Сезонная динамика накопления и расходования луговыми растениями запасных веществ. Основной запас углеводов у растений это
Для чего человеку нужны углеводы? Основные источники образования углеводов в организме. - Школа диабета
Углеводы являются веществами органического происхождения, они широко распространены в окружающей среде. Эти вещества представлены в составе всех клеток и тканей у растений и животных.
Углеводы являются веществами органического происхождения, они широко распространены в окружающей среде. Эти вещества представлены в составе всех клеток и тканей у растений и животных. Углеводы составляют примерно 80% сухого вещества живых организмов растительного происхождения и соответственно около 20% животного происхождения. В природе растения могут синтезировать углеводы из производных неорганического происхождения, таких как углекислый газ и вода.
 Углеводы главный поставщик энергии для организма человека. В организме человека всегда присутствует некоторый запас углеводов, примерно 500г. Здесь углеводы представлены веществом гликогеном. Основная часть этого запаса находится в клетках мышечной ткани, 1/3 часть хранится в печени. Когда мы не едим, гликоген расщепляется до глюкозы, что позволяет избежать выраженных изменений количества сахара в крови. Без поступления углеводов с пищей запасы гликогена расходуются в течение 12-18 часов, в зависимости от активности человека. Тогда организм человека включает механизм синтеза углеводов из промежуточных соединений белкового обмена. Такая изобретательность организма по обеспечению себя углеводами свидетельствует о том, насколько необходимы эти вещества для сохранения жизни организма в целом. Они жизненно необходимы получения энергии всеми тканями. Особенно углеводы важны для клеток мозга, которые получают энергию за счет глюкозы.
Углеводы главный поставщик энергии для организма человека. В организме человека всегда присутствует некоторый запас углеводов, примерно 500г. Здесь углеводы представлены веществом гликогеном. Основная часть этого запаса находится в клетках мышечной ткани, 1/3 часть хранится в печени. Когда мы не едим, гликоген расщепляется до глюкозы, что позволяет избежать выраженных изменений количества сахара в крови. Без поступления углеводов с пищей запасы гликогена расходуются в течение 12-18 часов, в зависимости от активности человека. Тогда организм человека включает механизм синтеза углеводов из промежуточных соединений белкового обмена. Такая изобретательность организма по обеспечению себя углеводами свидетельствует о том, насколько необходимы эти вещества для сохранения жизни организма в целом. Они жизненно необходимы получения энергии всеми тканями. Особенно углеводы важны для клеток мозга, которые получают энергию за счет глюкозы.
По химической структуре углеводы подразделяют на простые сахара, где выделяют моносахариды и дисахариды, а также сложные (полисахариды).
Глюкоза является важнейшим из моносахаридов. Она тот самый кирпичик, который является строительным материалом для синтеза подавляющего большинства дисахаридов и полисахаридов, которые человек получает с пищей. В процессе реакций расщепления и окисления из них образуются всё более простые вещества, вплоть до углекислого газа и воды, с выделением такой необходимой всему живому энергии.
Глюкоза в практически чистом виде содержится в таких овощах и фруктах, как виноград 7,8%, черешня и вишня её содержание около 5,5%, в землянике 2,7%, в сливе 2,5%, в арбузе 2,4%, в тыкве 2,6%, даже в капусте 2,6% и моркови 2,5%.
Фруктоза также моносахарид, она наиболее частый углевод из тех, которые обнаруживаются во фруктах. В отличие от глюкозы, этому веществу не нужна помощь инсулина, чтобы попасть из крови человека в клетки органов и тканей. Именно по этой причине фруктоза более показана для рациона питания диабетиков. Однако следует помнить, что в печени фруктоза превращается в глюкозу, что приводит к некоторому увеличению уровня глюкозы крови. Хотя фруктоза значительно слаще глюкозы, поэтому её можно употреблять в меньших количествах, чем глюкозу.
Фруктозу содержат виноград – 7,7%, яблоки – 5,5%, груши – 5,2%, вишня, черешня – 4,5%, арбузы – 4,3%, черная смородина – 4,2%, малина – 3,9%, земляника – 2,4%, дыня – 2,0%. В овощах её меньше – от 0,1% в свекле до 1,6% в белокочанной капусте. В меде её количество около 3,7%.
Лактоза это дисахарид (галактоза + глюкоза). Именно до таких веществ она расщепляется в желудке под действием лактазы. Лактоза является прекрасным питанием для микрофлоры кишечника. Галактоза в печени преобразуется в глюкозу.
В коровьем молоке лактозы 4,7%, в твороге – от 1,8% до 2,8%, в сметане – от 2,6 до 3,1%, в кефире – от 3,8 до 5,1%, в йогуртах – около 3%.
Сахароза также дисахарид (глюкоза + фруктоза). Из неё на 99,5% состоит сахар, за что его называют носителем пустых калорий. Сахар быстро распадается под действием ферментов желудка до глюкозы и фруктозы, они всасываются в кровь, принося энергию в организм и давая возможность пополнить запас гликогена и жиров. Больше всего сахарозы в свекле – 8,6%, персиках – 6,0%, дынях – 5,9%, сливах – 4,8%, мандаринах – 4,5%.
 Крахмал является перевариваемым полисахаридом. Им представлены около 80% всех углеводов, которые человек употребляет с пищей. Крахмал в наибольшем количестве содержится в крупах ( 60% в гречневой, 70% в рисовой крупе). Из злаков крахмала меньше в овсяной крупе - 49%. Макароны содержат 62-68% крахмала, хлеб из ржаной муки 33%-49%, изделия из пшеничной муки 35%-51% крахмала, мука – от 56 ( ржаная ) и 68% ( пшеничная высшего сорта ). В бобовых культурах крахмала 40% в чечевице, 44% в горохе. За высокое содержание крахмала (15-18%) картофель относится диетологами не к овощам, а к крахмалистым продуктам, таким же как злаковые и бобовые.
Крахмал является перевариваемым полисахаридом. Им представлены около 80% всех углеводов, которые человек употребляет с пищей. Крахмал в наибольшем количестве содержится в крупах ( 60% в гречневой, 70% в рисовой крупе). Из злаков крахмала меньше в овсяной крупе - 49%. Макароны содержат 62-68% крахмала, хлеб из ржаной муки 33%-49%, изделия из пшеничной муки 35%-51% крахмала, мука – от 56 ( ржаная ) и 68% ( пшеничная высшего сорта ). В бобовых культурах крахмала 40% в чечевице, 44% в горохе. За высокое содержание крахмала (15-18%) картофель относится диетологами не к овощам, а к крахмалистым продуктам, таким же как злаковые и бобовые.
Организм человека не может делать большие запасы глюкозы, Потому он нуждается в регулярном поступлении углеводов. Однако гораздо полезнее не чистая глюкоза, а полисахариды, например, крахмал. Кроме того, все продукты, которые являются источником крахмала, богаты витаминами, клетчаткой, микроэлементами и многими другими веществами, столь необходимыми организму человека. Полисахаридами должна быть представлена большая доля всех поступающих организм углеводов.
diabet.by
Углеводы
Углеводы— первичные продукты фотосинтеза и основные исходные продукты биосинтеза других веществ в растениях. Составляют существенную часть пищевого рациона. Обмен углеводов — совокупность процессов превращений углеводов в организме. Углеводы, поступающие в организм с пищевыми продуктами, представлены, главным образом, крахмалом и тростниковым сахаром. Крахмал — полисахарид растений, он состоит из цепочки соединенных между собой более простых молекул — моносахаров, главным из которых является глюкоза. По своей структуре крахмал подобен гликогену. Тростниковый сахар — это углевод, который преобладает в нашем рационе. По структуре это дисахарид, т.е. он состоит из двух молекул моносахаров — глюкозы и фруктозы. Глюкоза и фруктоза могут находиться в разных пищевых продуктах и в свободном виде, например, в меде и фруктах. В молочных продуктах содержится, в основном, такой углевод, как лактоза. В организме имеется «депо» углеводов — гликоген, образованный из молекул глюкозы.
Углеводы, прежде всего, источник энергии, в меньшей степени они выполняют
пластическую функцию. Организм человека не нуждается в определенных углеводах. Единственным «незаменимым» производным углеводов, которое обязательно должно поступать с пищей, является аскорбиновая кислота или витамин С, так как у человека отсутствует один из ферментов, необходимых для его синтеза. В сбалансированной диете примерно 50% необходимой человеку энергии должно поступать с углеводами Превращения углеводов связанные с дыхание и брожением
Процесс превращения углеводов начинается с переваривания их в ротовой полостипод влиянием слюны, затем некоторое время продолжается вжелудкеи заканчивается втонком кишечнике— основном месте гидролиза углеводов под влиянием ферментов, содержащихся в пищеварительном сокеподжелудочной железыи тонкого кишечника. Продукты гидролиза — моносахара — всасываются в кишечнике и поступают в кровь воротной вены, по которой моносахариды пищи поступают впечень, где они превращаются в глюкозу. Глюкоза далее поступает в кровь и может вступить в процессы, протекающие в клетках или переходит в гликоген печени.
Роль печени в углеводном обмене
Печень — главный орган, в клетках которого происходят биохимические превращения продуктов пищеварительного гидролиза углеводов и превращение их в глюкозу — форму, доступную для клеток организма. Печень — депо углеводов, так как часть глюкозы хранится здесь в виде гликогена. Печень поддерживает содержание глюкозы в крови на постоянном уровне — в этом состоит глюкостатическая функция печени. При избытке глюкозы в печени происходит синтез гликогена из глюкозы — гликогенез. После приема пищи богатой углеводами содержание гликогена может составлять до 8% веса печени. В среднем, запасы гликогена составляют около 5% веса печени, что у взрослого человека эквивалентно примерно 90 г глюкозы.
При повышении потребности организма в глюкозе происходит распад гликогена печени — гликогенолиз, который достаточен для удовлетворения нужд организма в первые 12-24 часа после приема пищи. Печень — один из главных органов, где происходит процесс ферментативного синтеза глюкозы из углеводных и неуглеводных продуктов — глюконеогенез. Причем клетки печени способны реагировать на возникновение потребности в глюкозе и в клетках других органов. При голодании, после истощения запасов гликогена, процессы глюконеогенеза идут с максимальной интенсивностью, поддерживая «сахар» крови на постоянном уровне. В печени также происходит гликолиз — ферментативный распад глюкозы с освобождением энергии, заключенной в ее молекуле и переводом ее в форму, доступную для организма — т.е. в аденозинтрифосфат (АТФ).
Превращение глюкозы в клетках
В клетках глюкоза может расщепляться как анаэробно (без участия кислорода), так и аэробно (с участием кислорода). В анаэробных условиях гликолиза из каждой молекулы расщепившейся глюкозы образуются 2 молекулы аденозинтрифосфата (АТФ) и 2 молекулы молочной кислоты. При аэробном гликолизе промежуточные продукты углеводного обмена, образующиеся в процессе анаэробного распада углеводов (пировиноградная кислота), не восстанавливаются до молочной кислоты, а окисляются в митохондриях в цикле трикарбоновых кислот до углекислого газа и воды с накоплением энергии в виде АТФ. Кроме того, промежуточные продукты гликолиза являются материалом для синтеза многих важных соединений и используются организмом как еще один источник материала для процессов ассимиляции.
Регуляция содержания глюкозы в крови
О состоянии обмена углеводов можно судить по содержанию сахара в крови. У здорового человека в крови поддерживается постоянная концентрация глюкозы 70-120 мг%. После приема пищи, содержащей углеводы, концентрация глюкозы в крови возрастает примерно до 150мг % и остается на этом уровне около 2 часов, а затем возвращается к норме. Содержание глюкозы в крови — одна из самых важных констант жидкой внутренней среды организма. Ведущая роль в поддержании этой константы на постоянном уровне благодаря идущим там процессам гликогенеза и гликогенолиза принадлежит печени. Длительное повышение содержания глюкозы в крови — гипергликемия стимулирует выделение в кровь инсулина. Инсулин снижаетсодержание глюкозы в крови после возрастания ее концентрации (гипергликемии).
У здорового человека в период между приемами пищи нормальное содержание глюкозы в крови поддерживается путем распада гликогена в печени с образованием свободной глюкозы — процессом гликогенолиза. При снижении сахара крови — гипогликемии, длящейся более длительное время, в кровь поступает глюкагон — гормон, выделяемыйподжелудочной железой. Инсулин, гормон поджелудочной железы, стимулирует процессы синтеза гликогена в печени — гликогенез, поглощение глюкозы клетками других тканей организма, подавляет образование глюкозы, т.е. процессы глюконеогенеза. Инсулин — главный гормон. Этот гормон обладает специфическим действием: он действует исключительно на процессы гликогенолиза, ускоряя образование глюкозы.
При голодании, длящемся более 24 часов, запасы гликогена в печени истощаются. В прессы регуляции включаются гормоны коры надпочечника— глюкокортикоиды. Глюкокортикоиды, во-первых, усиливают глюконеогенез в печени; во-вторых, обеспечивает процессы глюконеогенеза субстратом, усиливая распад белков в тканях организма, они предоставляют для глюконеогенеза углеродсодержащий субстрат. К гормонам, которые обеспечивают повышение сахара крови, относятся адреналин и гормон роста.
Адреналин — гормон мозгового вещества надпочечника. Он усиливает процессы перехода гликогена в глюкозу. Гормон роста, во-первых, подавляет использование глюкозы клетками тканей; во-вторых, при резком и длительном снижении сахара крови стимулирует распад жиров и образование из них углеводов. Дыхание. Все живые организмы дышат, т. е. поглощают кислород и выделяют углекислый газ и воду. При этом происходит разложение органических веществ и выделение энергии, необходимой для жизни каждой клетки, всего растения. В действительности этот процесс многоступенчатый. Он состоит из целого ряда последовательно идущих окислительно-восста-новительных реакций. В качестве органических веществ, необходимых для дыхания, служат в основном углеводы, белки и жиры. Типичным соединением, окисляемым в процессе дыхания, является глюкоза. Энергетически наиболее выгодным для дыхания веществом является жир. 1 г жира при окислении до СО2 и Н2О дает 9,2 ккал, белки — 5,7 ккал, углеводы — 4 ккал. Процесс превращения исходного органического вещества до более простых и затем до СО2 и Н2О требует большого числа различных ферментов.
В процессе фотосинтеза растения синтезируют углеводы, которые транспортируются из листьев в другие органы. На свету и в темноте клетки растения «дышат», окисляя углеводы молекулярным кислородом с образованием СО2 и воды. При этом освобождается большое количество свободной энергии: С6Н12О6 + 6О2 = 6СО2 + 6Н2О + энергия; ∆G = -2882 кДж/моль (-686 ккал/моль) Эта формула в общем виде отражает чрезвычайно сложный, а главное, контролируемый процесс, который условно можно разбить на три этапа: гликолиз, цикл трикарбоновых кислот и окислительное фосфорилирование в дыхательной цепи (рис. 1).
= -2882 кДж/моль (-686 ккал/моль) Эта формула в общем виде отражает чрезвычайно сложный, а главное, контролируемый процесс, который условно можно разбить на три этапа: гликолиз, цикл трикарбоновых кислот и окислительное фосфорилирование в дыхательной цепи (рис. 1).
Гликолиз и цикл трикарбоновых кислот — это биохимические пути окисления глюкозы, протекающие соответственно в цитозоле и матриксе митохондрий. В биохимических реакциях синтезируется небольшое количество АТФ, и главный их результат — образование соединений с высоким восстановительным потенциалом — НАДН и ФАДН2. На заключительном этапе восстановительные эквиваленты окисляются в электрон-транспортной цепи, локализованной во внутренней мембране митохондрий. Перенос электрона в цепи завершается восстановлением кислорода до воды. В процессе электронного транспорта на мембране образуется электрохимический протонный градиент ΔµἨ, энергия которого используется для синтеза АТФ из АДФ и Фн. Процесс, в котором работа дыхательной цепи сопряжена с синтезом АТФ, получил название окислительного фосфорилирования. Именно в этом процессе синтезируется основная масса АТФ, образуемого при дыхании. И у растений, и у животных дыхание выполняет три основные функции. Во-первых, освобождаемая при окислении углеводов энергия преобразуется в конвертируемые формы клеточной энергии — ΔµἨ и АТФ. Вторая, не менее важная функция — снабжение клетки метаболитами, которые образуются в ходе окисления глюкозы и используются в разнообразных биосинтезах. Третья функция связана с термогенезом, т. е. рассеиванием энергии в виде тепла. Процесс дыхания принципиально сходен у животных и растений, но у последних имеет свои особенности. Все вместе они отражают пластичность растительного метаболизма и связаны с функционированием, наряду с основными, альтернативных ферментов и реакций. Наличие альтернативных путей расширяет адаптивные возможности растений, но усложняет (с точки зрения исследователя) систему регуляции метаболических процессов.
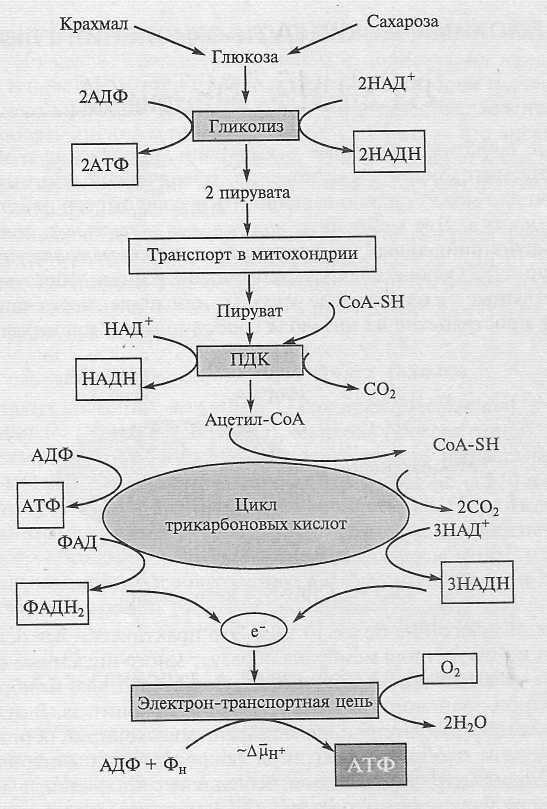
Рис. 1. Основные этапы дыхания
Окисление глюкозы в процессе гликолиза сопровождается восстановлением двух молекул НАД+, синтезом двух молекул АТФ и завершается образованием двух молекул пирувата. В митохондриях пируват подвергается полному окислению до СО2 в реакциях, катализируемых пируватдегидрогеназным комплексом (ПДК) и ферментами цикла трикарбоновых кислот (ЦТК). В этих процессах образуются 4НАДН, 1ФАДН2, а также одна молекула АТФ. Восстановительные эквиваленты окисляются, отдавая электроны в электрон-транспортную цепь, локализованную во внутренней митохондриальной мембране. Электронный транспорт приводит к восстановлению кислорода до воды и сопряжен с синтезом основной массы АТФ в процессе окислительного фосфорилирования.
Основные события, связанные с дыханием, происходят в митохондриях. Растительные митохондрии, как правило, сферической или цилиндрической формы, их число может сильно варьировать в зависимости от метаболической активности клетки. Две мембраны, наружная и внутренняя, делят митохондрию на два функциональных компартмента — межмембранное пространство и матрикс (рис. 2). 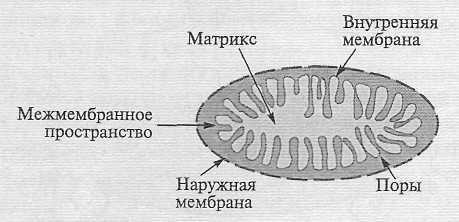 Рис. 2. Структура митохондрийОсобые белки, называемые поринами, образуют в наружной мембране крупные гидрофильные каналы, или поры, через которые в межмембранное пространство из цитозоля свободно могут проникать соединения с молекулярной массой не более 10 кДа. Это практически все основные метаболиты клетки. Внутренняя мембрана образует многочисленные складки, кри-сты, которые увеличивают ее поверхность. Во внутреннюю мембрану интегрированы электрон-транспортная цепь (ЭТЦ) и АТФ-синтаза. В отличие от других клеточных мембран внутренняя мембрана митохондрий обогащена белком (75 %) и содержит особый фосфолипид (дифосфатидилглицерол) — кардиолипин. Она пропускает газы, воду и небольшие липофильные молекулы, но непроницаема для заряженных молекул и ионов, что является обязательным условием ее функционирования как сопрягающей мембраны. Однако в мембране есть белки — транспортеры, с помощью которых возможен обмен метаболитами между матриксом и цитозолем . Матрикс, т. е. окруженное внутренней мембраной пространство, содержит ферменты цикла трикарбоновых кислот.
Рис. 2. Структура митохондрийОсобые белки, называемые поринами, образуют в наружной мембране крупные гидрофильные каналы, или поры, через которые в межмембранное пространство из цитозоля свободно могут проникать соединения с молекулярной массой не более 10 кДа. Это практически все основные метаболиты клетки. Внутренняя мембрана образует многочисленные складки, кри-сты, которые увеличивают ее поверхность. Во внутреннюю мембрану интегрированы электрон-транспортная цепь (ЭТЦ) и АТФ-синтаза. В отличие от других клеточных мембран внутренняя мембрана митохондрий обогащена белком (75 %) и содержит особый фосфолипид (дифосфатидилглицерол) — кардиолипин. Она пропускает газы, воду и небольшие липофильные молекулы, но непроницаема для заряженных молекул и ионов, что является обязательным условием ее функционирования как сопрягающей мембраны. Однако в мембране есть белки — транспортеры, с помощью которых возможен обмен метаболитами между матриксом и цитозолем . Матрикс, т. е. окруженное внутренней мембраной пространство, содержит ферменты цикла трикарбоновых кислот.
ГЛЮКОЗА — ОСНОВНОЙ СУБСТРАТ ДЫХАНИЯ У РАСТЕНИЙ Основным субстратом дыхания у растений являются глюкоза и ее производные, хотя в особых случаях дыхание могут поддерживать белки и жиры, запасенные в семенах. Глюкоза образуется в клетках растений при гидролизе крахмала и сахарозы — продуктов фотосинтеза. Крахмал представляет собой смесь двух полисахаридов — амилозы и амилопектина. Молекулы амилозы — это длинные, неразветвленные цепи α-D-глюкопиранозных остатков, соединенных гликозидными α(1→4)-связями. Молекулы амилопектина также представлены цепями α -D-глюкопиранозных остатков, которые в точке ветвления образуют а(1→6)-связь. Крахмал как запасный полисахарид накапливается в хлоропластах и пластидах гетеротрофных тканей. Некоторые растения — топинамбур (Heliantus tuberosus), георгин (Dahlia sp.) в качестве запасных углеводов могут использовать инулин и гемицеллюлозы. Сахароза — это дисахарид, образованный остатками глюкозы и фруктозы. Она синтезируется в цитозоле, из фотосинтезирующих клеток по апопласту листа и сосудам флоэмы транспортируется в другие органы растения. Крахмал расщепляется до моносахаридов при участии ряда ферментов (α- и β-амилазы, α-1,6-глюкозидазы, крахмалфосфорилазы и др.) с образованием D-глюкозы или D-глюкозо-1-фосфата. Распад сахарозы может идти при обращении реакций ее синтеза, но в основном происходит в результате гидролиза при участии фермента инвертазы: сахароза + Н2О → фруктоза + глюкоза
В геноме таких растений, как томат (Lycopersicon esculentum), кукуруза (Zea mays), арабидопсис (Arabidopsis thaliana), морковь (Dancus carota), обнаружено целое семейство ядерных генов, кодирующих разные изоформы инвертазы. Например, у моркови кислые инвертазы (оптимум рН 4,5 — 5,0) в пяти разных изоформах присутствуют в вакуоли и клеточной стенке. В цитозоле есть нейтральная инвертаза (оптимум рН 7,0—8,0), которая также может иметь несколько изоформ. Таким образом, у растений гидролиз сахарозы может идти в разных клеточных компартментах и контролируется сложным образом через активность инвертаз, обладающих разными свойствами.
Брожение- процесс анаэробного расщепления органических веществ, преимущественно углеводов, происходящий под влиянием микроорганизмов или выделенных из них ферментов. В ходе брожение в результате сопряженных окислительно-восстановительных реакций освобождается энергия, необходимая для жизнедеятельности микроорганизмов, и образуются химическиесоединения, которые микроорганизмы используют для биосинтеза аминокислот,белков,органических кислот, жиров и др. компонентов тела. Одновременно накапливаются конечные продуктыброжение. В зависимости от их характера различают брожение спиртовое, молочнокислое, маслянокислое, пропионовокислое, ацетоно-бутиловое, ацетоно-этиловое и др. виды. Характер брожение, его интенсивность, количественные соотношения конечных продуктов, а также направление брожение зависят от особенностей его возбудителя и условий, при которых брожение протекает (pH, аэрация, субстрат и др.).
Спиртовое Брожение. В 1836 французский учёный Каньяр де ла Тур установил, что спиртовое брожение связано с ростом и размножением дрожжей. Химическое уравнение спиртового брожение: C6h22O6 ® 2C2H5OH + 2CO2 было дано французскими химиками А. Лавуазье (1789) и Ж. Гей-Люссаком (1815). Л. Пастер пришёл к выводу (1857), что спиртовое брожение могут вызывать только живые дрожжи в анаэробных условиях («брожение — это жизнь без воздуха»). В противовес этому немецкий учёный Ю. Либих упорно настаивал на том, что брожение происходит вне живой клетки. На возможность бесклеточного спиртового брожение впервые (1871) указала русский врач-биохимик М. М. Манассеина.
Немецкий химик Э. Бухнер в 1897, отжав под большим давлением дрожжи, растёртые с кварцевым песком, получил бесклеточный сок, сбраживающий сахарс образованием спирта иCO2. При нагревании до 50°Cи выше сок утрачивал бродильные свойства. Всё это указывало на ферментативную природу активного начала, содержащегося в дрожжевом соке. Русский химик Л. А. Иванов обнаружил (1905), что добавленные к дрожжевому соку фосфаты в несколько раз повышают скоростьброжение.
Исследования отечественных биохимиков А. И. Лебедева, С. П. Костычева, Я. О. Парнаса и немецких биохимиков К. Нейберга, Г. Эмбдена, О. Мейергофа и др. подтвердили, что фосфорнаякислота участвует в важнейших этапах спиртовогоброжение 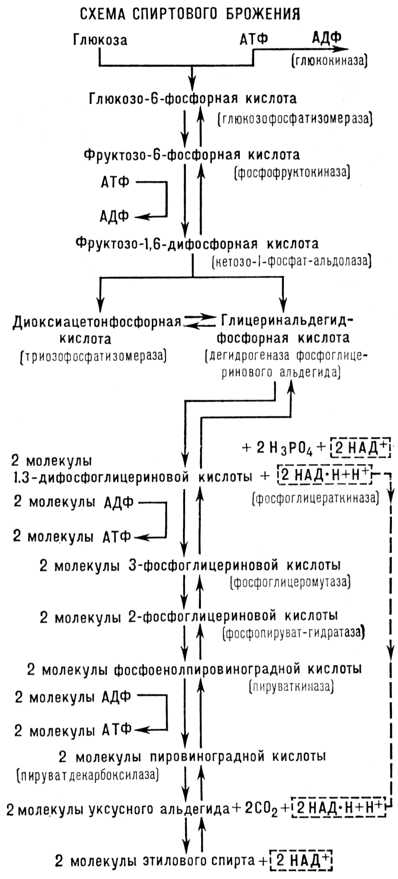 В дальнейшем многие исследователи детально изучили ферментативную природу и механизм спиртового брожение (см. схему). Первая реакция превращения глюкозыпри спиртовомброжение — присоединение к глюкозепод влиянием фермента глюкокиназы остаткафосфорнойкислоты отаденозинтрифосфорнойкислоты (АТФ, см.Аденозинфосфорные кислоты). При этом образуются аденозиндифосфорнаякислота (АДФ) иглюкозо-6-фосфорнаякислотата. Последняя под действием ферментаглюкозофосфати-зомеразыпревращается в фруктозо-6-фосфорнуюкислоту, которая, получая от новой молекулы АТФ (при участии фермента фосфофруктокиназы) ещё один остатокфосфорнойкислоты, превращается в фруктозо-1,6-дифосфорную кислоту. (Эта и следующая реакции, обозначенные встречными стрелками, обратимы, т. е. их направление зависит от условий — концентрации фермента, pH и др.) Под влиянием фермента кетозо-1-фосфатальдолазы фруктозо-1,6-дифосфорная кислота расщепляется на глицеринальдегидфосфорную и диоксиацетонфосфорную кислоты которые могут превращаться друг в друга под действием фермента триозофосфатизомеразы. Глицеринальдегидфосфорная кислота, присоединяя молекулу неорганическойфосфорнойкислоты и окисляясь под действием ферментадегидрогеназыфосфоглицеринальдегида, активной группой которого у дрожжей является никотинамидадениндинуклеотид (НАД), превращается в 1,3-дифосфоглицериновую кислоту. Молекула диоксиацетонфосфорной кислоты под действием триозофосфатизомеразы даёт вторую молекулу глицеринальдегидфосфорной кислоты, также подвергающуюся окислению до 1,3-дифосфоглицериновой кислоты; последняя, отдавая АДФ (под действием фермента фосфоглицераткиназы) один остатокфосфорнойкислоты, превращается в З-фосфоглицериновую кислоту, которая под действием фермента фосфоглицеро-мутазы превращается в 2-фосфоглицериновую кислоту, а она под влиянием фермента фосфопируват-гидратазы — в фосфоенол-пировиноградную кислоту. Последняя при участии фермента пируваткиназы передаёт остатокфосфорнойкислоты молекуле АДФ, в результате чего образуется молекула АТФ и молекула енолпировиноградной кислоты, которая весьма нестойка и переходит в пировиноградную кислоту. Эта кислота при участии имеющегося в дрожжах фермента пируватдекарбоксилазы расщепляется на уксусный альдегид и двуокисьуглерода.Уксусный альдегид, реагируя с образовавшейся при окислении глицеринальдегидфосфорной кислоты восстановленной формой никотинамидадениндинуклеотида (НАД-Н), при участии фермента алкогольдегидрогеназы превращается в этиловый спирт. Суммарно уравнение спиртовогоброжение может быть представлено в следующем виде: C6h22O6 + 2h4PO4 + 2АДФ ® 2Ch4Ch3OH + 2CO2 + 2АТФ. Т. о., при сбраживании 1 моля глюкозыобразуются 2моля этилового спирта, 2 моля CO2, а также в результате фосфорилирования2молей АДФ образуются 2 моля АТФ. Термодинамические расчёты показывают, что при спиртовом брожение превращение 1 моля глюкозыможет сопровождаться уменьшением свободной энергии примерно на 210кдж (50 000 кал), т. е. энергия, аккумулированная в 1 моле этилового спирта, на 210 кдж (50 000 кал) меньше энергии 1 моля глюкозы.При образовании 1моля АТФ (макроэргических — богатых энергией фосфатных соединений) используется 42 кдж (10 000 кал). Следовательно, значительная часть энергии, освобождающейся при спиртовом брожение, запасается в виде АТФ, обеспечивающей разнообразные энергетические потребности дрожжевых клеток. Такое же биологическое значение имеет процесс брожение и у др. микроорганизмов. При полном сгорании 1 моля глюкозы(с образованиемCO2 и h3O) изменение свободной энергии достигает 2,87 Мдж (686 000 кал). Иначе говоря, дрожжевая клетка использует лишь 7% энергии глюкозы.Это показывает малую эффективность анаэробных процессов по сравнению с процессами, идущими в присутствии кислорода. При наличии кислорода спиртовоеброжение угнетается или прекращается и дрожжи получают энергию для жизнедеятельности в процессе дыхания. Наблюдается тесная связь между брожением и дыханием микроорганизмов, растений и животных. Ферменты, участвующие в спиртовом брожение, имеются также в тканях животных и растений. Во многих случаях первые этапы расщепления сахаров,вплоть до образования пировиноградной кислоты, — общие дляброжение и дыхания. Большее значение процесс анаэробного распада глюкозыимеет и при сокращении мышц , первые этапы этого процесса также сходны с начальными реакциями спиртовогоброжение. Сбраживание углеводов (глюкозы,ферментативных гидролизатов крахмала, кислотных гидролизатов древесины) используется во многих отраслях промышленности: для получения этилового спирта, глицерина и др. технических и пищевых продуктов. На спиртовомБрожение основаны приготовление теста в хлебопекарной промышленности, виноделие и пивоварение. Молочнокислое Брожение. Молочнокислые бактерии подразделяют на 2 группы — гомоферментативные и гетероферментативные. Гомоферментативные бактерии (например, Lactobacillus delbrückii) расщепляют моносахариды с образованием двух молекул молочной кислоты в соответствии с суммарным уравнением: C6h22O6 = 2Ch4CHOH·COOH. Гетероферментативные бактерии (например, Bacterium lactis aerogenes) ведут сбраживание с образованием молочной кислоты, уксусной кислоты, этилового спирта и CO2, а также образуют небольшое количество ароматических. веществ — диацетила, эфиров и т.д. При молочнокислом брожение превращение углеводов, особенно на первых этапах, близко к реакциям спиртового брожение, за исключением декарбоксилирования пировиноградной кислоты, которая восстанавливается до молочной кислоты за счёт водорода, получаемого от НАД-Н. Гомоферментативное молочнокислое брожение используется для получения молочной кислоты, при изготовлении различных кислых молочных продуктов, хлеба и в силосовании кормов в сельском хозяйстве. Гетероферментативное молочнокислое брожение происходит при консервировании различных плодов и овощей путём квашения. Маслянокислое Брожение. Сбраживание углеводов с преимущественным образованием масляной кислоты производят многие анаэробные бактерии, относящиеся к роду Clostridium. Первые этапы расщепления углеводов при маслянокислом Брожение аналогичны соответстветственным этапам спиртового брожение, вплоть до образования пировиноградной кислоты, из которой при маслянокислом брожение образуется ацетил-кофермент A (Ch4CO-KoA). Ацетил-KoA может служить предшественником масляной кислоты, подвергаясь следующим превращениям: Маслянокислое Брожение применялось для получения масляной кислоты из крахмала. Ацетоно-бутиловое брожение бактерии Clostridium acetobutylicum сбраживают углеводы с преим. образованием бутилового спирта (Ch4Ch3Ch3Ch3OH) и ацетона (Ch4COCh4). При этом образуются также в сравнительно небольших количествах водород, CO2, уксусная, масляная кислоты, этиловый спирт. Первые этапы расщепления углеводов те же, что и при спиртовом брожении. Бутиловый спирт образуется путём восстановления масляной кислоты: Ch4Ch3Ch3COOH + 4H =Ch4Ch3Ch3Ch3OH + h3O. Ацетон же образуется декарбоксилированием ацетоуксусной кислоты, которая получается в результате конденсации двух молекул уксусной кислоты. Исследованиями В. Н. Шапошникова показано, что ацетоно-бутиловое брожение (как и ряд др., например пропионовокислое, маслянокислое) в опытах с растущей культурой происходит в две фазы. В первую фазу брожение параллельно с нарастанием биомассы накапливаются уксусная и масляная кислоты; во вторую фазу образуются преимущественно ацетон и бутиловый спирт. При ацетоно-бутиловом брожение сбраживаются моносахариды, дисахариды и полисахариды — крахмал, инсулин, но не сбраживаются клетчатка и гемицеллюлоза. Ацетоно-бутиловое брожение использовалось для промышленного получения бутилового спирта и ацетона, применяемых в химической и лакокрасочной промышленности. Сбраживание белков. Некоторые бактерии из рода Clostridium — гнилостные анаэробы— способны сбраживать не только углеводы, но и аминокислоты. Эти бактерии более приспособлены к использованию белков,расщепляемых ими при помощи протеолитических ферментов до аминокислот, которые затем подвергаютсяброжение. Процесс сбраживания белковимеет значение в круговороте веществ в природе. Пропионовокислое Брожение. Основные продукты пропионовокислого брожение, вызываемого несколькими видами бактерий из рода Propionibacterium, — пропионовая (Ch4Ch3OH) и уксусная кислоты и CO2. Химизм пропионовокислого брожение сильно изменяется в зависимости от условий. Это, по-видимому, объясняется способностью пропионовых бактерий перестраивать обмен веществ, например в зависимости от аэрации. При доступе кислорода они ведут окислительный процесс, а в его отсутствии расщепляют гексозы путём брожение. Пропионовые бактерии способны фиксировать CO2, при этом из пировиноградной к-ты и CO2 образуется щавелевоуксусная к-та, превращающаяся в янтарную к-ту, из которой декарбоксилированием образуется пропионовая к-та: Существуют брожение, которые сопровождаются и восстановительными процессами. Примером такого «окислительного» брожение служит лимоннокислое брожение. Многие плесневые грибы сбраживают сахарас образованием лимонной кислоты. Наиболее активные штаммы Aspergillus niger превращают до 90% потребленногосахарав лимонную кислоту. Значительная часть лимонной кислоты, используемой в пищевой промышленности, производится микробиологическим путём — глубинным и поверхностным культивированием плесневых грибов. Иногда по традиции и чисто окислительные процессы, осуществляемые микроорганизмами, называется брожение. Примерами таких процессов могут служить уксуснокислое и глюконовокислое брожение. Уксуснокислое Брожение. Бактерии, относящиеся к роду Acetobacter, окисляют этиловый спирт в уксусную кислоту в соответствии с суммарной реакцией: Промежуточное соединение при окислении спирта в уксусную кислоту — уксусный альдегид. Многие уксуснокислые бактерии, кроме окисления спирта в уксусную кислоту, осуществляют окисление глюкозыв глюконовую и кетоглюконовую кислоты. Глюконовокислое Брожение осуществляют и некоторые плесневые грибы, способные окислять альдегидную группу глюкозы,превращая последнюю в глюконовую кислоту: Кальциеваясоль глюконовой кислоты служит хорошим источником кальция для людей и животных.
В дальнейшем многие исследователи детально изучили ферментативную природу и механизм спиртового брожение (см. схему). Первая реакция превращения глюкозыпри спиртовомброжение — присоединение к глюкозепод влиянием фермента глюкокиназы остаткафосфорнойкислоты отаденозинтрифосфорнойкислоты (АТФ, см.Аденозинфосфорные кислоты). При этом образуются аденозиндифосфорнаякислота (АДФ) иглюкозо-6-фосфорнаякислотата. Последняя под действием ферментаглюкозофосфати-зомеразыпревращается в фруктозо-6-фосфорнуюкислоту, которая, получая от новой молекулы АТФ (при участии фермента фосфофруктокиназы) ещё один остатокфосфорнойкислоты, превращается в фруктозо-1,6-дифосфорную кислоту. (Эта и следующая реакции, обозначенные встречными стрелками, обратимы, т. е. их направление зависит от условий — концентрации фермента, pH и др.) Под влиянием фермента кетозо-1-фосфатальдолазы фруктозо-1,6-дифосфорная кислота расщепляется на глицеринальдегидфосфорную и диоксиацетонфосфорную кислоты которые могут превращаться друг в друга под действием фермента триозофосфатизомеразы. Глицеринальдегидфосфорная кислота, присоединяя молекулу неорганическойфосфорнойкислоты и окисляясь под действием ферментадегидрогеназыфосфоглицеринальдегида, активной группой которого у дрожжей является никотинамидадениндинуклеотид (НАД), превращается в 1,3-дифосфоглицериновую кислоту. Молекула диоксиацетонфосфорной кислоты под действием триозофосфатизомеразы даёт вторую молекулу глицеринальдегидфосфорной кислоты, также подвергающуюся окислению до 1,3-дифосфоглицериновой кислоты; последняя, отдавая АДФ (под действием фермента фосфоглицераткиназы) один остатокфосфорнойкислоты, превращается в З-фосфоглицериновую кислоту, которая под действием фермента фосфоглицеро-мутазы превращается в 2-фосфоглицериновую кислоту, а она под влиянием фермента фосфопируват-гидратазы — в фосфоенол-пировиноградную кислоту. Последняя при участии фермента пируваткиназы передаёт остатокфосфорнойкислоты молекуле АДФ, в результате чего образуется молекула АТФ и молекула енолпировиноградной кислоты, которая весьма нестойка и переходит в пировиноградную кислоту. Эта кислота при участии имеющегося в дрожжах фермента пируватдекарбоксилазы расщепляется на уксусный альдегид и двуокисьуглерода.Уксусный альдегид, реагируя с образовавшейся при окислении глицеринальдегидфосфорной кислоты восстановленной формой никотинамидадениндинуклеотида (НАД-Н), при участии фермента алкогольдегидрогеназы превращается в этиловый спирт. Суммарно уравнение спиртовогоброжение может быть представлено в следующем виде: C6h22O6 + 2h4PO4 + 2АДФ ® 2Ch4Ch3OH + 2CO2 + 2АТФ. Т. о., при сбраживании 1 моля глюкозыобразуются 2моля этилового спирта, 2 моля CO2, а также в результате фосфорилирования2молей АДФ образуются 2 моля АТФ. Термодинамические расчёты показывают, что при спиртовом брожение превращение 1 моля глюкозыможет сопровождаться уменьшением свободной энергии примерно на 210кдж (50 000 кал), т. е. энергия, аккумулированная в 1 моле этилового спирта, на 210 кдж (50 000 кал) меньше энергии 1 моля глюкозы.При образовании 1моля АТФ (макроэргических — богатых энергией фосфатных соединений) используется 42 кдж (10 000 кал). Следовательно, значительная часть энергии, освобождающейся при спиртовом брожение, запасается в виде АТФ, обеспечивающей разнообразные энергетические потребности дрожжевых клеток. Такое же биологическое значение имеет процесс брожение и у др. микроорганизмов. При полном сгорании 1 моля глюкозы(с образованиемCO2 и h3O) изменение свободной энергии достигает 2,87 Мдж (686 000 кал). Иначе говоря, дрожжевая клетка использует лишь 7% энергии глюкозы.Это показывает малую эффективность анаэробных процессов по сравнению с процессами, идущими в присутствии кислорода. При наличии кислорода спиртовоеброжение угнетается или прекращается и дрожжи получают энергию для жизнедеятельности в процессе дыхания. Наблюдается тесная связь между брожением и дыханием микроорганизмов, растений и животных. Ферменты, участвующие в спиртовом брожение, имеются также в тканях животных и растений. Во многих случаях первые этапы расщепления сахаров,вплоть до образования пировиноградной кислоты, — общие дляброжение и дыхания. Большее значение процесс анаэробного распада глюкозыимеет и при сокращении мышц , первые этапы этого процесса также сходны с начальными реакциями спиртовогоброжение. Сбраживание углеводов (глюкозы,ферментативных гидролизатов крахмала, кислотных гидролизатов древесины) используется во многих отраслях промышленности: для получения этилового спирта, глицерина и др. технических и пищевых продуктов. На спиртовомБрожение основаны приготовление теста в хлебопекарной промышленности, виноделие и пивоварение. Молочнокислое Брожение. Молочнокислые бактерии подразделяют на 2 группы — гомоферментативные и гетероферментативные. Гомоферментативные бактерии (например, Lactobacillus delbrückii) расщепляют моносахариды с образованием двух молекул молочной кислоты в соответствии с суммарным уравнением: C6h22O6 = 2Ch4CHOH·COOH. Гетероферментативные бактерии (например, Bacterium lactis aerogenes) ведут сбраживание с образованием молочной кислоты, уксусной кислоты, этилового спирта и CO2, а также образуют небольшое количество ароматических. веществ — диацетила, эфиров и т.д. При молочнокислом брожение превращение углеводов, особенно на первых этапах, близко к реакциям спиртового брожение, за исключением декарбоксилирования пировиноградной кислоты, которая восстанавливается до молочной кислоты за счёт водорода, получаемого от НАД-Н. Гомоферментативное молочнокислое брожение используется для получения молочной кислоты, при изготовлении различных кислых молочных продуктов, хлеба и в силосовании кормов в сельском хозяйстве. Гетероферментативное молочнокислое брожение происходит при консервировании различных плодов и овощей путём квашения. Маслянокислое Брожение. Сбраживание углеводов с преимущественным образованием масляной кислоты производят многие анаэробные бактерии, относящиеся к роду Clostridium. Первые этапы расщепления углеводов при маслянокислом Брожение аналогичны соответстветственным этапам спиртового брожение, вплоть до образования пировиноградной кислоты, из которой при маслянокислом брожение образуется ацетил-кофермент A (Ch4CO-KoA). Ацетил-KoA может служить предшественником масляной кислоты, подвергаясь следующим превращениям: Маслянокислое Брожение применялось для получения масляной кислоты из крахмала. Ацетоно-бутиловое брожение бактерии Clostridium acetobutylicum сбраживают углеводы с преим. образованием бутилового спирта (Ch4Ch3Ch3Ch3OH) и ацетона (Ch4COCh4). При этом образуются также в сравнительно небольших количествах водород, CO2, уксусная, масляная кислоты, этиловый спирт. Первые этапы расщепления углеводов те же, что и при спиртовом брожении. Бутиловый спирт образуется путём восстановления масляной кислоты: Ch4Ch3Ch3COOH + 4H =Ch4Ch3Ch3Ch3OH + h3O. Ацетон же образуется декарбоксилированием ацетоуксусной кислоты, которая получается в результате конденсации двух молекул уксусной кислоты. Исследованиями В. Н. Шапошникова показано, что ацетоно-бутиловое брожение (как и ряд др., например пропионовокислое, маслянокислое) в опытах с растущей культурой происходит в две фазы. В первую фазу брожение параллельно с нарастанием биомассы накапливаются уксусная и масляная кислоты; во вторую фазу образуются преимущественно ацетон и бутиловый спирт. При ацетоно-бутиловом брожение сбраживаются моносахариды, дисахариды и полисахариды — крахмал, инсулин, но не сбраживаются клетчатка и гемицеллюлоза. Ацетоно-бутиловое брожение использовалось для промышленного получения бутилового спирта и ацетона, применяемых в химической и лакокрасочной промышленности. Сбраживание белков. Некоторые бактерии из рода Clostridium — гнилостные анаэробы— способны сбраживать не только углеводы, но и аминокислоты. Эти бактерии более приспособлены к использованию белков,расщепляемых ими при помощи протеолитических ферментов до аминокислот, которые затем подвергаютсяброжение. Процесс сбраживания белковимеет значение в круговороте веществ в природе. Пропионовокислое Брожение. Основные продукты пропионовокислого брожение, вызываемого несколькими видами бактерий из рода Propionibacterium, — пропионовая (Ch4Ch3OH) и уксусная кислоты и CO2. Химизм пропионовокислого брожение сильно изменяется в зависимости от условий. Это, по-видимому, объясняется способностью пропионовых бактерий перестраивать обмен веществ, например в зависимости от аэрации. При доступе кислорода они ведут окислительный процесс, а в его отсутствии расщепляют гексозы путём брожение. Пропионовые бактерии способны фиксировать CO2, при этом из пировиноградной к-ты и CO2 образуется щавелевоуксусная к-та, превращающаяся в янтарную к-ту, из которой декарбоксилированием образуется пропионовая к-та: Существуют брожение, которые сопровождаются и восстановительными процессами. Примером такого «окислительного» брожение служит лимоннокислое брожение. Многие плесневые грибы сбраживают сахарас образованием лимонной кислоты. Наиболее активные штаммы Aspergillus niger превращают до 90% потребленногосахарав лимонную кислоту. Значительная часть лимонной кислоты, используемой в пищевой промышленности, производится микробиологическим путём — глубинным и поверхностным культивированием плесневых грибов. Иногда по традиции и чисто окислительные процессы, осуществляемые микроорганизмами, называется брожение. Примерами таких процессов могут служить уксуснокислое и глюконовокислое брожение. Уксуснокислое Брожение. Бактерии, относящиеся к роду Acetobacter, окисляют этиловый спирт в уксусную кислоту в соответствии с суммарной реакцией: Промежуточное соединение при окислении спирта в уксусную кислоту — уксусный альдегид. Многие уксуснокислые бактерии, кроме окисления спирта в уксусную кислоту, осуществляют окисление глюкозыв глюконовую и кетоглюконовую кислоты. Глюконовокислое Брожение осуществляют и некоторые плесневые грибы, способные окислять альдегидную группу глюкозы,превращая последнюю в глюконовую кислоту: Кальциеваясоль глюконовой кислоты служит хорошим источником кальция для людей и животных.
studfiles.net
Строение, примеры и функции углеводов
Углеводы — органические соединения, состав которых в большинстве случаев выражается общей формулой Cn(h3O)m (n и m ≥ 4). Углеводы подразделяются на моносахариды, олигосахариды и полисахариды.
Моносахариды — простые углеводы, в зависимости от числа атомов углерода подразделяются на триозы (3), тетрозы (4), пентозы (5), гексозы (6) и гептозы (7 атомов). Наиболее распространены пентозы и гексозы. Свойства моносахаридов — легко растворяются в воде, кристаллизуются, имеют сладкий вкус, могут быть представлены в форме α- или β-изомеров.
Рибоза и дезоксирибоза относятся к группе пентоз, входят в состав нуклеотидов РНК и ДНК, рибонуклеозидтрифосфатов и дезоксирибонуклеозидтрифосфатов и др. Дезоксирибоза (С5Н10О4) отличается от рибозы (С5Н10О5) тем, что при втором атоме углерода имеет атом водорода, а не гидроксильную группу, как у рибозы.
Глюкоза, или виноградный сахар (С6Н12О6), относится к группе гексоз, может существовать в виде α-глюкозы или β-глюкозы. Отличие между этими пространственными изомерами заключается в том, что при первом атоме углерода у α-глюкозы гидроксильная группа расположена под плоскостью кольца, а у β-глюкозы — над плоскостью.
Глюкоза — это:
- один из самых распространенных моносахаридов,
- важнейший источник энергии для всех видов работ, происходящих в клетке (эта энергия выделяется при окислении глюкозы в процессе дыхания),
- мономер многих олигосахаридов и полисахаридов,
- необходимый компонент крови.
Фруктоза, или фруктовый сахар, относится к группе гексоз, слаще глюкозы, в свободном виде содержится в меде (более 50%) и фруктах. Является мономером многих олигосахаридов и полисахаридов.
Олигосахариды — углеводы, образующиеся в результате реакции конденсации между несколькими (от двух до десяти) молекулами моносахаридов. В зависимости от числа остатков моносахаридов различают дисахариды, трисахариды и т. д. Наиболее распространены дисахариды. Свойства олигосахаридов — растворяются в воде, кристаллизуются, сладкий вкус уменьшается по мере увеличения числа остатков моносахаридов. Связь, образующаяся между двумя моносахаридами, называется гликозидной.
Сахароза, или тростниковый, или свекловичный сахар, — дисахарид, состоящий из остатков глюкозы и фруктозы. Содержится в тканях растений. Является продуктом питания (бытовое название — сахар). В промышленности сахарозу вырабатывают из сахарного тростника (стебли содержат 10–18%) или сахарной свеклы (корнеплоды содержат до 20% сахарозы).
Мальтоза, или солодовый сахар, — дисахарид, состоящий из двух остатков глюкозы. Присутствует в прорастающих семенах злаков.
Лактоза, или молочный сахар, — дисахарид, состоящий из остатков глюкозы и галактозы. Присутствует в молоке всех млекопитающих (2–8,5%).
Полисахариды — это углеводы, образующиеся в результате реакции поликонденсации множества (несколько десятков и более) молекул моносахаридов. Свойства полисахаридов — не растворяются или плохо растворяются в воде, не образуют ясно оформленных кристаллов, не имеют сладкого вкуса.
Крахмал (С6Н10О5)n — полимер, мономером которого является α-глюкоза. Полимерные цепочки крахмала содержат разветвленные (амилопектин, 1,6-гликозидные связи) и неразветвленные (амилоза, 1,4-гликозидные связи) участки. Крахмал — основной резервный углевод растений, является одним из продуктов фотосинтеза, накапливается в семенах, клубнях, корневищах, луковицах. Содержание крахмала в зерновках риса — до 86%, пшеницы — до 75%, кукурузы — до 72%, в клубнях картофеля — до 25%. Крахмал — основной углевод пищи человека (пищеварительный фермент — амилаза).
Гликоген (С6Н10О5)n — полимер, мономером которого также является α-глюкоза. Полимерные цепочки гликогена напоминают амилопектиновые участки крахмала, но в отличие от них ветвятся еще сильнее. Гликоген — основной резервный углевод животных, в частности, человека. Накапливается в печени (содержание — до 20%) и мышцах (до 4%), является источником глюкозы.
Целлюлоза (С6Н10О5)n — полимер, мономером которого является β-глюкоза. Полимерные цепочки целлюлозы не ветвятся (β-1,4-гликозидные связи). Основной структурный полисахарид клеточных стенок растений. Содержание целлюлозы в древесине — до 50%, в волокнах семян хлопчатника — до 98%. Целлюлоза не расщепляется пищеварительными соками человека, т.к. у него отсутствует фермент целлюлаза, разрывающий связи между β-глюкозами.
Инулин — полимер, мономером которого является фруктоза. Резервный углевод растений семейства Сложноцветные.
Гликолипиды — комплексные вещества, образующиеся в результате соединения углеводов и липидов.
Гликопротеины — комплексные вещества, образующиеся в результате соединения углеводов и белков.
Функции углеводов
| Функция | Примеры и пояснения |
| Энергетическая | Основной источник энергии для всех видов работ, происходящих в клетках. При расщеплении 1 г углеводов выделяется 17,6 кДж. |
| Структурная | Из целлюлозы состоит клеточная стенка растений, из муреина — клеточная стенка бактерий, из хитина — клеточная стенка грибов и покровы членистоногих. |
| Запасающая | Резервным углеводом у животных и грибов является гликоген, у растений — крахмал, инулин. |
| Защитная | Слизи предохраняют кишечник, бронхи от механических повреждений. Гепарин предотвращает свертывание крови у животных и человека. |
Похожие статьи:
poznayka.org
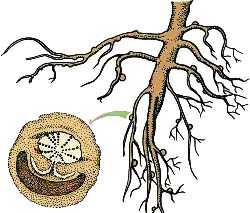
Сезонная динамика накопления и расходования луговыми растениями запасных веществ
Часть продуктов фотосинтеза сверх используемых на дыхание и построение новых органов растения откладывают в запас.
Они используются, когда потребность в энергетическом материале не покрывается текущим фотосинтезом (в начале весеннего отрастания, при отрастании после каждой дефолиации, в периоды, когда фотосинтетическая деятельность растений сильно снижается или прекращается). Запасные вещества представлены главным образом углеводами (растворимые углеводы, крахмал и др.) и белками и откладываются в приземных и подземных стеблевых органах (корневищах, зонах кущения, клубнях, луковицах в нижних частях надземных побегов и др.), а также в корнях. Из них наиболее изучены углеводы.
Обеспеченность луговых многолетних трав запасными веществами значительно изменяется в течение года. Об этом можно» судить при учете массы (веса) запасающих органов и содержания в них запасных веществ. По вегетационному сезону колеблется не только общая обеспеченность растений запасными углеводами, но и содержание различных форм углеводов. Осенью, перед уходом на зимовку, растения обычно хорошо обеспечены запасными веществами. Зимой часть этих веществ потребляется на дыхание, а в некоторых случаях и на ростовые процессы (зимой отмечен рост корней луговых растений), поэтому обеспеченность растений запасными веществами снижается. Особенно велики расходы запасных углеводов на дыхание в теплые зимы, когда растения под снегом вегетируют.
Снижение обеспеченности растений запасными веществами продолжается и ранней весной. Растения используют их как источник энергии и строительный материал при формировании новых листьев и побегов. С превышением же образования продуктов фотосинтеза над их расходованием у злаков в период весеннего кущения происходит накопление запасных углеводов. В фазы стеблевания, бутонизации, а иногда и начала цветения потребление углеводов может превышать их синтез, что ведет к снижению общего содержания их в органах запаса. Если число возникающих генеративных побегов невелико по сравнению с числом вегетативных побегов, то падения обеспеченности растений запасными углеводами может не происходить, приостанавливается. лишь их накопление. У ежи сборной от фазы «трубка» до начала колошения общее количество углеводов сохранилось на одном уровне. Начиная с фазы цветения углеводы образуются в больших количествах, чем они потребляются, а их постепенное накопление продолжается до осени. Лишь перед уходом на зимовку общее количество углеводов снижается, что может быть обусловлено различными причинами: переходом одних форм углеводов в другие, использованием их на образование новых побегов и корней, превышением расходования над синтезом.
Наибольшее накопление запасных углеводов происходите летне-осенний период, оно значительно превышает количество, накопленное весной. У отдельных видов луговых растений время второго максимума истощения запасных веществ приурочено то к фазе стеблевания, то к фазе бутонизации. В этот период растения особенно чувствительны к скашиванию или стравливанию. Это лучшее время для борьбы с сорняками путем подкашивания. Применение гербицидов наиболее эффективно в период интенсивного оттока ассимилятов из листьев в подземные органы, так как это ведет к поступлению гербицидов в многолетние органы растений и способствует их отмиранию. Во избежание угнетения ценных кормовых растений сроки их использования нельзя приурочивать к периодам максимального истощения запасных веществ, в частности «е следует их использовать ранней весной, когда органы запаса истощены. У особей, находящихся в вегетативном состоянии, если их рост не нарушается внешними воздействиями, от весны к осени происходит накопление запасных веществ.
Обычный ход динамики запасных веществ нарушается отчуждением надземных органов при скашивании или стравливании, которое прерывает накопление запасных веществ. При отрастании после дефолиации так же, как и при отрастании ранней весной, вначале запасные вещества расходуются на дыхание и построение вновь образующихся органов и соответственно этому происходит снижение содержания их в органах запаса. Например, снижение запаса углеводов после скашивания происходит у люцерны посевной примерно в течение 20, у клевера красного — 12—15, у райграса многолетнего — 7—11 дней. И только после этого срока происходит постепенное накопление запасных углеводов в растениях. В случае чрезмерно частого отчуждения надземных органов, когда растения не успевают накопить достаточное количество запасных веществ к следующему использованию обеспеченность ими прогрессивно снижается. Так, в опыте при двуукосном использовании клевера красного обеспеченность его запасными углеводами к осени возросла, а при четырехкратном — резко снизилась, падая от укоса к укосу.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info