
Что такое зародыш: строение, процесс формирования и развития. Зародыш растения
Из чего состоит зародыш семени растения? Строение зародыша семени
Прочитав эту статью, вы узнаете, из чего состоит зародыш семени. Кроме того, мы опишем основные стадии его развития.
Для того чтобы наглядно представить, о чем речь, обратимся к примеру. Затем перейдем к общей характеристике зародыша и стадий его развития.
Из чего состоит зародыш семени пшеницы

Если разрезать семя пшеницы, можно увидеть, что большую его часть составляет мучнистая белая ткань. Она называется эндоспермом. Ее клетки заполнены различными питательными веществами. Не так-то просто выяснить, из чего состоит зародыш семени пшеницы. Ведь он очень маленький. Его можно рассмотреть лишь с помощью лупы. Так же, как и у зародыша фасоли, у него есть стебелек, корешок и почечка. Однако семядоля у него лишь одна. Это тоненькая пластинка, плотно прилегающая к эндосперму.
Вот из чего состоит зародыш семени. Конечно, у разных видов растений он разный. Но на примере пшеницы можно представить его строение в общих чертах.
Разделение зародыша и суспензора

Ранние стадии развития зародыша у двудольных и однодольных очень сходны. Оно начинается с деления оплодотворенной яйцеклетки (зиготы) в зародышевом мешке семязачатка. У большинства цветковых плоскость первого деления проходит поперек (или почти поперек) ее продольной оси. При этом устанавливается полярность зародыша: верхний (халазальный) полюс - его главная ростовая зона, а нижний (микропилярный) образует своеобразную ножку — подвесок, или суспензор, заякоривающий зародыш у микропиле. После нескольких делений происходит окончательная дифференцировка на почти сферический собственно зародыш и суспензор.
Начало развития систем тканей
Из чего состоит зародыш семени, который только что сформировался? Из массы относительно недифференцированных клеток. Однако вскоре изменения его внутренней структуры приводят к началу развития систем растительных тканей. Будущая эпидерма (протодерма) формируется в ходе периклинальных делений наружных клеток собственно зародыша. Периклинальными называют деления, при которых клеточные пластинки между двумя дочерними клетками параллельны поверхности той части растения, где происходят деления.
Обособление прокабия и основной меристемы
В дальнейшем различия в степени вакуолизации и плотности клеток зародыша приводят к обособлению прокамбия и основной меристемы. Последняя, сильнее вакуолизированная и менее плотная, дает начало основной ткани, окружающей менее вакуолизированный и более плотный прокамбий, предшественник проводящих тканей — ксилемы и флоэмы.
Формирование семядолей
Протодерма, основная меристема и прокамбий (так называемые первичные меристемы), не прерываясь, переходят из семядолей в ось зародыша. Формирование семядолей может начинаться либо в процессе, либо после закладки первичных меристем (стадия развития до появления семядолей часто называется глобулярной). При этом глобулярный зародыш двудольных постепенно принимает двулопастную форму (эту стадию часто называют сердцевидной). Зародыш семени однодольных образует только одну семядолю. Поэтому он не имеет сердцевидной стадии.
Разрушение клеток суспензора
Затем семядоли и ось зародыша удлиняются (так называемая стадия торпеды) и первичные меристемы распределяются вдоль них. Вытягиваясь, зародыш остается прямым или искривляется. Единственная семядоля однодольных часто так сильно разрастается, что оказывается самой крупной зародышевой структурой. По мере дальнейшего роста зародыша клетки суспензора постепенно разрушаются.
Клеточные деления

На ранних стадиях эмбриогенеза клеточные деления происходят по всей массе молодого спорофита. Однако в ходе формирования зародыша появление новых клеток постепенно ограничивается апикальными меристемами побега и корня. У двудольных первая из них закладывается между двумя семядолями, а у однодольных с одной стороны семядоли и полностью окружена влагалище - подобным выростом ее основания. Апикальные меристемы имеют большое значение, так как являются в конечном итоге источником всех новых клеток, обеспечивающих развитие из зародыша проростка и взрослого растения.
Отделение семязачатка
В течение всего времени формирования зародыша питательные вещества постоянно перетекают от родительского растения к тканям семязачатка. В результате этого значительный их запас накапливается в эндосперме, перисперме или в семядолях развивающегося семени. В конце концов семязачаток отделяется от семяножки, связывающей его со стенкой завязи, и становится закрытой системой (в отношении питания). Семя высыхает, отдавая воду в окружающую среду, а семенная кожура отвердевает, как бы окружая "защитным панцирем" зародыш, а вместе с ним и запас питательных веществ.

Итак, вы узнали, из чего состоит зародыш семени. Как вы видите, по мере его развития происходит его трансформация. Поэтому строение зародыша семени отличается на разных этапах его существования.
fb.ru
Развитие эндосперма и зародыша у растений
Из оплодотворенной яйцеклетки, или зиготы, путем последовательных делений развивается зародыш, из оплодотворенной центральной клетки или клетки зачатка эндосперма развивается эндосперм.
После слияния спермия с яйцеклеткой, т. е. мужской и женской гаплоидных гамет, в оплодотворенной яйцеклетке восстанавливается диплоидное число хромосом. Первичное ядро эндосперма, являясь обычно продуктом слияния трех ядер — двух полярных и одного спермия — имеет триплоидное число хромосом. В редких случаях, когда эндосперм образуется без оплодотворения, его клетки имеют диплоидное число хромосом, а иногда и гаплоидное (представители семейства Onagraceae). Наконец, у тех растений, у которых при образовании вторичного ядра сливаются не два, а несколько ядер (3, 4, 7, 8 и даже 14), эндосперм может быть более или менее полиплоидным. После оплодотворения яйцеклетка и центральная клетка зародышевого мешка становятся способными к активному развитию. Двойное оплодотворение отсутствует у всех архегониальных растений и свойственно лишь покрытосеменным.
Во время развития зародыша и эндосперма зародышевый мешок нуждается в особенно обильном притоке питательных веществ, который осуществляется за счет извлечения их из окружающих материнских тканей, а также благодаря непосредственному потреблению зародышевым мешком тканей нуцеллуса и покровов семяпочки. Этому способствуют различного типа гаустории (присоски), развитые у многих растений. Под воздействием интенсивного обмена веществ размеры зародышевого мешка значительно увеличиваются. Наряду с этим стимулируется дальнейший рост и развитие семяпочки.
Период времени, протекающий от оплодотворения до образования зародыша и эндосперма, у разных растений неодинаков, колеблясь от 10—15 дней (у кок-сагыза, салата и др.) до нескольких недель (у злаков, хлопчатника и др.) и даже до нескольких месяцев (у черного саксаула и др.).
После двойного оплодотворения чаще всего первым делится первичное ядро эндосперма (продукт слияния вторичного — центрального ядра зародышевого мешка со спермием). Эндосперм у разных растений формируется неодинаково. Различают три основных типа развития эндосперма: ядерный, или нуклеарный, клеточный, или целлюлярный, и базальный, или тип Helobiae. В каждом из этих типов имеется ряд более или менее значительных модификаций.
Ядерный тип образования эндосперма характеризуется тем, что вслед за делением первичного ядра эндосперма не происходит немедленного образования клеточных перегородок, а возникает большее или меньшее число ядер, свободно лежащих в общей цитоплазме.

Формирование клеточных перегородок при ядерном типе развития эндосперма наступает позднее, начинаясь либо в микропилярной части зародышевого мешка, либо по всей периферии его. В редких случаях при ядерном типе эндосперма клеточные оболочки вообще не закладываются. Клеточный тип, напротив, характеризуется тем, что после первого и каждого из последующих делений ядер эндосперма немедленно появляются клеточные перегородки.

Базальный тип имеет промежуточный характер между ядерным и клеточным. Он отличается тем, что возникающая сразу после первого деления ядра эндосперма клеточная перегородка делит центральную клетку на две неравные части: большую верхнюю, или микропилярную, и маленькую нижнюю, или базальную. В микропилярной части эндосперма сначала образуется ряд свободных ядер и только позднее между ядрами появляются клеточные перегородки; в нижней же части либо образуются две клетки, либо небольшое число свободных ядер, либо делений вообще больше не происходит. Эта антиподальная часть эндосперма, или его базальный аппарат, функционирует часто как гаусторий.

Эндоспермальные гаустории формируются также при клеточном, а иногда и при ядерном типе развития эндосперма, возникая как в микропилярной, так и в халазальной и даже в латеральной части зародышевого мешка.

Гаусториями называются образования, состоящие из одно- или многоядерных клеток с густым содержимым и часто с гипертрофированными клеточными ядрами. В одних случаях гаустории состоят из нескольких более или менее разросшихся и разветвленных клеток и даже целой ткани, в других — только из одной клетки, сильно разрастающейся и дающей боковые отростки.
Наряду с эндоспермальными гаусториями в семяпочках покрытосеменных известны и другие типы гаусторий, образованных самим зародышевым мешком, антиподами, синергидами, подвеском зародыша. Гаустории весьма сильно варьируют по своему облику и строению, но роль их, по-видимому, одна и та же; они приспособлены к выполнению питательной и секреторной функции. Проникая своими отростками и боковыми ветвями глубоко в ткани семяпочки, плаценты и даже завязи, гаустории извлекают из материнского растения питательные вещества для роста и формирования зародыша. Эндоспермальные гаустории, как и гаустории любого другого типа, способствуют повышению интенсивности обмена веществ во время роста и развития зародыша, так как облегчают приток к нему питательных веществ из окружающих тканей материнского растения.
Примером гаусторий антиподального происхождения могут служить гаустории злаков. Комплекс антипод образует своеобразное подобие ткани, внедряющейся в нуцеллус. Клетки нуцеллуса, непосредственно примыкающие к клеткам антиподального комплекса, разрушаются.
При гибридизации и самоопылении перекрестников, а также под воздействием неблагоприятных внешних условий развитие эндосперма может преждевременно приостанавливаться или с самого начала происходит ненормально. Это, в свою очередь, сразу сказывается на формировании зародыша и приводит к образованию щуплых семян с недоразвитым зародышем.
В сформированном виде эндосперм зрелого семени (тех растений, у которых он сохраняется) представлен тканью, состоящей из более или менее крупных клеток, заполненных различными питательными веществами — крахмалом, маслом, сахарами, белком и т. д. Кроме того, в клетках эндосперма могут присутствовать дубильные вещества, алкалоиды, кристаллы оксалата кальция и т. д. Оболочки эндоспермальных клеток обычно тонкие, целлюлозные. У некоторых растений, у которых основное запасное вещество откладывается в оболочках, последние значительно утолщаются.
По своей консистенции эндосперм у одних растений более или менее жидкий или студенистый, у других — в той или иной степени твердый или даже роговой. Эндосперм бывает либо бесцветным, либо различно пигментированным.
В дальнейшем эндосперм претерпевает различные превращения. У одних растений он полностью ассимилируется зародышем в процессе развития последнего и в зрелом семени или отсутствует полностью, или представлен одним-двумя слоями клеток. К таким растениям принадлежат бобовые, тыквенные, сложноцветные и другие. В этих случаях запасные питательные вещества, необходимые для прорастания зародыша, накапливаются в клетках семядолей самого зародыша, и последний заполняет собой почти весь объем семени.
У других растений эндосперм сохраняется до созревания семени и потребляется зародышем лишь при его прорастании (злаковые, ситниковые, магнолиевые и др.). В этих случаях зародыши малы по объему, а основную часть семени занимает эндосперм. Между этими крайними примерами существует, конечно, много переходных форм, среди которых встречаются и такие, у которых объемы эндосперма и зародыша в зрелом семени одинаковые.
У некоторых растений (например, из семейства гвоздичных, перечноцветных) запасные вещества откладываются в нуцеллусе. Подобного рода запасная ткань называется периспермом.
Оплодотворенная яйцеклетка, или зигота, может делиться одновременно с делением первичного ядра эндосперма, но, как правило, зигота делится несколько позже (иногда значительно позже) — лишь после образования нескольких, а в некоторых случаях большого числа ядер или клеток эндосперма. Во время созревания зиготы она сама и ее ядро значительно увеличиваются в размерах, вакуоля постепенно исчезает, и вся клетка заполняется более или менее густым содержимым, концентрирующимся главным образом в апикальной ее части. По окончании периода созревания зигота начинает делиться, давая начало зародышу.
Имеется несколько классификаций типов развития зародыша, в основе которых лежат следующие признаки: способ заложения первой перегородки, делящей зиготу на две клетки, направления перегородок при последующих делениях, а также участие тех или иных клеток начинающего развиваться зародыша в формировании его органов.
При развитии зиготы покрытосеменного растения возникает клеточное тело, которое обычно уже на начальных фазах своего развития расчленено на базальную часть — подвесок, или суспензор, и терминальную — тело зародыша. Независимо от способа заложения клеточных перегородок зародыши покрытосеменных растений на самых ранних стадиях внешне довольно единообразны и являются радиально симметричными. В этот период нет различий даже между зародышами двудольных и однодольных растений, эти различия появляются позже. У двудольных растений тело зародыша постепенно принимает форму шара, который в дальнейшем несколько уплощается, после чего происходит дифференциация на семядоли и осевую часть. Через такой зародыш можно теперь провести только две плоскости симметрии.

При развитии зародыша однодольных растений шарообразная стадия отсутствует, а зародыш из радиально симметричного становится билатерально симметричным, т. е. через него можно провести лишь одну плоскость симметрии. В некоторых случаях, например у злаков, это объясняется тем, что здесь очень рано нарушается равномерность ритма деления клеток зародыша— на вентральной его стороне деления задерживаются, в то время как на противоположной дорзальной стороне и в апикальной части зародыша они протекают интенсивно. В результате зародыш изгибается, а это в свою очередь обусловливает иной по сравнению с двудольными характер заложения органов. Единственная семядоля закладывается терминально, а точка роста занимает боковое положение. В других случаях не исключены и другие пути, приводящие к становлению своеобразного зародыша однодольных.

Несколько особняком в отношении формирования зародыша среди покрытосеменных растений стоит семейство Леоновых. У его представителей деление зиготы и последующие деления производных ядер не сопровождаются заложением клеточных перегородок. В результате образуется предзародышевая ценоцитная структура, очень сходная с тем, что имеет место у голосеменных растений. Лишь позже между ядрами такого ценоцита закладываются перегородки и возникают меристематические очаги, дающие начало зародышам. Рост и дифференциация зародыша происходят в процессе активного обмена веществ, осуществляющегося между ним и окружающей его средой. Выше уже говорилось о том, что этому способствуют различного рода гаусториальные образования, формируемые различными структурами зародышевого мешка, в первую очередь эндоспермом. Такого рода гаусториальные функции свойственны также и самому зародышу, в частности его подвеску. При помощи подвеска зародыш продвигается в ткань эндосперма и получает наиболее благоприятные возможности для его использования; при помощи подвеска зародыш может получать питательные вещества и непосредственно из материнских тканей. Облегчая поступление к зародышу питательных веществ из окружающих его тканей семяпочки, подвесок способствует повышению интенсивности обмена веществ на ранних стадиях роста и развития зародыша.
Подвески зародышей бывают одноклеточными и многоклеточными, клетки в них могут быть одноядерными или многоядерными. В некоторых случаях подвесок сильно разрастается и разветвляется, образуя подвесочный гаусторий. Иногда же подвесок совершенно не развивается или развивается крайне слабо.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Зародыш - это... Что такое Зародыш?
1) у животных и человека З., или эмбрион, — организм в раннем периоде развития — от оплодотворённого яйца (зиготы (См. Зигота)) до появления способности к самостоятельному питанию и активному передвижению. Развитие З. происходит, как правило, в яйцевых оболочках или специальных органах материнского организма. См. Зародышевое развитие, Зародышевые оболочки. 2) У растений З., или эмбрио, — зачаток новой особи, развивающийся из зиготы, которая возникает в результате оплодотворения яйцеклетки зародышевого мешка (См. Зародышевый мешок) или из клеток нуцеллуса или интегумента (в случае Апомиксиса). Наиболее простой тип эмбрионального развития у зелёных водорослей, например у спирогиры, зигота которой не образует спорофитной нити, а сразу приступает к Мейозу. У бурых водорослей, например у фукуса, зигота прикрепляется к субстрату, делится, после чего образует эмбриональное тело с точкой роста Таллома (Рис.1). У мохообразных З. прорастает на материнском организме — Гаметофите, за счёт которого он питается, затем образует спорогенную ткань, где формируются споры (Рис.2). Такой тип З. не встречается среди др. групп высших растений. У древнейших высших растений — псилотовых — развитие З. протекает в ткани гаметофита, где происходит ряд последовательных делений зиготы. Из клеток нижней части формируется ножка, внедряющаяся в ткань гаметофита, в верхней — закладывается точка роста побега с двумя меристематическими бугорками (см. Меристема). З. здесь — поляризованная структура, один конец которой образует наземный побег, другой — корневище (Рис.3). У плауновидных З. имеет два симметричных бугорка будущих зародышевых листочков, между которыми находится точка роста в виде верхушечной клетки (Рис.4). Развитие З. у хвощей характеризуется правильным планом сегментации в виде 4 зон, дающих начало побегу, корню, первому листу и ножке (Рис.5). У папоротниковидных в первых 4 клетках, возникших в результате двух первых делении зиготы, определено место закладки верхушки побега, первого листа, корня и ножки (Рис.6). У голосеменных начальный цикл развития З. характеризуется свободноядерной, или ценоцитной, стадией (у саговников, например, свободных ядер 1024), которая сменяется образованием клеток; затем в формирующемся З. происходит закладка точек роста побега, корня и семядольных бугорков (у некоторых видов сосны — до 18) (Рис.7). У большинства покрытосеменных зигота делится на две клетки: апикальную, дающую начало собственно З., п базальную — подвеску, или суспензору. В Эмбриогенезе различают 2 фазы развития: предзародышевую (от зиготы до образования эмбриодермы) и зародышевую (закладка основных структур: точки роста побега, семядоли, гипокотиля и центрального зародышевого корешка). На основе первых этапов деления предзародыша были созданы классификации эмбриональных типов, используемые систематиками при решении вопросов эволюции и филогении растений. Предзародышевая стадия развития двудольных и однодольных одинакова. На более поздних стадиях у однодольных апикальная зона проэмбрио даёт начало только 1 семядольному бугорку; у двудольных таких бугорков может возникнуть 2, а у некоторых видов 3 и даже больше, что приводит к образованию многосемядольности. У покрытосеменных растений развитие З., в отличие от голосеменных, сопровождается образованием клеточных перегородок (Рис.8,9). Лишь у пионов обнаружен новый тип эмбриогенеза, при котором в зиготе вначале возникают свободные ядра, а затем такая многоядерная структура становится клеточной и на её периферии возникают проэмбриональные бугорки; из них в собственно З. развивается только один. По наличию или отсутствию у З. покрытосеменных зелёного пигмента — хлорофилла — они делятся на 2 группы: хлороэмбриофиты (бобовые, крестоцветные и др.) илейкоэмбриофиты (лютиковые, злаковые и др.). В процессе эволюции среди некоторых групп покрытосеменных, например у растений паразитов (заразиховые) и сапрофитов (орхидные), наблюдается -редукция З. Источником питания З. служит Эндосперм, который у одних видов (бобовые) поглощается целиком растущим З., у др. (злаки) — сохраняется. Иногда З. в семени окружен не только эндоспермом, но и др. запасной тканью — периспермом, развивающимся из клеток нуцеллуса (свёкла, чёрный перец и др.).Лит.: Баранов П. А., История эмбриологии растений в связи с развитием представлений о зарождении организмов, М.— Л., 1955; Яковлев М. С., Эмбриогенез и его значение для филогении растений, М. — Л., 1960; Поддубная-Арнольди В. А., Общая эмбриология покрытосеменных растений, М., 1964; Тахтаджян А. Л., Основы эволюционной морфологии покрытосеменных, М. — Л., 1964.
М. С. Яковлев.

Рис.1. Развитие зародыша у растений разных систематических групп. У бурой водоросли фукус (а — г).

Рис.2. Развитие зародыша у растений разных систематических групп. У мохообразных (Conocephalum conicum): a — зигота, б — первое деление зиготы горизонтальной перегородкой, в — образование поперечных перегородок, г — апикальный и базальный концы зародыша, дифференциация археспориальной ткани, д — зрелый зародыш, ставший спорогонием.

Рис.3. Развитие зародыша у растений разных систематических групп. У псилотовых (Tmesipteris tannensis): а — первое деление зиготы, б и в — проэмбрио, г — закладка апикальных клеток, д — начало образования дихотомически разветвленного зародыша.

Рис.4. Развитие зародыша у растений разных систематических групп. У плауновидных (Selaginella martensii): а, б, в — закладка апикальной точки роста (заштриховано), г — зрелый зародыш с прокамбиальным тяжом, точкой роста побега и листовыми зачатками, д — зародыш в ткани гаметофита.

Рис.5. Развитие зародыша у растений разных систематических групп. У хвощей (Equisetum debileu Е. arwense): а и б — начальные фазы эмбриогенеза, в — более поздняя фаза, в апикальной части видна верхушечная клетка стебля.
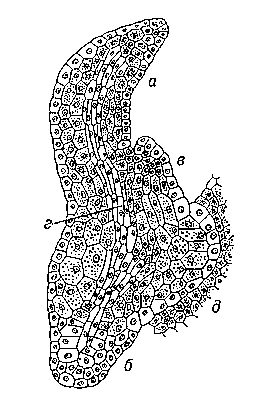
Рис.6. Развитие зародыша у растений разных систематических групп. У папоротника (Onoklea sp.): зрелый зародыш, хорошо развит первый лист (а) и корешок (б), закладывается второй лист (в), виден прокамбиальный тяж (г), дающий начало сосудистой системе, справа ножка (д), через которую в зародыш поступают питательные вещества из ткани гаметофита, или заростка.

Рис.7. Развитие зародыша у растений разных систематических групп. У голосеменных (Ginkgo biloba): а — проэмбрио в свободноядерной фазе, б — клеткообразование, в — проэмбрио, начало дифференциации клеток апикальной зоны, г — появление семядольных бугорков и точки роста, д — зрелый, хорошо дифференцированный зародыш.

Рис.8. Зародыш растений: 1 — зародыш (а) в семени кедровой сосны; 2—8 — различные стадии развития зародыша у некоторых видов табака: 2 — оплодотворённые яйцеклетка и центральная клетка зародышевого мешка, 3 — двухклеточный предзародыш, 4 — четырёхклеточный предзародыш, 5—6 — многоклеточные предзародыши с подвесками и эндоспермом.

Рис.9.: Зародыш растений: 7 — начало дифференциации зародыша (закладываются бугорки семядолей), 8 — дифференцированный зародыш, окруженный эндоспермом; у зародыша имеются две семядоли, зачатки стебля и корня, точки роста стебля и корня, корневой чехлик; 9—10 — продольный разрез через семя и зародыш овса Avena sativa; зародыш дифференцирован и имеет: а — одну семядолю (щиток), б — влагалище (колеоптиле), в — первые листочки (перышко), г — точку роста стебля, д — зачатки стебля и е — корня, ж — корневой чехлик, з — колеоризу, и — эпибласт; 11 — семя с недифференцированным зародышем у орхидеи Lycaste skinneri; 12 — зародыш с двумя складчатыми семядолями у хлопчатника Gossypium herbaceum.
dic.academic.ru
строение, процесс формирования и развития :: SYL.ru
Зарождение нового организма - это всегда загадка. Из нашей статьи вы узнаете, что такое зародыш. Эту структуру еще называют эмбрионом. Как же происходит процесс его развития?
Что такое зародыш
Сначала давайте разберемся, у каких организмов наблюдается формирование эмбриона. Среди растений это высшие споровые и семенные растения. У животных данный процесс происходит у многоклеточных представителей.
Что такое зародыш? Это зачаточный организм, который содержит все части взрослой особи. У растений зародышем называют зачаток нового спорофита. Так называют бесполое поколение в их жизненном цикле.
Типы оплодотворения животных
И у растений, и у животных зародыш образуется в результате слияния специализированных половых клеток - гамет. Этот процесс и называют оплодотворением.
Различают два типа этого процесса: внешнее и внутреннее. В первом случае оплодотворение происходит в окружающей среде. В основном оно характерно для водных обитателей. К примеру, двустворчатых моллюсков, раков, ланцетника, иглокожих, многощетинковых червей, земноводных и некоторых костных рыб.
Почему в других средах обитания не происходит этот процесс? Дело в том, что зародыш, который формируется вне материнского организма, подвержен многим неблагоприятным воздействиям экологических факторов. Это перепады температур, недостаток влаги, действие ультрафиолетового излучения. При этом яйцеклетки способны выделять особые вещества. Они привлекают половые клетки только своего биологического вида, что предотвращает межвидовое скрещивание.
Внутреннее оплодотворение происходит в половой системе самки. Этот процесс имеет массу преимуществ. Поскольку негативные условия окружающей среды утрачивают свое влияние, зародыш имеет больше шансов выжить. После проникновения сперматозоида в яйцеклетку ее оболочка становится непроницаемой для других мужских гамет. В результате оплодотворения образуется зигота, из которой развивается зародыш.

Зародыш семени
Как происходит оплодотворение и формирование зародыша растений? У представителей высших споровых этот процесс еще связан с водой. Для этих растений характерно чередование поколений в жизненном цикле. Из спор развивается особь полового поколения, в гаметангиях которых развиваются яйцеклетки и сперматозоиды. Они сливаются с помощью воды. Из оплодотворенной яйцеклетки развивается зародыш. Он прорастает и превращается в особь бесполого поколения.
Что такое зародыш у покрытосеменных растений? Это результат сразу двух процессов - опыления и оплодотворения. Первый заключается в переносе мужских гамет с пыльника тычинки на рыльце пестика. Оттуда пыльцевое зерно прорастает в виде трубки в зародышевый мешок. Здесь происходит оплодотворение. В его ходе принимают участие два спермия. Один сливается с женской гаметой, образуя зиготу.
Другой спермий соединяется с центральной зародышевой клеткой, образуя эндосперм. Это запасное питательное вещество. Его использует зародыш в ходе своего развития. Поэтому оплодотворение цветковых называют двойным.
Со временем развивается зародыш. Он содержит все части взрослого растения. Это зародышевый корешок, стебель, почечка и листья. Такое же строение имеют и зародыши пшеницы. Это растение из класса однодольные. Кроме злаков к ним относятся семейства лилейные и луковые. Все они имеют одну семядолю в зародыше семени.
Семейства розоцветные, пасленовые, астровые, капустные, бобовые относятся к классу двудольные. Их семена содержат два видоизмененных зародышевых листа или семядоли.

Биологическое значение
Зародыши пшеницы, как и других растений, кроме эндосперма снаружи защищены кожурой. Благодаря такому строению обеспечивается жизнеспособность будущего растения. Зародыш семени способен переживать даже продолжительные неблагоприятные периоды. К примеру, засуху и низкие температуры. Когда условия нормализируются, зародыш прорастает и дает начало новому организму.

Развитие эмбрионов
Онтогенез животных бывает трех типов: личиночный, яйцекладный и внутриутробный. В первом случае оплодотворенная яйцеклетка содержит ограниченный запас питательных веществ. Поэтому она быстро развивается в личинку. Такой тип развития встречается у земноводных, рыб и насекомых. Зародыш яйцекладущих млекопитающих, птиц и рептилий развивается без личиночной стадии. Их яйцеклетки богаты желтком.
Развитие зародыша человека - пример внутриутробного развития. Он задерживается в материнском организме, с которым связан особым органом - плацентой. В ходе эмбрионального развития происходит последовательное дробление зиготы. В результате образуется многоклеточная структура, которая дает начало всем органам взрослой особи.
Итак, что такое зародыш? Это организм, который находится на ранних стадиях развития. Он содержит зачатки будущей взрослой особи и имеет все необходимые для развития ресурсы.
www.syl.ru
Зародыш | Авторская платформа Pandia.ru
Зародыш, 1) у животных и человека З., или эмбрион, — организм в раннем периоде развития — от оплодотворённого яйца (зиготы) до появления способности к самостоятельному питанию и активному передвижению. Развитие З. происходит, как правило, в яйцевых оболочках или специальных органах материнского организма. См. Зародышевое развитие, Зародышевые оболочки.
2) У растений З., или эмбрио, — зачаток новой особи, развивающийся из зиготы, которая возникает в результате оплодотворения яйцеклетки зародышевого мешка или из клеток нуцеллуса или интегумента (в случае апомиксиса). Наиболее простой тип эмбрионального развития у зелёных водорослей, например у спирогиры, зигота которой не образует спорофитной нити, а сразу приступает к мейозу. У бурых водорослей, например у фукуса, зигота прикрепляется к субстрату, делится, после чего образует эмбриональное тело с точкой роста таллома (Рис.1). У мохообразных З. прорастает на материнском организме — гаметофите, за счёт которого он питается, затем образует спорогенную ткань, где формируются споры (Рис.2). Такой тип З. не встречается среди др. групп высших растений. У древнейших высших растений — псилотовых — развитие З. протекает в ткани гаметофита, где происходит ряд последовательных делений зиготы. Из клеток нижней части формируется ножка, внедряющаяся в ткань гаметофита, в верхней — закладывается точка роста побега с двумя меристематическими бугорками (см. Меристема). З. здесь — поляризованная структура, один конец которой образует наземный побег, другой — корневище (Рис.3). У плауновидных З. имеет два симметричных бугорка будущих зародышевых листочков, между которыми находится точка роста в виде верхушечной клетки (Рис.4). Развитие З. у хвощей характеризуется правильным планом сегментации в виде 4 зон, дающих начало побегу, корню, первому листу и ножке (Рис.5). У папоротниковидных в первых 4 клетках, возникших в результате двух первых делении зиготы, определено место закладки верхушки побега, первого листа, корня и ножки (Рис.6). У голосеменных начальный цикл развития З. характеризуется свободноядерной, или ценоцитной, стадией (у саговников, например, свободных ядер 1024), которая сменяется образованием клеток; затем в формирующемся З. происходит закладка точек роста побега, корня и семядольных бугорков (у некоторых видов сосны — до 18) (Рис.7). У большинства покрытосеменных зигота делится на две клетки: апикальную, дающую начало собственно З., п базальную — подвеску, или суспензору.
В эмбриогенезе различают 2 фазы развития: предзародышевую (от зиготы до образования эмбриодермы) и зародышевую (закладка основных структур: точки роста побега, семядоли, гипокотиля и центрального зародышевого корешка). На основе первых этапов деления предзародыша были созданы классификации эмбриональных типов, используемые систематиками при решении вопросов эволюции и филогении растений. Предзародышевая стадия развития двудольных и однодольных одинакова. На более поздних стадиях у однодольных апикальная зона проэмбрио даёт начало только 1 семядольному бугорку; у двудольных таких бугорков может возникнуть 2, а у некоторых видов 3 и даже больше, что приводит к образованию многосемядольности. У покрытосеменных растений развитие З., в отличие от голосеменных, сопровождается образованием клеточных перегородок (Рис.8,9). Лишь у пионов обнаружен новый тип эмбриогенеза, при котором в зиготе вначале возникают свободные ядра, а затем такая многоядерная структура становится клеточной и на её периферии возникают проэмбриональные бугорки; из них в собственно З. развивается только один. По наличию или отсутствию у З. покрытосеменных зелёного пигмента — хлорофилла — они делятся на 2 группы: хлороэмбриофиты (бобовые, крестоцветные и др.) илейкоэмбриофиты (лютиковые, злаковые и др.). В процессе эволюции среди некоторых групп покрытосеменных, например у растений паразитов (заразиховые) и сапрофитов (орхидные), наблюдается – редукция З. Источником питания З. служит эндосперм, который у одних видов (бобовые) поглощается целиком растущим З., у др. (злаки) — сохраняется. Иногда З. в семени окружен не только эндоспермом, но и др. запасной тканью — периспермом, развивающимся из клеток нуцеллуса (свёкла, чёрный перец и др.).
Лит.: Баранов П. А., История эмбриологии растений в связи с развитием представлений о зарождении организмов, М.— Л., 1955; Яковлев М. С., Эмбриогенез и его значение для филогении растений, М. — Л., 1960; Поддубная-Арнольди В. А., Общая эмбриология покрытосеменных растений, М., 1964; Тахтаджян А. Л., Основы эволюционной морфологии покрытосеменных, М. — Л., 1964.
М. С. Яковлев.

Рис.5. Развитие зародыша у растений разных систематических групп. У хвощей (Equisetum debileu Е. arwense): а и б — начальные фазы эмбриогенеза, в — более поздняя фаза, в апикальной части видна верхушечная клетка стебля.

Рис.9.: Зародыш растений: 7 — начало дифференциации зародыша (закладываются бугорки семядолей), 8 — дифференцированный зародыш, окруженный эндоспермом; у зародыша имеются две семядоли, зачатки стебля и корня, точки роста стебля и корня, корневой чехлик; 9—10 — продольный разрез через семя и зародыш овса Avena sativa; зародыш дифференцирован и имеет: а — одну семядолю (щиток), б — влагалище (колеоптиле), в — первые листочки (перышко), г — точку роста стебля, д — зачатки стебля и е — корня, ж — корневой чехлик, з — колеоризу, и — эпибласт; 11 — семя с недифференцированным зародышем у орхидеи Lycaste skinneri; 12 — зародыш с двумя складчатыми семядолями у хлопчатника Gossypium herbaceum.

Рис.1. Развитие зародыша у растений разных систематических групп. У бурой водоросли фукус (а — г).

Рис.8. Зародыш растений: 1 — зародыш (а) в семени кедровой сосны; 2—8 — различные стадии развития зародыша у некоторых видов табака: 2 — оплодотворённые яйцеклетка и центральная клетка зародышевого мешка, 3 — двухклеточный предзародыш, 4 — четырёхклеточный предзародыш, 5—6 — многоклеточные предзародыши с подвесками и эндоспермом.

Рис.4. Развитие зародыша у растений разных систематических групп. У плауновидных (Selaginella martensii): а, б, в — закладка апикальной точки роста (заштриховано), г — зрелый зародыш с прокамбиальным тяжом, точкой роста побега и листовыми зачатками, д — зародыш в ткани гаметофита.

Рис.7. Развитие зародыша у растений разных систематических групп. У голосеменных (Ginkgo biloba): а — проэмбрио в свободноядерной фазе, б — клеткообразование, в — проэмбрио, начало дифференциации клеток апикальной зоны, г — появление семядольных бугорков и точки роста, д — зрелый, хорошо дифференцированный зародыш.

Рис.3. Развитие зародыша у растений разных систематических групп. У псилотовых (Tmesipteris tannensis): а — первое деление зиготы, б и в — проэмбрио, г — закладка апикальных клеток, д — начало образования дихотомически разветвленного зародыша.

Рис.2. Развитие зародыша у растений разных систематических групп. У мохообразных (Conocephalum conicum): a — зигота, б — первое деление зиготы горизонтальной перегородкой, в — образование поперечных перегородок, г — апикальный и базальный концы зародыша, дифференциация археспориальной ткани, д — зрелый зародыш, ставший спорогонием.

Рис.6. Развитие зародыша у растений разных систематических групп. У папоротника (Onoklea sp.): зрелый зародыш, хорошо развит первый лист (а) и корешок (б), закладывается второй лист (в), виден прокамбиальный тяж (г), дающий начало сосудистой системе, справа ножка (д), через которую в зародыш поступают питательные вещества из ткани гаметофита, или заростка.
pandia.ru










