
Плотность и численность популяций. Возрастной состав популяций растений
Плотность и численность популяций
В любой природной системе поддерживается та численность особей в популяциях, которая в наибольшей степени отвечает интересам воспроизводства. Режим численности зависит от постоянно действующих регулирующих экологических факторов.
Для того, чтобы сравнить численность отдельных популяций или изменение численности одной и той же популяции в разные отрезки времени, целесообразно пользоваться таким относительным показателем, как плотность, то есть численность популяции, отнесенная к единице занимаемого ею пространства или среднее число особей на единицу площади или объема.
Зная изменение плотности во времени или пространстве, можно установить, увеличивается или уменьшается численность особей, представляет или нет данная популяция угрозу хозяйственным интересам. Динамика плотности популяций отражает сложные закономерности взаимоотношений между различными животными, между животными и растениями, поскольку все они являются биотическими факторами по отношению друг к другу. Кроме того, плотность зависит и от колебаний абиотических факторов среды.
Иногда бывает важно различать среднюю плотность, то есть численность (или биомассу) на единицу всего пространства, иудельную, илиэкологическую, плотность, то есть численность (или биомассу) на единицу обитаемого пространства (доступной площади или объема, которые фактически могут быть заняты популяцией).
Чем ниже трофический уровень, тем выше плотность, а чем крупнее животные внутри данного уровня, тем больше их биомасса. Так как у крупных животных интенсивность метаболизма на единицу массы меньше, чем у мелких, на данной энергетической базе может поддерживаться большая биомасса крупных животных.
Возрастной состав популяции
Возрастной состав популяции– это соотношение разных возрастных групп в популяции. Он определяет ее способность к размножению в данный момент и показывает, чего можно ожидать в будущем. Обычно в быстро растущих популяциях значительную долю составляют молодые особи. В популяции, находящейся в стационарном состоянии, возрастное распределение более равномерно, а в популяции, численность которой снижается, будет содержаться большая доля старых особей.
Возрастная структура популяции может изменяться и без изменения ее численности.
По мере развития человеческого общества от состояния освоения новых территорий и быстрого роста численности к зрелому состоянию стабильной популяции процент населения в младших возрастных классах уменьшается. Это изменение возрастной структуры и увеличение процента населения старших возрастных классов оказывает глубокое воздействие на жизненный уклад общества и экономику.
Упрощенно в популяции можно выделить три экологические возрастные группы: пререпродуктивную,репродуктивнуюипострепродуктивную. Длительность этих возрастов по отношению к общей продолжительности жизни сильно варьирует у разных организмов. У современного человека три эти возрастные периоды приблизительно одинаковы, на каждый из них приходится около трети жизни. У первобытных людей пострепродуктивный период был намного короче. Для многих животных и растений характерен очень длительный пререпродуктивный период. У некоторых животных, в особенности у насекомых, пререпродуктивный период чрезвычайно длителен, репродуктивный период очень короткий, а пострепродуктивный период отсутствует совсем.
studfiles.net
Возрастной состав популяций
(син. - возрастной спектр популяции онтогенетический состав популяции, онтогенетический спектр популяции), соотношение групп особей разного абсолютного или биологического возраста в составе популяции. Возрастной состав популяций представляет собой один из важнейших признаков популяции, определяющий ее устойчивость и способность к самоподдержанию. Возрастной состав популяций животных чаще всего определяется по соотношению числа особей разного абсолютного возраста, возрастной состав популяций растений - по соотношению числа особей или их частей (кустов, побегов) с разным возрастным состоянием. Различают 3 типа возрастного состава популяций: 1) инвазионная (внедряющаяся, расширяющаяся) включает зачатки, преобладают проростки и молодые особи, генеративные особи практически или полностью отсутствуют. Пример такой популяции - густой еловый подрост в березовом лесу, ель в этом случае представляет инвазионную популяцию; 2) нормальная, представленная всеми возрастными группами. Напр., сосна в сухом сосновом бору, где представлены как взрослые особи, так и хорошо развитый подрост, или ненарушенные еловые древостой, образованные разными поколениями ели - от подроста до спелых и перестойных; 3) регрессивная - преобладают сенильные или генеративные особи, не дающие полноценных зачатков {плодов, семян, спор и т. д.). Напр., березовый лес с густым еловым подростом. Береза, образующая верхний ярус, не возобновляется, поэтому ее популяция может считаться регрессивной. Инвазионные популяции находятся в стадии становления и, в зависимости от онтогенетического состава и численности особей, с одной стороны, и эколого-ценотических условий - с другой, имеют более или менее вероятные перспективы развития в нормальные. Последние полностью способны к спонтанному самоподдержанию семенным и/или вегетативным путем. Лесной биогеоценоз, находящийся в стадии климакса, характеризуется нормальными популяциями всех видов деревьев, входящих в его состав. Отсутствие отдельных онтогенетических групп в спектре нормальных популяций может быть связано с периодичностью плодоношения и, как правило, не является свидетельством неустойчивого состояния вида в сообществе. Популяции становятся регрессивными в тех случаях, когда старые растения прекращают плодоношение, либо условия в сообществе препятствуют развитию подроста. Помимо перечисленных вариантов в нарушенных лесных сообществах популяции могут быть представлены отдельными особями некоторых возрастных состояний (фрагментарный спектр популяции). Обычно это свидетельствует об эпизодическом приживании вида при крайне низком уровне численности, и свойственно популяциям видов-асеектаторов. Перспективы развития таких популяций оценить очень трудно. Анализ возрастного состава популяций наряду с учетом биологических и экологических особенностей вида позволяет дать прогноз дальнейшего развития популяций, а также позволяет подойти к оценке сукцессионного состояния сообщества (см. Сукцессия).
Лит: Ценопопуляции растений: Основные понятия и структура. - М., 1976; Ценопопуляции растений (Очерки популяционной биологии). — М., 1988.
www.derev-grad.ru
8.3.2. Возрастная структура популяций. Общая экология
8.3.2. Возрастная структура популяций
С возрастом требования особи к среде и устойчивость к отдельным ее факторам закономерно и весьма существенно изменяются. На разных стадиях онтогенеза могут происходить смена сред обитания, изменение типа питания, характера передвижения, общей активности организмов. Нередко возрастные экологические различия в пределах вида выражены в значительно большей степени, чем различия между видами. Травяные лягушки на суше и их головастики в водоемах, гусеницы, грызущие листья, и крылатые бабочки, сосущие нектар, сидячие морские лилии и их планктонные личинки долиолярии – всего лишь разные онтогенетические стадии одних и тех же видов. Возрастные различия в образе жизни часто приводят к тому, что отдельные функции целиком выполняются на определенной стадии развития. Например, многие виды насекомых с полным превращением не питаются в имагинальном состоянии. Рост и питание осуществляются на личиночных стадиях, тогда как взрослые особи выполняют только функции расселения и размножения.
Возрастные различия в популяции существенно усиливают ее экологическую неоднородность и, следовательно, сопротивляемость среде. Повышается вероятность того, что при сильных отклонениях условий от нормы в популяции сохранится хотя бы часть жизнеспособных особей и она сможет продолжить свое существование. Возрастная структура популяций имеет приспособительный характер. Она формируется на основе биологических свойств вида, но всегда отражает также силу воздействия факторов окружающей среды.
Возрастная структура популяций у растений. У растений возрастная структура ценопопуляции, т. е. популяции конкретного фитоценоза, определяется соотношением возрастных групп. Абсолютный, или календарный, возраст растения и его возрастное состояние – понятия не тождественные. Растения одного календарного возраста могут находиться в разных возрастных состояниях. Возрастное, или онтогенетические состояние особи– это этап ее онтогенеза, на котором она характеризуется определенными отношениями со средой. Полный онтогенез, или большой жизненный цикл растений, включает все этапы развития особи – от возникновения зародыша до ее смерти или до полного отмирания всех поколений ее вегетативно возникшего потомства (рис. 97).

Рис. 97. Возрастные состояния овсяницы луговой (А), василька сибирского (Б):
р– проростки; j – ювенильные растения; im – имматурные; v – вирги-нильные; g1 – молодые генеративные; g2 – средневозрастные генеративные; g3 – старые генеративные; ss – субсенильные; s – сенильные
Проростки имеют смешанное питание за счет запасных веществ семени и собственной ассимиляции. Это маленькие растения, для которых характерно наличие зародышевых структур: семядолей, начавшего расти зародышевого корня и, как правило, одноосного побега с небольшими листьями, имеющими часто более простую форму, чем у взрослых растений.
Ювенильные растения переходят к самостоятельному питанию. У них отсутствуют семядоли, но организация еще проста, часто сохраняется одноосность и листья иной формы и меньшего размера, чем у взрослых.
Имматурные растения имеют признаки и свойства, переходные от ювенильных растений к взрослым вегетативным. У них часто начинается ветвление побега, что приводит к увеличению фотосинтетического аппарата.
У взрослых вегетативных растений появляются черты типичной для вида жизненной формы в структуре подземных и наземных органов и строение вегетативного тела принципиально соответствует генеративному состоянию, но репродуктивные органы пока отсутствуют.
Переход растений в генеративный период определяется не только появлением цветков и плодов, но и глубокой внутренней биохимической и физиологической перестройкой организма. В генеративном периоде в растениях безвременника великолепного примерно вдвое больше колхамина и вдвое меньше колхицина, чем в молодых и старых вегетативных особях; у свербиги восточной резко повышается содержание всех форм фосфорных соединений, а также активность каталазы, интенсивность фотосинтеза и транспирации; у жабрицы порезниковой содержание РНК увеличивается в 2 раза, а общего азота – в 5 раз.
Молодые генеративные растения зацветают, образуют плоды, происходит окончательное формообразование взрослых структур. В отдельные годы могут быть перерывы в цветении.
Средневозрастные генеративные растения обычно достигают наибольшей мощности, имеют наибольший ежегодный прирост и семенную продукцию, также могут иметь перерыв в цветении. В этом возрастном состоянии у клонообразующих видов часто начинает проявляться дезинтеграция особей, возникают клоны.
Старые генеративные растения характеризуются резким снижением репродуктивной функции, ослаблением процессов побего– и корнеобразования. Процессы отмирания начинают преобладать над процессами новообразования, усиливается дезинтеграция.
Старые вегетативные (субсенилъные) растения характеризуются прекращением плодоношения, снижением мощности, усилением деструктивных процессов, ослаблением связи между побеговыми и корневыми системами, возможно упрощение жизненной формы, появление листьев имматурного типа.
Сенилъные растения характеризуются крайней дряхлостью, уменьшением размеров, при возобновлении реализуются немногие почки, вторично появляются некоторые ювенильные черты (форма листьев, характер побегов и т. д.).
Отмирающие особи – крайняя степень выражения сенильного состояния, когда у растения остаются живыми лишь некоторые ткани и в отдельных случаях – покоящиеся почки, которые не могут развить надземные побеги.
У некоторых деревьев (дуба черешчатого, бука лесного, клена полевого и др.) описано квазисенильное возрастное состояние (термин предложен Т. А. Работновым). Это угнетенные, низкорослые растения, описанные как торчки (рис. 98). Они приобретают со временем черты старого вегетативного растения, так и не пройдя генеративную фазу.

Рис. 98. Онтогенез дуба черешчатого в благоприятных условиях (сверху) и при недостатке света (по О. В. Смирновой, 1998)
Распределение особей ценопопуляции по возрастным состояниям называется ее возрастным, или онтогенетическим спектром. Он отражает количественные отношения разных возрастных уровней.
Для определения численности каждой возрастной группы у разных видов используют различные счетные единицы. Счетной единицей могут быть отдельные особи, если в течение всего онтогенеза они остаются пространственно обособленными (у однолетников, стержнекорневых моно– и поликарпических трав, многих деревьев и кустарников) или являются четко разграниченными частями клона. У длиннокорневищных и корнеотпрысковых растений счетной единицей могут быть парциальные побеги или парциальные кусты, так как при физической целостности подземной сферы они оказываются нередко физиологически разобщенными, что установлено, например, для ландыша майского при использовании радиоактивных изотопов фосфора. У плотнодерновинных злаков (щучка, типчак, ковыли, змеевка и др.) счетной единицей наряду с молодыми особями может быть компактный клон, который в отношениях со средой выступает как единое целое.
Число семян в почвенном запасе, хотя этот показатель и очень важен, при построении возрастного спектра ценопопуляции обычно не учитывается, так как подсчет их очень трудоемок и получить статистически надежные величины практически невозможно.
Если в возрастном спектре ценопопуляции в момент ее наблюдения представлены только семена или молодые особи, ее называют инвазионной. Такая ценопопуляция не способна к самоподдержанию, и существование ее зависит от поступления зачатков извне. Часто это молодая ценопопуляция, только что внедрившаяся в биоценоз. Если ценопопуляция представлена всеми или почти всеми возрастными группами (некоторые возрастные состояния у конкретных видов могут быть не выражены, например, имматурное, субсенильное, ювенильное), то она называется нормальной. Такая популяция независима и способна к самоподдержанию семенным или вегетативным путем. В ней могут преобладать те или иные возрастные группы. В связи с этим различают молодые, средневозрастные и старые нормальные ценопопуляции.
Нормальная ценопопуляция, состоящая из особей всех возрастных групп, называется полночленной, а если особи каких-либо возрастных состояний отсутствуют (в неблагоприятные годы временно могут выпадать отдельные возрастные группы), то популяция называется нормальной неполночленной.
Регрессивная ценопопуляция представлена только сенильными и субсенильными или также генеративными, но старыми, не образующими всхожих семян. Такая ценопопуляция не способна к самоподдержанию и зависит от заноса зачатков извне.
Инвазионная ценопопуляция может перейти в нормальную, а нормальная – в регрессивную.
Возрастная структура ценопопуляции во многом определяется биологическими особенностями вида: периодичностью плодоношения, числом продуцируемых семян и вегетативных зачатков, способностью вегетативных зачатков к омоложению, скоростью перехода особей из одного возрастного состояния в другое, способностью образовывать клоны и др. Типичный возрастной спектр называется базовым (рис. 99). Проявление всех этих биологических особенностей, в свою очередь, зависит от условий внешней среды. Меняется и ход онтогенеза, который может протекать у одного вида во многих вариантах (поливариантность онтогенеза), что сказывается на структуре возрастного спектра ценопопуляции (рис. 100).

Рис. 99. Базовый тип спектра ценонопуляций (по Л. Б. Зауголыювой, 1976) А – бурачок ленский; Б – анабазис безлистный; В – овсяница луговая; Г – типчак.
1 – базовый спектр; 2 – пределы изменения базового спектра
Разные размеры растений отражают различную жизненность особей в пределах каждой возрастной группы. Жизненность особи проявляется в мощности ее вегетативных и генеративных органов, что соответствует количеству накопленной энергии, и в устойчивости к неблагоприятным воздействиям, что определяется способностью к регенерации. Жизненность каждой особи меняется в онтогенезе по одновершинной кривой, возрастая на восходящей ветви онтогенеза и уменьшаясь на нисходящей. У многих видов особи одного возрастного состояния в одной ценопопуляции могут иметь разную жизненность. Эта дифференциация особей по жизненности может вызываться разнокачественностью семян, разными сроками их прорастания, микроусловиями среды, воздействием животных и человека, конкурентными отношениями. Высокая жизненность может сохраняться до смерти особи во всех возрастных состояниях или снизиться в ходе онтогенеза. Растения высокого уровня жизненности нередко проходят все возрастные состояния ускоренными темпами. В ценопопуляциях часто преобладают растения среднего уровня жизненности. Одни из них проходят онтогенез полностью, а другие пропускают часть возрастных состояний, переходя перед отмиранием на более низкий уровень жизненности. Растения низшего уровня жизненности имеют сокращенный онтогенез и часто переходят в сенильное состояние, едва приступив к цветению.

Рис. 100. Варианты развития ежи сборной в разных экологических условиях (по Л. А. Жуковой, 1985). Латинскими буквами обозначены возрастные состояния растений, а пунктирными линиями – их возможная последовательность
Особи одной ценопопуляции могут развиваться и переходить из одного возрастного состояния в другое с разной скоростью. По сравнению с нормальным развитием, когда возрастные состояния сменяют друг друга в обычной последовательности, может наблюдаться ускорение или задержка в развитии, выпадение отдельных возрастных состояний или целых периодов, наступление вторичного покоя, часть особей может омолаживаться или отмирать. Многие луговые, лесные, степные виды при выращивании их в питомниках или посевах, т. е. на лучшем агротехническом фоне, сокращают свой онтогенез, например овсяница луговая и ежа сборная – с 20–25 до 4 лет, горицвет весенний – со 100 до 10–15 лет, жабрица порезниковая – с 10–18 до 2 лет. У других растений при улучшении условий онтогенез может удлиняться, как, например, у тмина обыкновенного.
В засушливые годы и при усилении выпаса у степного вида овсеца Шелля отдельные возрастные состояния выпадают. Например, взрослые вегетативные особи могут сразу пополнять группу субсенильных, реже – старых генеративных. Клубне-луковичные растения безвременника великолепного в центральных частях компактных клонов, где условия менее благоприятные (хуже освещение, увлажнение, минеральное питание, проявляется токсическое действие отмерших остатков), быстрее переходят в сенильное состояние, чем периферические особи. У свербиги восточной при усиленной пастбищной нагрузке, когда повреждаются почки возобновления, молодые и зрелые генеративные особи могут иметь перерывы в цветении, тем самым как бы омолаживаясь и продлевая свой онтогенез.
У ежи сборной в разных условиях реализуется от 1–2 до 35 путей онтогенеза, а у подорожника большого от 2–4 до 100. Возможность менять путь онтогенеза обеспечивает адаптацию к меняющимся условиям среды и расширяет экологическую нишу вида.
У двух видов степного овсеца – Шелля и опушенного – в Пензенской области четко прослежено циклическое изменение возрастных спектров в многолетней динамике. В засушливые годы популяции овсецов стареют, а во влажные – молодеют. Флюктуации возрастного спектра ценопопуляций вслед за погодными условиями особенно характерны для растений поемных лугов.
Возрастной спектр может варьировать не только вследствие внешних условий, но и в зависимости от реактивности и устойчивости самих видов. К выпасу у растений разная сопротивляемость: у одних выпас вызывает омолаживание, так как растения отмирают, не достигнув старости (например, у полыни равнинной), у других – способствует старению ценопопуляции изза снижения возобновления (например, у степного вида жабрицы Ледебура).
У некоторых видов на всем протяжении ареала в широком диапазоне условий нормальные ценопопуляции сохраняют основные черты возрастной структуры (ясень обыкновенный, типчак, овсяница луговая и др.). Такой возрастной спектр зависит преимущественно от биологических свойств вида. В нем сохраняются прежде всего соотношения во взрослой, наиболее стабильной части. Число вновь возникающих и отмирающих особей в каждой возрастной группе сбалансировано, и общий спектр оказывается постоянным до значительных изменений условий существования. Такие базовые спектры чаще всего имеют ценопопуляции видов-эдификаторов в устойчивых сообществах. Им противопоставляются ценопопуляции, относительно быстро меняющие возрастной спектр вследствие не установившихся отношений с окружающей средой.
Чем крупнее особь, тем значительнее сфера и степень ее воздействия на среду и на соседние растения («фитогенное поле», по А. А. Уранову). Если в возрастном спектре ценопопуляции преобладают особи взрослые вегетативные, молодые и средневозрастные генеративные, то вся популяция в целом будет занимать более прочное положение среди других.
Таким образом, не только численность, но и возрастной спектр ценопопуляции отражает состояние и приспособленность ее к меняющимся условиям внешней среды и определяет позиции вида в биоценозе.
Возрастная структура популяций у животных. В зависимости от особенностей размножения члены популяции могут принадлежать к одной генерации или к разным. В первом случае все особи близки по возрасту и примерно одновременно проходят очередные этапы жизненного цикла. Примером может служить размножение многих видов нестадных саранчовых. Весной из яиц, перезимовавших в кубышках, отложенных в землю, появляются личинки первого возраста. Отрождение личинок несколько растягивается под влиянием микроклиматических и иных условий, но в целом протекает довольно дружно. В это время популяция состоит только из молодых насекомых. Через 2–3 недели в связи с неравномерностью развития отдельных особей в ней могут одновременно встречаться личинки смежных возрастов, но постепенно вся популяция переходит в имагинальное состояние и к концу лета состоит только из взрослых половозрелых форм. К зиме, отложив яйца, они погибают. Такова же возрастная структура популяций у дубовой листовертки, слизней рода Deroceras и других видов с однолетним циклом развития, размножающихся один раз в жизни. Сроки размножения и прохождения отдельных возрастных стадий обычно приурочены к определенному сезону года. Численность таких популяций, как правило, неустойчива: сильные отклонения условий от оптимума на любой стадии жизненного цикла действуют сразу на всю популяцию, вызывая значительную смертность.
Виды с одновременным существованием различных генераций можно разделить на две группы: размножающиеся один раз в жизни и размножающиеся многократно.
У майских жуков, например, самки вскоре после откладки яиц весной погибают. Личинки развиваются в почве и окукливаются на четвертый год жизни. Одновременно в популяции присутствуют представители четырех генераций, каждая из которых появляется через год после предыдущей. Ежегодно завершает свой жизненный цикл одна генерация и появляется новая. Возрастные группы в такой популяции разделяются четким интервалом. Соотношение их по численности зависит от того, насколько благоприятными оказались условия при появлении и развитии очередного поколения. Например, генерация может оказаться малочисленной, если поздние заморозки погубят часть яиц или холодная дождливая погода помешает лету и копуляции жуков.

Рис. 101. Соотношение возрастных групп сельди за 14 лет. «Урожайные» поколения прослеживаются в течение нескольких лет (по Ф. Швердпфегеру, 1963)
У видов с однократным размножением и короткими жизненными циклами в течение года сменяется несколько поколений. Одновременное существование разных генераций обусловлено растянутостью откладки яиц, роста и полового созревания отдельных особей. Это происходит как в результате наследственной неоднородности членов популяции, так и под влиянием микроклиматических и других условий. Например, у свекловичной моли, вредящей сахарной свекле в южных районах СССР, зимуют гусеницы разных возрастов и куколки. За лето развивается 4–5 генераций. Одновременно встречаются представители двух и даже трех смежных поколений, но одно из них, очередное по срокам, всегда преобладает.

Рис. 102. Возрастная структура популяций у животных (по Ю. Одуму, 1975; В. Ф. Осадчих и Е. А. Яблонской, 1968):
А – общая схема, Б – лабораторные популяции полевки Microtus agrestis, В – сезонные изменения соотношения возрастных групп моллюска Adaena vitrea в Северном Каспии.
Различная штриховка – разные возрастные группы:
1– растущая, 2 – стабильная, 3 – сокращающаяся популяции
Еще сложнее возрастная структура популяций у видов с повторным размножением (рис. 101, 102). При этом возможны две крайние ситуации: 1) продолжительность жизни во взрослом состоянии невелика и 2) взрослые живут долго и размножаются многократно. В первом случае ежегодно сменяется значительная часть популяции. Численность ее неустойчива и может резко изменяться в отдельные годы, благоприятные или неблагоприятные для очередного поколения. Возрастная структура популяции сильно варьирует.
У полевки-экономки возрастная структура популяции за летний сезон постепенно усложняется. Сначала население состоит лишь из особей прошлого года рождения, затем добавляются молодые первого и второго пометов. К периоду появления третьего и четвертого приплодов наступает половая зрелость у представителей первых двух и в популяцию вливаются генерации внучатого поколения. Осенью популяция состоит преимущественно из разновозрастных особей текущего года рождения, так как старшие погибают.
Во втором случае возникает относительно устойчивая структура популяции, с длительным сосуществованием различных поколений. Так, индийские слоны достигают половой зрелости к 8-12 годам и живут до 60–70 лет. Самка рождает одного, реже двух слонят, примерно раз в четыре года. В стаде обычно взрослые животные разного возраста составляют около 80 %, молодняк – около 20 %. У видов с более высокой плодовитостью соотношение возрастных групп может быть иным, но общая структура популяции всегда остается достаточно сложной, включая представителей разных поколений и их разновозрастное потомство. Колебания численности таких видов происходят в небольших пределах.
Длительно размножающуюся часть популяции часто называют запасом. От размеров популяционного запаса зависят возможности восстановления численности. Та часть молодых, которые достигают половой зрелости и увеличивают запас, – это ежегодное пополнение популяции. У видов с одновременным существованием только одной генерации запас практически равен нулю и размножение осуществляется целиком за счет пополнения. Виды со сложной возрастной структурой характеризуются значительной величиной запаса и небольшой, но стабильной долей пополнения.
При эксплуатации человеком природных популяций животных учет их возрастной структуры имеет важнейшее значение (рис. 103). У видов с ежегодным большим пополнением можно изымать более значительную часть популяции без угрозы подорвать ее численность. Если же уничтожить много взрослых в популяции со сложной возрастной структурой, то это сильно замедлит ее восстановление. Например, у горбуши, созревающей на второй год жизни, возможен вылов до 50–60 % нерестящихся особей без угрозы дальнейшего снижения численности популяции. Для кеты, созревающей позднее и имеющей более сложную возрастную структуру, нормы изъятия из половозрелого стада должны быть меньше.

Рис. 103. Возрастная структура таймырской популяции дикого северного оленя в период умеренного (А) и чрезмерного (Б) охотничьего промысла (по А. А. Колпащикову, 2000)
Анализ возрастной структуры помогает прогнозировать численность популяции на протяжении жизни ряда ближайших поколений. Такие анализы широко применяют, например, в рыбном хозяйстве для предвидения динамики промысловых стад. Используют довольно сложные математические модели с количественным выражением воздействия на отдельные возрастные группы всех поддающихся учету факторов среды. Если выбранные показатели возрастной структуры совершенно правильно отражают реальное влияние среды на природную популяцию, получают высокодостоверные прогнозы, позволяющие планировать вылов на ряд лет вперед.
Поделитесь на страничкеСледующая глава >
bio.wikireading.ru
Глава 8. Популяции
8.1. Понятие о популяции в экологии
Популяцией в экологии называют группу особей одного вида, находящихся во
Поддержание оптимальной в данных условиях численности называют гомеостазом популяции. Гомеостатические возможности популяций по-разному выражены у различных видов. Осуществляются они также через взаимоотношения особей.
Таким образом, популяции, как групповые объединения, обладают рядом специфических свойств, которые не присущи каждой отдельно взятой особи.
Основные характеристики популяций:
1) численность– общее количество особей на выделяемой территории;
2) плотность популяции – среднее число особей на единицу площади или объема занимаемого популяцией пространства; плотность популяции можно выражать также через массу членов популяции в единице пространства;
3) рождаемость– число новых особей, появившихся за единицу времени в результате размножения;
4) смертность – показатель, отражающий количество погибших в популяции особей за определенный отрезок времени;
5) прирост популяции– разница между рождаемостью и смертностью; прирост может быть как положительным, так и отрицательным;
6) темп роста – средний прирост за единицу времени.
Популяции свойственна определенная организация. Распределение особей по территории, соотношения групп по полу, возрасту, морфологическим, физиологическим, поведенческим и генетическим особенностям отражают структуру популяции.
8.3.1. Половая структура популяций
Экологические и поведенческие различия между особями мужского и женского пола могут быть сильно выражены. Например, самцы комаров семейства Culicidae, в отличие от кровососущих самок, в имагинальный период либо не питаются совсем, либо ограничиваются слизыванием росы, либо потребляют нектар растений. Но даже если образ жизни самцов и самок сходен, они различаются по многим физиологическим признакам: темпам роста, срокам полового созревания, устойчивостью к изменениям температуры, голоданию и т. п.
Особенно наглядно влияние условий среды на половую структуру популяций у видов с чередованием половых и партеногенетических поколений. Дафнии Daphnia magna при оптимальной температуре размножаются партеногенетически, но при повышенной или пониженной температуре в популяциях появляются самцы. На появление обоеполого поколения у тлей могут влиять изменения длины светового дня, температуры, увеличение плотности населения и другие факторы.
У некоторых видов пол изначально определяется не генетическими, а экологическими факторами. Так, у растений Arisaema japonica пол зависит от накопления запасов питательных веществ в клубнях. Из крупных клубней вырастают экземпляры с женскими цветками, из мелких – с мужскими.
8.3.2. Возрастная структура популяций
С возрастом требования особи к среде и устойчивость к отдельным ее факторам закономерно и весьма существенно изменяются. На разных стадиях онтогенеза могут происходить смена сред обитания, изменение типа питания, характера передвижения, общей активности организмов.
Возрастная структура популяций имеет приспособительный характер. Она формируется на основе биологических свойств вида, но всегда отражает также силу воздействия факторов окружающей среды.
Возрастная структура популяций у растений. У растений возрастная структура ценопопуляции, т. е. популяции конкретного фитоценоза, определяется соотношением возрастных групп. Абсолютный, или календарный, возраст растения и его возрастное состояние – понятия не тождественные. Растения одного календарного возраста могут находиться в разных возрастных состояниях.Возрастное, или онтогенетическиесостояние особи– это этап ее онтогенеза, на котором она характеризуется определенными отношениями со средой. Полный онтогенез, или большой жизненный цикл растений, включает все этапы развития особи – от возникновения зародыша до ее смерти или до полного отмирания всех поколений ее вегетативно возникшего потомства (рис. 97).
Рис. 97. Возрастные состояния овсяницы луговой (А), василька сибирского (Б):
р– проростки; j– ювенильные растения;im– имматурные;v– вирги-нильные;g1 – молодые генеративные;g2– средневозрастные генеративные;g3– старые генеративные;ss– субсенильные;s– сенильные
Проростки имеют смешанное питание за счет запасных веществ семени и собственной ассимиляции. Это маленькие растения, для которых характерно наличие зародышевых структур: семядолей, начавшего расти зародышевого корня и, как правило, одноосного побега с небольшими листьями, имеющими часто более простую форму, чем у взрослых растений.
Ювенильные растения переходят к самостоятельному питанию. У них отсутствуют семядоли, но организация еще проста, часто сохраняется одноосность и листья иной формы и меньшего размера, чем у взрослых.
Имматурные растения имеют признаки и свойства, переходные от ювенильных растений к взрослым вегетативным. У них часто начинается ветвление побега, что приводит к увеличению фотосинтетического аппарата.
У взрослых вегетативных растений появляются черты типичной для вида жизненной формы в структуре подземных и наземных органов и строение вегетативного тела принципиально соответствует генеративному состоянию, но репродуктивные органы пока отсутствуют.
Молодые генеративные растения зацветают, образуют плоды, происходит окончательное формообразование взрослых структур. В отдельные годы могут быть перерывы в цветении.
Средневозрастные генеративные растения обычно достигают наибольшей мощности, имеют наибольший ежегодный прирост и семенную продукцию, также могут иметь перерыв в цветении. В этом возрастном состоянии у клонообразующих видов часто начинает проявляться дезинтеграция особей, возникают клоны.
Старые генеративные растения характеризуются резким снижением репродуктивной функции, ослаблением процессов побего– и корнеобразования. Процессы отмирания начинают преобладать над процессами новообразования, усиливается дезинтеграция.
Старые вегетативные (субсенилъные) растения характеризуются прекращением плодоношения, снижением мощности, усилением деструктивных процессов, ослаблением связи между побеговыми и корневыми системами, возможно упрощение жизненной формы, появление листьев имматурного типа.
Сенилъные растения характеризуются крайней дряхлостью, уменьшением размеров, при возобновлении реализуются немногие почки, вторично появляются некоторые ювенильные черты (форма листьев, характер побегов и т. д.).
Отмирающие особи– крайняя степень выражения сенильного состояния, когда у растения остаются живыми лишь некоторые ткани и в отдельных случаях – покоящиеся почки, которые не могут развить надземные побеги.
Распределение особей ценопопуляции по возрастным состояниям называется ее возрастным, или онтогенетическим спектром. Он отражает количественные отношения разных возрастных уровней.
Если в возрастном спектре ценопопуляции в момент ее наблюдения представлены только семена или молодые особи, ее называют инвазионной. Такая ценопопуляция не способна к самоподдержанию, и существование ее зависит от поступления зачатков извне. Если ценопопуляция представлена всеми или почти всеми возрастными группами (некоторые возрастные состояния у конкретных видов могут быть не выражены, например, имматурное, субсенильное, ювенильное), то она называетсянормальной. Такая популяция независима и способна к самоподдержанию семенным или вегетативным путем. В ней могут преобладать те или иные возрастные группы. В связи с этим различают молодые, средневозрастные и старые нормальные ценопопуляции.
Нормальная ценопопуляция, состоящая из особей всех возрастных групп, называется полночленной, а если особи каких-либо возрастных состояний отсутствуют (в неблагоприятные годы временно могут выпадать отдельные возрастные группы), то популяция называетсянормальной неполночленной.
Регрессивная ценопопуляция представлена только сенильными и субсенильными или также генеративными, но старыми, не образующими всхожих семян. Такая ценопопуляция не способна к самоподдержанию и зависит от заноса зачатков извне.
Инвазионная ценопопуляция может перейти в нормальную, а нормальная – в регрессивную.
Возрастная структура популяций у животных.
Виды с одновременным существованием различных генераций можно разделить на две группы: размножающиеся один раз в жизни и размножающиеся многократно.
У майских жуков, например, самки вскоре после откладки яиц весной погибают. Личинки развиваются в почве и окукливаются на четвертый год жизни. Одновременно в популяции присутствуют представители четырех генераций, каждая из которых появляется через год после предыдущей. Ежегодно завершает свой жизненный цикл одна генерация и появляется новая. Возрастные группы в такой популяции разделяются четким интервалом.
У видов с однократным размножением и короткими жизненными циклами в течение года сменяется несколько поколений. Одновременное существование разных генераций обусловлено растянутостью откладки яиц, роста и полового созревания отдельных особей.
При эксплуатации человеком природных популяций животных учет их возрастной структуры имеет важнейшее значение (рис. 103). У видов с ежегодным большим пополнением можно изымать более значительную часть популяции без угрозы подорвать ее численность. Если же уничтожить много взрослых в популяции со сложной возрастной структурой, то это сильно замедлит ее восстановление. Например, у горбуши, созревающей на второй год жизни, возможен вылов до 50–60 % нерестящихся особей без угрозы дальнейшего снижения численности популяции. Для кеты, созревающей позднее и имеющей более сложную возрастную структуру, нормы изъятия из половозрелого стада должны быть меньше.
Анализ возрастной структуры помогает прогнозировать численность популяции на протяжении жизни ряда ближайших поколений. Такие анализы широко применяют, например, в рыбном хозяйстве для предвидения динамики промысловых стад.
studfiles.net
Возрастная структура популяций
С возрастом требования особи к среде и устойчивость к отдельным ее факторам закономерно и весьма существенно изменяются. На разных стадиях онтогенеза могут происходить смена сред обитания, изменение типа питания, характера передвижения, общей активности организмов. Нередко возрастные экологические различия в пределах вида выражены в значительно большей степени, чем различия между видами. Травяные лягушки на суше и их головастики в водоемах, гусеницы, грызущие листья, и крылатые бабочки, сосущие нектар, сидячие морские лилии и их планктонные личинки долиолярии – всего лишь разные онтогенетические стадии одних и тех же видов. Возрастные различия в образе жизни часто приводят к тому, что отдельные функции целиком выполняются на определенной стадии развития. Например, многие виды насекомых с полным превращением не питаются в имагинальном состоянии. Рост и питание осуществляются на личиночных стадиях, тогда как взрослые особи выполняют только функции расселения и размножения.
Возрастные различия в популяции существенно усиливают ее экологическую неоднородность и, следовательно, сопротивляемость среде. Повышается вероятность того, что при сильных отклонениях условий от нормы в популяции сохранится хотя бы часть жизнеспособных особей и она сможет продолжить свое существование. Возрастная структура популяций имеет приспособительный характер. Она формируется на основе биологических свойств вида, но всегда отражает также силу воздействия факторов окружающей среды.
Возрастная структура популяций у растений.У растений возрастная структура ценопопуляции, т. е. популяции конкретного фитоценоза, определяется соотношением возрастных групп. Абсолютный, или календарный, возраст растения и его возрастное состояние – понятия не тождественные. Растения одного календарного возраста могут находиться в разных возрастных состояниях. Возрастное, или онтогенетические состояние особи–это этап ее онтогенеза, на котором она характеризуется определенными отношениями со средой. Полный онтогенез, или большой жизненный цикл растений, включает все этапы развития особи – от возникновения зародыша до ее смерти или до полного отмирания всех поколений ее вегетативно возникшего потомства (рис. 97).

Рис. 97. Возрастные состояния овсяницы луговой (А), василька сибирского (Б):
р– проростки; j – ювенильные растения; im – имматурные; v – вирги-нильные; g1 – молодые генеративные; g2 – средневозрастные генеративные; g3 – старые генеративные; ss – субсенильные; s – сенильные
Проростки имеют смешанное питание за счет запасных веществ семени и собственной ассимиляции. Это маленькие растения, для которых характерно наличие зародышевых структур: семядолей, начавшего расти зародышевого корня и, как правило, одноосного побега с небольшими листьями, имеющими часто более простую форму, чем у взрослых растений.
Ювенильные растения переходят к самостоятельному питанию. У них отсутствуют семядоли, но организация еще проста, часто сохраняется одноосность и листья иной формы и меньшего размера, чем у взрослых.
Имматурные растения имеют признаки и свойства, переходные от ювенильных растений к взрослым вегетативным. У них часто начинается ветвление побега, что приводит к увеличению фотосинтетического аппарата.
У взрослых вегетативных растений появляются черты типичной для вида жизненной формы в структуре подземных и наземных органов и строение вегетативного тела принципиально соответствует генеративному состоянию, но репродуктивные органы пока отсутствуют.
Переход растений в генеративный период определяется не только появлением цветков и плодов, но и глубокой внутренней биохимической и физиологической перестройкой организма. В генеративном периоде в растениях безвременника великолепного примерно вдвое больше колхамина и вдвое меньше колхицина, чем в молодых и старых вегетативных особях; у свербиги восточной резко повышается содержание всех форм фосфорных соединений, а также активность каталазы, интенсивность фотосинтеза и транспирации; у жабрицы порезниковой содержание РНК увеличивается в 2 раза, а общего азота – в 5 раз.
Молодые генеративные растения зацветают, образуют плоды, происходит окончательное формообразование взрослых структур. В отдельные годы могут быть перерывы в цветении.
Средневозрастные генеративные растения обычно достигают наибольшей мощности, имеют наибольший ежегодный прирост и семенную продукцию, также могут иметь перерыв в цветении. В этом возрастном состоянии у клонообразующих видов часто начинает проявляться дезинтеграция особей, возникают клоны.
Старые генеративные растения характеризуются резким снижением репродуктивной функции, ослаблением процессов побего– и корнеобразования. Процессы отмирания начинают преобладать над процессами новообразования, усиливается дезинтеграция.
Старые вегетативные (субсенилъные) растения характеризуются прекращением плодоношения, снижением мощности, усилением деструктивных процессов, ослаблением связи между побеговыми и корневыми системами, возможно упрощение жизненной формы, появление листьев имматурного типа.
Сенилъные растения характеризуются крайней дряхлостью, уменьшением размеров, при возобновлении реализуются немногие почки, вторично появляются некоторые ювенильные черты (форма листьев, характер побегов и т. д.).
Отмирающие особи – крайняя степень выражения сенильного состояния, когда у растения остаются живыми лишь некоторые ткани и в отдельных случаях – покоящиеся почки, которые не могут развить надземные побеги.
У некоторых деревьев (дуба черешчатого, бука лесного, клена полевого и др.) описано квазисенильное возрастное состояние (термин предложен Т. А. Работновым). Это угнетенные, низкорослые растения, описанные как торчки (рис. 98). Они приобретают со временем черты старого вегетативного растения, так и не пройдя генеративную фазу.

Рис. 98. Онтогенез дуба черешчатого в благоприятных условиях (сверху) и при недостатке света (по О. В. Смирновой, 1998)
Распределение особей ценопопуляции по возрастным состояниям называется ее возрастным, или онтогенетическим спектром. Он отражает количественные отношения разных возрастных уровней.
Для определения численности каждой возрастной группы у разных видов используют различные счетные единицы. Счетной единицей могут быть отдельные особи, если в течение всего онтогенеза они остаются пространственно обособленными (у однолетников, стержнекорневых моно– и поликарпических трав, многих деревьев и кустарников) или являются четко разграниченными частями клона. У длиннокорневищных и корнеотпрысковых растений счетной единицей могут быть парциальные побеги или парциальные кусты, так как при физической целостности подземной сферы они оказываются нередко физиологически разобщенными, что установлено, например, для ландыша майского при использовании радиоактивных изотопов фосфора. У плотнодерновинных злаков (щучка, типчак, ковыли, змеевка и др.) счетной единицей наряду с молодыми особями может быть компактный клон, который в отношениях со средой выступает как единое целое.
Число семян в почвенном запасе, хотя этот показатель и очень важен, при построении возрастного спектра ценопопуляции обычно не учитывается, так как подсчет их очень трудоемок и получить статистически надежные величины практически невозможно.
Если в возрастном спектре ценопопуляции в момент ее наблюдения представлены только семена или молодые особи, ее называют инвазионной. Такая ценопопуляция не способна к самоподдержанию, и существование ее зависит от поступления зачатков извне. Часто это молодая ценопопуляция, только что внедрившаяся в биоценоз. Если ценопопуляция представлена всеми или почти всеми возрастными группами (некоторые возрастные состояния у конкретных видов могут быть не выражены, например, имматурное, субсенильное, ювенильное), то она называется нормальной. Такая популяция независима и способна к самоподдержанию семенным или вегетативным путем. В ней могут преобладать те или иные возрастные группы. В связи с этим различают молодые, средневозрастные и старые нормальные ценопопуляции.
Нормальная ценопопуляция, состоящая из особей всех возрастных групп, называется полночленной, а если особи каких-либо возрастных состояний отсутствуют (в неблагоприятные годы временно могут выпадать отдельные возрастные группы), то популяция называется нормальной неполночленной.
Регрессивная ценопопуляция представлена только сенильными и субсенильными или также генеративными, но старыми, не образующими всхожих семян. Такая ценопопуляция не способна к самоподдержанию и зависит от заноса зачатков извне.
Инвазионная ценопопуляция может перейти в нормальную, а нормальная – в регрессивную.
Возрастная структура ценопопуляции во многом определяется биологическими особенностями вида: периодичностью плодоношения, числом продуцируемых семян и вегетативных зачатков, способностью вегетативных зачатков к омоложению, скоростью перехода особей из одного возрастного состояния в другое, способностью образовывать клоны и др. Типичный возрастной спектр называется базовым (рис. 99). Проявление всех этих биологических особенностей, в свою очередь, зависит от условий внешней среды. Меняется и ход онтогенеза, который может протекать у одного вида во многих вариантах (поливариантность онтогенеза), что сказывается на структуре возрастного спектра ценопопуляции (рис. 100).

Рис. 99. Базовый тип спектра ценонопуляций (по Л. Б. Зауголыювой, 1976) А – бурачок ленский; Б – анабазис безлистный; В – овсяница луговая; Г – типчак.
1 – базовый спектр; 2 – пределы изменения базового спектра
Разные размеры растений отражают различную жизненность особей в пределах каждой возрастной группы. Жизненность особи проявляется в мощности ее вегетативных и генеративных органов, что соответствует количеству накопленной энергии, и в устойчивости к неблагоприятным воздействиям, что определяется способностью к регенерации. Жизненность каждой особи меняется в онтогенезе по одновершинной кривой, возрастая на восходящей ветви онтогенеза и уменьшаясь на нисходящей. У многих видов особи одного возрастного состояния в одной ценопопуляции могут иметь разную жизненность. Эта дифференциация особей по жизненности может вызываться разнокачественностью семян, разными сроками их прорастания, микроусловиями среды, воздействием животных и человека, конкурентными отношениями. Высокая жизненность может сохраняться до смерти особи во всех возрастных состояниях или снизиться в ходе онтогенеза. Растения высокого уровня жизненности нередко проходят все возрастные состояния ускоренными темпами. В ценопопуляциях часто преобладают растения среднего уровня жизненности. Одни из них проходят онтогенез полностью, а другие пропускают часть возрастных состояний, переходя перед отмиранием на более низкий уровень жизненности. Растения низшего уровня жизненности имеют сокращенный онтогенез и часто переходят в сенильное состояние, едва приступив к цветению.

Рис. 100. Варианты развития ежи сборной в разных экологических условиях (по Л. А. Жуковой, 1985). Латинскими буквами обозначены возрастные состояния растений, а пунктирными линиями – их возможная последовательность
Особи одной ценопопуляции могут развиваться и переходить из одного возрастного состояния в другое с разной скоростью. По сравнению с нормальным развитием, когда возрастные состояния сменяют друг друга в обычной последовательности, может наблюдаться ускорение или задержка в развитии, выпадение отдельных возрастных состояний или целых периодов, наступление вторичного покоя, часть особей может омолаживаться или отмирать. Многие луговые, лесные, степные виды при выращивании их в питомниках или посевах, т. е. на лучшем агротехническом фоне, сокращают свой онтогенез, например овсяница луговая и ежа сборная – с 20–25 до 4 лет, горицвет весенний – со 100 до 10–15 лет, жабрица порезниковая – с 10–18 до 2 лет. У других растений при улучшении условий онтогенез может удлиняться, как, например, у тмина обыкновенного.
В засушливые годы и при усилении выпаса у степного вида овсеца Шелля отдельные возрастные состояния выпадают. Например, взрослые вегетативные особи могут сразу пополнять группу субсенильных, реже – старых генеративных. Клубне-луковичные растения безвременника великолепного в центральных частях компактных клонов, где условия менее благоприятные (хуже освещение, увлажнение, минеральное питание, проявляется токсическое действие отмерших остатков), быстрее переходят в сенильное состояние, чем периферические особи. У свербиги восточной при усиленной пастбищной нагрузке, когда повреждаются почки возобновления, молодые и зрелые генеративные особи могут иметь перерывы в цветении, тем самым как бы омолаживаясь и продлевая свой онтогенез.
У ежи сборной в разных условиях реализуется от 1–2 до 35 путей онтогенеза, а у подорожника большого от 2–4 до 100. Возможность менять путь онтогенеза обеспечивает адаптацию к меняющимся условиям среды и расширяет экологическую нишу вида.
У двух видов степного овсеца – Шелля и опушенного – в Пензенской области четко прослежено циклическое изменение возрастных спектров в многолетней динамике. В засушливые годы популяции овсецов стареют, а во влажные – молодеют. Флюктуации возрастного спектра ценопопуляций вслед за погодными условиями особенно характерны для растений поемных лугов.
Возрастной спектр может варьировать не только вследствие внешних условий, но и в зависимости от реактивности и устойчивости самих видов. К выпасу у растений разная сопротивляемость: у одних выпас вызывает омолаживание, так как растения отмирают, не достигнув старости (например, у полыни равнинной), у других – способствует старению ценопопуляции изза снижения возобновления (например, у степного вида жабрицы Ледебура).
У некоторых видов на всем протяжении ареала в широком диапазоне условий нормальные ценопопуляции сохраняют основные черты возрастной структуры (ясень обыкновенный, типчак, овсяница луговая и др.). Такой возрастной спектр зависит преимущественно от биологических свойств вида. В нем сохраняются прежде всего соотношения во взрослой, наиболее стабильной части. Число вновь возникающих и отмирающих особей в каждой возрастной группе сбалансировано, и общий спектр оказывается постоянным до значительных изменений условий существования. Такие базовые спектры чаще всего имеют ценопопуляции видов-эдификаторов в устойчивых сообществах. Им противопоставляются ценопопуляции, относительно быстро меняющие возрастной спектр вследствие не установившихся отношений с окружающей средой.
Чем крупнее особь, тем значительнее сфера и степень ее воздействия на среду и на соседние растения («фитогенное поле», по А. А. Уранову). Если в возрастном спектре ценопопуляции преобладают особи взрослые вегетативные, молодые и средневозрастные генеративные, то вся популяция в целом будет занимать более прочное положение среди других.
Таким образом, не только численность, но и возрастной спектр ценопопуляции отражает состояние и приспособленность ее к меняющимся условиям внешней среды и определяет позиции вида в биоценозе.
Возрастная структура популяций у животных.В зависимости от особенностей размножения члены популяции могут принадлежать к одной генерации или к разным. В первом случае все особи близки по возрасту и примерно одновременно проходят очередные этапы жизненного цикла. Примером может служить размножение многих видов нестадных саранчовых. Весной из яиц, перезимовавших в кубышках, отложенных в землю, появляются личинки первого возраста. Отрождение личинок несколько растягивается под влиянием микроклиматических и иных условий, но в целом протекает довольно дружно. В это время популяция состоит только из молодых насекомых. Через 2–3 недели в связи с неравномерностью развития отдельных особей в ней могут одновременно встречаться личинки смежных возрастов, но постепенно вся популяция переходит в имагинальное состояние и к концу лета состоит только из взрослых половозрелых форм. К зиме, отложив яйца, они погибают. Такова же возрастная структура популяций у дубовой листовертки, слизней рода Deroceras и других видов с однолетним циклом развития, размножающихся один раз в жизни. Сроки размножения и прохождения отдельных возрастных стадий обычно приурочены к определенному сезону года. Численность таких популяций, как правило, неустойчива: сильные отклонения условий от оптимума на любой стадии жизненного цикла действуют сразу на всю популяцию, вызывая значительную смертность.
Виды с одновременным существованием различных генераций можно разделить на две группы: размножающиеся один раз в жизни и размножающиеся многократно.
У майских жуков, например, самки вскоре после откладки яиц весной погибают. Личинки развиваются в почве и окукливаются на четвертый год жизни. Одновременно в популяции присутствуют представители четырех генераций, каждая из которых появляется через год после предыдущей. Ежегодно завершает свой жизненный цикл одна генерация и появляется новая. Возрастные группы в такой популяции разделяются четким интервалом. Соотношение их по численности зависит от того, насколько благоприятными оказались условия при появлении и развитии очередного поколения. Например, генерация может оказаться малочисленной, если поздние заморозки погубят часть яиц или холодная дождливая погода помешает лету и копуляции жуков.

Рис. 101. Соотношение возрастных групп сельди за 14 лет. «Урожайные» поколения прослеживаются в течение нескольких лет (по Ф. Швердпфегеру, 1963)
У видов с однократным размножением и короткими жизненными циклами в течение года сменяется несколько поколений. Одновременное существование разных генераций обусловлено растянутостью откладки яиц, роста и полового созревания отдельных особей. Это происходит как в результате наследственной неоднородности членов популяции, так и под влиянием микроклиматических и других условий. Например, у свекловичной моли, вредящей сахарной свекле в южных районах СССР, зимуют гусеницы разных возрастов и куколки. За лето развивается 4–5 генераций. Одновременно встречаются представители двух и даже трех смежных поколений, но одно из них, очередное по срокам, всегда преобладает.

Рис. 102. Возрастная структура популяций у животных (по Ю. Одуму, 1975; В. Ф. Осадчих и Е. А. Яблонской, 1968):
А – общая схема, Б – лабораторные популяции полевки Microtus agrestis, В – сезонные изменения соотношения возрастных групп моллюска Adaena vitrea в Северном Каспии.
Различная штриховка – разные возрастные группы:
1– растущая, 2 – стабильная, 3 – сокращающаяся популяции
Еще сложнее возрастная структура популяций у видов с повторным размножением (рис. 101, 102). При этом возможны две крайние ситуации: 1) продолжительность жизни во взрослом состоянии невелика и 2) взрослые живут долго и размножаются многократно. В первом случае ежегодно сменяется значительная часть популяции. Численность ее неустойчива и может резко изменяться в отдельные годы, благоприятные или неблагоприятные для очередного поколения. Возрастная структура популяции сильно варьирует.
У полевки-экономки возрастная структура популяции за летний сезон постепенно усложняется. Сначала население состоит лишь из особей прошлого года рождения, затем добавляются молодые первого и второго пометов. К периоду появления третьего и четвертого приплодов наступает половая зрелость у представителей первых двух и в популяцию вливаются генерации внучатого поколения. Осенью популяция состоит преимущественно из разновозрастных особей текущего года рождения, так как старшие погибают.
Во втором случае возникает относительно устойчивая структура популяции, с длительным сосуществованием различных поколений. Так, индийские слоны достигают половой зрелости к 8-12 годам и живут до 60–70 лет. Самка рождает одного, реже двух слонят, примерно раз в четыре года. В стаде обычно взрослые животные разного возраста составляют около 80 %, молодняк – около 20 %. У видов с более высокой плодовитостью соотношение возрастных групп может быть иным, но общая структура популяции всегда остается достаточно сложной, включая представителей разных поколений и их разновозрастное потомство. Колебания численности таких видов происходят в небольших пределах.
Длительно размножающуюся часть популяции часто называют запасом. От размеров популяционного запаса зависят возможности восстановления численности. Та часть молодых, которые достигают половой зрелости и увеличивают запас, – это ежегодное пополнение популяции. У видов с одновременным существованием только одной генерации запас практически равен нулю и размножение осуществляется целиком за счет пополнения. Виды со сложной возрастной структурой характеризуются значительной величиной запаса и небольшой, но стабильной долей пополнения.
При эксплуатации человеком природных популяций животных учет их возрастной структуры имеет важнейшее значение (рис. 103). У видов с ежегодным большим пополнением можно изымать более значительную часть популяции без угрозы подорвать ее численность. Если же уничтожить много взрослых в популяции со сложной возрастной структурой, то это сильно замедлит ее восстановление. Например, у горбуши, созревающей на второй год жизни, возможен вылов до 50–60 % нерестящихся особей без угрозы дальнейшего снижения численности популяции. Для кеты, созревающей позднее и имеющей более сложную возрастную структуру, нормы изъятия из половозрелого стада должны быть меньше.

Рис. 103. Возрастная структура таймырской популяции дикого северного оленя в период умеренного (А) и чрезмерного (Б) охотничьего промысла (по А. А. Колпащикову, 2000)
Анализ возрастной структуры помогает прогнозировать численность популяции на протяжении жизни ряда ближайших поколений. Такие анализы широко применяют, например, в рыбном хозяйстве для предвидения динамики промысловых стад. Используют довольно сложные математические модели с количественным выражением воздействия на отдельные возрастные группы всех поддающихся учету факторов среды. Если выбранные показатели возрастной структуры совершенно правильно отражают реальное влияние среды на природную популяцию, получают высокодостоверные прогнозы, позволяющие планировать вылов на ряд лет вперед.
lektsia.com
Возрастной состав популяции
Возрастная структура (возрастной состав) популяции — соотношение в популяции особей разных возрастных групп.[ ...]
Возрастной состав популяции имеет большое значение для ее существования и процветания. При благоприятных условиях в популяции наличествуют все возрастные группы и поддерживается более или менее стабильный возрастной состав. В быстро растущих популяциях доминируют интенсивно размножающиеся молодые особи, а в сокращающихся — старые, уже неспособные интенсивно размножаться. Такие популяции малопродуктивны, недостаточно устойчивы. Возрастной состав популяции зависит от продолжительности жизни особей, периода достижения ими половой зрелости, числа генераций.[ ...]
ВОЗРАСТНОЙ СОСТАВ ПОПУЛЯЦИИ (В.с.п.) - соотношение численности разных когорт, т. е. групп особей разного возраста. При постоянном «популяционном потоке», когда часть особей вымирает, а часть занимает освободившееся место, популяция представлена особями разного возраста. В быстро растущих популяциях обычно велика доля молоди, в популяциях, численность которых сокращается, — доля взрослых и стареющих особей. Популяции растений по соотношению когорт разделяют на инвазионные (состоящие в основном из молодых особей), нормальные (более или менее равномерно представленные всеми когортами) и регрессивные (с преобладанием когорты старых особей).[ ...]
Возрастной состав популяции выражают в процентах особей (из общей выборки), отнесенных к той или иной возрастной пли онтогенетической группе, зафиксированной во всей выборке, взятой в данном биотопе. Морфофизиологическпе показатели, в том числе н массу тела, выражают средними максимальными и минимальным величинами для выборки из данного биотока. Кроме того, отмечают процент особей, имеющих показатели выше и ниже средних оптимальных для данного вида или географической популяции. Количество развивающихся или диаиаузирующнх особей в выборке выражают в процентах от общего числа.[ ...]
| Возрастной состав популяции дуба белого (Quercus alba) в спелом дубово-гикориевом лесу. Распределение числа деревьев по возрастным классам с 50-летним интервалом показывает уменьшение этого числа по мере перехода от молодых возрастов к старым. График в линейном масштабе {слева) представляет собой убывающую J-образную кривую возрастного состава. В логарифмическом масштабе (справа) распределение близко к прямой линии, что позволяет предполагать, что константная часть деревьев отмирает при переходе из одного возрастного класса в другой. Распределения получены по измерениям диаметров 1525 стволов деревьев на площади 74 га; интерпретация возрастных классов выполнена по меньшему объему данных (56 деревьев). (Данные R. S. Miller, «Illinois Nat. | 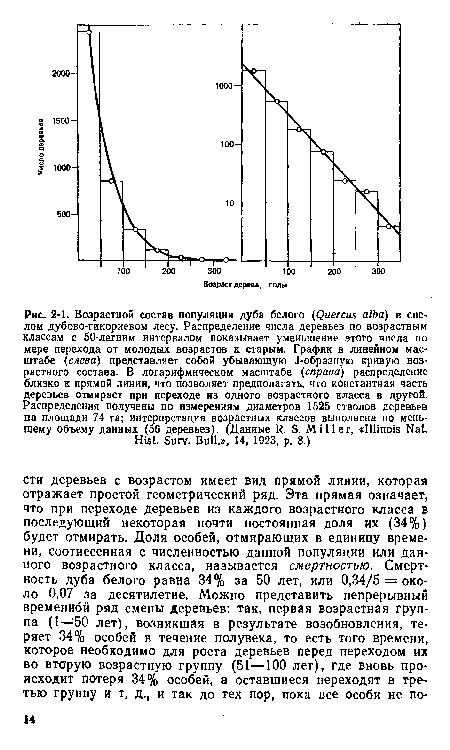 |
| Возрастной состав популяций Acacia burkittii в двух местностях в Южной Австралии. (По Crisp, Lange, 1976.) |  |
| 45 |  |
В сокращающихся популяциях преобладают старые особи, которые уже не способны интенсивно размножаться. Данная возрастная структура свидетельствует о неблагоприятных условиях. В быстро растущих популяциях преобладают интенсивно размножающиеся молодые особи. В стабильных популяциях это соотношение, как правило, составляет 1:1. При благоприятных условиях в популяции имеются все возрастные группы и поддерживается сравнительно стабильный уровень численности. На возрастной состав популяции, помимо общей продолжительности жизни, влияют длительность периода размножения, число генераций в сезон, плодовитость и смертность разных возрастных групп. Так, у полевок (СМеЛпопошув) взрослые особи могут давать потомство три раза в год и более, а молодые особи способны размножаться через 2—3 месяца (рис. 9.7).[ ...]
На разных фазах динамики складывается своеобразный возрастной состав популяции. На фазах спада численности и депрессии он наиболее однотипен, а на фазах подъема численности и массового размножения (эпифитотий) наиболее многообразен. На каждом этане онтогенеза существенно различаются реакции особей вида иа среду, т. е. диапазон оптимальных пределов состояния факторов среды. Так, у полевок молодые особи слабо приспособлены к перенесению низких температур но сравнению со взрослыми, но они лучше переносят засуху и питание недостаточно влажным кормом. В итоге общий диапазон переживания популяцией неблагоприятных условий значительно расширяется: зимние условия переживают взрослые, а летнюю засуху — молодые.[ ...]
Е. В. Бурмакин и Л. А. Жаков (1961) высказали предположение, что в тех озерах, где, кроме окуня, нет других рыб, возрастной состав популяции окуня приобретает описанный выше необычный вид под влиянием каннибализма. Старшие особи поедают младших, формируя тем самым одно или несколько поколений с сильно уменьшенной численностью. Когда эти малочисленные поколения достигают возраста, при котором каннибализм является единственным способом добывания пищи, пресс хищников на младшие возрастные группы существенно уменьшается и снова появляются условия для существования многочисленного поколения. Таким образом возникает циклическая смена многочисленных и малочисленных поколений с периодом порядка продолжительности жизни окуня. Именно проверка этой гипотезы и послужила первоначальным поводом к построению модели популяции окуня в виде кибернетической системы.[ ...]
Развитие и рост (рис. 62). В последние годы численность щуки неуклонно снижается. В уловах 1998-1999 гг. было обследовано 79 экземпляров. Большую часть составляли особи в возрасте 3+ - 5+. Табл. 45 отображает возрастной состав популяции в процентах.[ ...]
ru-ecology.info










