельность групп населения в расовом, этническом и экономическом отношениях. Это проявляется в поразительном размахе изменчивости, например по соматическим признакам. Тем не менее преобладающие экологические факторы, особенно климатический, способствовали образованию определенного комплекса морфофизиологических признаков обитателей тропиков и субтропиков. К характерным признакам тропического типа относят удлиненную форму тела, сниженную мышечную массу, относительное уменьшение массы тела при увеличении длины конечностей, уменьшение окружности грудной клетки, более интенсивное потоотделение за счет повышенного количества потовых желез на 1 см2 кожи, низкие показатели основного обмена и синтеза жиров, сниженную концентрацию холестерина в крови.
Антропологическое изучение современных обитателей зоны умеренного климата под углом зрения формирования биологических механизмов адаптации к природным условиям затруднено, так как значительная часть людей проживает в промышленно развитых странах с большой долей городского населения. Результаты наблюдения позволяют, однако, судить о том, что и в этом случае в процессе исторического развития человеческих популяций сформировался комплекс признаков, соответствующий особому адаптивному типу умеренного пояса.
По соматическим показателям, уровню основного обмена население умеренного пояса занимает промежуточное положение между коренными жителями арктического и тропического регионов. Это соответствует условиям биогеографической среды в зоне умеренного климата. Для нее характерны неравномерное распределение районов, отличающихся по количеству тепла и влаги, типу растительности (от сухих степей и полупустынь до тайги), богатству животного мира. Вместе с тем температура и влажность воздуха здесь не достигают экстремальных величин, хорошо выражен сезонный ритм биоклиматических
167
условий.
Условия высокогорья для человека во многих отношениях экстремальны. Их характеризуют низкое атмосферное давление, сниженное парциальное давление кислорода, холод, относительное однообразие пищи. Основным экологическим фактором формирования горного адаптивного типа явилась,по-видимому,гипоксия. У жителей высокогорья независимо от климатической зоны, расовой и этнической принадлежности наблюдаются повышенный уровень основного обмена, относительное удлинение длинных трубчатых костей скелета, расширение грудной клетки, повышение кислородной емкости крови за счет увеличения количества эритроцитов, содержания гемоглобина и относительной легкости его перехода в оксигемоглобин.
15.4.3. Происхождение адаптивных экологических типов
Человечество возникло в тропической зоне Африки, и следует полагать, что поэтому наиболее древним и исходным для остальных экологических типов человека является тропический тип. Об этом свидетельствует и наибольший полиморфизм морфофункциональных признаков человека в рамках этого типа на территории Центральной иСеверо-ВосточнойАфрики. В связи с этим уместно обратиться к учению Н.И. Вавилова о центрах происхождения и многообразия культурных растений, в соответствии с которым именно в зонах первоначального вовлечения биологических видов в социальную среду наблюдается наиболее выраженный наследственный полиморфизм местных популяций по большому количеству признаков.
Об этом свидетельствует и то, что именно в тропической Африке в непосредственной близости друг от друга, т.е. под действием сходных экологических факторов, обитают наиболее низкорослые и наиболее высокорослые племена, например пигмеи, готтентоты и бушмены, с одной стороны, и массаи — с другой. Пределы изменчивости африканских популяций, например, по росту и массе тела таковы, что в них укладываются все известные на Земле человеческие популяции. Основные же черты тропического типа в Африке, отмеченные выше, остаются неизменными. Они же характеризуют монголоидные популяции Индокитая, Малайского архипелага и некоторые группы индейских племен зоны влажных тропических лесов Центральной и Южной Америки.
Это указывает на то, что тропический адаптивный тип развивается в результате асинхронного параллелизма в эволюции человека, причем вначале на африканском континенте, а потом и в других областях (см. раздел 3.13). Отсюда следует и еще один вывод: адаптивный тип формируется на фоне расогенеза и вне зависимости от него. В процессе адаптогенеза в популяциях человека можно проследить и проявление закона гомологических рядов (см. разд. 13.3.5). Это выражается, например, в том, что в бассейне Меконга в Индокитае и на острове Суматра имеются пигмеоидные популяции, по антропометрическим признакам соответствующие африканским пигмеям.
168
Экологический тип умеренного пояса сформировался на базе исходного генетического и фенотипического полиморфизма тропического типа при расселении популяций человека в умеренных зонах Евразии и позже — Северной Америки. Он оформился в рамках двух больших рас: европеоидной и монголоидной.
При заселении человеком арктической зоны в Евразии и Северной Америке произошло формирование арктического типа. Независимо от него среди индейцев Южной Патагонии и Огненной Земли в приантарктической зоне Южной Америки возникли популяции индейцев, по основному комплексу признаков соответствующие арктическому типу. Это еще один убедительный пример параллелизма эволюции человеческих популяций и реализации закона гомологических рядов, а также доказательство вторичности адаптивных типов по отношению к большим расам человечества.
Формирование горного адаптивного типа иллюстрирует общие закономерности адаптогенеза, отмеченные выше. Этот тип также развился независимо от расовой и этнической принадлежности популяций — среди европеоидов Альп, Кавказа, Памира и Гималаев, а также в монголоидных популяциях Тибета,Тянь-Шаняи Анд. В связи с тем что высокогорья заселялись человеком в последнюю очередь, горный экологический тип является по происхождению самым молодым. Интересно, что, несмотря на особенно выраженную расовую и этническую разнородность этого типа, комплекс основных признаков его является монолитным. Однако в отличие от других типов, вероятно, именно горный проявляется в основном только на фенотипическом уровне и не имеет наследственной природы. Об этом свидетельствует то, что количество эритроцитов в крови и объем грудной клетки людей, переселяющихся в условия высокогорья и обратно, могут меняться на протяжении жизни одного поколения.
Следовательно, адаптивные типы человека не только отражают его прошлое, но формируются и в настоящем, а разные типы имеют и разную по длительности историю.
В настоящее время пока еще на фенотипическом уровне идет формирование адаптивного типа человека городской среды, который характеризуется широкой лабильностью психических реакций, обеспечивающих способность переживать состояния постоянного стресса, и рядом морфофизиологических особенностей, оптимальных для жизни в специфических условиях города (см. разд. 17.3.2).
Формирование экологических типов человека в значительной степени обеспечило всесветное расселение людей. Меняющаяся среда обитания под действием антропогенных факторов ставит перед популяциями человека новые задачи, решаемые и сегодня за счет не только социальных, но и биологических адаптации. Схему эволюционных взаимоотношений больших рас и адаптивных типов человека см. на рис. 15.10.
169

Рис. 15.10. Адаптивные типы человека и большие расы
170
studfiles.net






