
Строение клеточной стенки продуктов растительного происхождения. Строение клеточной стенки растений
Биология для студентов - 39. Строение и свойства клеточных стенок растительных клеток и бактерий
Клеточная стенка — оболочка клетки, расположенная снаружи от цитоплазматической мембраны и выполняющая структурные, защитные и транспортные функции. Обнаруживается у большинства бактерий, грибов и растений. Животные и многие простейшие не имеют клеточной стенки.
Клеточные стенки бактерий состоят из пептидогликана (муреина) и бывают двух типов:
- грамположительного,
- грамотрицательного.
Клеточная стенка грамположительного типа состоит исключительно из толстого слоя пептидогликана, плотно прилегающего к клеточной мембране и пронизанного тейхоевыми и липотейхоевыми кислотами.
При грамотрицательном типе слой пептидогликана существенно тоньше, между ним и плазматической мембраной находится периплазматическое пространство, а снаружи клетка окружена ещё одной мембраной, представленной липополисахаридоми, являющаяся пирогенным эндотоксином грамотрицательных бактерий.
Клеточные стенки грибов состоят из хитина и глюканов.
Большинство водорослей имеют клеточную стенку из целлюлозы и различных гликопротеинов. Включения дополнительных полисахаридов имеют большое таксономическое значение. Диатомовые водоросли синтезируют свою клеточную стенку из кремнезёма.
Важнейшей отличительной особенностью растительной клетки является наличие прочной клеточной стенки, основным компонентом которой является целлюлоза. Клеточная стенка высших растений представляет собой сложноорганизованный, преимущественно полимерный, внеклеточный матрикс, окружающий каждую клетку. Растительная клетка, лишённая клеточной стенки, обозначается термином протопласт. В клеточных стенках растений существуют углубления — поры, через которые проходят цитоплазматические канальца — плазмодесмы, осуществляющие контакт соседних клеток и обмен веществами между ними.
Химический состав и пространственная организация полимеров клеточной стенки отличаются у разных видов, клеток разных тканей одного растения и иногда у разных частей стенки вокруг одного протопласта.
Кроме того, строение клеточной стенки изменяется в онтогенезе растительного организма.
Первичная клеточная стенка формируется при делении и сохраняется во время роста клетки. Формирование вторичной клеточной стенки происходит с внутренней стороны от первичной стенки и связано с окончанием роста и специализацией (дифференцировкой) клеток растения. Снаружи от первичной клеточной стенки, между первичными стенками двух соседних клеток, располагается срединная пластинка (состоит преимущественно из кальциевых и магниевых солей пектиновых веществ).
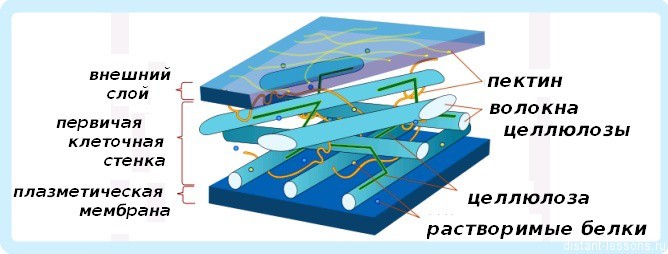
Первичная клеточная стенка высших растений состоит из трёх взаимодействующих, но структурно независимых трехмерных сетей полимеров:
- Основная сеть состоит из фибрилл целлюлозы и связывающих их гемицеллюлоз (или сшивочных гликанов).
- Вторая сеть состоит из пектиновых веществ.
- Третья сеть представлена, как правило, структурными белками клеточной стенки.
Также следует отметить, что у растений клады commelinids (группа в системах APG) и у представителей семейства Маревыепервичная клеточная стенка содержит значительное количество ароматических веществ (гидроксикоричные кислоты, главным образом феруловая и п-кумаровая). При этом у представителей клады commelinids гидроксикоричные кислоты присоединяются к сшивочным гликанам (к глюкуроноарабиноксиланам), а у семейства Маревые к пектиновым веществам (к рамногалактуронанам I).
Растительные клеточные стенки выполняют целый ряд функций:
- они обеспечивают жесткость клетки для структурной и механической поддержки,
- придают форму клетке, направление её роста и в конечном счете морфологию всему растению.
Клеточная стенка также противодействует тургору, то есть осмотическому давлению, когда дополнительное количество воды поступает в растения. Клеточные стенки защищают от патогенов, проникающих из окружающей среды, и запасают углеводы для растения.
vseobiology.ru
Функции, строение, химический состав и рост клеточной стенки.
Клеточная оболочка — типичный компонент растительной клетки, является продуктом жизнедеятельности протопласта.
Функции:
1. Прочные и жесткие клеточные оболочки, служат механической опорой для органов растения.
2. Оболочка ограничивает растяжение протопласта вакуолью, а размер и форма зрелой клетки перестают изменяться.
3. В наружных тканях клеточные оболочки, защищают лежащие глубже клетки от высыхания.
4. По клеточным стенкам, примыкающим к друг другу, могут передвигаться различные вещества и вода от клетки к клетке (путь через апопласт).
5. Они оказывают влияние на поглощение, транспирацию и секрецию.
Клеточные стенки, как правило, бесцветны и легко пропускают солнечный свет. Стенки соседних клеток скреплены пектиновой срединной пластинкой. Срединная пластинка — единый слой, общий для двух соседних клеток. Она представляет собой несколько видоизмененную клеточную пластинку, возникшую в процессе цитокинеза. Срединная пластинка менее обводнена, в ней могут присутствовать молекулы лигнина. Углы клеточных стенок в результате внутриклеточного давления могут округляться, и между соседними клетками образуются межклетники. Все стенки клеток растения, связанные одна с другой и примыкающие к заполненным водой межклетникам, обеспечивают существование сплошной обводненной среды, в которой свободно передвигаются водорастворимые вещества.
Строение и химический состав.
Первичная клеточная стенка.
Первоначально кнаружи от плазмалеммы возникает первичная клеточная стенка.
Состав: целлюлоза, гемицеллюлоза, пектин и вода.
Первичные клеточные стенки соседних клеток соединены протопектиновой срединной пластинкой. В клеточной стенке линейные очень длинные (несколько микрон) молекулы целлюлозы, состоящие из глюкозы, собраны в пучки - мицеллы, которые, в свою очередь, объединяются в микрофибриллы – тончайшие (1,5…4 нм) волоконца неопределенной длины, а затем в макрофибриллы. Целлюлоза образует многомерный каркас, который погружен в аморфный сильно обводненный матрикс из нецеллюлозных углеводов: пектинов, гемицеллюлоз и др. Именно целлюлоза обеспечивает прочность клеточной стенки. Микрофибриллы эластичны и по прочности на разрыв сходны со сталью. Полисахариды матрикса определяют такие свойства стенки, как высокая проницаемость для воды, растворенных мелких молекул и ионов, сильная набухаемость. Благодаря матриксу по стенкам, примыкающим к друг другу, могут передвигаться вода и вещества от клетки к клетке (путь через апопласт по «свободному пространству»). Некоторые гемицеллюлозы могут откладываться в стенках клеток семян в качестве запасных веществ.
Рост стенки.
При делении клеток создается заново лишь клеточная пластинка. На нее обе дочерние клетки откладывают собственные стенки, состоящие главным образом из гемицеллюлозы. При этом образование стенки происходит и на внутренней поверхности остальных стенок, принадлежащих материнской клетке. Клеточная пластинка преобразуется в срединную, она обычно тонка и почти неразличима. После деления клетка вступает в фазу растяжения за счет поглощения клеткой воды и роста центральной вакуоли. Тургорное давление растягивает стенку, в которую внедряются мицеллы целлюлозы и вещества матрикса. Такой способ роста носит название интуссусцепции, внедрения. Оболочки делящихся и растущих клеток называют первичными. Они содержат воды до 90 %, в сухом веществе преобладают полисахариды матрикса: у двудольных пектины и гемицеллюлозы в равном соотношении, у однодольных – в основном гемицеллюлозы; содержание целлюлозы не превышает 30 %. Толщина первичной стенки не более 0,1…0,5 мкм.
К моменту, когда рост клетки заканчивается, рост клеточной стенки может продолжаться, но уже в толщину. Этот процесс носит название вторичного утолщения. При этом на внутренней поверхности первичной клеточной стенки откладывается вторичная клеточная стенка. Рост вторичной клеточной стенки происходит в результате аппозиции, наложения новых мицелл целлюлозы на внутреннюю поверхность клеточной стенки. Таким образом, наиболее молодые слои клеточной стенки ближе всего к плазмаллеме.
Для некоторых типов клеток (многие волокна, трахеиды, членики сосудов) образование вторичной стенки – основная функция протопласта, после завершения вторичного утолщения он отмирает. Однако это не обязательно. Вторичная стенка выполняет главным образом механические, опорные функции. В ее составе значительно меньше воды и преобладают микрофибриллы целлюлозы (40…50 % сухого вещества). Во вторичных стенках волокон льна и волосков хлопчатника содержание целлюлозы может достигать 95 %.
Механизм построения клеточной стенки. Клеточная стенка образуется в результате деятельности протопласта. В соответствии с этим вещества поступают в стенку изнутри, со стороны протопласта. Строительные материалы – молекулы целлюлозы пектина, лигнина и других веществ — накапливаются и частично синтезируются в цистернах аппарата Гольджи. Упакованные в пузырьки аппарата Гольджи, они транспортируются к плазмалемме. Разорвав ее, пузырек лопается, и содержимое его оказывается снаружи плазмалеммы. Мембрана пузырька восстанавливает целостность плазмалеммы. Благодаря ферментной активности плазмалеммы идет сборка фибрилл целлюлозы строение клеточной стенки. Образуемые плазмалеммой фибриллы накладываются изнутри, не переплетаясь. В их ориентации большая роль принадлежит микротрубочкам, располагающимся под плазмалеммой параллельно формирующимся фибриллам.
2. Поры. Видоизменения клеточной стенки.
Поры. При образовании первичной клеточной стенки в ней выделяются более тонкие участки, где фибриллы целлюлозы лежат более рыхло. Канальцы эндоплазматической цепи проходят здесь через клеточные стенки, соединяя соседние клетки. Эти участки называются первичными поровыми полями, а канальцы эндоплазматической сети, проходящие в них, — плазмодесмами.
Рост в толщину происходит у клеточной стенки неравномерно, неутолщенными остаются небольшие участки первичной клеточной стенки в местах расположения первичных поровых полей (поровых каналов). Поровые каналы двух соседних клеток располагаются обычно друг против друга и разделяются замыкающей пленкой поры — двумя первичными клеточными стенками с межклеточным веществом между ними. В пленке сохраняются субмикроскопические отверстия, через которые проходят плазмодесмы. Таким образом, пора — это два поровых канала и замыкающая пленка между ними.
Плазмодесмы пронизывают замыкающие пленки пор. В каждой клетке имеется от нескольких сотен до десятков тысяч плазмодесм. Плазмодесмы встречаются только - в растительных клетках, там, где имеются твердые клеточные стенки. Плазмодесмы образуются из канальцев ЭР, которые остаются в клеточной пластинке между двумя дочерними клетками. При воссоздании ЭР обеих клеток они оказываются соединенными через плазмодесмы.
Плазмодесма проходит через плазмодесменный канал в замыкающей пленке поры. Плазмалемма, выстилающая канал, и гиалоплазма между ней и плазмодесмой непрерывны с плазмалеммами и гиалоплазмами смежных клеток. Таким образом, протопласты соседних клеток связаны между собой каналами плазмодесм и плазмодесмами. По ним происходит межклеточный транспорт ионов и молекул, а также гормонов. Объединенные плазмодесмами протопласты клеток в растении образуют единое целое — симпласт. Транспорт веществ через плазмодесмы получил название симпластического в отличие от апопластического транспорта по клеточным стенкам и межклетникам.
В процессе жизнедеятельности клетки целлюлозная клеточная стенка может претерпевать видоизменения.
Не нашли то, что искали? Воспользуйтесь поиском гугл на сайте:
zdamsam.ru
Строение клеточной стенки продуктов растительного происхождения.
Клетка любого организма, представляет собой целостную живую систему. Она состоит из трех неразрывно связанных между собой частей: оболочки, цитоплазмы и ядра. Оболочка клетка осуществляет непосредственное взаимодействие с внешней средой и взаимодействие с соседними клетками (в многоклеточных организмах).
Оболочка клеток. Оболочка клеток имеет сложное строение. Она состоит из наружного слоя и расположенной под ним плазматической мембраны. Основными составляющими клеточной стенки можно выделить 3 компонента:
- структурные компоненты (целлюлоза)
- компоненты матрикса (наполнители оболочки) – ГМЦ, пектиновые вещества, белки (экстенсин), липиды;
- инкрустаторы клеточной стенки (выстилающие ее изнутри) – лигнин.
Срединная пластинка представляет собой тонкий слой пектиновых веществ (пектаты магния и кальция), главным образом протопектина.
Клеточная оболочка окружает собственно клетку со всех сторон и служит связующим звеном между ней и соседними клетками, обеспечивая, таким образом, единство и целостность всего растительного организма. В жестких оболочках растительных клеток образуются каналы, в которых располагаются тончайшие тяжи цитоплазмы – плазмодесмы. Благодаря этому, осуществляются межклеточные взаимодействия. Иными словами, у растений клеточные оболочки призваны обеспечивать те функции, которые у животных выполняют скелет, кожа и система кровообращения (т.е. опорную, защитную и транспортную.) Не удивительно поэтому, что в ходе эволюции у растений возникли весьма разнообразные по структуре и химическому составу типы клеточных стенок. Собственно говоря, растительные клетки во многом различают и классифицируют именно по форме и природе клеточных стенок.
Оболочка, как правило, бесцветна и прозрачна. Она легко пропускает солнечный свет. Оболочки соседних клеток как бы сцементированы межклеточными веществами, образующими так называемую срединную пластинку. Вследствие этого соседние клетки оказываются отделёнными друг от друга стенкой, образованной двумя оболочками и срединной пластинкой. Это и даёт основание называть оболочку также клеточной стенкой.
Клеточная стенка растительных клеток состоит, главным образом, из полисахаридов. Оболочку эмбриональных тканей и клеток, растущих растяжением, называют первичной. В этот период оболочка достаточно эластична. После прекращения роста клетки изнутри на первичную клеточную стенку начинают откладываться новые слои и образуется вторичная клеточная стенка, придающая клетке жёсткость и прочность, где начинает откладываться инкрустирующее вещество лигнин, образованное производными ароматических соединений. (правда лигнин начинает откладываться еще на ранних стадиях, но все-таки значительный его рост происходит непосредственно во 2 клет оболочке)
Схема строения клеточной стенки:



 соседняя клетка Срединная пластинка Первичная клеточная стенка Вторичная клеточная стенка
соседняя клетка Срединная пластинка Первичная клеточная стенка Вторичная клеточная стенка



 непрерывный матрикс
непрерывный матрикс

 Микрофибриллы целлюлозы
Микрофибриллы целлюлозы


 ГМЦ (гемицеллюлозы)
ГМЦ (гемицеллюлозы)

 Лигнин
Лигнин

 Пектин.вещества
Пектин.вещества
Первичная клеточная стенка состоит из волокон (микрофибрилл) целлюлозы, которые занимают менее 20% объема гидратированной стенки. Располагаясь в клеточных стенках параллельно, целлюлозные волокна образуют мицеллы, которые имеют правильную почти кристаллическую упаковку. Одна мицелла целлюлозы может отстоять от другой на расстоянии, равном ее десяти диаметрам. Пространство между мицеллами целлюлозы заполнено матриксом, состоящим из пектиновых веществ, гемицеллюлоз (ксилоглюкан и арабиногалактан) и структурного белка, связанного с тетрасахаридами.
Первичная стенка клетки рассматривается как целая макромолекула, компоненты которой тесно взаимосвязаны (рис 5.). Между мицеллами целлюлозы и ксилоглюканом имеется значительное количество водородных связей. В свою очередь ксилоглюкан ковалентно связан с пектиновыми веществами через их боковые галактановые цепи. С другой стороны, пектиновые вещества через арабиногалактан ковалентно связаны со структурным белком.

Рисунок 5 - Структура первичной клеточной стенки (по Альберсхейму):
1 – микрофибрила целлюлозы; 2 – ксилоглюкан; 3 – главные рамногалактуроновые цепи пектиновых веществ; 4 – боковые галактановые цепи пектиновых веществ; 5 - структурный белок с арабинозными тетрасахаридами; 6 – арабиногалактан.
ГМЦ – сложный полисахарид, образованный нейтральными и кислыми и нейтральными компонентами, в состав которых входят ксиланы, арабинаны, маннаны и галактаны. Большинство полисахаридов ГМЦ относятся к гетерополимерам, построенным из различных по составу и содержанию моносахаридов несольких видов. Они являются производными пентоз ( ксилоза, арабиноза) и гексоз (глюкоза,галактоза). Значительное количество гидроксильных групп в каждом остатке моносахаридов, формирующих полисахариды ГМЦ, обуславливает их гидрофильность.
Что касается непосредственно всего матрикса в целом, то он построен по принципу взаимопроникающих сеток. Одну из них образуют полисахариды за счет H-H связей, 2 – ГМЦ, связанные ковалентно с пектиновыми веществами через их боковые галактановые цепи. Связь между матриксом и микрофибриллами целлюлозы осуществляется макромолекулами ГМЦ , образующими переходный слой на поверхности элементарных фибрилл и соединенными с молекулами целлюлозы водородными связями. Пектин пронизывает матрикс клетки. Гликопротеины плотно вплетены в ткань клеточной стенки.
 Обязательный компонент растительной клетки – вода, которая способна проникать в матрикс клетки, но не проходящая через кристаллические области микрофибрилл. Количество воды в клеточной стенки зависит от хим природы полисахаридов матрикса и степени их гидрофильности. С увеличением доли гидрофобных соединений в матриксе снижается его гидрофильность меньше воды содержится в клеточной стенке.
Обязательный компонент растительной клетки – вода, которая способна проникать в матрикс клетки, но не проходящая через кристаллические области микрофибрилл. Количество воды в клеточной стенки зависит от хим природы полисахаридов матрикса и степени их гидрофильности. С увеличением доли гидрофобных соединений в матриксе снижается его гидрофильность меньше воды содержится в клеточной стенке.
Изменения при тепловой обработке:
В процессе тепловой кулинарной обработки механическая прочность овощей и плодов уменьшается, изделия приобретают мягкую консистенцию. Это связано в основном с ослаблением связей между клетками в растительной ткани вследствие деструкции срединных пластинок, а также с деструкцией матрикса в клеточной оболочке (ПП, части ГМЦ, экстенсина).
Деструкция протопектина (ПП)



 Разрушение срединных пластинок проходит вследствие расщепление ПП под действием горячей воды с образованием растворимых в воде пектиновых кислот. Гидролиз ПП начинается с 60 град и интенсифицируется с 80 град и выше. Деструкция происходит за счет разрыва пектиновых связей между этарифицированными остатками галактуроновых кислот и хелатных связей (солевые мостики). Если водородные связи можно разрушить добавлением определенного количества воды, то хелатные связи разрушаются в процессе ионобменной реакции:
Разрушение срединных пластинок проходит вследствие расщепление ПП под действием горячей воды с образованием растворимых в воде пектиновых кислот. Гидролиз ПП начинается с 60 град и интенсифицируется с 80 град и выше. Деструкция происходит за счет разрыва пектиновых связей между этарифицированными остатками галактуроновых кислот и хелатных связей (солевые мостики). Если водородные связи можно разрушить добавлением определенного количества воды, то хелатные связи разрушаются в процессе ионобменной реакции:
 СОО COOK (Na)
СОО COOK (Na)

 Са (Мg) +
Са (Мg) +  (
( ) +
) + 



 COO COOK (Na)
COO COOK (Na)
Поскольку это обратимая, то, чтобы она протекала с разрушением солевых мостиков, необходимо выводить ионы кальция из реакции, превращая их в малорастворимые или нерастворимые соли ( к примеру, крупу предварительно отваривают в воде, а потом уже начинается варка каши в молоке. Если сначала варить крупу в молоке, то хелатные связи укрепляются)
Чем больше содержание органических кислот, тем интенсивнее деструкция в ПП.
Содержание ГМЦ и белка экстенсина в результате тепловой оболочке также уменьшается (деструкция). Их продукты растворимы в воде (в результате этого происходит разрыхление матрикса и уменьшение механ прочности). Деструкция ГМЦ (70 – 80 град), экстенсин – 50 гард.
Клеточные оболочки после тепловой обработке не разрушаются, несмотря на их разрыхление.
foodteor.ru
Структура клеточных стенок растений
Клеточные стенки растений обладают необычайной прочностью, и в процессе роста растения меняют свою структуру и состав. Основными компонентами клеточных стенок растений являются полисахариды, причем среди них преобладает целлюлоза, которая в значительной мере определяет архитектуру стенки.
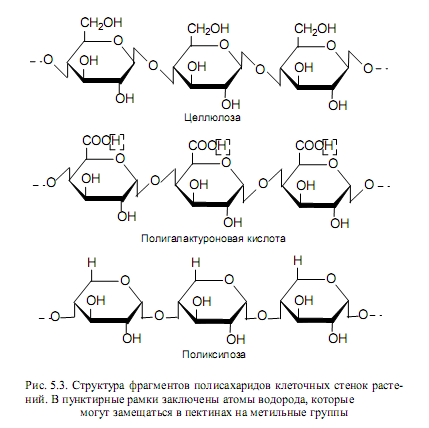
Целлюлоза. Этот гомополисахарид является самым распространенным на Земле углеводом (растения образуют в год до 1011 т целлюлозы). Мономерами целлюлозы служат остатки глюкозы, соединенные в длинные цепочки (до 10 000 остатков глюкозы в каждой) с помощью b(1→4)-гликозидных связей (рис. 5.3). В такой молекуле отсутствует полная свобода вращения вокруг 1С—О- и О— 4С-связей, и полимер приобретает конформацию, благоприятную для образования межцепочечных водородных связей, в случае, когда цепочки располагаются антипараллельно. В результате молекулы целлюлозы объединяются в микрофибриллы толщиной примерно от 10 до 25 нм. Микрофибриллы перевиваются и образуют тонкие нити, которые, в свою очередь, могут обматываться одна вокруг другой, как пряди в канате, формируя макрофибриллы. Каждая макрофибрилла имеет толщину около 0,5 мкм и может достигать в длину 6—8 мкм. Прочность макрофибрилл сопоставима с прочностью равной по толщине стальной проволоки. Кроме того, отдельные участки микрофибрилл имеют упорядоченное строение и придают клеточной стенке кристаллические свойства. Таким образом, можно отметить сложность и высокую упорядоченность целлюлозы в составе клеточных стенок, что неслучайно: этот полимер выполняет защитную и опорную функции в растении.
В таком виде полисахариды недоступны действию собственных ферментов, и целлюлоза не может использоваться растением в качестве резервного вещества. Лишь немногие организмы (некоторые бактерии, грибы, простейшие и редкие животные) обладают ферментными системами, способными расщеплять целлюлозу.
Микро- и макрофибриллы целлюлозы в клеточной стенке растений погружены в матрикс, который также состоит в основном из полисахаридов и меняет свою структуру в процессе роста растения. На начальных этапах развития матрикс состоит из пектиновых веществ, а в дальнейшем в нем появляются ксиланы и различные нейтральные полисахариды («гемицеллюлоза»). Пектиновые вещества представляют собой полимеры a-галактуроновой кислоты, в которых некоторые водородные атомы замещены метильными группами (-СН3) (рис. 5.3). Ксиланы представляют собой полимеры ксилозы (рис. 5.3).
На более поздних этапах развития, когда происходит одревеснение клеточных стенок, в клетках откладывается лигнин — химически устойчивый полимер, содержащий большое число ароматических колец. Кроме этого, в составе клеточных стенок растений обнаруживаются небольшие количества гликопротеинов, нерастворимых липидных полимеров различного строения и восков.
Клеточные стенки некоторых растений содержат редкие полисахариды, имеющие необычное строение. Например, в стенках и межклеточном
веществе морских красных водорослей содержится сложный гетерополисахарид агар, представляющий собой смесь сульфатированных полисахаридов —агарозы и агаропектина. Агароза построена из чередующихся остатков D-галактозы и 3,6-ангидро-L-лактозы, связанных попеременно b(1→4)- и a(1→3)-связями. Агаропектин имеет более сложное строение: в его состав входят D-галактоза, 3,6-ангидрогалактоза, уроновые кислоты и сульфат. Агар используется в качестве наиболее распространенного уплотнителя для твердых сред, незаменимых в микробиологии, а также в пищевой промышленности для желирования продуктов. Следует отметить, что подавляющее большинство микроорганизмов не способно расщеплять агар, и это одно из главных его преимуществ перед другим уплотнителем питательных сред — желатиной. Агароза находит широкое применение в биохимических исследованиях: она в водной среде образует гель с большими порами, размер которых определяется ее концентрацией. Агарозные гели используются для фрак-ционирования белков и нуклеиновых кислот, а также для иммобилизации клеток.
Похожие статьи:
poznayka.org
Клеточные стенки строение - Справочник химика 21
ГЕМИЦЕЛЛЮЛОЗЫ — полисахариды, входящие в состав клеточной стенки растительной ткани наряду с целлюлозой и лигнином. Часть Г. волокнистого строения является устойчивой к действию щелочи. Присутствие Г. в целлюлозной массе ухудшает качество целлюлозы наоборот, в производстве бумаги Г. улучшает процесс размола и механические свойства бумаги. [c.68] Все методы выделения лигнина можно подразделить на две группы методы, основанные на удалении полисахаридов, с получением лигнина в виде нерастворимого остатка методы, основанные на переводе лигнина в раствор с последующим осаждением (получение препаратов растворимых лигнинов). Нерастворимые препараты лигнина, имеющие подобно природному лигнину сетчатую структуру, более или менее сохраняют морфологическое строение клеточной стенки, но, естественно, более рыхлое. Растворимые лигнины после осаждения, очистки и сушки имеют вид порошков. [c.367]Г. Строение клеточных стенок бактерий [c.388]
Клеточная стенка (оболочка клетки) состоит из нескольких слоев, отличающихся друг от друга строением, физическими свойствами и химическим составом. Подробнее всего изучено строение клеточной стенки трахеид (рис. 8.10, а). [c.214]
Функции солода и культур плесневых грибов не ограничиваются осахариванием крахмала, в них входят накопление в сусле достаточного количества органического азота для питания дрожжей и частичное растворение клеточных стенок эндосперма сырья. В осуществлении этих процессов, а также в выращивании солода и плесневых грибов участвуют многочисленные ферменты, поэтому необходимо знание их химической природы, строения и механизма действия. [c.114]
Строение и состав клеточной стенки [c.214]
Целью работы является исследование строения структурных полимеров клеточной стенки мицелиальных грибов и разработка научных основ использования их биомассы для получения ценных в практическом отношении продуктов. [c.162]
Эта схема была подробно рассмотрена в ряде работ. Например, в работе [58] было высказано предположение о возможности протекания этих процессов как в протоплазме, так и в клеточных стенках растений. Последняя точка зрения была признана рядом исследователей, которые указанными выше переходами объясняли наблюдаемые изменения в составе клеточных стенок в процессе их развития. Так, на основе приведенных выше схем объяснялось относительное увеличение содержания в клеточных стенках растений пентозанов, высказывалось предположение о существований в макромолекулах целлюлозы дефектных звеньев, состоявших из остатков глюкуроновой кислоты и пентоз. Эта точка зрения в настоящее время считается неправильной, и объясняется этот вывод различным строением макромолекул полисахаридов, входящих в состав гемицеллюлоз. Эти различия в основном сводятся к следующему [c.331]
Препараты ЛМР считают наиболее пригодными для исследования строения лигнина и моделирования его химических реакций, однако и эти препараты не идентичны с природным лигнином, и, кроме того, их нельзя считать представительными для всего лигнина клеточной стенки. Размол древесины увеличивает доступность лигнина и вызывает его механическую деструкцию. При механической деструкции происходят реакции гомолитического расщепления связей лигнина с гемицеллюлозами и связей С-О-С и С-С в сетке лигнина с возникновением промежуточных свободных радикалов, которые вступают в реакции рекомбинации и инициируют реакции полимеризации и окисления кислородом воздуха. Поэтому ЛМР, хотя и близок к природному лигнину, все же химически изменен. Первоначальную методику Бьеркмана в дальнейшем неоднократно модифицировали. [c.371]
Ранее мы рассмотрели способы, при помощи которых белковые субъединицы могут соединяться друг с другом, образуя замкнутые олигомеры и длинные опирали. Другой чрезвычайно важный способ упаковки белков и липидов приводит к образованию пластинчатых структур, или мембран [1—10], которые с молекулярной точки зрения можно рассматривать как практически безграничные двумерные поверхности. Эта глава посвящена строению, химическим свойствам и функциям биологических мембран, а также клеточных стенок бактерий, грибов и растений. [c.337]
РИС. 5-12. Предполагаемое строение стенки растительных клеток. Относительные размеры различных компонентов стенки не соблюдены, однако соотношения между ними близки к истинным. Расстояние между пучками целлюлозных волокон увеличено, чтобы можно было изобразить соединяющие структуры В первичной клеточной стенке одной клетки может находиться от 10 до 100 пучков целлюлозных волокон [115] [c.396]
Содержимое всех живых клеток отделено от окружающей среды специальными структурами - биомембранами, которые обычно называют прото-плазматическими мембранами. У растений и бактерий наряду с такими мембранами снаружи клетки еще имеется клеточная стенка. Для эукариотических клеток характерно деление внутреннего содержимого клетки на отдельные отсеки, или компартменты. Они представляют собой субклеточные органеллы, ограниченные мембранами, например, ядро митохондрии, аппарат Гольджи. Однако мембраны служат не только поверхностями раздела. По существу, мембраны представляют собой сложные по строению и разнообразные по функциям биохимические системы. [c.106]
В учебнике в достаточно компактной четкой форме излагается на современном уровне обширный по тематике материал. Особое внимание уделено строению макромолекул и физической структуре полимеров как основе для понимания структуры и свойств синтетических полимеров и высокомолекулярных компонентов древесины. Рассмотрены процессы синтеза полимеров, в том числе биосинтеза природных полимеров. Детально излагаются свойства синтетических полимеров, используемых при получении разнообразных материалов и изделий на основе древесины и продуктов ее переработки. Учебник содержит необходимые сведения по анатомии древесины и строению клеточной стенки. Значительное место отводится изложению теоретических основ процессов химической переработки древесины и ее компонентов. [c.2]
Слои вторичной стенки хорошо различимы на микрофотографиях, полученных в поляризованном свете, благодаря различной ориентации в разных слоях микрофибрилл целлюлозы (см. 8.6.2), обладающей вследствие кристаллической структуры двойным лучепреломлением. Слои 8 , 82 и 8з(Т) существенно различаются по толщине 8 и 8з(Т) тонкие, а 8а толстый и образует основную массу клеточной стенки. Во всех этих слоях уже преобладает целлюлоза. Слой 81 имеет толщину 0,1...0,3 мкм в зависимости от части годичного кольца (поздняя или ранняя) и древесной породы. Толщина слоя 82 составляет в среднем 2...6 мкм с колебаниями от 1 мкм (в ранней древесине) до 7...9 мкм (в поздней древесине). Слой 8з(Т) самый тонкий (0,1.. .0,2 мкм) строение его в значительной степени зависит от древесной породы. [c.216]
Полисахариды. В растениях полисахариды участвуют в образовании клеточной стенки (структурные полисахариды) и, кроме того, используются для создания запаса связанного углерода и энергии (резервные полисахариды). Структурные полисахариды формируют срединную пластинку, первичную и вторичные стенки, различающиеся функциональным назначением, строением и составом. Клеточная стенка является природным полимерным композиционным материалом, в котором полимерная дисперсная фаза - микрофибриллы распределена в полимерной дисперсионной среде - углеводной матрице (матриксе). Следовательно, при биосинтезе полисахаридов должна одновременно закладываться и структура клеточной стенки. [c.334]
Строение клеточных стенок волокон либриформа и волокнистых трахеид в древесине лиственных пород примерно такое же, как у трахеид поздней древесины хвойных. Распределение слоев по массе приблизительно следующее у волокон либриформа Р 81 82 83 = 1 10 87 2 у волокнистых трахеид 5 16 67 12. По строению клеточных стенок паренхимные клетки и сосуды отличаются от трахеид и волокон либриформа. У сосудов также существуют первичная Р и вторичная 8 (81 + 82 + 83) [c.216]
Гетерогенные процессы у полисахаридов отличаются от гетерогенных реакций НМС. На характер гетерогенных процессов у полисахаридов, как и других полимеров, влияет их надмолекулярная структура, а у полисахаридов в древесине также ультраструктура клеточной стенки и анатомическое строение древесины. Все эти детали структуры определяют доступность полисахарида для химического реагента. Результаты гетерогенного процесса будут зависеть поэтому не только от скорости самой химической реакции, но и от скорости диффузии реагента в глубь клеточной стенки древесины или в глубь волокна технической целлюлозы. Класси- [c.281]
Все рассмотренные закономерности гидролитической деструкции относятся к влиянию на скорость гидролиза различий в химическом строении полисахаридов. При гидролизе полисахаридов непосредственно в древесине решающее влияние на начальных стадиях гидролиза оказывает доступность полисахаридов, зависящая от ультраструктуры клеточной стенки и, главным образом, от надмолекулярной структуры полисахаридов. [c.290]
Между гелифицированными и фюзенизованными микрокомпонентами нельзя провести резкой границы. Фюзенизация может происходить на начальной стадии изменения растительных тканей или после того, как уже начался процесс гели-фикации. В первом случае получается типичный фюзен, в котором ткани полностью сохраняют клеточное строение. Часто клеточная структура фюзена может быть нарушена из-за разрыва клеточных стенок. [c.74]
Стенка клетки имеет слоистое строение (рис. 71). На поверхности ее расположен липоидный слой с выступами и бугорками, под ним — липополисахарндный слой, ниже неплотно упакованные молекулы белка и прилегающий к цитоплазматической мембране плотный гликопептидный слой. Вся масса пронизана каналами, поэтому клеточная стенка проницаема для солей и других многочисленных молекулярных соединений. Вотличие от нее цито- [c.247]
К настоящему времени многие О-гликозид-гидролазы получены в высокоочищенном и в кристаллическом состоянии, для целого ряда карбогидраз получены данные о первичной структуре (всей белковой молекулы или ее фрагментов). Именно среди кар-богидраз был выбран фермент — лизоцим, для которого впервые в энзимологии было расшифровано пространственное строение с помощью рентгеноструктурных методов анализа. Карбогидразы широко используются для изучения структуры многих биологически важных соединений — гликоконьюгатов, компонентов клеточной стенки и т. д. [c.22]
Строение клетки определяется теми веществами, из которых образованы стенки (мембраны) клетки, представляющие ее каркас, и веществами, содержащимися внутри клеток. Углевод целлюлоза — важнейшая составная часть клеточных стенок растений. В организмах животных основными структурообразующими материалами являются белки. Кроме того, внутриклеточные вещества состоят в значительной мере из белков. Так, красная клетка крови граничена тонкой мембраной, внут- [c.383]
Инфицирование клетки Е. соИ бактериофагом происходит следующим путем фаг впрыскивает свою ДНК через клеточную стенку в цитоплазму. Приблизительно через 20 мин после этого клетка лопается, и из нее выходит около 100 полностью готовых копий исходной вирусной частицы. Такая высокая скорость размножения позволяет проводить в пробирке в течение 20 мин генетические эксперименты, для которых потребовалось бы все население земного шара, если бы эти опыты проводились на людях. Главные принципы, лежащие в основе этого метода, были ясно изложены Бензером [130], который впервые составил карту тонкого строения гена. Частицы бактериофагов, подобно бактериям, можно посеять в чашке с агаром. Отличие заключается лишь в том, что агар должен содержать однородную суспензию бактерий, чувствительных к вирусу. В какой бы участок чашки ни попали вирусные частицы, они заражают какую-либо бактерию. Вокоре инфекция распространяется на соседние бактерии и в результате образуется стерильное пятно (рис. 15-20). Число основных вирусных частиц, содержащихся в суспензии, можно легко определить, сосчитав число стерильных пятен, образовавшихся в результате посева. [c.248]
Т.к. клеточных стенок (их содержание может достигать 75% от сухой массы этих стенок) ковалентно связаны с пептидогликанами, причем область связывания представляет собой олигомерное звено, содержап(ее аминосахара и глицерофосфат, но отличающееся по строению от повторяющихся звеньев Т.к. Как правило, ближайшим к пепти-догликану является остаток глюкозамина атом С-1 этого моносахарида связан фосфодиэфирной связью с атомом С-6 остатка мурамовой к-ты пептидогликана, а с др. стороны звена (ближайшей к Т.к.) остаток глицерофосфата фосфодиэфирной связью соединен с остатком полиола Т. к. [c.509]
Толстая стенка растительной клетки (рис. 1-3) устроена необычайно сложно [ИЗ—116]. Благодаря ее сложному строению растения обладают прочностью и жесткостью, а их клетки способны к быстрому удлинению в период роста. Норткот [ИЗ] сравнил строение стенки растений с фибраглассом — пластиком, армированным стекловолокном. Так, в стенке клетки находятся микрофибриллы, состоящие из целлюлозы и других полисахаридов, которые погружены в матрикс, также состоящий в основном из полисахаридов. На ранних стадиях роста зеленых растений закладывается первичная клеточная стенка, содержащая свободно переплетенные целлюлозные волокна диаметром приблизительно 10 нм, центральная часть которых (- 4 нм) имеет кристаллическую структуру. Такие целлюлозные волокна содержат 8000—12 000 остатков глюкозы. [c.395]
Клеточная стенка анатомических элементов древесины, волокон технической целлюлозы и других волокнистых полуфабрикатов имеет сложное строение, связанное с распределением в клеточной стенке высокомолекулярных химических компонентов. Для изучения этих вопросов применяют, кроме световой, микроскопию в ультрафиолетовом и поляризованном свете, а также флюоресцентную микроскопию. Для исследования тонкого строения клеточной стенки - ультраструктуры (субмикроструктуры) используют главным образом электронную микроскопию (см. 5.4) с применением просвечивающих (ПЭМ) и растровых, или сканирующих, электронных микроскопов (РЭМ). Эти исследования имеют важное значение для понимания изменений, происходящих с анатомическими элементами древесины и другого растительного сырья, а также в клеточной стенке в процессах делигнификации и других процессах химической и химико-механической переработки древесины. [c.214]
Целлюлоза - наиболее распространенный в природе полисахарид. Кроме древесины, в большом количестве она содержится в семенных волосках хлопка (96...99%), в лубяных волокнах таких текстильных растений, как лен, рами (80...90%), соломе злаков и др. Свойства целлюлозы -физические, физико-химические и химические зависят как от химического строения целлюлозы, так и от ее физической структуры - формы макромолекул, межмолекулярного взаимодействия, надмолекулярной структурь[ и фазового и релаксационного (физического) состояний. Целлюлоза, будучи основным компонентом клеточных стенок, во многом определяет строение и свойства древесины. [c.225]
На сорбцию паров воды древесиной алияют химическое строение компонентов древесины, их надмолекулярная структура, а также ультраструктура клеточных стенок и анатомическое строение древесных тканей. Выделенные из древесины компоненты по сорбционной способности могут значительно отличаться от компонентов в древесине в зависимости от метода выделения. Выделенная из древесины целлюлоза набухает в воде в большей степени, чем сама древесина. [c.265]
Эти необычные полимеры, содержащие остатки фосфорной кислоты, составляют до 50 % сухой массы клеточных стенок некоторых грамположительных бактерий. Они являются также мембранными и внутриклеточными компонентами бактерий. Тейхоевые кислоты прочно закреплены в клеточной стенке, и для их экстракции необходим такой реагент, как трнхлоруксусная кислота. Их распространению, строению и свойствам посвящен обзор [136]. Известны тейхоевые кислоты двух типов, один из которых содержит цепи из остатков D-глицерина, связанных фосфодиэфирными связями второй тип вместо D-глицерина содержит D-рибит. Рибит-тейхоевые кислоты содержат углеводные остатки, присоединенные гликозидной связью в глицеринтейхоевых кислотах углеводные остатки имеются лишь в некоторых случаях. [c.251]
В углеводной части древесины главным образом содержатся полисахариды различного строения и в небольшом количестве полиурониды. В зависимости от растворимости и функций в древесине полисахариды (основную часть) относят к структурным (структурообразующим) компонентам, участвующим в построении клеточной стенки, или же к экстрактивным веществам (водорастворимые полисахариды и полиурониды). Четкое разделение на такие две части провести, однако, невозможно. Структурные полисахариды, не извлекаемые из древесины нейтральными растворителями и в том числе водой, объединяют общим понятием холоцеллюлоза. [c.268]
В древесине гемицеллюлозы, водорастворимые полисахариды и полиурониды выполняют различные функции. Гемицеллюлозы являются структурными компонентами клеточной стенки, тогда как водорастворимые полисахариды - резервными питательными веществами. Однако следует подчеркнуть, что четкое отнесение того или иного индивидуального полисахарида к одной из этих двух фупп часто становится затруднительным, поскольку в древесине присутствуют некоторые нецеллюлозные полисахариды, относящиеся по химическому строению (по главному составляющему моносахариду) к одному и тому же типу, но отличающиеся по функции и растворимости. [c.269]
При действии концентрированных кислот на измельченную древесину кислота проникает в полости клеток, а затем в клеточные стенки и вызывает набухание древесины. Далее начинается растворение полисахаридов. В первую очередь растворяются и гидролизуются гемицеллюлозы, а затем уже целлюлоза с целлюлозанами. На скорость процесса гидролитической деструкции оказывает влияние не только надмолекулярная структура полисахаридов, но и строение клеточной стенки и древесной ткани в целом. [c.294]
Строение клеточной стенки Staphylo o us aureus схематически изображено на рис. 23.5.1. Здесь X и Y представляют остатки N-ацетилглюкозамина и N-ацетилмурамовой кислоты соответственно. Пустыми кружками обозначены аминокислоты, которых в возникающих пептидогликанах содержится пять. Однако когда [c.339]
Вароссио и Бреджер [140] анализировали образцы ели, сосны и березовой пихты, взятые из свай, балок и половиц строений, имевших возраст 75—600 лет. Древесина, которая была утоплена или погружена в грунт, имела пониженное содержание целлюлозы при явном увеличении содержания лигнина. Это показывало, что лигнин является наиболее устойчивым компонентом органического вещества клеточной стенки. Однако в наиболее старых образцах лигнин был частично разложен. [c.672]
Когда срезы предварительно обрабатывали солью цианистоводородной кислоты, реакция пероксидазы не происходила. Прибавление перекиси водорода к ткани не приводило к образованию лигниноподобных материалов. Это показывало, что неспособность тяговой древесины к лигнификации может вызываться различным строением предшественников лигнина на обеих сторонах стебля. Изменение клеточного метаболизма, ведущее к образованию высококристаллической целлюлозы в клеточной стенке, может не дать мест, с которыми лигнин мог бы образовать связь. По-видимому в лигнифицированных клетках лигнин ассоциирован с нецеллюлозной фракцией клеточной стенки. [c.768]
chem21.info
Строение и состав клеточной стенки
ГЕМИЦЕЛЛЮЛОЗЫ — полисахариды, входящие в состав клеточной стенки растительной ткани наряду с целлюлозой и лигнином. Часть Г. волокнистого строения является устойчивой к действию щелочи. Присутствие Г. в целлюлозной массе ухудшает качество целлюлозы наоборот, в производстве бумаги Г. улучшает процесс размола и механические свойства бумаги. [c.68] Рассмотрены вопросы строения клеточной стенки у различных типов микрооргяниамов, химический состав и строение мембран, а также транспорт веществ через эти структуры с позиции биохимии. Дай раздел, посвященный метаболизму превращений в процессе роста и развития микроорганизмов. Детально освещены пути биосинтеза аминокислот, антибиотиков, витаминов, липидов, токсического начала микробных средств защиты растений, ксенобиотиков, нуклеотидов и нуклеозидов, их производных и флавинов. Рассмотрены некоторые аспекты синтеза биологически активных веществ у микробов, способных к биологической фиксации азота, а также у фотосинтезирующих и метилотрофных микроорганизмов. Кратко показаны взаимосвязи различных биосинтетических путей. [c.2]В состав клеточной стенки эндосперма зерновых культур входят углеводы и белки, хотя содержание последних составляет лишь 1-6%. Полипептиды образуют своего рода матрицу, взаимодействующую с углеводами. Последние можно разделить на углеводы,.образованные из глюкозных остатков с р-связями (глюканы и целлюлозу), и на углеводы, содержащие пентозные сахара — гемицеллюлозу и пентозаны (в разных пропорциях). Среди пентозанов злаковых культур доминируют полимеры арабиноксилана (арабиноза). Подробнее о строении клеточной стенки у злаковых культур см. [31]. [c.24]
Клетка прокариот обладает рядом принципиальных особенностей, касающихся как ее ультраструктурной, так и химической организации (рис. 4). Структуры, расположенные снаружи от ЦПМ (клеточная стенка, капсула, слизистый чехол, жгутики, ворсинки), называют обычно поверхностными структурами. Термином клеточная оболочка часто обозначают все слои, располагающиеся с внешней стороны от ЦПМ (клеточная стенка, капсула, слизистый чехол). ЦПМ вместе с цитоплазмой называется протопластом. Рассмотрим сначала строение, химический состав и функции поверхностных клеточных структур. [c.27]
Высшие растения состоят из огромного числа клеток, определенным образом скрепленных друг с другом окружающими их клеточными стенками. Многие характерные свойства растений прямо или косвенно связаны с наличием этих клеточных стенок. Состав и внешний вид клеточных стенок непосредственно определяются тем, к какому типу принадлежит данная клетка и каковы ее функции. Вместе с тем основные принципы построения всех клеточных стенок поразительно сходны жесткие волокна целлюлозы погружены в матрикс, содержащий множество поперечных сшивок и состоящий из таких полисахаридов, как пектины и гемицеллюлозы, а также из гликопротеинов. Благодаря такому строению первичная клеточная стенка обладает большим запасом прочности при растяжении и способна пропускать лишь молекулы относительно небольшого размера. Если растительную клетку, лишенную клеточной стенки (протопласт), поместить в воду, то она осмотическим путем наберет воду, набухнет и лопнет. В то же время живое содержимое клетки, заключенное в оболочку, набухает и давит на последнюю, в результате чего возникает давление, известное под названием тургорного. Тургор строго регулируется и жизненно необходим как для увеличения размеров клетки, так и для механической жесткости молодого растения. [c.398]
Строение и состав клеточной стенки [c.214]
Эта схема была подробно рассмотрена в ряде работ. Например, в работе [58] было высказано предположение о возможности протекания этих процессов как в протоплазме, так и в клеточных стенках растений. Последняя точка зрения была признана рядом исследователей, которые указанными выше переходами объясняли наблюдаемые изменения в составе клеточных стенок в процессе их развития. Так, на основе приведенных выше схем объяснялось относительное увеличение содержания в клеточных стенках растений пентозанов, высказывалось предположение о существований в макромолекулах целлюлозы дефектных звеньев, состоявших из остатков глюкуроновой кислоты и пентоз. Эта точка зрения в настоящее время считается неправильной, и объясняется этот вывод различным строением макромолекул полисахаридов, входящих в состав гемицеллюлоз. Эти различия в основном сводятся к следующему [c.331]
В состав клеточной стенки низших растений входят полисахариды разнообразного строения (см. гл. 20). [c.601]
Клеточная стенка. Клеточная стенка — важный и обязательный структурный элемент прокариотной клетки (исключение миконлазмы и Ь-формы), располагающийся под капсулой или слизистым чехлом или же непосредственно контактирующий с окружающей средой (у клеток, не содержащих этих слоев клеточной оболочки). На долю клеточной стенки приходится от 5 до 50 % сухих веществ клетки. Клеточная стенка служит механическим барьером между протопластом и внешней средой и придает клеткам определенную, присущую им форму. Концентрация солей в клетке, как правило, намного выше, чем в окружающей среде, и поэтому между ними существует большое различие в осмотическом давлении. Клеточная стенка чисто механически защищает клетку от проникновения в нее избытка воды. По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотных организмов. В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. [c.12]
Реакция гидролиза полисахаридов, входящих в состав клеточных стенок растений, в чистой воде без участия ферментов при нормальной температуре не протекает. Сложность химического гидролиза целлюлозы и других полисахаридов обусловливается высокой степенью организации структуры их молекул. Поэтому целесообразно подробнее остановиться на строении основных полисахаридных компонентов, содержащихся в растениях. [c.132]
Под влиянием внешних условий роста не только может варьировать толщина клеточной стенки, но могут возникать также определенные различия в субмикроскопическом строении клеток и их химическом составе. Например, состав ГМЦ и содержание других химических компонентов в клеточных стенках — в реакционной древесине, т. е. в древесине, образованной под влиянием механического воздействия, — отличаются от их содержания и состава в нормальной древесине [54]. Полисахариды тяговой древесины в древесине бука содержат больше галактозы (6,6%) и глюкозы (73,5%) по сравнению с нормальной (1,6 и 57,4% соответственно). В то же время ксилозы в гидролизатах полисахаридов тяговой древесины меньше (17,3%), чем в нормальной (35,1%). Характерным свойством клеточной стенки тяговой древесины является то, что она изнутри покрыта желатиноподобным слоем [8, 36], обозначаемым О или 4. Предполагается, что этот слой состоит почти целиком из целлюлозы. По-видимому, механизм [c.42]
У грамотрицательных прокариот строение клеточной стенки намного сложнее, чем у грамположительных (рис. 1.4). В ее состав входит гораздо большее число макромолекул разного химического типа. [c.16]
Некоторые элементы морфологического строения грибов К грибам относятся организмы, очень разнообразные по своей форме. В отличие от высших растений грибы не имеют корней, стволов или стеблей, листьев, а также хлорофилла и не способны к самостоятельной ассимиляции. В связи с этим подавляющее большинство видов грибов — паразитические организмы, живущие за счет готовых органических веществ в растениях и животных. Грибы имеют клеточную структуру, и клетки содержат хорошо заметные ядра. Размножаются они как половым, так п бесполым путем. Тело грибов (thallus) состоит из грибницы — сплетения нитей, гиф, которые густой сеткой покрывают объекты, служащие источником питания гриба. Гифы разделены перегородками на клетки или на большего или меньшего размера многоядерные объединенные клетки — синцитии, в целом образующие мицелий, или грибницу. Оболочки, стенки клеток образованы из калозы, грибной целлюлезы — углевода, близкого к лигнину. У некоторых видов грибов стенки клеток состоят из хитиноподобных веществ — грибного хитина. Состав клеточных стенок изменяется с возрастом грибницы, а также под влиянием условий среды произрастания гриба. [c.281]
Отрыв индивидуальной макромолекулы целлюлозы от этих волокон весьма затруднен, поэтому целлюлоза нерастворима в воде и химически достаточно инертна. Все эти особенности строения и свойств целлюлозы делают ее структурным полисахаридом. Из целлюлозы состоят клеточные стенки растений. В состав древесины входит от 50 до 70 % целлюлозы. [c.247]
Клетки актиномицетов покрыты плотной клеточной оболочкой, имеют протоплазму и ядерное вещество, или нуклеоиды, которые, как и у других бактерий, не покрыты ядерной оболочкой. Клеточная оболочка 10—30 нм толщиной, многослойная или двух-трехслойная. По химическому составу весьма близка к оболочкам грамположительных бактерий. Гексозамины и мукопротеиды — компоненты мукополисахаридов — входят в состав оболочек актиномицетов и прочих грамположительных бактерий. Особенностью оболочки мицелия актиномицетов является мелкопористое строение с диаметром пор около 5 нм. Оболочка воздушного мицелия и спор актиномицетов имеет фибриллярное строение, которое исчезает после обработки ли-зоцимом, смесью этанола, хлороформа и этилового эфира либо другими растворителями липидов это свидетельствует о том, что фибриллярность обусловлена липоидами. По химическому составу клеточной стенки различаются между собой не только [c.17]
Ксилоза является главной составной частью пентозанов, входящих в состав одревесневших клеточных стенок различных растительных тканей. Она представляет собой моносахарид следующего строения [c.364]
Состав и строение клеточной стенки взрослого растения не являются чем-то неизменным составляющие части могут добавляться и удаляться, а связи между компонентами меняться. Локальное удаление материала стенки при развитии горцевых стенок в сосудах ксилемы и ситовидных трубок флоэмы представляют собой поразительный пример таких изменений (рис. 20-16). [c.395]
Обнаруженные у термофильных ацидофилов особенности химического строения мембран нельзя, однако, прямо связать со свойством ацидофильности этих организмов, так как они одновременно находятся под двойным стрессовым воздействием (температуры и кислотности среды), тем более что их мезофильные аналоги (представители рода Thioba illus) имеют довольно типичный химический состав клеточной стенки и мембран. [c.140]
Туберкулезная палочка имеет особенности строения и химического состава клеточной стенки, которые отражаются на всех биологических свойствах. Главная особенность — в клеточной стенке содержится большое количество липидов (до 60%). Большинство из них — миколовые кислоты, которые входят в каркас клеточной стенки, где находятся в виде свободных гликопептидов, входящих в состав корд-факторов. Корд-факторы обуславливают характер роста в виде жгутов. [c.110]
В состав клеточных стенок эндосперма и оболочки семян входит ряд представителей ГМЦ. К растворимым полисахаридам относятся арабииоксиланы, арабиногалактаны. Последние имеют разветвленное строение и ковалентно связаны с пептидом. [c.110]
По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотных организмов. В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. В зависимости от строения клеточной стенки прокариоты, относящиеся к эубак-териям, делятся на две большие фуппы. Было обнаружено, что если фиксированные клетки эубактерий обработать сначала кристаллическим фиолетовым, а затем йодом, образуется окрашенный комплекс. При последующей обработке спиртом в зависимости от строения клеточной стенки судьба комплекса различна у так называемых грамположительных видов этот комплекс удерживается клеткой, и последние остаются окрашенными, у грамотрицательных видов, наоборот, окрашенный комплекс вымывается из клеток, и они обесцвечиваются. У некоторых эубактерий положительная реакция при окрашивании описанным выше способом свойственна только клеткам, находящимся в стадии активного роста. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки. [c.28]
Клеточная стенка. Большинство прокариот имеет ригидную клеточную стенку, под которой расположена ЦПМ. Состав и строение клеточной стенки — важный систематический признак, по которому все прокариоты подразделяются на следующие группы грамположительные, грамотрицательные и не имеющие клеточной стенки. Своеобразным строением и составом клеточной стенки характеризуются археи, которые не синтезируют пептидогликан, но некоторые из них образуют псевдомуреин. [c.34]
Формирование клеточной стенки у разных злаков занимает разное время у риса — 9 дней после оплодотворения, у пшеницы — 20 дней, а у ячменя — 30 дней, причем клеточная стенка эндосперма у риса и кукурузы тоньше, чем у пшеницы и ячменя. Состав клеточных стенок в зерне одной культуры существенно варьирует. У ячменя клеточная стенка в алейроновом слое на 65-67% состоит из пентозана и на 26-29% — из глюкана, тогда как в эндосперме она состоит на 20% из пентозана и на 70% — из глюкана. У пшеницы, ячменя и риса клеточные стенки в алейроновом слое толще, чем в эндосперме, и состоят из двух разных слоев. Более тонкий внутренний слой в процессе прорастания практически не меняется, тогда как внешний слой, имеющий бороздчатое или пластинчатое строение, по мере развития зерна сильно сморщивается , в нем обнаруживаются щелочные экстракты и микрофибриллы целлюлозы. В клеточной стенке в алейроновом слое наблюдаются широкие межклеточные каналы, по которым, как полагают, осуществляется межклеточная коммуникация и которые могут способствовать перемещению ферментов. [c.24]
Виноградные ягоды, а также сусло и вина содержат малоизученные ГМЦ. Они оказывают определенное влияние на качество и технологию произаодства вин. Среди ГМЦ наиболее изучены водорастворимые полисахариды. В их состав входят разнообразные монозы галактоза, глюкоза, манноза, арабиноза, ксилоза, глюкуроновая кислота [59, 60]. В составе клеточных стенок виноградной ягоды найдены арабиногалактан и маннан. Молекулярная масса первого иосле фракционирования колеблется в широком диапазоне — от 7760 до 12 300,второго составляет величину порядка 46 200. Макромолекула этого маннана отличается линейностью строения, состоит из остатков а-О- и р-Д-маннозы. Предиоложено, что основная цепь включает участки, в которых остатки маннозы соединены (1-связями (1—>-3) и (1—>-6), а боковые сформированы. из остатков а-/)-маннозы, соединенных с маниоииранозами основной цепи связью (1— -б). Изучена [59] динамика изменения ГМЦ. в процессе созревания винограда и производства различных вин. [c.113]
Следует различать основные макромолекулярные компоненты клеточной стенки — целлюлозу, полиозы (темицеллюлозы) и лигнин, которые присутствуют в древесине всех видов, и низкомолекулярные компоненты — экстрактивные и минеральные вещества, которые содержатся в меньших количествах и по природе и количеству зависят от ботанического вида дерева. Относительное содержание и химический состав лигнина и полиоз в древесине хвойных и лиственных пород различны, тогда как строение целлюлозы одинаково во всех древесных породах. Химические компоненты древесины представлены на схеме 3.1. Более подробное их обсуждение дано в аналитической части этой главы и особенно в гл. 4—7. [c.17]
Из трех основных полимеров растительного происхождения — целлюлозы, лигнина и гемицеллюлозы — менее всего изучен лигнин Лигнин наряду с целлюлозой - наиболее распространенное в природе органическое вещество, входит в состав одревесневших клеточных стенок всех наземных растений (от 15 до 36% их массы) и может быть охарактеризован как полифункциональный гетеро-цепной хаотически построенный природный полимер ароматической природы, не гидролизуюшийся до мономеров [106-1101 Получено много данных, свидетельствующих о неоднородности природного лигнина, что позволяет считать его групповым веществом, объединяющим природные полимеры близкой структуры с общим типом строения (108, ПО] [c.28]
Клеточная стенка грамотрицательных эубактерий. У фамотрицательных эубактерий строение клеточной стенки намного сложнее, чем у грамположительных (см. рис. 5). В ее состав входит гораздо большее число макромолекул разного химического типа (см. табл. 3). Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к ЦПМ. Для разных видов фамотрицательных эубактерий содержание этого гетерополимера колеблется в широких пределах. У большинства видов он образует одно- или двухслойную структуру, характеризующуюся весьма редкими поперечными связями между гетерополимерными цепями (рис. 9). [c.33]
Важным шагом в развитии систематики прокариот явилось использование признаков, даюших информацию о химическом строении клетки состав оснований ДНК, ДНК—ДНК- и ДНК— РНК-гомологии, аминокислотная последовательность белков, строение рибосом, компонентов клеточной стенки и т.д. [c.157]
Определение химического состава древесиаы имеет большое значение как с теоретической, так и с практической точек зрения. Особенно важно знать химический состав древесины при использовании ее в качестве сырья для химической переработки в связи с тем, что различные отрасли производства предъявляют к древесному сырью различные требования. Однако определение химического состава древесины связано с большими трудностями из-за сложности строения клеточных стенок и существования тесной связи между отдельными компонентами древесины. До сих пор еще не найдено совершенных методов, позволяющих выделять эти компоненты в неизмененном состоянии. Трудности выделения отдельных составных частей древесины осложняются их высокомолекулярной природой. При выделении отдельных веществ древесины (например, целлюлозы, лигнина) приходится прибегать к сравнительно жестким методам химического воздействия, которые вызывают изменение химического состава и молекулярного веса вследствие гидролитических, окислительных и других химических реакций. Кроме того, выделенные вещества, как правило, содержат примеси других компонентов и продуктов их разлонметоды анализа, основанные на выделении отдельных компонентов, не всегда точно характеризуют их количественное содержание в древесине. Следует также отметить, что определение некоторых компонентов древесины (например, пентозанов) основано на косвенных методах с применением эмпирических расчетных формул. [c.8]
Каждая клетка имеет оболочку — клеточную стенку и полость. Клеточные стенки состоят в основном из органических веществ различного строения, образующих вещество древесины. Они включают в себя углеводы — полисахариды (в среднем около 70%) с различной длиной цепных молекул и вещества ароматической природы, составляющие до 30% от древесины. Вся углеводная часть древесины называется холо-целлюлозой, а комплекс ароматических веществ — лигнином. В состав холоцеллюлозы входят целлюлоза и гемицеллюлозы (гексозаны, пентозаны, связанные с пентоза-нами уроновые кислоты и пектиновые вещества). Клеточные стенки удерживают в своем составе незначительные количества минеральных веществ. Полости клеток часто содержат сложные химические вещества, которые можно извлекать из древесины (экстрагировать) нейтральными растворителями (водой и органическими растворителями). Поэтому они получили название экстрактивных веществ. Иногда эти вещества пропитывают и стенки клеток. [c.7]
Большинство бактерий, так же как водоросли и грибы, имеют ригидную клеточную стенку. Но ее состав иной, чем у эукариот. Тпнич1пэ1м компонентом клеточной стенки большинства прокариот, относящихся к эубактериям, является пептидгликан (му-реин), состоящий из Ы-ацетилглюкозамина и Ы-ацетилмурамо-вой кислоты. Ни у одного из эукариот такой полимер не обнаружен. Имеются различия в строении жгутиков, которые обусловливают подвижность ряда эукариот и прокариот, в составе липидов и некоторых других компонентов клеток. [c.18]
Кальций (Са). Этот элемент, так же как и калий, не входит в состав основных органических веществ, но участвует в строении клеточных стенок. В отличие от калия он повышает вязкость белковых веществ. Вещества, находящиеся в коллоидальном состоянии, в присутствии кальция становятся менее подвижными. Таким. одействие друг друга. По этой [c.29]
Разработаны приемы освобождения растительных клеток от твердых клеточных оболочек для получения культуры изолированных протопластов, отграниченных от окружающей среды одной только плазмалеммой. Изолированные протопласты получают в результате комбинированного действия ряда ферментов (пектиназы и целлулазы), которые гидролизуют клеточные оболочки. В результате возникает возможность более детального изучения внутреннего строения клетки. Культивирование протопластов приводит в дальнейшем к ресинтезу клеточных стенок и образованию обычной культуры клеток, из которой затем можно вновь регенерировать целое растение. Изолированные протопласты представляют также большой научный и практический интерес, поскольку, изменяя соответствующим образом состав питательной среды, можно стимулировать их слияние друг с другом, осуществляя таким образом процесс так называемой соматической (неполовой) гибридизации растительных клеток. Культивируемые затем в определенных условиях гибридные протопласты могут дать начало новому растению с признаками, унаследованными от обоих родителей. Соматическая гибридизация может применяться во всех случаях, когда получение гибридов обычным (половым) путем невозможно из-за ряда физиологических или цитогенетических барьеров между растениями, например при отдаленной гибридизации. [c.10]
Первичная клеточная стенка состоит из целлюлозных фибрилл, погруженньгх в матрикс, в состав которого входят другие полисахариды. Целлюлоза тоже представляет собой полисахарид (ее химическое строение описано в разд. З.2.З.). Она обладает высокой прочностью на разрыв, сравнимой с прочностью стали. Матрикс состоит из полисахаридов, которые для удобства описания делят обычно на пектины и гемицеллюлозы. Пектины — это кислые полисахариды с относительно высокой растворимостью. Срединная пластинка, скрепляющая стенки соседних ютеток, состоит из клейких студнеобразных пектатов (солей пектина) магния и кальция. [c.204]
chem21.info










